










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.43 n.2 Pretoria 2013
REVIEW ARTICLE
Herbal extracts and organic acids as natural feed additives in pig diets
L.B. CostaI, #; F.B. LucianoI; V.S. MiyadaII; F.D. GoisIII
IPUC-PR - School of Agricultural Sciences and Veterinary Medicine, Pontifícia Universidade Católica do Paraná, 83010-500, Brazil
IIUSP/ESALQ - Department of Animal Science, Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, Piracicaba, SP, 13418-900, Brazil
IIIUESC - Department of Agricultural and Environmental Sciences, Universidade Estadual de Santa Cruz, Ilhéus, BA, 45662-900, Brazil
ABSTRACT
This article summarizes experimental knowledge of the efficacy, modes of action and applications of herbal extracts and organic acids as feed additives for pigs. Herbal extracts comprise a wide variety of herbs, spices and, most importantly, essential oils. Organic acids, characterized as short-chain weak acids, are widely distributed in nature as normal constituents of herbal or animal tissues. Herbal extracts and organic acids can stimulate feed intake and the production of endogenous secretions and nutrient absorption, protect against the colonization of pathogenic microorganisms in the animal gut, reduce the fermentation process and production of toxic metabolites, and present a beneficial effect on the intestinal microbiota. Therefore, these compounds could replace antibiotics as growth promoters for pig production. However, a systematic approach to the efficacy and safety of herbal extracts and organic acids as feed additives for swine is still non-existent.
Keywords: Antimicrobial, botanicals, organic acids, swine
1. Introduction
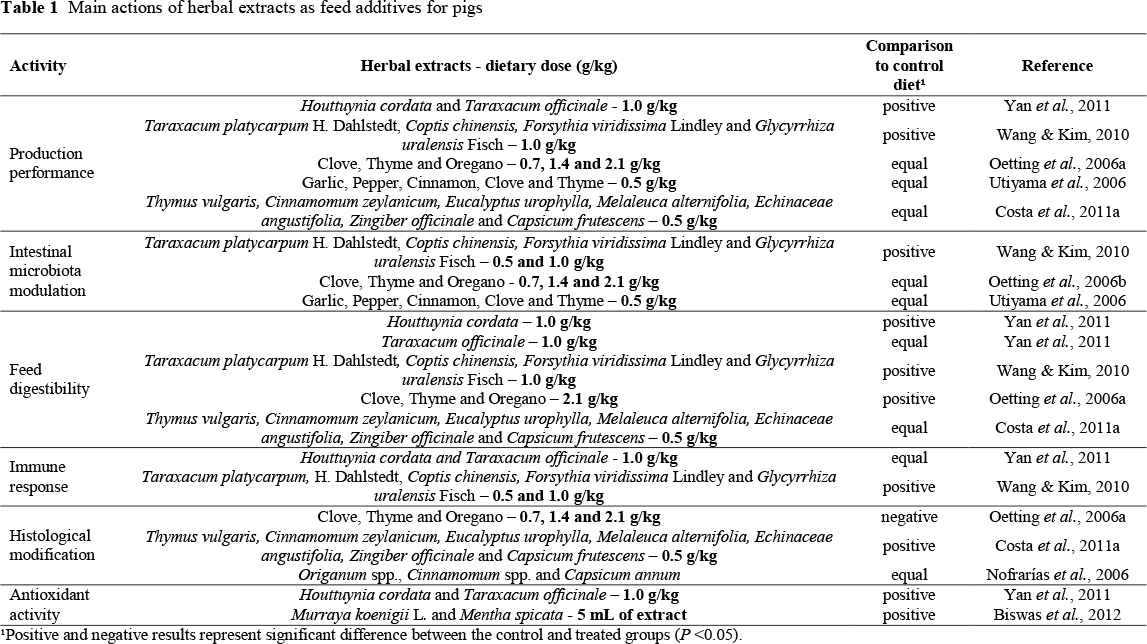
Antimicrobial properties of medicinal and aromatic herbals and their extracts have been observed since ancient times, but it was only in the early 1900s that these activities were first characterized in a laboratory. The benefits of herbal extracts to animals may be related to the increase in digestive secretions, improvement of digestibility and absorption of nutrients, modification of intestinal microbiota, stimulation of the immune system and antibacterial activities, coccidiostatic, anthelmintic, anti-viral or anti-inflammatory and antioxidant properties (Costa et al., 2007; 2011a).
