










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkWater SA
versão On-line ISSN 1816-7950
versão impressa ISSN 0378-4738
Water SA vol.49 no.2 Pretoria Abr. 2023
http://dx.doi.org/10.17159/wsa/2023.v49.i2.3956
RESEARCH PAPER
AquaSens: exploring the use of 16S rRNA next-generation sequencing to determine bacterial composition of various water matrices
Nisreen HoosainI; Jeanne KorsmanII; Peter O KimathiII; Paidamoyo KachambwaII; Rembu MagobaI; Shane L MurrayII
IScientific Services Branch, City of Cape Town (CoCT), Wastewater Avenue, Athlone, Cape Town 7764, South Africa
IIThe Centre for Proteomic and Genomic Research (CPGR), Upper Level, St Peter's Mall, Corner of Anzio and Main Road, Observatory, Cape Town 7925, South Africa
ABSTRACT
Access to clean water, one of the United Nation's Sustainable Development Goals, is challenged by an increase in the presence of emerging microbial and other contaminants due to urbanization, among other factors. Traditionally, the presence of indicator microorganisms is determined using culturing methods. However, these classical methods cannot be used to determine the identities of 'unknown' bacteria and is limited to isolating the culturable state of microorganisms. Thus with culturing, the identities of many bacteria, particularly novel or non-culturable, may remain unknown. The use of a DNA-based method, 16S rRNA next-generation sequencing (NGS), can assist with determining the identities of bacterial populations in a water sample. The objective of this 16S rRNA NGS study was to investigate the bacterial community composition and diversity in a range of water sources. Water samples comprising of potable, surface, ground, marine, aquaculture, rain, wetland and swimming bath water matrices were subjected to 16S rRNA NGS using the Illumina 16S rRNA Metagenomics analysis pipeline. Operational taxonomic units were analysed and the identities of bacterial genera determined. In this study, genera of Acinetobacter, Mycobacterium, Pseudomonas, Legionella, Burkholderia, Yersinia, Staphylococcus and Vibrio were spread across the water matrices. Alpha (within sample) and beta (between samples) diversities for each bacterial community within the tested samples were also determined.
Keywords: bacterial genera, 16S rRNA next-generation, sequencing, water matrices
INTRODUCTION
Approximately 70% of the Earth consists of water. Of this, about 68.7% is not available as it is present in glaciers and ice caps, and about 30.1% is in groundwater, while roughly 0.3% is found in surface water, which includes rivers, swamps and lakes (National Geographic, 2022). Water is the basis of all forms of life and is used for various purposes such as domestic (cooking and cleaning), farming, industry and recreation (Evans, 2019). Water is also an important habitat for bacteria and serves as a means for their dissemination in nature (Vaz-Moreira et al., 2014). Some bacteria have been found to evade disinfection in the drinking water treatment process, either through resistance or via biofilm protection (Simoes and Simöes, 2013; Li et al., 2017). As a result, access to clean water has been adopted as one of the United Nation's Sustainable Development Goals (United Nations, 2023). In 2017, the World Health Organisation (WHO) published their Guidelines for Drinking Water Quality as a tool to ensure that drinking water of good quality is maintained. Currently in South Africa, drinking water is routinely monitored to ensure that it complies with the South African National Standard for drinking water (SANS 241:2015, SABS, 2015). However, exposure to waterborne bacteria is not limited to potable water.
The human gastrointestinal microbiome, shed in faeces, consists of a mixed population of bacteria, some of which are commensals; others exhibit a mutualistic relationship, while some may cause harm in the host (Almeida et al., 2019). Worldwide, inadequate human hygiene practices, open defecation, lack of access to clean water, and inadequately treated water have notably contributed to various opportunistic infections (Carr, 2001; WHO, 2019). Several anthropogenic activities as well as human and animal waste can contaminate surface (rivers and streams), coastal and groundwater sources (Páll et al., 2013; Itchon and Bruni, 2020). In South Africa, wastewater treatment plants treat sewage according to licence agreements instituted by the national Department of Water and Sanitation (DWS) to reduce the pollution risk of effluents to the environment. Growing populations, loadshedding or rolling blackouts, sewer line failures and blockages, sewer pump station overflows and breakages as well as increases in emerging contaminants could have an indirect effect on the efficacy of wastewater treatment (City of Cape Town, 2020).
Traditional culture-based techniques determine the presence and number of microorganisms such as bacteria in a tested sample. However, the method is limited to culturable microorganisms (Verhille, 2013), is laborious and less sensitive than molecular techniques such as the quantitative polymerase chain reaction (qPCR). Despite being a sensitive technique, qPCR can only detect and quantitate the target microorganisms such as bacteria that the reaction has been designed for. Thus, unknown/non-suspected bacteria may remain undetected. Urban waters have a diverse array of microorganisms, many of which are still unknown (Jin et al., 2018), thus identifying them using specific approaches such as culturing or qPCR is impractical. This could be overcome with the use of screening molecular techniques such as next-generation sequencing (NGS). 16S rRNA NGS has transformed the field of bacterial ecology (Bautista-de los Santos et al., 2016; Brandt et al., 2018; Zhang and Liu, 2019). The composition of a bacterial population can be determined using the unique 16S rRNA-encoding DNA sequences in samples and through comparing it to all known 16S rRNA sequences in a database to determine the presence, abundance and taxonomy level of the bacteria present. Alpha (within sample) and beta (between samples) diversity can be calculated for each bacterial community giving an indication of the diversity and abundance within the bacterial community (Bautista-de los Santos et al., 2016).
This study aimed to use 16S rRNA NGS to investigate the bacterial composition in an array of filtered water samples such as potable, surface, ground, marine, aquaculture, rain, wetland and swimming baths. This study also determined the alpha (within sample) and beta (between samples) diversity for each bacterial community within the tested samples.
METHODS
Sample collection
Water samples were mainly collected from the Cape Metropole area and consisted of municipal samples as well as citizen science samples. Municipal potable water samples were collected in disposable 250mL PET sterile (gamma radiated) bottles with 3% so dium thiosulphate; while swimming bath samples were collected in pre-autoclaved bottles, transported on ice and immediately filtered upon arrival at the laboratory. Citizen science samples were collected in a sterile manner at the water source using sterile disposable gloves. The stipulated instructions for water filtration and storage were supplied with the water collection kit. Each water sample (three 60 mL aliquots) was aseptically filtered using a syringe-filtering unit (cellulose acetate membranes with 0.2 μm pores; GVS Filter Technologies, UK) and touching of the syringe and filter attachment points was avoided. All processed filters were placed in sterile plastic zip-lock bags and stored at -20°C until delivered to CPGR. Frozen filters were transported on ice to CPGR and stored at -20°C prior to DNA extraction. Water samples were collected (50 from the municipality and 50 from a variety of water sources supplied by members of the public, see Supplementary Material I) between 2019/10/28 and 2019/11/26. In tandem, the municipal samples were subjected to standard microbial analyses and assessed according to the required guidelines (WHO, 2006; DoH, 2015; SANS, 2015).
DNA extraction and QC
Filters were sterilely removed and DNA extractions carried out using the ZymoBIOMICS 96 MagBead DNA Kit (Zymo Research, USA). As a negative control, DNA was extracted from a sterile filter. DNA quantification was carried out on the GloMax Multi Detection System (Promega, USA) using the fluorescent-based QuantiFluor dsDNA System (Promega, USA).
