










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.43 n.3 Pretoria Jul. 2017
http://dx.doi.org/10.4314/wsa.v43i3.03
Genetic characterization of Salmonella and Shigella spp. isolates recovered from water and riverbed sediment of the Apies River, South Africa
Mutshiene Deogratias Ekwanzala; Akebe Luther King Abia; Jitendra Keshri; Maggy Ndombo Benteke Momba*
Department of Environmental, Water and Earth Sciences, Tshwane University of Technology, Arcadia Campus, Private Bag X680, Pretoria 0001, South Africa
ABSTRACT
Riverbed sediment is a vital component of river ecosystems and plays an important role in many geomorphological and ecological processes. However, when re-suspension occurs, pathogenic bacteria associated with sediment particles may be released into the water column, thus creating a health risk to those who use such water for drinking, household and recreational purposes. The aim of this study was to investigate the presence of bacterial pathogens Salmonella spp. and Shigella spp. in the Apies River and to ascertain whether there was any level of genetic relatedness between river water and riverbed sediment isolates of these pathogenic bacteria. A total of 124 water and sediment samples were collected from a site located on the Apies Rivers upstream of the Daspoort Wastewater Treatment Works, Pretoria, Gauteng, South Africa, between August and November 2014. In order to detect and identify the target bacteria, samples were analysed by culture-dependent and culture-independent techniques (quantitative real-time PCR). Genetic relatedness was established using Sanger sequencing of the invA gene of Salmonella spp. and ipaH of Shigella spp. Results of this study displayed the presence of the target bacteria both in the water and sediment of the river. The phylogenetic tree of Salmonella spp. revealed a ≥ 99% and 99% genetic relatedness between river water and riverbed sediment isolates for Salmonella spp. and Shigella spp., respectively. The degree of genetic relatedness between sediment and water pathogen isolates suggests that these organisms could possibly have a common origin and that there could be possible movement of microorganisms between the water column and the sediments.
Keywords: Salmonella spp., Shigella spp., river water, riverbed sediment, genetic relatedness
INTRODUCTION
Many waterborne pandemics are associated with Shigella spp. and Salmonella spp. (Threlfall, 2002). The genus Shigella contains four species, namely S. flexneri, S. sonnei, S. boydii and S. dysenteriae. All four species are responsible for shigellosis or bacillary dysentery, a disease that causes high fever, neurological disturbances and mucus-pyo-hemorrhagic dysentery (Sansonetti, 2001). The global burden of shigellosis has been estimated to be 150 million cases, with 1 million deaths per year recorded in developing countries (Parsot, 2005). Shigella spp. are normally found in water polluted with human excrement (Saha et al., 2009). The presence of Shigella spp. in drinking water indicates human faecal contamination. This bacterium is of fundamental public health significance because of its great pathogenicity. Outbreaks of shigellosis have been associated with water treatment failures (at times inefficient treatment) in water supply systems (Karanis et al., 2007). Such waterborne outbreaks often lead to a considerable number of individuals being simultaneously affected, and in most cases the outbreak subsides when the water supply is adequately treated (Pillsbury, 2010).
The genus Salmonella consists of two species, namely S. enterica and S. bongori, each of which contains multiple serotypes. Most of the disease-causing serovars are from Subspecies I, with the most important serovars in human health being Typhimurium and Typhi (Lan et al., 2009). Typhoid fever is recognised as a devastating disease in several regions in Asia, Africa and South America, while the disease is rare in developed countries. In 2000, the global burden was estimated to be more than 21 million cases, with up to 200 000 deaths (Crump et al., 2004). Enteric fever is widespread in poor nations, affecting around 12.5 million people yearly (CDC, 2005). Waterborne illnesses associated with Salmonella spp. are more commonly due to increased faecal pollution of water bodies (Levantesi et al., 2012). In South Africa, the MDG (Millennium Development Goals) 2013 report pointed out that some 3-5 million people still depend on untreated water from rivers and springs (UN, 2013). An epidemiological study by Niehaus et al. (2011), following an outbreak of food-borne salmonellosis after a school function in Durban, KwaZulu-Natal, reported that Salmonella enteritidis isolated from patients and food samples could not be distinguished phenotypically and genotypically. The authors suggested a point-source as the origin of the outbreak, with a possibility of continued transmission through the water supply.
Several studies have reported on the occurrence of Salmonella spp. (Tobias and Heinemeyer, 1994; Touron et al., 2005) and Shigella spp. (Baums et al., 2007; Skariyachan et al., 2015; Xiong et al., 2015) in different aquatic sediments, even when these were not isolated in the overlying water. A study by Baudart et al. (2000) showed that Salmonella trapped in sediment particles accumulated in the riverbed during low water levels downstream of a river course and were re-suspended during storm events. In another study, the loads of Salmonella spp. were found to be high in water resources during severe or frequent disturbance events (Walters et al., 2007). The presence of these pathogenic bacteria of faecal origin poses severe threats to environmental and human health, particularly when sediments go through natural or human-induced re-suspension, resulting in them being transported toward downstream areas (Chapman, 2013). The presence of pathogenic bacteria in riverbed sediments highlights the need to investigate the similarity between river sediment and water isolates at genetic level to better understand the microbial dynamics between these two matrices.
Real-time quantitative PCR (qPCR) has been used in monitoring studies to compare the data acquired by culture-based techniques, which have been generally used for evaluation of bacterial pathogens in environmental samples (Mackay, 2004; Robertson and Nicholson, 2005; Kaushik and Balasubramanian, 2012; Yamahara et al., 2012; Garrido et al., 2013). These molecular methods, when applied to DNA purified from environmental samples, permit the enumeration of pathogenic microorganisms even when dead or in a viable, but not yet cultivable, state (Noble and Weisberg, 2005; Lee et al., 2006; Rantsiou et al., 2013). Genetic approaches also provide additional information such as pathogenicity, especially if the genes targeted are those associated with the disease-causing abilities of the organism (Pathak et al., 2012). As a result, qPCR has been used to enumerate bacteria in different sample types, including food (Elizaquível et al., 2012), soil (Prévost-Bouré et al., 2011), wastewater (Sidhu et al., 2013) and marine sediments (Luna et al., 2012; Schippers et al., 2012). Despite these numerous applications, the use of qPCR in riverbed sediment studies is still in its early stages. Sediments may be problematic when it comes to the effective utilization of the qPCR methods due to the presence of inhibitory substances co-extracted with DNA, which can hinder the polymerase chain response, hence reducing the capacity to amplify target DNA (Vezzulli et al., 2009; Sidstedt et al., 2015).
The aim of the present study was to establish the genetic relatedness between Salmonella spp. and Shigella spp. isolated from water and those isolated from riverbed sediments of the Apies River using culture-based and culture-independent methods in a bid to understand genetic similarity among isolates of water and sediments.
MATERIALS AND METHODS
Study area
The Apies River and its tributaries have previously been described by Abia et al. (2015b). Briefly, this river is situated in the Gauteng Province of South Africa, and flows through the city of Pretoria. It falls within the Crocodile (West) Marico Water Management Area, within the Apies River basin. The Apies rises in the Fountains Valley in Pretoria and flows through Gauteng, North-West and Limpopo, where it ultimately joins the Pienaars River to the north of Hammanskraal (Tuwani, 2011). The Apies River has a total flow of approximately 500 m3/year, about 12% of which originates from wastewater treatment works around the river, such as Daspoort, Rooiwal and Mogwase Wastewater Treatment Plants (WWTPs) (Venter, 2007). For the purpose of this study, the sampling point was upstream of the Daspoort Wastewater Treatment Works (WWTW) effluent discharge point where the river is channelled through Pretoria Central and the National Zoological Garden of Pretoria.
Sample collection
Water and riverbed sediment samples were concurrently collected on a weekly basis from August to November, 2014 from the sampling point located in the Apies River (immediately upstream of the Daspoort WWTP discharge point), resulting in a total of 62 water and 62 sediment samples. Water and sediment samples were collected using 1 L sterile containers as previously described by Abia et al. (2015b). Samples were transported to the laboratory in cooler boxes containing ice and analyses were performed within 3 h of the time of collection.