Organic acids, which are characterized as weak and short-chain acids, are widely distributed in nature as constituents of herbal and animal tissues. However, these compounds are also obtained through the fermentation of carbohydrates, which is a common occurrence in the large intestine of pigs. Organic acids and their salts have the ability to lower the gastric pH, resulting in an increased gastric retention time and improved activity of proteolytic enzymes. They can also reduce the diet's buffering capacity, inhibit the proliferation and colonization of undesirable microorganisms in raw materials, feed and gastrointestinal tract of animals, act on the physiology of the gastrointestinal mucosa, and ease the availability of nutrients in the diet, improving their digestion, absorption and retention (Costa et al., 2011a).
Herbal extracts and organic acids have been tested in the diets of pig as potential alternatives to antibiotics on growth promotion. However, many questions about the effectiveness of this substitution still need to be clarified. Therefore, the aim of this review is to research and evaluate recent information on the use of these additives, with the emphasis on their modes of action to improve pig performance.
2. Herbal extracts
Herbal extracts are defined as herbal-derived compounds that are incorporated into animal diets in order to promote growth performance and the quality of products. They are classified according to their origins and processes: extracts, spices, essential oils (lipophilic compounds extracted by steam distillation of the ground herbals) and oleoresins (compounds extracted by non-aqueous solvents) (Windisch et al., 2007).
Modern laboratory techniques enable the isolation and characterization of bioactive substances from herbal sources. Bioactive substances are chemical compounds that are present in all parts of an herbal or are found in specific locations, which give the herbal material a therapeutic activity. Moreover, these molecules usually have a low molecular weight and are derived from the secondary metabolism of herbals.
The bioactive principles vary widely in concentration and antibacterial activity from one botanical species to another, but few have presented similar growth-promoting activity in comparison with the classic antibiotics. These compounds, for example capsaicin (pepper), cinnamaldehyde (cinnamon), eugenol (clove) and carvacrol (oregano), are produced in the plant as defence mechanisms against external factors such as physiological stress (that is, lack of water or nutrients), environmental factors (climate changes) and protection from predators and pathogens (Oetting et al., 2006a).
The content of these active substances in herbal extracts varies greatly, depending on the herbal part (seeds, leaves, stems, roots or bark), geographic origin, herbal maturity stage, preservation methods and period, storage and extraction methods (Windisch et al., 2007). Usually, these substances are not present in a pure state within the herbal cells, but in the form of complexes, which in some instances have better antimicrobial activity in the animal body.
Recent research has demonstrated the existence of a synergistic effect between the primary and secondary active principles of herbals. For example, in oregano, the carvacrol is present in a high concentration (primary active principle) and the thymol in a lower concentration (secondary active principle). The secondary components, which are active principles that are found in small concentrations, act as enhancers of the primary compounds (Kamel, 2000). However, antagonistic effects have also been observed between the active principles present in phytogenic compounds, as well as between the various additives used as promoters of animal growth (Costa et al., 2011b). Different combinations or different levels of herbal extracts must be considered in order to have control of the balance between synergism and antagonism, as well as animal health and growth versus possible toxicity of the additives, which might reflect directly in reduced or improved animal performance (Mellor, 2000).
The elucidation of the mode of action will provide the scientific basis to establish the efficacy and safety of these additives, thus allowing the development of long-term strategies in formulating specific and improved diets. Only then, would the industry and consumers benefit from the use of these products and utilize them as standard practice, similar to the antibiotics used as growth promoters.