Sequencing
In addition to all extracted DNA samples, the ZymoBIOMICS Microbial Community DNA Standard (Zymo Research, USA) positive control and a no-template control were included in library preparation and sequencing. The 16S V3-V4 rRNA amplicons were produced according to the Illumina 16S Metagenomics Sequencing Library Preparation guide using KAPA HiFi HotStart ReadyMix (Roche, USA) and the Nextera XT Index Kit v2 (Illumina, USA). Libraries were normalised to 5 ng/μL or left at their original concentration if lower than 5 ng/μL. Up to 10 ng of each sequencing library was pooled. The concentration of the final library pool was confirmed by qPCR using the KAPA Illumina Library Quantification Kit (Roche, USA) and diluted to a concentration of 4nM, denatured, diluted to 5 pM and combined with the 10% PhiX control (Illumina, USA). Sequencing was performed on an Illumina MiSeq (Illumina, USA) using an Illumina MiSeq Reagent Kit v2 (500 cycles).
Data analysis
The Illumina 16S rRNA Metagenomics analysis pipeline classified sequencing reads using an Illumina-curated version of the Greengenes database (DeSantis et al., 2006) and run on all samples using the MiSeq Reporter software version 2.6.2.3 (Illumina, USA). The resulting summary text files for each sample contained operational taxonomic unit (OTU) classifications, the related number and percentage of OTU reads. OTUs that were not assigned to a taxonomic classification were labelled 'Unclassified, along with the related number and percentage of reads. Programming scripts were used to (i) filter OTUs with less than 1% of the total number of reads; (ii) sum and list OTUs to genus level; while (iii) remaining OTUs with a classification higher than genus were labelled 'Other, level > genus'; (iv) identify bacterial genera based on WHO, 2006; USEPA, 2012 and WHO, 2017 guidelines; and (v) calculate the Shannon Diversity Index (H) as follows: H = Σ - (Pi x ln Pi), where Pi = proportion of the sample represented by Genus (Spellerberg and Fedor, 2003). Further data analysis and graphical representation was carried out using Excel, the on-line Venn diagram tool (Ghent University, 2023) or JAMOVI version 1.6.13 (Jamovi, 2023). After testing for normality using the Shapiro Wilk test, Welch's one-way ANOVA with a Games-Howell post-hoc test was used to determine statistically significant differences between Shannon diversities of the different water matrices.
FASTQ files were uploaded to the CyVerse Data Store (Cyverse, 2023) and analysed using the DNA Subway (Cold Spring Harbor Laboratory, 2023) Metabarcoding pipeline with a simplified version of the QIIME 2 pipeline (Bolyen et al., 2019). The Greengenes database was used for classification. Beta diversity was analysed by principal component analysis (PCA) of the unweighted UniFrac distance which measures the community dissimilarity and incorporates phylogenetic relationships between the features (Lozupone and Knight, 2005).
Sequencing reads were deposited on the NCBI Sequence Read Archive with Project Number PRJNA775828.
RESULTS AND DISCUSSION
Samples, DNA extraction and library preparation
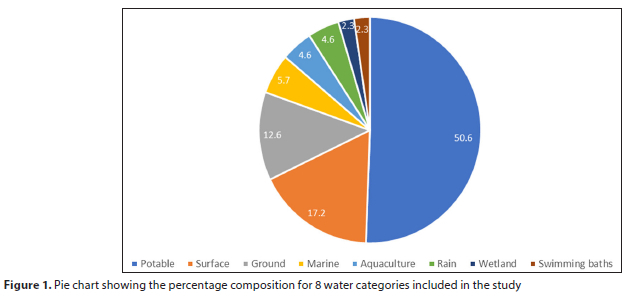
A mixture of filtered water samples were received from the municipality and from members of the public, which included samples from an abalone farm, an aquaponics system, boreholes, bottled water, household taps, freshwater and marine tanks, harbour and sea samples, rainwater tanks, rivers, springs, an estuary, a vlei and wellpoints. Municipal potable water and swimming bath samples were subjected to standard microbial analyses. All municipal samples tested were compliant (Supplementary Material I), except for two desalination plants which exceeded the SANS241:2015 limits (Supplementary Material I). Both swimming bath samples complied with the national and WHO operational guidelines for microbial testing (Supplementary Material I) (WHO, 2006; DoH, 2015).
DNA concentrations from the extracted water samples ranged from <1 ng/μL to 37 ng/μL (Supplementary Material I). To test the utility of the DNA across this concentration range, 16S rRNA sequencing libraries were produced from all samples and quantified. The concentration of the sterile filter negative control library was 0.317 ng/μL (Supplementary Material I). Thirteen filtered water samples had library DNA concentrations <0.317 ng/μL and were therefore excluded from further analysis. Thus only 87 of the 100 samples were subjected for further analyses in the study. These water samples were divided into 8 categories: potable water, surface water (from dams and rivers), groundwater (from springs, wellpoints and boreholes), marine water (from sea, harbour and marine water tanks), aquaculture samples (from abalone and aquaponics systems), rainwater tanks, wetland samples (vlei and estuary) and swimming baths (Fig. 1, Supplementary Material I).
Sequencing and data analysis
The MiSeq sequencing run resulted in 7.72 Gb non-indexed sequence data. The Q-score distribution plot generated a Q30 score of 76.0% for the sequencing run, indicating that the quality of the sequencing data was acceptable.
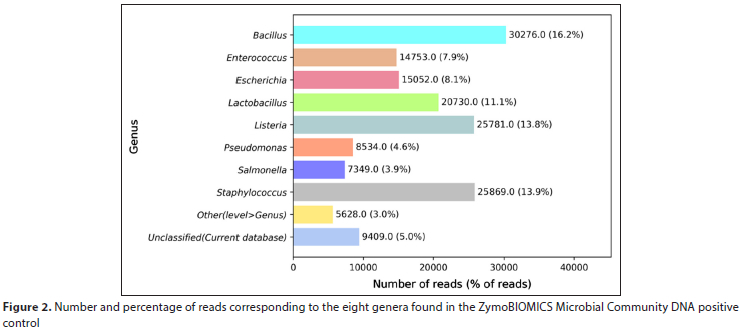
The sequencing output was run through Illumina's 16S rRNA metagenomics pipeline. The number of sequencing reads for the detected genera based on WHO (2006), EPA (2012) and WHO (2017) were identified (Supplementary Material I). Supplementary Material II lists the genera identified in each water sample. A very low number of reads was obtained for the no-template and sterile filter negative controls (885 and 7 540 filtered reads, respectively). The percentage of filtered reads classified at the genus level with high confidence for the ZymoBIOMICS positive control are shown in Fig. 2. The ZymoBIOMICS Microbial Community DNA Standard consists of 8 bacterial genera and all 8 genera were detected, ranging from 3.9% of filtered total reads (for Salmonella spp.) to 16.2% of filtered total reads (for Bacillus spp.).