Culture-based enumeration of Salmonella spp. and Shigella spp.
For the culture-based isolation and enumeration of Salmonella spp. and Shigella spp., collected sediment samples were tested using the membrane filtration (MF) methods according to the procedure described in APHA (2001). Prior to the isolation and enumeration of these target pathogens, attached bacteria were dislodged from the sediments using the method developed by Abia et al. (2015a). Briefly, sediment samples were gradually transferred to a graduated 1 L Durham bottle containing 900 mL of 1 × PBS until the 1 000 mL mark was reached, to obtain a 10% dilution (v/v). Thereafter the suspension was vigorously shaken manually for 2 min as described by Abia et al. (2015a), who point out that the water-displacement method is important to dislodge attached microorganisms from the sediment matrix. In addition, use of this method does not influence the growth of target bacteria in the subsequent membrane-filtration step. 100 mL aliquots, along with 10-fold serial dilutions of the resulting phosphate buffer solution, were then analysed using the membrane filtration technique according to standard methods (US EPA, 2002). Two 0.45 µm filters were used; one was placed onto xylose lysine deoxycholate agar (XLD) (Biorad, South Africa) and/or Salmonella-Shigella agar (SS) (Merck, South Africa) and incubated at a temperature of 37°C for 24 h under aerobic conditions, and the other was preserved in 1 mL of 15% glycerol at 0°C for genetic analysis. Since the desired counts required were of the actual water and sediment samples, no enrichment step was needed to enhance and multiply the initial bacterial counts, as also performed by Wolffs et al. (2006) and Marathe et al. (2012). The abundance of cultured Salmonella spp. and Shigella spp. was recorded as colony-forming units (CFU) per 100 mL of water or sediment. Colonies were counted based on their morphological features. The culture-based method was used for presumptive enumeration only. No isolate from this method was used for genetic analysis.
Total bacterial DNA extraction
The preserved filters were thawed and centrifuged for 1 min at 12 000 r/min; then total bacterial DNA from pellets was extracted using InstaGene matrix (BioRad, South Africa) for water samples and ZR Soil Microbe DNA MicroPrep (Zymo Research, USA) for riverbed sediment samples using their respective manufacturer's instructions. The quality and quantity of the isolated DNA was determined by means of the NanoDrop 2000 spectrophotometer (Thermo Scientific, South Africa).
Standard curve
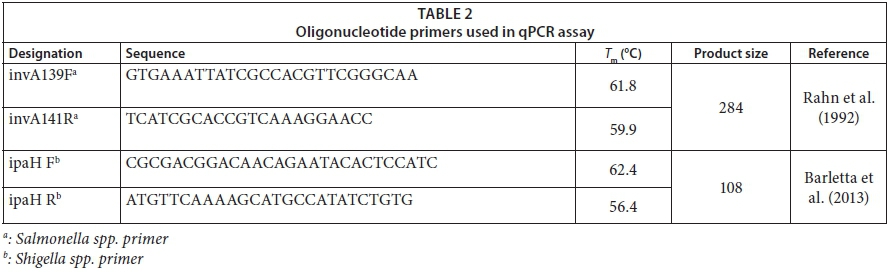
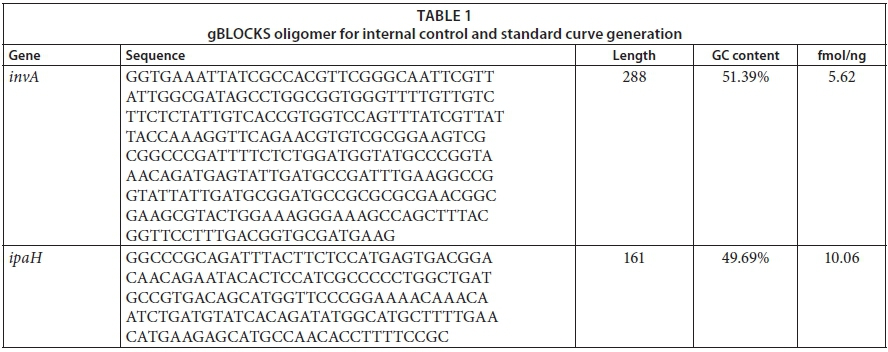
Standard curves converting cycle threshold (Ct) values to bacterial invA and ipaH gene copy numbers were generated as follows: a double-stranded DNA oligomer gBLOCKS was synthesized (Integrated DNA Technologies, Inc., USA) to span the region of the invA and ipaH gene covered by the forward and reverse qPCR primers (Table 2) as listed in Table 1. The lyophilized gBLOCKS were reconstituted to prepare a 20 nM gBLOCKS stock solution that was 10-fold serially diluted and used in the range of 1 ng to 10 fg per reaction (Gunawardana et al., 2014) to produce 10 standards spanning the 20-2.0 × 10−8 nM concentration range (6.8 × 1010-68 for Salmonella spp. and 7.4 × 1011-74 Shigella spp. copy numbers). The standard curves were always performed in duplicate. A graph of Ct values versus log10 (copy number) afforded linear calibration curves with typical R2 values of 0.981 for Salmonella spp. and 0.998 for Shigella spp.
Enumeration of total Salmonella spp. and Shigella spp. by qPCR
The qPCR assays were applied for the determination of the total abundance of Shigella spp. and Salmonella spp. in both river water and riverbed sediment samples. For each type of sample, a volume of 100 mL was filtered through 0.45 μm nitrocellulose membrane filters. The membrane filters were then placed in a cuvette containing 1.5 mL of phosphate buffer. The cuvette was then vortexed for 10 min to dislodge bacteria from the membrane for DNA extraction. Bacterial DNA from water and riverbed sediments was extracted using commercial kits InstaGene Matrix (Bio-Rad, South Africa) and ZR Soil Microbe DNA MiniPrep kit (Zymo Research, USA), respectively. The extracted DNA was quantified using the NanoDrop 2000 spectrophotometer (Thermo Scientific, South Africa). For the purpose of qPCR, primers (synthesized by Inqaba Biotec, South Africa), listed in Table 2, were used where ipaH targeted the invasive plasmid antigen of Shigella spp. and invA targeted the invasive gene of Salmonella spp. The ipaH target gene is carried by all Shigella spp. as well as by enteroinvasive Escherichia coli (E. coli) (EIEC); thus it is used for the diagnosis of dysentery (Sethabutr, 1993). The invA target gene is located on the Salmonella pathogenicity island 1 (SPI1), which encodes proteins of a type III secretion system (Malorny et al., 2003). Reactions were run using SsoFast EvaGreen Supermix on the CFX96 Touch real-time PCR detection system (Bio-Rad, South Africa). All reactions were run in a total volume of 20 μL, containing 10 μL of Supermix, 1 μL of each primer (final concentration 1 μM), 5 μL of template DNA and 3 μL nuclease-free (NF) water (Fermentas, Germany). The qPCR conditions were optimised in terms of the following thermal cycling parameters: 98°C for 2 min for the enzyme activation, followed by 40 amplification cycles of denaturation at 98°C for 5 s, annealing of primers with the genomic DNA (gDNA) template at 60°C for Salmonella spp. and 59°C for Shigella spp., and a primer extension at 72°C for 5 s followed by melt curve analysis steps from 65 to 95°C in 0.5 s increments for 5 s. Cycle threshold (Ct) values were automatically calculated by the Bio-Rad CFX Manager software (Ver. 3.0). Reactions were run in duplicate, using the isolated DNA extracted from both water and riverbed sediment. Purified DNA of Shigella dysenteriae (ATCC 11835) and Salmonella subsp. enterica serovar Typhimurium (ATCC 14028) obtained from the TUT (Tshwane University of Technology) Water Research Group bacteria stock collections were used as positive control. For each reaction and primer set, negative controls were run in duplicate, consisting of primers, PCR Supermix and nuclease-free water instead of gDNA template. The specificity of the assay was assessed by the analysis of the melting curve (Varga and James, 2005; D'Souza et al., 2009). Melting was performed from 54°C to 95°C and 60°C to 95°C for Shigella spp. and Salmonella spp., respectively, at increments of 0.5°C/10 s. The melting temperature (Tm) was defined as the peak of fluorescence in the generated melting curve.