2.1. Modes of action of herbal extracts
The possible mechanisms of action of herbal extracts in the animal organism for growth promotion include changes in the intestinal microbiota (Dorman & Deans, 2000); increased digestibility and nutrient absorption (Oetting et al., 2006a); enhanced nitrogen absorption (Alcicek et al., 2004); improvement of the immune response (Namkung et al., 2004); morphological and histological modifications of the gastrointestinal tract (Utiyama et al., 2006); and antioxidant activity (Botsoglou et al., 2002b; 2004). However, most of these studies were performed in controlled laboratorial conditions, and in practice most herbal extracts should be included in higher doses to the animal diet than those used in vitro to present the same bactericidal or bacteriostatic effect. Therefore, in the animal, the mode of action and place of action of the active components in herbal extracts (phyto-components or phyto-molecules) depend on their structure, metabolism and level of inclusion (Costa et al., 2011a).
2.2. Antimicrobial activity
Herbal extracts and metabolites are known to exert an in vitro antimicrobial effect against many zoonotic and foodborne pathogens (Essawi & Srour, 2000; Nascimento et al., 2000; Araújo & Leon, 2001; Harris et al., 2001; Karanika et al., 2001; Salvat et al., 2001; Srinivasan et al., 2001; Vieira et al., 2001; Si et al., 2006; Ozer et al., 2007).
Studies based on minimum inhibitory concentration (MIC) methods have demonstrated that herbal extracts present similar effects to most commercial antibiotics (Kamel, 2000); but need to be added in greater concentrations to demonstrate their effects (Burt, 2004). Many studies have shown that herbal extracts and isolated bioactive compounds have a wide spectrum of in vitro bactericidal activity. The mechanism by which the majority of herbal extracts exert their antibacterial effect is by acting in the bacterial cell wall structure, denaturing and coagulating proteins. More specifically, the essential oils act by altering the permeability of the cytoplasmic membrane to hydrogen ions (H+) and potassium (K+). This change leads to the disruption of essential cellular processes such as electron transport, protein translocation, oxidative phosphorylation and other enzyme-dependent reactions, resulting in the loss of chemiosmotic control and, consequently, in bacterial death (Dorman & Deans, 2000). According to these authors, the disruption of the bacterial cytoplasmic membrane is owing to the lipophilic nature of essential oils that accumulate in the membranes. Gram-negative bacteria have an outer membrane that contains lipopolysaccharides, which form a hydrophilic surface that creates a barrier to the permeability of hydrophobic substances such as essential oils. This could explain the higher resistance of gram-negative bacteria to the antimicrobial effect of some essential oils, compared with gram-positive bacteria (Oetting et al., 2006b). Other mechanisms of action may be related to the inhibition of nutrient absorption, enzymatic inhibition, synthesis of DNA, RNA and synthesis of proteins by the bacterial cells (Lambert et al., 2001; Luciano & Holley, 2009).
The effects of herbal extracts on microorganisms have been demonstrated in vitro. Some in vivo studies with pigs (Jug-Chizzola et al., 2005; Hagmüller et al., 2006; Li et al., 2012; Yan et al., 2012) have demonstrated the antimicrobial potential of herbal extracts to replace antibiotics in feeds. However, the data are too limited to allow reliable conclusions on the efficacy of phytogenic feed additives.
2.3. Influence on diet palatability and gut functions
Herbal extracts are often used in animal diets to improve palatability and productive performance in particular. However, studies testing the palatability of the diets with the addition of phytogenics are limited. Most studies have observed only the effect of phytogenics on feed intake and weight gain through performance tests (Biagi et al., 2006; 2007; Oetting, et al., 2006a; Costa et al., 2011). Schöne et al. (2006) and Yan et al. (2012) reported reduced palatability of the diet in weaned piglets as a result of the inclusion of essential oils. The authors related the lower feed intake to the high level of essential oils in this study and to the organoleptic properties of the herbal extract mixture. Essential oils may have strong odours or sharp flavours if used in high concentrations. On the other hand, essential oils in lower doses may contribute to feed intake (Costa et al., 2007), improving flavour and palatability, and resulting in higher feed consumption by pigs. The initial effect of adding herbals to feed for pigs is the stimulation of appetite. The aroma of these additives can excite the olfactory nerves and taste buds, resulting in a positive effect on feed intake.