Bacterial diversity of varied water sources
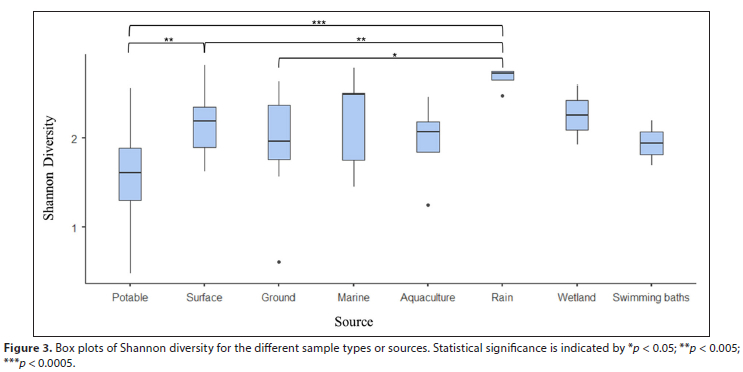
The Shannon Diversity Index was used to calculate an overall score of diversity for each sample at the genus level (Supplementary Material I). To compare diversity ranges within the eight water categories, the mean Shannon Diversity Index was calculated (Fig. 3). The potable water category had the lowest average Shannon diversity. This was significantly lower than rain and surface water, whereas the rain tank category (reflecting samples harvested from rooves and stored prior to purification) had the highest Shannon diversity (Fig. 3).
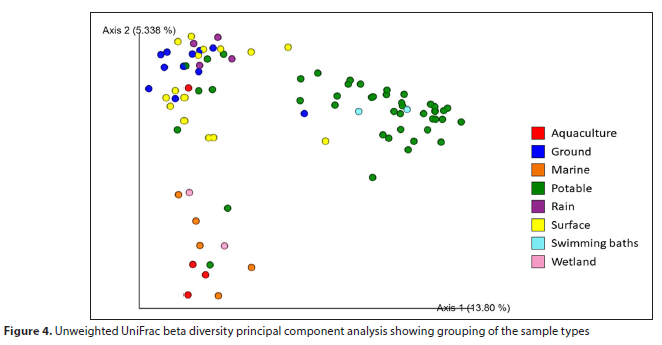
Beta diversity of the sample types was determined using an unweighted UniFrac PCA. This measured bacterial community differences between the tested samples (Lozupone and Knight, 2005). The majority of the potable samples, together with the swimming bath samples, broadly separated from the other sample types based on the first component contributing to the variation (13.8% of the variation), whereas the rain tank and the majority of the ground and surface water samples grouped further from the other sample types based on the second component (5.338% of the variation) (Fig. 4).
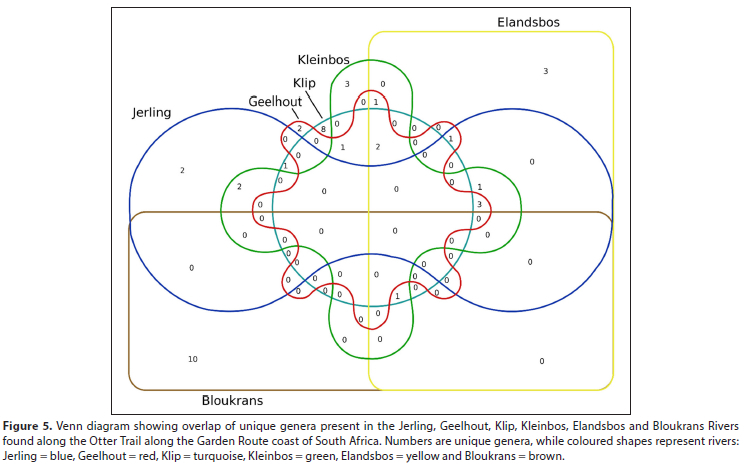
Within the surface water category, 6 river samples were collected within a few days of each other from clearly defined river crossings along the Otter Trail, a hiking trail along the Garden Route coast of South Africa. This dataset offered an opportunity to study the diversity of different genera in nearby rivers over a very short time. Comparison of the genera identified for each sample (Supplementary Material II, Fig. 5) indicated that there was very little overlap between the different samples. This suggests that the composition of the bacterial populations in these river samples may be influenced by distance to the river mouth, localised geology and upstream farming practices (Doherty et al., 2017; Chen et al., 2018).
Identification of bacterial genera
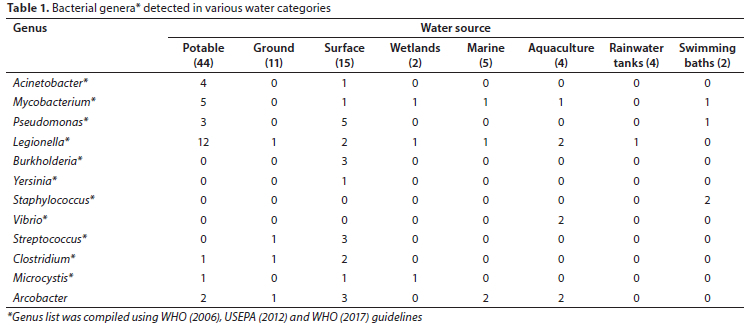
Filtered sequencing data were further analysed to determine the genera based on WHO (2006), USEPA (2012) and WHO (2017) guidelines, in each water category (Table 1). Sequencing reads corresponding to 8 genera (Acinetobacter, Mycobacterium, Pseudomonas, Legionella, Burkholderia, Yersinia, Staphylococcus and Vibrio) were detected in at least one matrix (Table 1; WHO, 2017). In addition, sequencing reads corresponding to the genera Streptococcus, Clostridium, Microcyctis, and Arcobacter were detected in more than one water matrix (Table 1; WHO, 2006; USEPA, 2012).
Currently, the Acinetobacter genus consists of 74 different species (LPSN, 2022a; Parte et al., 2020), is considered to be ubiquitous in nature, and its presence in sewage, soil, water, food and clinical environments is noted (Towner, 2006; WHO, 2017). Acinetobacter spp. have been linked to enhanced biological phosphorus removal from wastewater (Wagner et al., 1994; Yang et al., 2015) and have bioremediation potential (Paisio et al., 2016; Al-Hadithi et al., 2017) suggesting a further possibility for their presence in waterbodies. To date, Acinetobacter spp. have been found in potable water (Carvalheira et al., 2021), rivers (Kittinger et al., 2018), wastewater (Yang et al., 2015), groundwater (Pindi et al., 2013) and seawater (Yoon et al., 2007). In this study, Acinetobacter was detected in 4 potable water samples (2 tap and 2 desalination water samples) and 1 surface (river) water sample (Table 1; Supplementary Material I).
The Mycobacterium genus consists of 3 groups: (1) obligate pathogens, (2) potential pathogens and (3) saprophytes (Vaerewijck et al., 2005). Group 1, the M. tuberculosis complex (MTC), causes disease in humans and animals and is generally not found in the environment. On the other hand, Group 2 (potential pathogens) consists of the Mycobacterium avium complex (MAC) (Hunter, 1997) and is found in terrestrial and aquatic environments. In comparison, Group 3 (saprophytes) are rarely pathogenic (Vaerewijck et al., 2005). Groups 2 and 3 are also known as non-tuberculosis/atypical Mycobacterium (NTM), are naturally occurring in a range of water environments, and are able to grow and persist if conditions are suitable (Le Dantec et al., 2002; WHO, 2017). To date, there are 195 validly published species of Mycobacterium (LPSN, 2022b). In our study, the genus Mycobacterium was identified in 1 sample in each of the following categories: aquaculture, marine, surface, wetland and swimming bath; as well as in 5 potable water samples (Table 1; Supplementary Material I). These findings are in agreement with international studies where Mycobacterium was found in aquaculture (Yanong and Pouder, 2010; Gcebe et al., 2018; Mugetti et al., 2020; Mataragka et al., 2022), marine environments (Hashish et al., 2018; Mataragka et al., 2022), surface water (Eun-Sook et al., 2008), wetlands (Kopecky et al., 2011; Tortone et al., 2018), swimming baths (Torvinen et al., 1999; D'Ancona et al., 2014) and extensively found in potable water sources worldwide (Bailey et al., 1970; Von Reyn et al., 1993; Schwartz et al., 1998; Vaerewijck et al., 2005; Tortone et al., 2018; Lin et al., 2020). Furthermore, chlorine disinfection has been found to be ineffective for Mycobacterium in potable water samples, with the presence of biofilms adding an extra mechanism for its persistence in water (Simoes and Simöes, 2013; Li et al., 2017).