Genetic analysis of Salmonella spp. and Shigella spp. isolated from riverbed sediment and river water samples
Amplification DNA product for Sanger sequencing
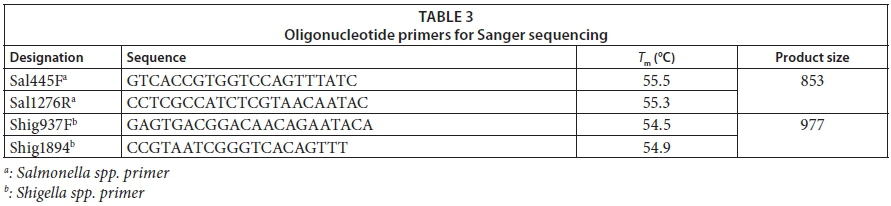
For the amplification process, the following primer sets, as listed in Table 3, were generated using the PrimerQuest Tool software available at www.idtdna.com (IDT, USA) targeting the invA of Salmonella spp. and ipaH gene of Shigella spp.
Gel electrophoresis
For quality control purposes only, amplicons were run through gel electrophoresis of 1% (w/v) agarose gel stained with ethidium bromide, followed by imaging under ultraviolet light. The FastRulerlow range DNA ladder (Fermentas, Germany) was included in all gels as a size marker as well as positive control (DNA of ATCC cultures). These results were captured using a gel documentation system (Syngene, Cambridge, U.K.)
Sanger sequencing of the invA and ipaH gene
Following the gel image, all amplicons showing the desired size of Salmonella spp. and Shigella spp. were sent for sequencing at Inqaba Biotech (South Africa). The dideoxy Sanger sequencing in forward directions only was used with the primer sets listed in Table 3. For this procedure, Big Dye Terminator Cycle Sequencing Kit for ABI3130XL was used according to the manufacturer's instructions and the gel was run on a 3130XL sequencer. Sequences were analysed by comparing them with known invA and ipaH sequences using the BLASTn (Basic Location Alignment Search Tool for nucleotide) algorithm (http://blast.ncbi.nlm.nih.gov/Blast.cgi) to find the closest match in GenBank, EMBL, DDBJ (DNA Data Bank of Japan) and PDB sequence data. Similar type species with 97% resemblance (<3% diversity) to the sequences of isolates were selected as matching species. The invA of Salmonella spp. and ipaH of Shigella spp. sequences were aligned with Clustal X2 (Larkin et al., 2007) and then were edited using BioEdit v.7.2.5 software (Hall, 1999). The distances of relatedness for each invA and ipaH gene were calculated by the neighbour-joining method and phylogenetic trees were created by using MEGA (Tamura et al., 2004). Nucleotide distance matrices of invA and ipaH were calculated by the neighbour-joining method with the Kimura 2-parameter model of substitution on datasets of 500 bootstrap replicates (Tamura et al., 2004). All locations having gaps and missing data were removed from the dataset using the complete deletion option.
All of the newly-sequenced bacteria were deposited in the DNA Data Bank of Japan (DDBJ) and the accession numbers are listed in the supplementary material.
Statistical analysis
The data were statistically analysed using Microsoft Excel 2010. Mean bacterial counts between the river water and riverbed sediments were assessed by two-way ANOVA to test for differences in the abundance of Salmonella spp. and Shigella spp., followed by Tukey's test when significant differences were encountered (p < 0.05).
RESULTS
Salmonella and Shigella culture counts
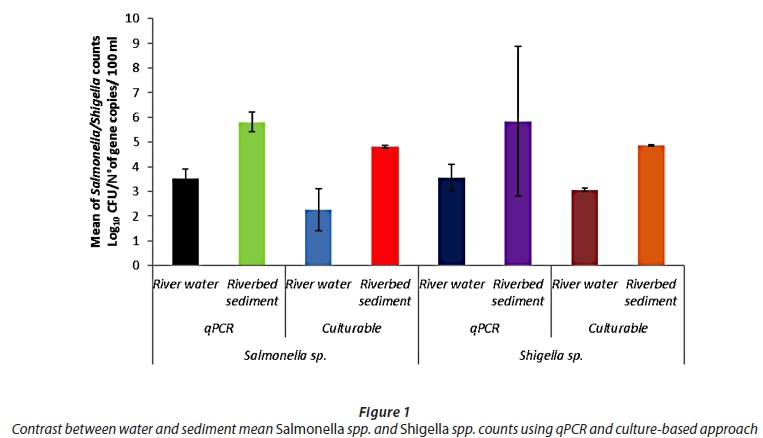
A total of 124 samples (62 water and 62 sediment samples) were collected from the described sampling site on the Apies River for analysis of the abundance of Salmonella spp. and Shigella spp. The mean counts of Salmonella spp. in water and sediment were, respectively, 2.60 log10 and 4.82 log10 CFU/100 mL. The mean counts of Shigella spp. in water and sediment were, respectively, 3.05 log10 and 4.87 log10 CFU/100 mL. Culture counts of Salmonella spp. and Shigella spp. in water were respectively in the range of zero (9.6%) to 2.98 log10 CFU/100 mL and 2.93 to 3.16 log10 CFU/100 mL, while in the sediment samples the counts were in the range of 4.70 to 4.91 log10 CFU/100 mL for Salmonella spp. and 4.78 to 4.92 log10 CFU/100 mL for Shigella spp.
For the qPCR counts, the mean counts of Salmonella spp. invA gene copies in water and sediment were respectively 3.52 log10 and 5.81 log10. The mean counts of Shigella spp. ipaH gene copies in water and sediment were respectively 3.55 log10 and 5.59 log10. The number of invA gene copies of Salmonella spp. and ipaH gene copies of Shigella spp. varied from zero (3.2%) to 1.52 log10 CFU/100 mL and zero (9.6%) to 1.55 log10 CFU/100 mL, respectively, in the water samples, while the number of invA gene copies of Salmonella spp. and ipaH gene copies of Shigella spp. was 'not detected' (3.2%) to 5.82 log10 CFU/100 mL and zero (22.6%) to 5.86 log10 CFU/100 mL, respectively, in the sediment samples of the Apies River.
The contrast between qPCR and culture-based methods revealed that the abundances of Salmonella spp. (5.81 log10) and Shigella spp. (5.59 log10) in the riverbed sediment samples obtained by quantification of gene copies were consistently and significantly higher than those obtained using the culture-based approach (Salmonella - 4.82 log10 CFU/100 mL; Shigella - 4.87 log10 CFU/100 mL) (p < 0.05; Fig. 1). In particular, loads determined by qPCR of Salmonella spp. and Shigella spp. were between 10 and 30 times higher than the loads obtained using culture-based techniques.