Herbal extracts have shown significant benefits in the digestive tract of rats, producing laxative effects and preventing flatulence, stimulating digestive secretions (bile and mucus), and increasing enzymatic activity (Platel & Srinivasan, 2004). Many herbal-derived additives have active ingredients that increase salivary gland secretion and gastric and pancreatic juices (Mellor, 2000). Capsaicin, the active component of Capsicum annum (cayenne pepper), has been shown to be effective in stimulating salivation (producing amylase) (Platel & Srinivasan, 1996) and increasing the secretion of pancreatic and intestinal enzymes in non-ruminants (Costa et al., 2011a). Another active ingredient, cinnamaldehyde, the main component of Cinnamomum spp. (cinnamon), showed a stimulating effect on pancreatic enzymes and increased feed retention time in the stomach owing to the promotion of reduced gastric motility in pigs (Manzanilla et al., 2004). As a result, the increased activity of digestive enzymes promoted improvement in the digestibility and availability of nutrients (Costa et al., 2011a)
Herbal extracts promote gastric secretion, which activates pepsin. In addition, they increase the production of pancreatic juice, which leads to increased activity of digestive enzymes (for example trypsin, chymotrypsin, carboxypeptidase and lipase). Although those mechanisms that help to explain the improvement in feed digestibility generated by the phytoactive compounds have been studied most, other mechanisms might be involved. Modulation of the gut microbiota and maintenance of intestinal epithelium integrity may be important effects that are exerted by herbal extracts, as happens with other growth promoters.
During the digestion process, oxygen radicals (superoxide radicals) can be produced by an event known as autoxidation. Such reactive oxygen radicals can attack the surface of the intestinal mucosa, impairing the absorption of nutrients. Enzymes such as superoxide dismutase, glutathione S-transferase and catalase are responsible for converting superoxide radicals into water and molecular oxygen, thus maintaining intestinal health and allowing greater absorption of nutrients by the microvilli (Costa et al., 2007).
Herbal extracts seem to stimulate mucus secretion in the stomach and intestine. This effect prevents the adhesion of pathogens and contributes to the stabilization of favourable microbiota, protecting the intestinal villi and, therefore, improving the digestion and absorption of nutrients (Jamroz et al., 2005; 2006).
2.4. Antioxidant activity
Lipid oxidation of meat and meat products leads to the formation of unpleasant flavours, decreasing product acceptance by consumers (Lee & Shibamoto, 2002). In recent years, the potent antioxidant activity of herbal extracts has attracted interest from the food industry (Racanicci et al., 2004). These can be added to meat and meat products and replace synthetic products such as BHT (butylated hydroxytoluene) and BHA (butylated hydroxyanisole), two antioxidants that are widely used in animal feed.
Oxygen is essential to the metabolism and growth of animals and herbals. However, it is also responsible for uncontrolled oxidation reactions, known as autoxidation. These reactions cause the destruction of essential molecules in animal diet and damage to cellular tissues in living organisms. Autoxidation is responsible for the formation of reactive oxygen species (ROS), which cause a wide variety of diseases (Kamel, 2000).
The antioxidant activity of essential oils is related mainly to the presence of phenolic compounds. However, other compounds such as flavonoids (found in oregano and thyme) and terpenoids (such as thymol, carvacrol and eugenol, the active principles of thyme, oregano and cloves, respectively) protect food, tissues and cells against the deleterious effect of autoxidation reactions. These compounds can scavenge and neutralize free radicals. Moreover, they can chelate iron and copper ions, preventing the spread of the oxidation process (Kamel, 2000; Oetting et al., 2006a). Essential oils, after they have been absorbed systemically, are distributed and subsequently retained in animal tissues in small concentrations that already exert significant antioxidant activity (Botsoglou et al., 2004).
Although they possess strong antioxidant activity, some herbal extracts are made up of compounds with extremely sharp aromas, such as eugenol, thymol and carvacrol. Thus, their addition to animal feed in large quantities must be avoided, especially for pigs because of their sensitive palates (Madsen et al., 1997). This may limit their use as antioxidants in certain products.