Pseudomonas are free-living micro-organisms and ubiquitous in nature. The genus is found in moist environments, rivers and soils, therapy pools and surrounding areas, and is often associated with biofilm production (Wingender and Flemming, 2011; Koeck et al., 2018). To date, there are 310 validly published species of Pseudomonas (LPSN, 2022c). In our study, the genus Pseudomonas was detected in 3 potable water samples, 1 swimming bath and 5 surface water samples, 4 of which were found along the Otter Trail (Table 1; Supplementary Material I).
Of interest, 20 samples (23%) in this study contained Legionella. Twelve of these were potable water samples (2 from water treatment plants and 10 from distribution points or taps) (Table 1; Supplementary Material I). This is possible as Legionella can grow in water, pipe biofilms and soil. High levels of biodegradable organic carbon, warmer temperatures, stagnant water, low residual chlorine concentrations and biofilms may further exacerbate their growth in water (WHO, 2017). Other water categories where Legionella were detected in our study included: 2 samples from an abalone farm and 1 sample each from a dam and river (surface water), spring, wetlands, marine, and rainwater tank (Table 1; Supplementary Material I). Legionella is an environmental microorganism and may proliferate in suitable conditions (WHO, 2017). Currently, there are more than 58 different Legionella spp. with more than 70 serotypes, (Ditommaso et al, 2021; Legionella Control International Ltd., 2021).
Burkholderia was detected in 3 river samples; 2 of which were along the Otter trail (Table 1; Supplementary Material I). Burkholderia was co-detected with Yersinia in 1 river sample. The principle reservoirs of Yersinia spp. are domestic and wild animals but they can also be found in sewage and polluted waters (WHO, 2017). In 2015, a study by Prentice indicated that a species of Yersinia could enter the viable but non-culturable state in water and could also persist in soil, thus emphasizing the importance for molecular monitoring over culturing for this genus. The presence of Burkholderia spp. has been documented in soil and muddy water in tropical regions (WHO, 2017), coastal environments and mussels (Maravic et al., 2012), a water treatment plant (Inglis et al., 2000) and also maintained its culturable state after several years' incubation in distilled water (Wuthiekanun et al., 1995).
In this study, Staphylococcus was detected in 2 swimming bath samples (Table 1; Supplementary Material I). Staphylococci are ubiquitous in the environment and commonly found in mucous membranes and on human and animal skins (Leroy et al., 2016). As a result of its presence on the skin, it can be transferred to swimming baths, recreational water and spa pools (Koeck et al., 2018). Although this microorganism has been detected in drinking water, there is no evidence that it can be transmitted through water consumption (WHO, 2017). Staphylococci are occasionally found in the gastrointestinal tracts of humans; thus its presence in sewage is also possible (WHO, 2017).
There are currently 138 validly published species of Vibrio (LPSN, 2022d). These species are abundant in aquatic environments as well as in the organs and/or tissues of marine algae and animals, including corals, sponges, bivalves, abalone, shrimp, fish, squid, and zooplankton (Nicolas et al., 2002; Cai et al., 2006; Pichon et al., 2013; Economopoulou et al., 2017), which may negatively impact the infected species and may also have a zoonotic potential (Economopoulou et al., 2017). In our study, the Vibrio genus was detected in 2 aquaculture samples where abalone was farmed.
Although Escherichia spp. sequencing reads were not reported in various water samples of this study (Table 1), it was reported in the ZymoBIOMICS control (Fig. 2). Escherichia spp. and Shigella spp. are closely related and belong to the Enterobacteriaceae family (Devanga Ragupathi et al., 2017). Of the municipal samples collected, one treated desalination plant sample (ASV041) contained 1 CFU/100 mL E. coli (Supplementary Material I). Additionally, poor correlation between 16S microbiome sequencing data and culture-based methods may reflect the limits of detection using 16S sequencing, as low numbers of sequencing reads associated with low concentrations of micro-organisms in communities may be removed during down-stream data analysis steps; alternatively deeper sequencing may be needed to detect micro-organisms in very low abundance (Hamady and Knight, 2009; Schang et al. 2016).
Intestinal enterococci are also used to monitor water quality (WHO, 2017). They consist of2 genera, Enterococcus and Streptococcus, and are used as indicators of faecal pollution. Intestinal enterococci are highly prevalent in sewage and sewage spills as well as human and animal waste. They survive for longer periods in water compared to E. coli, and are resistant to chlorination and dry conditions (WHO, 2017). Limayem et. al. (2019) found Streptococcus to be the second-most predominant genus in influent and treated effluent samples collected at reclamation facilities in the USA. Streptococcus were also found in the Nile River (Goja, 2013) as well as in a groundwater sample in South Africa (Ateba and Moloantoa, 2013). In our study, Enterococcus was not detected but Streptococcus was identified in 3 river samples and 1 groundwater sample (Supplementary Material I).
Clostridium spp. are ubiquitous in nature and often found in the gastro-intestinal tracts of humans and animals. These species can enter surface water systems via runoff in agricultural settings or through treated wastewater effluents (Fourie, 2017). The genus Clostridium was detected in 4 samples in this study, consisting of 2 surface (river), 1 groundwater and 1 potable water sample (Supplementary Material I, II); these surface and groundwater samples also contained Streptococcus spp. The two surface water samples (ASV005 and ASV006) were submitted with a note of 'may contain sewage', suggesting that the presence of both Streptococcus spp. and Clostridium spp. may be linked to faecal contamination, while an explanation for the presence of both genera in a single groundwater sample (ASV016) in our study is unclear.
Cyanobacteria are found in a range of environments, including hot springs, soil, artificially illuminated cave systems, ice layers in polar regions as well as in marine, brackish and freshwater systems (Harding and Paxton, 2001; WHO, 2017; USEPA, 2020). These aquatic organisms produce pigments and generally occur in low numbers in most surface waters; however, eutrophication can result in bloom development. Microcystis is a genus of freshwater cyanobacteria and is capable of causing harmful/toxic as well as non-toxic blooms in ecosystems globally (Park et al., 2018; Pérez-Carrascal et al., 2019). In our study, Microcystis was detected in 1 wetland, 1 potable water (citizen science sample) and 1 surface water (raw sample: water treatment plant) sample (Supplementary Material I, II). All samples were colourless and odourless, suggesting low growth of Microcystis. No sequencing reads corresponding to other cyanotoxic bacteria were detected.