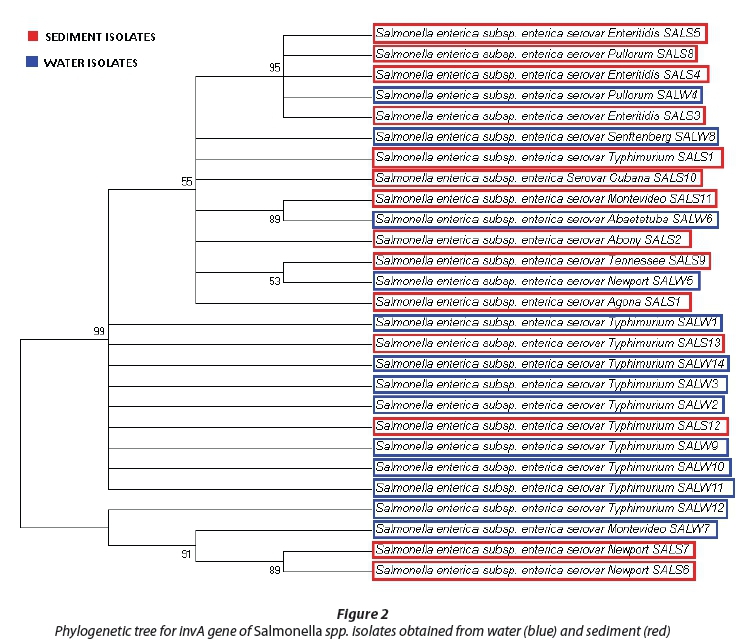
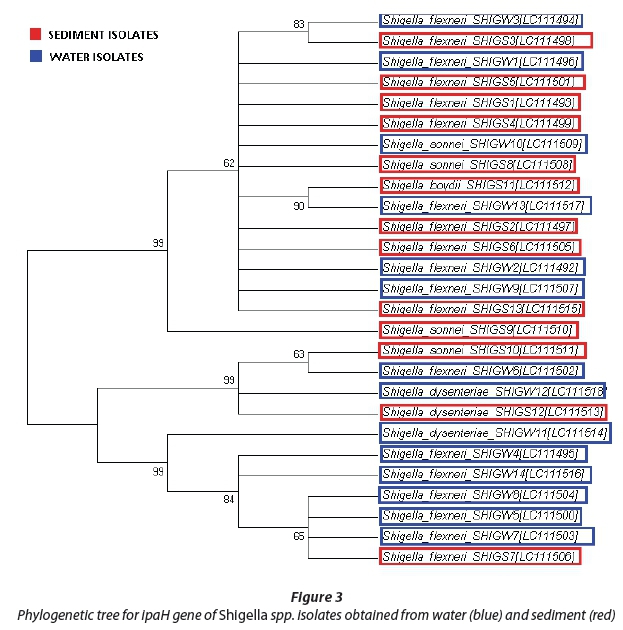
Phylogenetic analysis of isolates
The evolutionary history was gathered using the neighbour-joining system (Saitou and Nei, 1987). The phylogenetic trees with the sum of branch lengths = 1.12364635 (Salmonella spp.) and 2.25308283 (Shigella spp.) are shown in Fig. 2 and 3. The values of replicate trees in which the related taxa clustered together in the bootstrap test (500 replicates) are shown next to the branches (Felsenstein, 1985). The evolutionary distances were processed using the Kimura-2 parameter method (Kimura, 1980) and are in the units of the number of base replacements per location. The analysis involved 28 Salmonella spp. and 28 Shigella spp. nucleotide sequences and first, second and third codon positions were included. All positions containing gaps and missing information were eliminated. There was a sum of 2 072 (Salmonella spp.) and 717 (Shigella spp.) positions in the last dataset. Evolutionary analyses were performed using MEGA6 (Tamura et al., 2013).
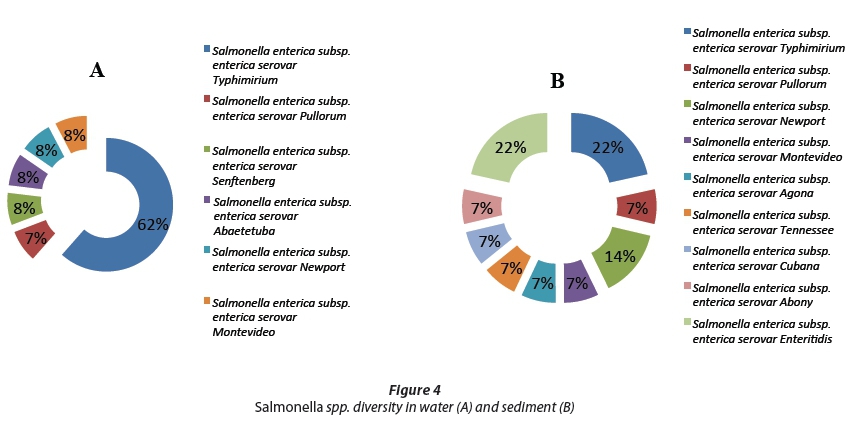
We analysed the invA gene sequences in Salmonella spp.; all fragments belonged to Salmonella enterica with diverse serovars. In water, the most abundant strains were Salmonella enterica subsp. enterica serovar Typhimurium at 62% of all the water isolate fragments, followed by 8% of the other isolate fragments, as shown in Fig. 4. The sediment isolates exhibited a more diverse Salmonella spp. community than the water, with both Salmonella enterica subsp. enterica serovar Typhimurium and Salmonella enterica subsp. enterica serovar Enteritidis having 21.4% of all the isolated sediment fragments, followed by Salmonella enterica subsp. enterica serovar Newport with 14% and the other serovars with 7% each, as shown in Fig. 4. The only other strains found in water samples were Salmonella enterica subsp. enterica serovar Senftenberg and Abaetetuba, while the sediment samples were found to contain Salmonella enterica subsp. enterica serovar Agona, Abony, Cubana, Tennessee and Enteritidis.
As can be seen in Fig. 2, both sediment and water isolates are in a phylogenetic tree with two primary clades. The first clade contains many fragments from water and sediment at 99% gene sequence similarity. The second clade has three fragments, one water isolate and two sediment isolates at 91% genetic similarity. Both water and sediment isolates are clustered together. Although the other observed similarity levels were secondary, no less than 53% sequence similarity was observed.
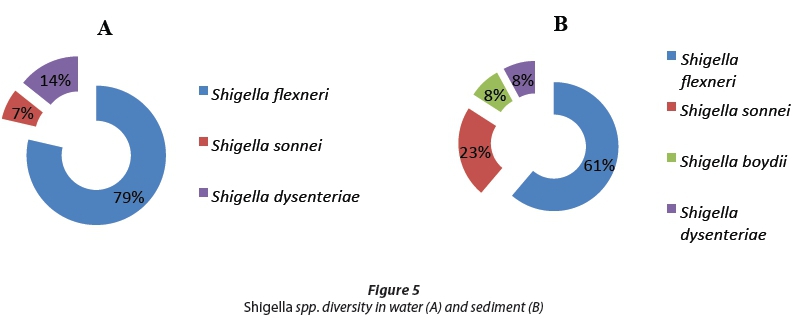
The ipaH gene sequences of Shigella spp. were analysed, and Shigella flexneri was found to be most abundant both in water and sediment, at 79% and 61%, respectively, of all sequences (Fig. 3). In water, two strains were identified as Shigella sonnei and Shigella dysenteriae with sequence similarities of 7% and 14%, respectively, while in the sediments three other strains were isolated, namely, Shigella sonnei (23%), Shigella boydii (8%) and Shigella dysenteriae (8%) as shown in Fig. 5.
In Fig. 3, three primary clades can be observed. All three clades have 99% similarity among the DNA sequences analysed; DNA sequence clustering was observed in both water and sediment samples at no less than 63% of genetic similarity.
DISCUSSION
Pathogen concentrations
Salmonellosis and shigellosis continue to be a major public health problem worldwide, with 93.8 million and 163.2 million cases, respectively, reported annually (Majowicz et al., 2010; Kotloff et al., 2013). Many interrelated factors contributing to this incidence include increased urbanization, inadequate supplies of clean water, antibiotic resistance and increased regional movement (Boehmer et al., 2009). Real-time PCR is a generally used procedure that permits the precise quantification of a particular gene in ecosystem samples (Luna et al., 2012). This method can likewise be used for the determination of the bacterial loads, including microorganisms of faecal source (Noble and Weisberg, 2005). On the other hand, while this technique has been efficiently used for the quantification of pathogenic bacteria in seawater, soil, food and wastewater specimens (Ibekwe and Grieve, 2003; Haugland et al., 2005; Fukushima et al., 2007; Shannon et al., 2007), its application in riverbed sediment tests has still not been extensively researched (Cébron et al., 2008; Vezzulli et al., 2009). For the effective use of qPCR-based examinations on riverbed sediment samples, numerous scientific steps are required, particularly those required for minimizing the presence of inhibitory substances which are associated with nucleic acids (Vezzulli et al., 2009). Inhibitors should be removed from the sediment samples as these substances interfere with the extraction of DNA and prevent the amplification of the nucleic acids through the PCR. Therefore, extraction and refinement steps are of vital importance in sediment sample preparation, as high levels of organic matter, biological pollutants and trace metals act as PCR inhibitors through interaction with DNA or interference with the DNA polymerase enzyme (Fortin et al., 2004).