Several in vitro studies have investigated the antioxidant action of herbals and satisfactory results have been observed (Racanicci et al., 2004; 2008; Juntachote et al., 2006; Rodtjer et al., 2006; Marianne et al., 2007). Thus, a new line of research has emerged to assess whether the antioxidant effect in meat can be increased by supplementing animal diets with herbal extracts and essential oils in. Feed supplementation with rosemary and sage extracts has shown positive effects against lipid oxidation (Kamel, 2000). Studies with oregano essential oil have also shown this effect (Botsoglou et al., 2002a; b; 2003a; b; 2004; Papageorgiou et al., 2003; Young et al., 2003), contributing to a longer shelf life of meat products.
Lopez-Bote et al. (1998) studied the addition of sage and rosemary extracts to the diet of broilers. The authors found that supplementation with the extracts altered the composition of body fatty acids in these animals. Sage and rosemary extract increase the percentage of polyunsaturated fatty acids (PUFAs). Higher levels of PUFAs may predispose the animal's organism to oxidative reactions. However, phenols, carotenoids and flavonoids, which are present in high concentrations in these herbals, can protect cells and tissues against the deleterious effects of ROS at the same level as vitamin E (Kamel, 2000), compensating for the increase of PUFAs in the animal organism.
Some studies have shown that herbal extracts added to the animal diet can introduce natural antioxidants within the phospholipids of the plasmatic membrane, where they may effectively inhibit oxidative reactions, thus preserving meat products for longer periods (Lauridsen et al., 1997; Botsoglou et al., 2004). However, more studies should be carried out to confirm this activity, which may greatly benefit the meat-processing industry.
2.5. Other effects of herbal extracts
Some reports showed improvements in pig growth performance when they received a mixture of oregano, cinnamon and Mexican pepper (Matysiak et al., 2012) in their feed as replacements for antibiotics. This may be partially derived from the effect of these additives on controlling pathogenic microorganisms, which increases resistance to digestive disorders, especially in newly weaned piglets. With better intestinal health, the animals are less exposed to microbial toxins and other undesirable microbial metabolites such as ammonia and biogenic amines. In addition, these growth-promoting additives may help the immune response in critical situations and improve the availability of essential nutrients for absorption, enabling better use of the animal genetic potential for growth (Windisch et al., 2007).
A possible improvement in the digestive capacity of the small intestine is considered an indirect effect of herbal extracts through the stabilization of the intestinal microbiota. An increase in pre-caecal digestibility reduces the flow of fermentable material in the intestine and reduces post-ileal microbial growth and excretion of bacterial mass in the faeces. Owing to the representative participation of bacterial protein in faecal total protein, increased pre-caecal digestive capacity should result in improved digestibility of protein in the diet. This observation supports the hypothesis that herbal extracts should stabilize digestive functions (Windisch et al., 2007; Li et al., 2012).
Phytogenic compounds were found to modulate the intestinal microbiota of pigs (Manzanilla et al., 2004). A mixture of essential oils and formic acid was fed to early weaned pigs that were challenged by social and diet stressors. The natural compounds were able to decrease the total volatile fatty acid (VFA) content in the colon. VFAs are the main end-products of the bacterial metabolism in the large intestine of pigs, and the phytogenics acted on the gut microbiota, promoting their death or growth stabilization, reducing microbial fermentation and consequent production of VFAs. Interestingly, the experimental additives increased the lactobacilli : enterobacteria ratio in the gut, which may enhance the health status of the animals, since many pathogens belong to the second group of bacteria. The main actions of herbal extracts are shown in Table 1.
Currently in certain studies herbal extracts are tested as possible growth promoters for pigs. These studies, however, are not conclusive and do not effectively demonstrate their modes of action. Therefore, more studies should be carried out so that, in practice, herbal extracts can effectively replace antimicrobial growth promoters.