Arcobacter, a genus of emerging concern, has been associated with 3 waterborne outbreaks and its persistence in wastewater is suggestive of an ecological reservoir (Banting and Figueras Salvat, 2017). In our study, Arcobacter was detected in 2 potable, 3 surface, 2 marine, 1 ground and 2 aquaculture water samples, all of which were citizen science samples. This finding is not new as Arcobacter spp. have been identified in several water sources worldwide. These include: tap and river water in Asia (Marita et al., 2004) and Europe (Moreno et al., 2003), seawater in Spain (Collado et al., 2008), groundwater in South Africa and Turkey (Diergaardt et al., 2004; Ertas et al., 2010), and it has also been isolated from abalone in Japan (Mizutani et al., 2019).
Importantly, the viability status of the microorganisms detected in our study was not determined and was not part of the scope of the current study. Rather, it was used to determine the bacteriological composition and diversity within and among the tested water samples.
CONCLUSION
This 16S rRNA amplicon microbiome sequencing study identified genera which included Acinetobacter, Mycobacterium, Pseudomonas, Legionella, Burkholderia, Yersinia, Staphylococcus, Vibrio, Streptococcus, Clostridium, Microcystis and Arcobacter. In addition, this study uncovered a wide range of bacterial diversity at the genus level, both within and between the different water types, with municipal treated potable samples and samples from rain tanks displaying the lowest and highest within-sample type diversity, respectively. Of note, the limit of detection of 16S NGS compared to culture techniques is also an important factor to consider as the indicator micro-organism, E. coli, could be present in lower concentrations compared to the more abundant genera. Thus E. coli may not be detected with NGS but could be cultured using selective media or detected using a rapid technology like IDEXX Colilert.
ACKNOWLEDGEMENTS
Gratitude is extended to CPGR for funding and to members of the public for providing water samples to incorporate a citizen science component in the study. Additionally, municipal staff are thanked for sampling, sample filtration and microbial analyses of the municipal samples.
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
REFERENCES
AL-HADITHI HT, AL-RAZZAQ EA and FADHIL GF (2017) Bioremediation of polycyclic aromatic hydrocarbons by Acinetobacter species isolated from ecological sources. J. Environ. Biol. 38 785-789. https://doi.org/10.22438/jeb/38/5/MRN-422 [ Links ]
ALMEIDA A, MITCHELL AL, BOLAND M, FORSTER SC, GLOOR GB, TARKOWSKA A, LAWLEY TD and FINN RD (2019) A new genomic blueprint of the human gut microbiota. Nature 568 499-504. https://doi.org/10.1038/s41586-019-0965-1 [ Links ]
ATEBA CN and MOLOANTOA M (2013) Isolation of Group B Streptococcus in groundwater in the North West Province, South Africa. Life Sci. J. 10 (3) 444-449. [ Links ]
BAILEY RK, WYLES S, DINGLEY M, HESSE F and KENT GW (1970) The isolation of high catalase Mycobacterium kansasii from tap water. Am. Rev. Respirat. Dis. 101 (3) 430-431. [ Links ]
BANTING G and FIGUERAS SALVAT MJ (2017) Arcobacter. In: Pruden A, Ashbolt N and Miller J (eds) Part 3: Specific Excreted Pathogens: Environmental and Epidemiology Aspects - Section 2: Bacteria. In: Rose JB and Jiménez-Cisneros (eds) Water and Sanitation for the 21st Century: Health and Microbiological Aspects of Excreta and Wastewater Management (Global Water Pathogen Project). Michigan State University, E Lansing, MI, UNESCO. https://doi.org/10.14321/waterpathogens.22 [ Links ]
BAUTISTA-DE LOS SANTOS OM, SCHROEDER JL, SEVILLANO-RIVERA MC, SUNGTHONG R, IJIAZ UZ, SLOAN WT and PINTO AJ (2016) Emerging investigators series: microbial communities in full-scale drinking water distribution systems -a meta-analysis. Environ. Sci. Water Res. Technol. 2 (4) 631-644. https://doi.org/10.1039/C6EW00030D [ Links ]
BRANDT J and ALBERTSEN M (2018) Investigation of detection limits and the influence of DNA extraction and primer choice on the observed microbial communities in drinking water samples using 16 S rRNA gene amplicon sequencing. Front. Microbiol. 9 (2140) https://doi.org/10.3389/fmicb.2018.02140 [ Links ]
BOLYEN E, RIDEOUT JR, DILLON MR, BOKULICH NA, ABNET CC, AL-GHALITH GA, ALEXANDER H, ALM EJ, ARUMUGAM M, ASNICAR F and co-authors (2019) Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37 852-857. https://doi.org/10.1038/s41587-019-0209-9 [ Links ]
CAI J, HAN Y and WANG Z (2006) Isolation of Vibrio parahaemolyticus from abalone (Haliotis diversicolor supertexta L.) postlarvae associated with mass mortalities. Aquaculture 257 (1-4) 161-166. https://doi.org/10.1016/j.aquaculture.2006.03.007 [ Links ]
CA RR R (2001) Excreta-related infections and the role of sanitation in the control of transmission. In: Fewtrell L and Bartram J (eds) World Health Organization (WHO), Water Quality: Guidelines, Standards and Health. IWA Publishing, London. ISBN: 1900222280. [ Links ]
CARVALHEIRA A, SILVA J and TEIXEIRA P (2021) Acinetobacter spp. in food and drinking water - A review. Food Microbiol. 95 103675. https://doi.org/10.1016/j.fm.2020.103675 [ Links ]
CHEN W, WILKES G, KHAN IUH, PINTAR KDM, THOMAS JL, LÉVESQUE CA, CHAPADOS JT, TOPP E and LAPEN DR (2018) Aquatic bacterial communities associated with land use and environmental factors in agricultural landscapes using a metabarcoding approach. Front. Microbiol. 9 2301. https://doi.org/10.3389/fmicb.2018.02301 [ Links ]
CITY OF CAPE TOWN (2020) Know your coast: Key findings from over 10 000 sample bacterial tests at 99 sites along 307 km of coastline. URL: https://resource.capetown.gov.za/documentcentre/Documents/City%20research%20reports%20and%20review/Know_Your_Coast_2020.pdf (Accessed 30 June 2021). [ Links ]
COLLADO L, INZA I, GUARRO J and FIGUERAS MJ (2008) Presence of Arcobacter spp. in environmental waters correlates with high levels of faecal pollution. Environ. Microbiol. 10 1635-1640. https://doi.org/10.1111/j.1462-2920.2007.01555.x [ Links ]
COLD SPRING HARBOR LABORATORY (2023) DNA Subway. URL: https://dnasubway.cyverse.org (Accessed 11 April 2023). [ Links ]