When contrasting quantification by culture methods versus qPCR, results of this study revealed that qPCR produced higher values compared to the culture-based method. The real-time PCR examinations gave much higher (up to 3 logs) estimations of pathogenic bacteria loads than those obtained using culture-based techniques. Similar results were also observed by Luna et al. (2012) when they conducted qPCR techniques coupled with cultivable enumeration. The authors quantified E. coli, Enterococcus spp. and Salmonella spp. from harbour marine sediments using both culture-based and qPCR techniques. The qPCR technique revealed a higher concentration than the culture-based methods. They found that total E. coli and Enterococcus spp. (qPCR) counts were respectively 65 to 1 571 and 11 to 51 times higher than that from cultured-based quantification. In a study by Su et al. (2013), the high bacterial counts were attributed to viable but non-cultivable bacteria that are found in the environment. In another study, it was found that high qPCR counts may also result from DNA of dead cells in the environment (Wolffs et al., 2005). Culture-based methods are known to seriously underestimate the bacterial pathogen counts in seawater and marine sediment (Shannon et al., 2007; Luna et al., 2010). A similar higher sensitivity of the qPCR method when compared to culture-based approaches has previously been reported in harbour sediment (Luna et al., 2012) and beach sands (Yamahara et al., 2009). These data are vital for an understanding of the potential dangers related to the presence of pathogenic bacteria in sediments. Since the presence of pathogenic bacteria can bring about human diseases with counts of as low as 1.7 × 101 CFU/mL for Salmonella typhimurium (Srinivasan et al., 1982) and 1 × 104 CFU/mL for Shigella spp. (Kothary and Babu, 2001), such a low identification breaking point makes the real-time PCR assay particularly valuable for surveying the microbiological nature of riverbed sediments. Although qPCR does not necessarily produce results comparable to culture-based methods because qPCR measures a genetic, rather than a growth, endpoint, it remains clear that the sensitivity of qPCR is needed in analysing bacteria of public and environmental interest.
In a study conducted by Lindsay et al. (2013), the authors observed that at 4.38 log10 copies/mL of ipaH gene present in the stools of infants, a moderate to severe diarrheal infection occurred. Results in our study are substantially above this value; this may suggest that if Apies River water is consumed, this can lead to diarrhoeal infections in infants. Pathogenic bacteria in sediments have been reported by Atiribom et al. (2007); these authors have reported the isolation of pathogenic organisms such as Aeromonas hydrophila, pathogenic E. coli, S. typhi and Vibrio from sediment and the surrounding areas of the Kainji Lake, Nigeria.
Phylogenetic analysis
The Salmonella spp. isolated during this study included all Salmonella enterica with serovars including Typhimurium, Enteritidis, Pullorum, Senftenberg, Cubana, Montevideo, Abaetetuba, Abony, Tennessee, Newport and Agona. Salmonella enterica serovar Typhimurium was the most frequently detected in both water and sediment samples followed by Salmonella enterica serovar Enteritidis. The only serovar isolated in water was Salmonella enterica serovar Senftenberg, while many other serovars were present only in sediment; this includes: Salmonella enterica serovar Cubana, Abaetetuba, Abony, Tennessee and Agona. These results may suggest that sediments of the Apies River harboured more diversity than the water. All these pathogenic bacteria are linked to gastro-intestinal infections worldwide (Weinberger and Keller, 2005) and are reported to be multidrug resistant (Gordon et al., 2008). The Salmonella enterica isolated exhibited a 99% genetic relatedness which may suggest a common ancestry and origin between isolates from river water and riverbed sediment, as suggested by Grant et al. (2001); Boehm et al. (2002); Kim et al. (2004); Noble and Xu (2004) and Ekwanzala et al. (2017) - i.e., that faecal bacteria from these environments have a high level of similarity. In a recent study by Njage and Buys (2015) on genetic relatedness between commensal and pathogenic strains of E. coli from lettuce and irrigation water using the phylogenetic tree, they found high genetic relatedness despite the sites being 246 km apart.
The Shigella spp. phylogenetic tree analysis shown in Fig. 5 revealed that four Shigella strains were isolated from water and sediments of the Apies River. These include: Shigella flexneri, Shigella sonnei, Shigella boydii and Shigella dysenteriae. Shigella flexneri was the most predominant species found in both river water and riverbed sediments. With a bootstrap value of 99%, this may suggest that Shigella spp. isolated from river water and riverbed sediment are also strongly related at a genetic level. Skariyachan et al. (2015) found that where Shigella sonnei is isolated from sediment this is suggestive of natural or environmental sources of contamination in the overlying water
CONCLUSIONS
Overall, results from this study indicate that the qPCR technique, being highly specific and sensitive, may present a powerful tool which can be routinely used for a true assessment of the pathogenic contamination of water and riverbed sediments. The phylogenetic typing characteristics of isolate sequencing show that pathogenic bacteria isolated from water and sediment samples were closely related (99%). This study recommends future studies to be conducted on multi-locus sequencing or whole genome sequencing techniques in order to emphasize the findings of this study. The presence of pathogenic Salmonella spp. and Shigella spp. is a matter of concern for the communities along the Apies River, who currently depend on this water sources for multiple purposes, including drinking, bathing, recreational and agricultural purposes. An urgent intervention is required by the local Water Service Authority to supply safe drinking water in order to prevent a severe outbreak of waterborne diseases within these communities.
ACKNOWLEDGEMENTS
This work received funding from the Department of Science and Technology (DST)/ the National Research Foundation (NRF)/South African Research Chairs Initiative (SARChI) in Water Quality and Wastewater management. Opinions expressed and conclusions arrived at are those of the authors and are not necessarily to be attributed to the NRF.
REFERENCES
ABIA LKA, UBOMBA-JASWA E, SSEMAKALU CC and MOMBA MNB (2015a) Development of a rapid approach for the enumeration of Escherichia coli in riverbed sediment: case study, the Apies River, South Africa. J. Soil Sediment. 15 (12) 2425-2432. https://doi.org/10.1007/s11368-015-1081-y [ Links ]
ABIA LKA, UBOMBA-JASWA E and MOMBA MNB (2015) Impact of seasonal variation on Escherichia coli concentrations in the riverbed sediments in the Apies River, South Africa. Sci. Total Environ. 537 462-469. https://doi.org/10.1016/j.scitotenv.2015.07.132 [ Links ]
APHA (2001) Compendium of Methods for the Microbiological Examination of Foods. Frances PD, Keith I (eds). APHA, Washington, DC. [ Links ]
ATIRIBOM RY, OVIE SI and AJAYI O (2007) Bacteriological quality of water and fish samples from Kainji Lake and the effects of animal and human activities. In: Fisheries Society of Nigeria (FISON) Conference Proceedings 2007, 12 November 2007. 209-218. [ Links ]
BARLETTA F, MERCADO EH, LLUQUE A, RUIZ J, CLEARY TG and OCHOA TJ (2013) Multiplex real-time PCR for detection of Campylobacter, Salmonella, and Shigella. J. Clin. Microbiol. 51 (9) 2822-2829. https://doi.org/10.1128/JCM.01397-13 [ Links ]
BAUDART J, GRABULOS J, BARUSSEAU JP and LEBARON P (2000) Salmonella spp. and faecal coliform loads in coastal waters from a point vs. nonpoint source of pollution. J. Environ. Qual. 29 (1) 241-250. https://doi.org/10.2134/jeq2000.00472425002900010031x [ Links ]
BAUMS IB, GOODWIN KD, KIESLING T, WANLESS D, DIAZ MR and FELL JW (2007) Luminex detection of faecal indicators in river samples, marine recreational water, and beach sand. Mar. Pollut. Bull. 54 (5) 521-536. https://doi.org/10.1016/j.marpolbul.2006.12.018 [ Links ]
BOEHM AB, SANDERS BF and WINANT CD (2002) Cross-shelf transport at Huntington Beach. Implications for the fate of sewage discharged through an offshore ocean outfall. Environ. Sci. Technol. 36 1899-1906. https://doi.org/10.1021/es0111986 [ Links ]
BOEHMER TK, BAMBERG WM, GHOSH TS, CRONQUIST A, FORNOF ME, CICHON MK, GERSHMAN K, and VOGT RL (2009) Health care-associated outbreak of Salmonella Tennessee in a neonatal intensive care unit. Am. J. Infect. Control 37 (1) 49-55. https://doi.org/10.1016/j.ajic.2007.12.012 [ Links ]
CÉBRON A, NORINI MP, BEGUIRISTAIN T and LEYVAL C (2008) Real-time PCR quantification of PAH-ring hydroxylating dioxygenase (PAH-RHD_) genes from Gram positive and Gram negative bacteria in soil and sediment samples. J. Microbiol. Methods 73 148-159. https://doi.org/10.1016/j.mimet.2008.01.009 [ Links ]
CDC (Center for Disease Control and Prevention) (2005) Preliminary FoodNet data on the incidence of infection with pathogens transmitted commonly through food--10 sites, United States, 2004. MMWR. Morbid. Mortal. Wkly Rep. 54 (14) 352. [ Links ]
CHAPMAN PM, WANG F and CAEIRO SS (2013) Assessing and managing sediment contamination in transitional waters. Environ. Int. 55 71-91. https://doi.org/10.1016/j.envint.2013.02.009 [ Links ]
CRUMP JA, LUBY SP and MINTZ ED (2004) The global burden of typhoid fever. Bull. World Health Organ. 82 (5) 346-353. [ Links ]
D'SOUZA DH, CRITZER FJ and GOLDEN DA (2009) Real-time reverse-transcriptase polymerase chain reaction for the rapid detection of Salmonella using invA primers. Foodborne Pathog. Dis. 6 (9) 1097-1106. https://doi.org/10.1089/fpd.2009.0322 [ Links ]
EKWANZALA MD, ABIA ALK, UBOMBA-JASWA E, KESHRI J and MOMBA MNB (2017) Genetic relatedness of faecal coliforms and enterococci bacteria isolated from water and sediments of the Apies River, Gauteng, South Africa. AMB Expr. 7(1) 20. https://doi.org/10.1186/s13568-016-0319-4 [ Links ]
ELIZAQUÍVEL P, SÁNCHEZ G and AZNAR R (2012) Quantitative detection of viable foodborne E. coli O157: H7, Listeria monocytogenes and Salmonella in fresh-cut vegetables combining propidium monoazide and real-time PCR. Food Control 25 (2) 704-708. https://doi.org/10.1016/j.foodcont.2011.12.003 [ Links ]
FELSENSTEIN J (1985) Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39 783-791. https://doi.org/10.1111/j.1558-5646.1985.tb00420.x [ Links ]
FORTIN N, BEAUMIER D, LEE K and GREER CW (2004) Soil washing improves the recovery of total community DNA from polluted and high organic content sediments. J. Microbiol. Methods 56 181-191. https://doi.org/10.1016/j.mimet.2003.10.006 [ Links ]
FUKUSHIMA H, KATSUBE K, HATA Y, KISHI R and FUJIWARA S (2007) Rapid separation and concentration of food-borne pathogens in food samples prior to quantification by viable-cell counting and real-time PCR. Appl. Environ. Microbiol. 73 (1) 92-100. https://doi.org/10.1128/AEM.01772-06 [ Links ]
GARRIDO A, CHAPELA MJ, ROMÁN B, FAJARDO P, LAGO J, VIEITES JM and CABADO AG (2013) A new multiplex real-time PCR developed method for Salmonella spp. and Listeria monocytogenes detection in food and environmental samples. Food Control 30 (1) 76-85. https://doi.org/10.1016/j.foodcont.2012.06.029 [ Links ]
GORDON MA, GRAHAM S M, WALSH AL, WILSON L, PHIRI A, MOLYNEUX E and MOLYNEUX ME (2008) Epidemics of invasive Salmonella enterica serovar enteritidis and S. enterica serovar typhimurium infection associated with multidrug resistance among adults and children in Malawi. Clin. Infect. Dis. 46 (7) 963-969. https://doi.org/10.1086/529146 [ Links ]
GRANT SB, SANDERS BF, BOEHM AB, REDMAN JA, KIM JH, MRSE RD, CHU AK, GOULDIN MM, MCGEE CD, GARDINER NA, JONES BH, SVEJKOVSKY J, LEIPZIG GV and BROWN A (2001) Generation of enterococci bacteria in a coastal saltwater marsh and its impact on surf zone water quality. Environ. Sci. Technol. 35 2407-2416. https://doi.org/10.1021/es0018163 [ Links ]
GUNAWARDANA M, CHANG S, JIMENEZ A, HOLLAND-MORITZ D, HOLLAND-MORITZ H, LA VAL TP, LUND C, MULLEN M, OLSEN J, SZTAIN TA, YOO J, MOSS JA and BAUM MM (2014) Isolation of PCR quality microbial community DNA from heavily contaminated environments. J. Microbiol. Methods 102 1-7. https://doi.org/10.1016/j.mimet.2014.04.005 [ Links ]
HALL TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41 98-98. [ Links ]
HAUGLAND RA, SIEFRING SC, WYMER LJ, BRENNER KP and DUFOUR AP (2005) Comparison of Enterococcus measurements in freshwater at two recreational beaches by quantitative polymerase chain reaction and membrane filter culture analysis. Water Res. 39 559-568. https://doi.org/10.1016/j.watres.2004.11.011 [ Links ]
IBEKWE AM and GRIEVE CM (2003) Detection and quantification of Escherichia coli O157: H7 in environmental samples by real‐time PCR. J. Appl. Microbiol. 94 (3) 421-431. https://doi.org/10.1046/j.1365-2672.2003.01848.x [ Links ]
KARANIS P, KOURENTI C and SMITH H (2007) Waterborne transmission of protozoan parasites: a worldwide review of outbreaks and lessons learnt. J. Water Health 5 (1) 1-38. https://doi.org/10.2166/wh.2006.002 [ Links ]
KAUSHIK R and BALASUBRAMANIAN R (2012) Assessment of bacterial pathogens in fresh rainwater and airborne particulate matter using real-time PCR. Atmos. Environ. 46 131-139. https://doi.org/10.1016/j.atmosenv.2011.10.013 [ Links ]
KIM JH, GRANT SB, MCGEE CD, SANDERS BF and LARGIER JL (2004) Locating sources of surf zone pollution: a mass budget analysis of fecal indicator bacteria at Huntington Beach, California. Environ. Sci. Technol. 38 2626-2636. https://doi.org/10.1021/es034831r [ Links ]