CYVERSE (2023) The open science workspace for collaborative data-driven discovery. URL: https://cyverse.org (Accessed 11 April 2023). [ Links ]
D'ANCONA FP, KANITZ EE, MARINELLI L, SINAGRA JL, PRIGNANO G, CEROCCHI C, BONADONNA L, TORTOLI E, CAPITANIO B, COTTARELLI A and DE GIUSTI M (2014) Non tuberculous cutaneous mycobacteriosis in a primary school in Rome: epidemiological and microbiological investigation. Ann. Igiene Med. Prevent. Comun. 26 (4) 305-310. https://doi.org/10.7416/ai.2014.1990 [ Links ]
DoH (Department of Health, South Africa) (2015) National environmental health norms and standards for premises and acceptable monitoring standards for environmental health practitioners. Government Gazette No. 39561. URL: www.gpwonline.co.za [ Links ]
DESANTIS TZ, HUGENHOLTZ P, LARSEN N, ROJAS M, BRODIE EL, KELLER K, HUBER T, DALEVI D, HU P and ANDERSEN GL (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72 (7) 5069-5072. https://doi.org/10.1128/AEM.03006-05 [ Links ]
DEVANGA RAGUPATHI NK, MUTHUIRULANDI SETHUVEL DP, INBANATHAN FY and VEERARAGHAVAN B (2018) Accurate differentiation of Escherichia coli and Shigella serogroups: challenges and strategies. New Microbes New Infect. 21 58-62. https://doi.org/10.1016/j.nmni.2017.09.003 [ Links ]
DIERGAARDT SM, VENTER SN, SPREETH A, THERON J and BROZEL VS (2004) The occurrence of Campylobacters in water sources in South Africa. Water Res. 38 2589-2595. https://doi.org/10.1016/j.watres.2004.03.004 [ Links ]
DITOMMASO S, GIACOMUZZI M, MEMOLI G, GARLASCO J and ZOTTI CM (2021) Comparison of BCYEa+AB agar and MWY agar for detection and enumeration of Legionella spp. in hospital water samples. BMC Microbiol. 21 48. https://doi.org/10.1186/s12866-021-02109-1 [ Links ]
DOHERTY M, YAGER PL, MORAN MA, COLES VJ, FORTUNATO CS, KRUSCHE AV, MEDEIROS PM, PAYET JP, RICHEY JE, SATINSKY BM, SAWAKUCHI HO, WARD ND and CRUMP BC (2017) Bacterial biogeography across the Amazon River-ocean continuum. Front. Microbiol. 8 882. https://doi.org/10.3389/fmicb.2017.00882 [ Links ]
ECONOMOPOULOU A, CHOCHLAKIS D, ALMPAN MA, SANDALAKIS V, MARAKI S, TSELENTIS Y and PSAROULAKI A (2017) Environmental investigation for the presence of Vibrio species following a case of severe gastroenteritis in a touristic island. Environ. Sci. Pollut. Res. 24 4835-4840. https://doi.org/10.1007/s11356-016-8231-7 [ Links ]
ERTAS N, DOGRUER Y, GONULALAN Z, GUNER A and ULGER I (2010). Prevalence of Arcobacter species in drinking water, spring water, and raw milk as determined by multiplex PCR. J. Food Protect. 73 2099-2102. https://doi.org/10.4315/0362-028X-73.11.2099 [ Links ]
EUN-SOOK L, LEE M-Y, HAN S-H and KA J-O (2008) Occurrence of molecular differentiation of environmental mycobacteria in surface waters. J. Microbiol. Biotechnol. 18 (7) 1207-12015. [ Links ]
EVANS M (2019) How we use water and why we should conserve more. URL: https://www.thebalancesmb.com/how-do-we-use-water-and-why-conserve-3157863 (Accessed 19 December 2022). [ Links ]
FOURIE JCJ (2017) Characterization of Clostridium spp. isolated from selected surface water systems and aquatic sediment. MSc thesis, North-West University. [ Links ]
GCEBE N, MICHEL AL and HLOKWE TM (2018) Non-tuberculous Mycobacterium species causing mycobacteriosis in farmed aquatic animals of South Africa. BMC Microbiology 18 32 https://doi.org/10.1186/s12866-018-1177-9 [ Links ]
GOJA AM (2013) Bacterial genera and their some species of Nile water. Asian J. Biol. Sci. 6 116-123. https://doi.org/10.3923/ajbs.2013.116.123 [ Links ]
GHENT UNIVERSITY (2023) Bioinformatics and evolutionary genomics. URL: http://bioinformatics.psb.ugent.be/webtools/Venn/ (Accessed 11 April 2023). [ Links ]
HAMADY M and KNIGHT R (2009) Next-generation DNA sequencing/review: microbial community profiling for human microbi-ome projects: tools, techniques and challenges. Genome Res. 19 1141-1152. https://doi.org/10.1101/gr.085464.108 [ Links ]
HARDING W and PAXTON B (2001) Cyanobacteria in South Africa: A review. WRC Report No: TT 153/01. Water Research Commission, Pretoria. URL: http://www.wrc.org.za/wp-content/uploads/mdocs/TT153-01.pdf (Accessed 12 August 2022). [ Links ]
HASHISH E, MERWAD A, ELGAML S, AMER A, KAMAL H, ELSADEK A, MAREI A and SITOHY M (2018) Mycobacterium marinum infection in fish and man: Epidemiology, pathophysiology and management; a review. Vet. Q. 38 35-46. https://doi.org/10.1080/01652176.2018.1447171 [ Links ]
HUNTER PR (1997) Waterborne Disease: Epidemiology and Ecology. New York, Wiley. ISBN: 978-0-471-96646-3 384. [ Links ]
INGLIS TJ, GARROW SC, HENDERSON M, CLAIR A, SAMPSON J, O' REILLY L and CAMERON B (2000) Burkholderia pseudomallei traced to water treatment plant in Australia. Emerg. Infect. Dis. 6 (1) 56-59. [ Links ]
ITCHON GS and BRUNI M (2020) Pathogens and contaminants. URL: https://sswm.info/arctic-wash/module-2-environment-pollution-levels-implications/further-resources-environment-and/pathogens-%26-contaminants- (Accessed 19 August 2022). [ Links ]
JAMOVI (2023) Open statistical software for the desktop and cloud. URL: https://www.jamovi.org (Accessed 11 April 2023). [ Links ]
JIN D, KONG X, CUI B, JIN S, XIE Y, WANG X and DENG Y (2018) Bacterial communities and potential waterborne pathogens within typical urban surface waters. Sci. Rep. 8 13368. https://doi.org/10.1038/s41598-018-31706-w [ Links ]
KITTINGER C, KIRSCHNER A, LIPP M, BAUMERT R, MASCHER F, FARNLEITNER AH and ZARFEL GE (2018) Antibiotic resistance of Acinetobacter spp. isolates from river Danube: susceptibility stays high. Int. J. Environ. Res. Publ. Health 15 (1) 52. https://doi.org/10.3390/ijerph15010052 [ Links ]
KOECK DE, HUBER S, HANIFI N, KÖSTER M, SCHIERLING MB and HÖLLER C (2018) Occurrence of antibiotic-resistant bacteria in therapy pools and surrounding surfaces. Int. J. Environ. Res. Publ. Health 15 (12) 2666. https://doi.org/10.3390/ijerph15122666 [ Links ]
KOPECKY J, KYSELKOVA M, OMELKA M, CERMAK, L, NOVOTNA J, GRUNDMANN G, MOËNNE-LOCCOZ and SAGOVA-MARECKOVA M (2011) Environmental mycobacteria closely related to the pathogenic species evidenced in an acidic forest wetland. Soil Biol. Biochem. 43 697-700. https://doi.org/10.1016/j.soilbio.2010.11.033 [ Links ]