KIMURA M (1980) A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Gen. Evol. 16 111-120. https://doi.org/10.1007/BF01731581 [ Links ]
KOTHARY MH and BABU US (2001) Infective dose of foodborne pathogens in volunteers: a review. J. Food Saf. 21 (1) 49-68. https://doi.org/10.1111/j.1745-4565.2001.tb00307.x [ Links ]
KOTLOFF KL, WINICKOFF JP, IVANOFF B, CLEMENS JD, SWERDLOW DL, SANSONETTI PJ, ADAK GK and LEVINE MM (2013) Global burden of Shigella infections: implications for vaccine development and implementation of control strategies. Bulletin of the World Health Organization 77 (8) 651-666. [ Links ]
LAN R, REEVES PR and OCTAVIA S (2009) Population structure, origins and evolution of major Salmonella enterica clones. Inf. Gen. Evol. 9 (5) 996-1005. https://doi.org/10.1016/j.meegid.2009.04.011 [ Links ]
LARKIN MA, BLACKSHIELDS G, BOWN NP, CHENNA R, MCGETTIGAN PA, GIBSON TJ, LOPEZ R, MCWILLIAM H, THOMPSON JD, HIGGINS DG, VALENTIN F and ZALLACE I (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23 (21) 2947-2948. https://doi.org/10.1093/bioinformatics/btm404 [ Links ]
LEE C, KIM J, SHIN SG and HWANG S (2006) Absolute and relative qPCR quantification of plasmid copy number in Escherichia coli. J. Biotechnol., 123 (3) 273-280. https://doi.org/10.1016/j.jbiotec.2005.11.014 [ Links ]
LEVANTESI C, BONADONNA L, BRIANCESCO R, GROHMANN E, TOZE S and TANDOI V (2012) Salmonella in surface and drinking water: occurrence and water-mediated transmission. Food Res. Int. 45 (2) 587-602. https://doi.org/10.1016/j.foodres.2011.06.037 [ Links ]
LINDSAY B, OCHIENG JB, IKUMAPAYI UN, TOURE, A, AHMED D, LI S, PANCHALINGAM S, LEVINE MM, KOTLOFF K, RASKO DA, MORRIS CR, JUMA J, FIELDS BS, DIONE M, MALLE D, BECKER SM, HOUPT ER, NATARO JP, SOMMERFELT H, POP M, OUNDO J, ANTONIO M, HOSSAIN A, TAMBOURA B and STINE OC (2013) Quantitative PCR for detection of Shigella improves ascertainment of Shigella burden in children with moderate-to-severe diarrhea in low-income countries. J. Clin. Microbiol. 51 (6) 1740-1746. https://doi.org/10.1128/JCM.02713-12 [ Links ]
LUNA GM, DELL'ANNO A, PIETRANGELI B and DANOVARO R (2012) A new molecular approach based on qPCR for the quantification of faecal bacteria in contaminated marine sediments. J. Biotechnol. 157 (4) 446-453. https://doi.org/10.1016/j.jbiotec.2011.07.033 [ Links ]
MACKAY IM (2004) Real-time PCR in the microbiology laboratory. Clin. Microbiol. Infect. 10 190-212. https://doi.org/10.1111/j.1198-743X.2004.00722.x [ Links ]
MAJOWICZ SE, MUSTO J, SCALLAN E, ANGULO FJ, KIRK M, O'BRIEN SJ, JONES TF, FAZIL A and HOEKSTRA RM (2010) The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. 50 (6) 882-889. https://doi.org/10.1086/650733 [ Links ]
MALORNY B, HOORFAR J, BUNGE C and HELMUTH R (2003) Multicenter validation of the analytical accuracy of Salmonella PCR: towards an international standard. Appl. Environ. Microbiol. 69 290-296. https://doi.org/10.1128/aem.69.1.290-296.2003 [ Links ]
MARATHE SA, CHOWDHURY R, BHATTACHARYA R, NAGARAJAN AG and CHAKRAVORTTY D (2012) Direct detection of Salmonella without pre-enrichment in milk, ice-cream and fruit juice by PCR against hilA gene. Food Control 23 (2) 559-563. https://doi.org/10.1016/j.foodcont.2011.08.005 [ Links ]
UN (United Nations) (2013) Annual Report 24 May 2012 - 1 May 2013. United Nations Publications, New York. eBook Collection (EBSCOhost), viewed 10 March 2016. [ Links ]
NIEHAUS AJ, APALATA T, COOVADIA YM, SMITH AM and MOODLEY P (2011) An outbreak of foodborne salmonellosis in rural KwaZulu-Natal, South Africa. Foodborne Pathog. Dis. 8 (6) 693-697. https://doi.org/10.1089/fpd.2010.0749 [ Links ]
NJAGE PM and BUYS EM (2015) Pathogenic and commensal Escherichia coli from irrigation water show potential in transmission of extended spectrum and AmpC β-lactamases determinants to isolates from lettuce. Microb. Biotechnol. 8 (3) 462-473. https://doi.org/10.1111/1751-7915.12234 [ Links ]
NOBLE M and XU J (2004) Huntington beach shoreline contamination investigation, phase III, final report: Coastal circulation and transport patterns: the likelihood of OCSD's plume impacting the Huntington beach, CA shoreline: U.S. Geological Survey Open-File Report 2004-1019. [ Links ]
NOBLE R and WEISBERG R (2005) A review of technologies for rapid detection of bacteria in recreational waters. J. Water Health 3 381-392. https://doi.org/10.2166/wh.2005.051 [ Links ]
PARSOT C (2005) Shigella spp. and enteroinvasive Escherichia coli pathogenicity factors. FEMS Microbiol. Lett. 252 (1) 11-18. https://doi.org/10.1016/j.femsle.2005.08.046 [ Links ]
PATHAK S, AWUH JA, LEVERSEN NA, FLO TH and ASJØ B (2012) Counting mycobacteria in infected human cells and mouse tissue: a comparison between qPCR and CFU. PLoS One 7 (4) e34931. https://doi.org/10.1371/journal.pone.0034931 [ Links ]
PILLSBURY L, MILLER EA, BOON C and PRAY L (eds) (2010) Providing healthy and safe foods as we age. Workshop Summary. National Academies Press, Washington DC. [ Links ]
PRÉVOST-BOURÉ NC, CHRISTEN R, DEQUIEDT S, MOUGEL C, LELIEVRE M, JOLIVET C, SHAHBAZKIA HR., GUILLOU L, ARROUAYS D and RANJARD L (2011) Validation and application of a PCR primer set to quantify fungal communities in the soil environment by real-time quantitative PCR. PLoS One 6 (9) e24166. https://doi.org/10.1371/journal.pone.0024166 [ Links ]
RAHN K, DE GRANDIS SA, CLARKE RC, MCEWEN SA, GALAN JE, GINOCCHIO C, CURTISS, R III and GYLES CL (1992) Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Gen. Cell. Probes 6 (4) 271-279. https://doi.org/10.1016/0890-8508(92)90002-F [ Links ]
RANTSIOU K, COCOLIN L and SOFOS J (2013) Second-generation polymerase chain reaction (PCR) and DNA microarrays for in vitro and in situ study of foodborne pathogens. Adv. Microb. Food Saf. 1 193-201. https://doi.org/10.1533/9780857098740.3.193 [ Links ]
ROBERTSON BH and NICHOLSON JK (2005) New microbiology tools for public health and their implications 1. Annu. Rev. Public Health 21 (26) 281-302. https://doi.org/10.1146/annurev.publhealth.26.021304.144522 [ Links ]
SAHA T, MURHEKAR M, HUTIN Y J and RAMAMURTHY T (2009) An urban, water-borne outbreak of diarrhoea and shigellosis in a district town in eastern India. Natl. Med. J. India 22 237-239. [ Links ]
SAITOU N and NEI M (1987) the neighbour-joining method: A new method for reconstructing phylogenetic trees. Gen. Biol. Evol. 4 406-425. [ Links ]
SANSONETTI PJ (2001) Rupture, invasion and inflammatory destruction of the intestinal barrier by Shigella, making sense of prokaryote-eukaryote cross-talks. FEMS Microbiol. Rev. 25 (1) 3-14. https://doi.org/10.1016/s0168-6445(00)00060-7 [ Links ]
SCHIPPERS A, KOCK D, HÖFT C, KÖWEKER G and SIEGERT M (2012) Quantification of microbial communities in subsurface marine sediments of the Black Sea and off Namibia. Front. Microbiol. 3 16. https://doi.org/10.3389/fmicb.2012.00016 [ Links ]
SETHABUTR O, VENKATESAN M, MURPHY GS, EAMPOKALAP B, HOGE CW and ECHEVERRIA P (1993) Detection of Shigellae and enteroinvasive Escherichia coli by amplification of the invasion plasmid antigen H DNA sequence in patients with dysentery. J. Infect. Dis. 167 (2) 458-461. https://doi.org/10.1093/infdis/167.2.458 [ Links ]