LE DANTEC C, DUGUET J-P, MONTIEL A, DUMOUTIER N, DUBROU S and VINCENT V (2002) Occurrence of Mycobacteria in water treatment lines and in water distribution systems. J. Appl. Environ. Microbiol. 68 (11) 5318-5325. https://doi.org/10.1128/AEM.68.11.5318-5325.2002 [ Links ]
LEGIONELLA CONTROL INTERNATIONAL LTD (2021) How many Legionella species exist and which ones cause Legionnaires' disease? URL: https://legionellacontrol.com/legionella/legionella-species/ (Accessed 11 April 2023). [ Links ]
LEROY S, VERMASSEN A and TALON R (2016) Staphylococcus: Occurrence and properties. In: Caballero B, Finglas PM and Toldrá F (eds) Encyclopaedia of Food and Health. Academic Press, Oxford. 140-145. https://doi.org/10.1016/B978-0-12-384947-2.00656-5 [ Links ]
LI Q, YU S, LI L, LIU G, GU Z, LIU M, LIU Z, YE Y, XIA Q and REN L (2017) Microbial communities shaped by treatment processes in a drinking water treatment plant and their contribution and threat to drinking water safety. Front. Microbiol. 8 2465. https://doi.org/10.3389/fmicb.2017.02465 [ Links ]
LIMAYEM A, WASSON S, MEHTA M, POKHREL AR, PATIL S, NGUYEN M, CHEN J and NAYAK B (2019) High-throughput detection of bacterial community and its drug-resistance profiling from local reclaimed wastewater plants. Front. Cell. Infect. Microbiol. https://doi.org/10.3389/fcimb.2019.00303 [ Links ]
LIN W, YE C, GUO L, HU D and YU X (2020) Analysis of microbial contamination of household water purifiers. Appl. Microbiol. Biotechnol. 104 (10) 4533-4545. https://doi.org/10.1007/s00253-020-10510-5 [ Links ]
LOZUPONE C and KNIGHT R (2005) UniFrac: a New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 71 (12) 8228. https://doi.org/10.1128/AEM.71.12.8228-8235.2005 [ Links ]
LPSN (List of Prokaryotic Names with Standing in Nomenclature) (2022a) Genus Acinetobacter. URL: https://www.bacterio.net/genus/acinetobacter (Accessed 28 October 2022). [ Links ]
LPSN (List of Prokaryotic Names with Standing in Nomenclature) (2022b) Genus Mycobacterium. URL: https://www.bacterio.net/genus/mycobacterium (Accessed on 31 October 2022). [ Links ]
LPSN (List of Prokaryotic Names with Standing in Nomenclature) (2022c) Genus Pseudomonas. URL: https://www.bacterio.net/genus/pseudomonas (Accessed on 03 November 2022). [ Links ]
LPSN (List of Prokaryotic Names with Standing in Nomenclature) (2022d) Genus Vibrio. URL: https://www.bacterio.net/genus/vibrio (Accessed on 31 October 2022). [ Links ]
MARAVIC A, SKOCIBUSIC M, SPRUNG M, SAMANIC I, PUIZINA J and PAVELA-VRANCIC (2012) Occurrence and antibiotic susceptibility profiles of Burkholderia cepacia complex in coastal marine environments. Int. J. Environ. Health Res. 22 (6) 531-542. https://doi.org/10.1080/09603123.2012.667797 [ Links ]
MARITA Y, MARUYAMA S, KABEYA H, BOONMAR S, NIMSUPHAN B, NAGAI A, KOZAWA K, NAKAJIMA T, MIKAMI T and KIMURA H (2004) Isolation and phylogenetic analysis of Arcobacter spp. in ground chicken meat and environmental water in Japan and Thailand. Microbiol. Immunol. 48 527-533. https://doi.org/10.1111/j.1348-0421.2004.tb03548.x [ Links ]
MATARAGKA A, TZIMOTOUDIS N, KOLYGAS M, KARAVANIS E and IKONOMOPOULOS J (2022) Diagnostic investigation for the detection of mycobacteria in samples of fish feeds and tissue from sea bream and sea bass with severe granulomatous lesions Aquaculture 546 737283. https://doi.org/10.1016/j.aquaculture.2021.737283 [ Links ]
MIZUTANI Y, IEHATA S, MORI T, OH R, FUKUZAKI S and TANAKA R (2019) Diversity, enumeration and isolation of Arcobacter spp. in the giant abalone, Haliotis gigantea. Microbiol. Open 8 e890. URL: https://doi.org/10.1002/mbo3.890 [ Links ]
MORENO Y, BOTELLA S, ALONSO JL, FERRÚS MA and HERNANDEZ J (2003) Specific detection of Arcobacter and Campylobacter strains in water and sewage by PCR and fluorescent in situ hybridization. Appl. Environ. Microbiol. 69 1181-1186. https://doi.org/10.1128/AEM.69.2.1181-1186.2003 [ Links ]
MUGETTI D, VARELLO K, GUSTINELLI A, PASTORINO P, MENCONI V, FLORIO D, FIORAVANTI ML, BOZZETTA E, ZOPPI S, DONDO A and PREARO M (2020) Mycobacterium pseudoshottsii in Mediterranean fish farms: new trouble for European aquaculture? Pathogens 9 (8) 610. https://doi.org/10.3390/pathogens9080610 [ Links ]
NATIONAL GEOGRAPHIC (2022) Earth's freshwater. In: Resource Library. URL: https://education.nationalgeographic.org/resource/earths-fresh-water (Accessed 2 June 2022). [ Links ]
NICOLAS JL, BASUYAUX O, MAZURIÉ J and THÉBAULT A (2002) Vibrio carchariae, a pathogen of the abalone Haliotis tuberculate. Dis. Aquat. Organ. 50 (1) 35-43. https://doi.org/10.3354/dao050035 [ Links ]
PAISIO CE, TALANO MA, GONZALEZ PS, MAGALLANES-NOGUERA C, KURINA-SANZ M and AGOSTINI E (2016) Biotechnological tools to improve bioremediation of phenol by Acinetobacter sp. RTE14. Environ. Technol. 37 (18) 2379-2390. https://doi.org/10.1080/09593330.2016.1150352 [ Links ]
PÁLL E, NICULAE M, KISS T, SANDRU CD and SPlNU M (2013) Human impact on the microbiological water quality of the rivers. J. Med. Microbiol. 62 1635-1640. https://doi.org/10.1099/jmm.0.055749-0 [ Links ]
PARK BS, LI, Z, KANG Y-H, SHIN HH, JOO J-H and HAN M-S (2018) Distinct bloom dynamics of toxic and non-toxic Microcystis (Cyanobacteria) subpopulations in Hoedong Reservoir (Korea) Microb. Ecol. 75 163-173. https://doi.org/10.1007/s00248-017-1030-y [ Links ]
PARTE AC, SARDÄ CARBASSE, J, MEIER-KOLTHOFF JP, REIMER LC and GÖKER M (2020). List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ. Int. J. of Syst. Evol. Microbiol. 70 5607-5612. https://doi.org/10.1099/ijsem.0.004332 [ Links ]
PÉREZ-CARRASCAL OM, TERRAT Y, GIANI A, FORTIN N, GREER CW, TROMAS N and SHAPIRO BJ (2019) Coherence of Microcystis species revealed through population genomics. ISME J. 13 (12) 2887-2900. https://doi.org/10.1038/s41396-019-0481-1 [ Links ]
PICHON D, CUDENNEC B, HUCHETTE S, DJEDIAT C, RENAULT T, PAILLARD C and AUZOUX-BORDENAVE S (2013) Characterization of abalone Heliotis tuberculata - Vibrio harveyi interactions in gill primary cultures. Cytotechnology 65 (5) 759-772. https://doi.org/10.1007/s10616-013-9583-1 [ Links ]
PINDI PK, YADAV PR and SHANKER AS (2013) Identification of opportunistic pathogenic bacteria in drinking water samples of different rural health centers and their clinical implications on humans. BioMed Res. Int. Article ID 348250. 10 pp. https://doi.org/10.1155/2013/348250 [ Links ]