SHANNON KE, LEE DY, TREVORS JT and BEAUDETTE LA (2007) Application of real time quantitative PCR for the detection of selected bacterial pathogens during municipal wastewater treatment. Sci. Total Environ. 382 121-129. https://doi.org/10.1016/j.scitotenv.2007.02.039 [ Links ]
SIDHU JP, AHMED W and TOZE S (2013) Sensitive detection of human adenovirus from small volume of primary wastewater samples by quantitative PCR. J. Virol. Methods 187 (2) 395-400. https://doi.org/10.1016/j.jviromet.2012.11.002 [ Links ]
SIDSTEDT M, JANSSON L, NILSSON E, NOPPA L, FORSMAN M, RÅDSTRÖM P, HEDMAN J (2015) Humic substances cause fluorescence inhibition in real-time polymerase chain reaction. Anal. Biochem. 487 30-37. https://doi.org/10.1016/j.ab.2015.07.002 [ Links ]
SKARIYACHAN S, MAHAJANAKATTI AB, GRANDHI NJ, PRASANNA A, SEN B, SHARMA N, VASIST KS and NARAYANAPPA R (2015) Environmental monitoring of bacterial contamination and antibiotic resistance patterns of the faecal coliforms isolated from Cauvery River, a major drinking water source in Karnataka, India. Environ. Monit. Assess. 187 (5) 1-13. https://doi.org/10.1007/s10661-015-4488-4 [ Links ]
SRINIVASAN A, FOLEY J, RAVINDRAN R and MCSORLEY, SJ (2004) Low-dose Salmonella infection evades activation of flagellin-specific CD4 T cells. J. Immunol. 173 (6) 4091-4099. https://doi.org/10.4049/jimmunol.173.6.4091 [ Links ]
SU X, CHEN X, HU J, SHEN C and DING L (2013) Exploring the potential environmental functions of viable but non-culturable bacteria. World J. Microbiol. Biotechnol. 29 (12) 2213-2218. https://doi.org/10.1007/s11274-013-1390-5 [ Links ]
TAMURA K, NEI M and KUMAR S (2004) Prospects for inferring very large phylogenies by using the neighbour-joining method. Proc. Natl. Acad. Sci. 101 (30) 11030-11035. https://doi.org/10.1073/pnas.0404206101 [ Links ]
TAMURA K, STECHER G, PETERSON D, FILIPSKI A and KUMAR S (2013) MEGA6: Genetic Evolutionary Genetics Analysis version 6.0. Gen. Biol. Evol. 30 2725-2729. https://doi.org/10.1093/molbev/mst197 [ Links ]
THRELFALL EJ (2002) Antimicrobial drug resistance in Salmonella: problems and perspectives in food- and water-borne infections. FEMS Microbiol. Rev. 26 (2) 141-148. https://doi.org/10.1111/j.1574-6976.2002.tb00606.x [ Links ]
TOBIAS H and HEINEMEYER EA (1994) Occurrence of Salmonella in coastal North Sea water and their hygienic relation to indicator bacteria and sources of contamination. Zbl. Hyg. Umweltmed. 195 (5-6) 495-508. [ Links ]
TOURON A, BERTHE T, PAWLAK B and PETIT F (2005) Detection of Salmonella in environmental water and sediment by a nested-multiplex polymerase chain reaction assay. Res. Microbiol. 156 (4) 541-553. https://doi.org/10.1016/j.resmic.2005.01.001 [ Links ]
TUWANI NP (2011) Success factors for the development of natural based resorts: a comparative analysis of Mphephu, Sagole and Tshipise thermal springs, Limpopo, South Africa. PhD Thesis, UNISA, South Africa. [ Links ]
USEPA (United States Environmental Protection Agency) (2002) Method 1603: Escherichia coli (E. coli) in water by membrane filtration using modified membrane-thermotolerant Escherichia coli agar (modified mTEC). EPA 821-R-02-023. US Environmental Protection Agency, Washington DC. [ Links ]
VARGA A and JAMES D (2005) Detection and differentiation of Plum pox virus using real-time multiplex PCR with SYBR Green and melting curve analysis: a rapid method for strain typing. J. Virol. Methods 123 213-220. https://doi.org/10.1016/j.jviromet.2004.10.005 [ Links ]
VENTER A (2007) 'Water delivery: public or private? J.W. Visser & C. Mbazira (Eds): book review'. S. Afr. Geogr. J. 2 172. [ Links ]
VEZZULLI L, PEZZATI E, MORENO M, STAUDER M, FABIANO M and PRUZZO C (2009) Genetic ecology of marine sediments: determination of Real-Time PCR efficiency for quantifying microbial cells. Chem. Ecol. 25 285-292. https://doi.org/10.1080/02757540903062509 [ Links ]
WALTERS SP, GANNON VP and FIELD KG (2007) Detection of Bacteroidales fecal indicators and the zoonotic pathogens E. coli O157: H7, Salmonella, and Campylobacter in river water. Environ. Sci. Technol. 41 (6) 1856-1862. https://doi.org/10.1021/es0620989 [ Links ]
WEINBERGER M and KELLER N (2005) Recent trends in the epidemiology of non-typhoid Salmonella and antimicrobial resistance: the Israeli experience and worldwide review. Curr. Opin. Infect. Dis. 18 (6) 513-521. https://doi.org/10.1097/01.qco.0000186851.33844.b2 [ Links ]
WOLFFS P, NORLING B and RÅDSTRÖM P (2005) Risk assessment of false-positive quantitative real time PCR results in food, due to detection of DNA originating from dead cells. J. Microbiol. Methods 60 (3) 315-323. https://doi.org/10.1016/j.mimet.2004.10.003 [ Links ]
WOLFFS PG, GLENCROSS L, THIBAUDEAU R and GRIFFITHS MW (2006) Direct quantification of salmonellae in biological samples without enrichment, using two step filtration and real-time PCR. Appl. Environ. Microbial. 72 (6) 3896-3900. https://doi.org/10.1128/AEM.02112-05 [ Links ]
XIONG W, SUN Y, DING X, WANG M and ZENG Z (2015) Selective pressure of antibiotics on ARGs and bacterial communities in manure-polluted freshwater-sediment microcosms. Front. Microbiol. 6 194. https://doi.org/10.3389/fmicb.2015.00194 [ Links ]
YAMAHARA KM., SASSOUBRE LM, GOODWIN KD and BOEHM AB (2012) Occurrence and persistence of bacterial pathogens and indicator organisms in beach sand along the California coast. Appl. Environ. Microbiol. 78 (6) 1733-1745. https://doi.org/10.1128/AEM.06185-11 [ Links ]
YAMAHARA, KM, WALTERS, SP and BOEHM AB (2009) Growth of enterococci in unaltered, unseeded beach sands subjected to tidal wetting. Appl. Environ. Microbiol. 75 (6) 1517-1524. https://doi.org/10.1128/AEM.02278-08 [ Links ]
Received 18 April 2016
Accepted in revised form 2 June 2017
* To whom all correspondence should be addressed. e-mail: mombamnb@tut.ac.za
APPENDIX
All the newly-sequenced bacteria were deposited in the DNA Data Bank of Japan (DDBJ) with the following accession numbers: LC111465 (SALW1), LC111466 (SALW2), LC111467 (SALW3), LC111468 (SALW13), LC111469 (SALW14), LC111470 (SALW12), LC111471 (SALW11), LC111472 (SALW10), LC111473 (SALW9), LC111474 (SALS12), LC111475 (SALW01), LC111476 (SALW8), LC111477 (SALS1), LC111478 (SALS2), LC111479 (SALW7), LC111480 (SALS11), LC111481 (SALS10), LC111482 (SALW6), LC111483 (SALS9), LC111484 (SALS8), LC111485 (SALS7), LC111486 (SALS6), LC111487 (SALS5), LC111488 (SALS4), LC111489 (SALW5), LC111490 (SALS3) LC111491 (SALW4), LC111492 (SHIGW2), LC111493 (SHIGS1), LC111494 (SHIGW3), LC111495 (SHIGW4), LC111496 (SHIGW1), LC111497 (SHIGS2), LC111498 (SHIGS3), LC111499 (SHIGS4), LC111500 (SHIGW5), LC111501 (SHIGS5), LC111502 (SHIGW6), LC111503 (SHIGW7), LC111504 (SHIGW8), LC111505 (SHIGS6), LC111506 (SHIGS7), LC111507 (SHIGW9), LC111508 (SHIGS8), LC111509 (SHIGW10), LC111510 (SHIGS9), LC111511 (SHIGS10), LC111512 (SHIGS11), LC111513 (SHIGS12), LC111514 (SHIGW11), LC111515 (SHIGS13), LC111516 (SHIGW14), LC111517 (SHIGW13) and LC11151 (SHIGW12)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}