PRENTICE M (2015) Yersinia and yersiniosis. Global Water Pathogen Project. URL: https://www.waterpathogens.org/node/126 [ Links ]
SCHANG C, HENRY R, KOLOTELO PA, PROSSER T, CROSBIE N, GRANT T, COTTAM D, O'BRIEN P, COUTTS S, DELETIC A and co-authors (2016) Evaluation of techniques for measuring microbial hazards in bathing waters: A comparative study. PLos One 11 (5) e0155848. https://doi.org/10.1371/journal.pone.0155848 [ Links ]
SCHWARTZ T, KALMBACH S, HOFFMANN S, SZEWZYK U and OBST U (1998) PCR-based detection of mycobacteria in biofilms from a drinking water distribution system. J. Microbiol. Meth. 34 (2) 113-123. https://doi.org/10.1016/S0167-7012(98)00081-5 [ Links ]
SABS (South African Bureau of Standards) (2015) SANS (South African National Standard). Drinking water: Part 1: Microbiological, physical, aesthetic and chemical determinants. South African Bureau of Standards, Pretoria. ISBN 978-0-626-29841-8. [ Links ]
SIMOES LC and SIMÖES M (2013) Biofilms in drinking water: problems and solutions. R. Soc. Chem. Adv. 3 (8) 2520-2533. https://doi.org/10.1039/C2RA22243D [ Links ]
TORTONE CA, ZUMÁRRAGA MJ, GIOFFRÉ AK and ORIANI DS (2018) Utilization of molecular and conventional methods for the identification of nontuberculous mycobacteria isolated from different water sources. Int. J. Mycobacteriol. 7 (1) 53-60. [ Links ]
TORVINEN E, NORTHRUP J, ARBEIT R, RISTOLA M, MATTILA L and REYN C (1999) Isolation of mycobacteria from indoor swimming pools in Finland. APMIS 107 193-200. https://doi.org/10.1111/j.1699-0463.1999.tb01544.x [ Links ]
TOWNER K (2006) The genus Acinetobacter. In: Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E (eds) The Prokaryotes. Springer, New York. https://doi.org/10.1007/0-387-30746-X_25 [ Links ]
UNITED NATIONS (2023) The 17 goals. United Nations Department of Economic and Social Affairs Sustainable Development. URL: https://sdgs.un.org/goals (Accessed 11 April 2023). [ Links ]
USEPA (United States Environmental Protection Agency) (2012) Microbial risk assessment guideline: pathogenic microorganisms with focus on food and water. EPA/100/J-12/001. United States Environmental Protection Agency, Washington DC. [ Links ]
USEPA (United States Environmental Protection Agency) (2020) Health effects from cyanotoxins. URL: https://www.epa.gov/cyanohabs/health-effects-cyanotoxins (Accessed 13 August 2021). United States Environmental Protection Agency, Washington DC. [ Links ]
VAEREWIJCK MJM, HUYS G, PALOMINO JC, SWING J and PORTAELS F (2005) Mycobacteria in drinking water distribution systems: ecology and significance for human health. FEMS Microbiol. Rev. 29 (5) 911-934. https://doi.org/10.1016/j.femsre.2005.02.001 [ Links ]
VAZ-MOREIRA I, NUNES OC and MANAIA CM (2014) Bacterial diversity and antibiotic resistance in water habitats: searching the links with the human microbiome. FEMS Microbiol. Rev. 38 (4) 761-778. https://doi.org/10.1111/1574-6976.12062 [ Links ]
VERHILLE S (2013) Understanding microbial indicators for drinking water assessment: interpretation of test results and public health significance. National Collaborating Centre for Environmental Health. URL: https://www.ncceh.ca/sites/default/files/Microbial_Indicators_Jan_2013_0.pdf (Accessed 11 April 2023). [ Links ]
VON REYN CF, WADDELL RD, EATON T, ARBEIT RD, MASLOW JN, BARBER TW, BRINDLE RJ, GILKS CF, LUMIO J and LÄHDEVIRTA J (1993) Isolation of Mycobacterium avium complex from water in the United States, Finland, Zaire, and Kenya. J. Clin. Microbiol. 31 (12) 3227-3230. https://doi.org/10.1128/jcm.31.12.3227-3230.1993 [ Links ]
WAGNER M, ERHART R, MANZ W, AMANN R, LEMMER H, WEDI D and SCHLEIFER K-H (1994) Development of an rRNA-targeted oligonucleotide probe specific for the genus Acinetobacter and its application for in situ monitoring in activated sludge. Appl. Environ. Microbiol. 60 (3) 792-800. https://doi.org/10.1128/aem.60.3.792-800.1994 [ Links ]
WHO (World Health Organization) (2006) Guidelines for safe recreational water environments. Volume 2: Swimming pools and similar environments. WHO, Geneva. ISBN: 92 4 154680 8. URL: http://apps.who.int/iris/bitstream/handle/10665/43336/9241546808_eng.pdf?sequence=1 (Accessed 11 April 2023). [ Links ]
WHO (World Health Organization) (2017) Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the First Addendum. WHO, Geneva. ISBN: 978-92-154995-0. [ Links ]
WHO (World Health Organization) (2019) 1 in 3 people globally do not have access to safe drinking water. UNICEF/WHO, Geneva. URL: https://www.who.int/news/item/18-06-2019-1-in-3-people-globally-do-not-have-access-to-safe-drinking-water-unicef-who (Accessed 12 August 2022). [ Links ]
WINGENDER J and FLEMMING HC (2011) Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health 214 (6) 417-423. https://doi.org/10.1016/j.ijheh.2011.05.009 [ Links ]
WUTHIEKANUN V, SMITH MD and WHITE NJ (1995) Survival of Burkholderia pseudomallei in the absence of nutrients. Trans. R. Soc. Trop. Med. Hyg. 89 (5) 491. https://doi.org/10.1016/0035-9203(95)90080-2 [ Links ]
YANG L, REN Y-X, LIANG, X, ZHAO S-Q, WANG J-P and XIA Z-H (2015) Nitrogen removal characteristics of a heterotrophic nitrifier Acinetobacter junii YB and its potential application for the treatment of high-strength nitrogenous wastewater. Bioresour. Technol. 193 227-233. https://doi.org/10.1016/j.biortech.2015.05.075 [ Links ]
YANONG RPE and POUDER DB (2010) Association of Mycobacteria in recirculating aquaculture systems and mycobacterial disease in fish. J. Aquat. Anim. Health 22 219-223. [ Links ]
YOON J-H, KIM IG and OH T-K (2007) Acinetobacter marinus sp. Nov. and Acinetobacter seohaensis sp. Nov., isolated from seawater of the Yellow Sea in Korea. J. Microbiol. Biotechnol. 17 (11) 1743-1750. [ Links ]
ZHANG Y and LIU W-T (2019) The application of molecular tools to study the drinking water microbiome - current understanding and future needs. Crit. Rev. Environ. Sci. Technol. 49 (13) 1188-1235. https://doi.org/10.1080/10643389.2019.1571351 [ Links ]
 Correspondence:
Correspondence:
Nisreen Hoosain
Email: nisreen.hoosain@capetown.gov.za
Received: 29 October 2021
Accepted: 11 April 2023


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}