3. Organic acids
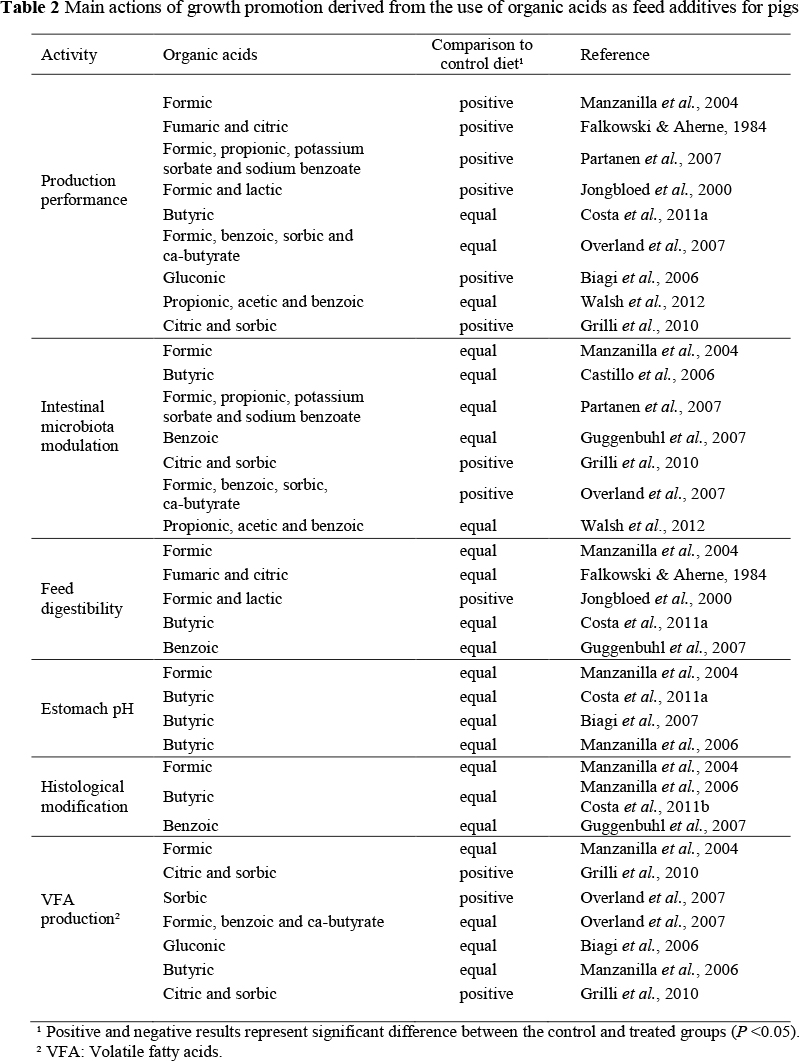
The organic acids, characterized as weak short-chain acids, are widely distributed in nature as natural constituents of herbals and animal tissues. Some may be formed by the fermentation of carbohydrates, predominantly in the large intestine, and others in the intermediary metabolism of pigs (Partanen & Mroz, 1999). Many are found commercially in the form of salts, with fewer undesirable odours and greater ease of use in feed manufacturing because they are solid, less volatile and less corrosive.
Organic acids were first used as effective preservatives. Their primary bacteriostatic action against many microbial species occurs by reducing the pH of the diet. In addition, when the acid is not dissociated, it can diffuse freely through the microbial plasmic membrane and reach the cytoplasm. Once inside the cell, where the pH is kept near to 7, the acid will dissociate into ions and protons, reducing the cytoplasmic pH, causing cell death by inhibiting enzymatic reactions, and denaturating proteins and DNA (Cherrington et al., 1991). The antimicrobial effect of the acids can also be the result of the accumulation of anions in the cell. The efficiency of the acid in the inhibition of microorganisms depends on its pKa value, which is the pH where 50% of the acid is dissociated. Organic acids with high pKa values are more effective preservatives, and their antimicrobial activity is generally improved with the increase of the carbon chain length and level of unsaturation (Foegeding & Busta, 1991).
The absorption of the acid depends on its pKa and the environmental pH. When the lumen pH is lower than the pKa of the acids, they are quickly absorbed. As the intestinal pH is normally higher than the pKa of these acids, they remain predominantly in the dissociated form that is poorly absorbed. In the digestive tract, non-dissociated organic acids are absorbed by the intestinal epithelium by passive diffusion through the favourable electrochemical gradient between the lumen and epithelial cells (Partanen & Mroz, 1999).
The efficiency of organic acids on pathogenic bacteria depends on their time of exposure, concentration, the type of acid, the composition of the basal diet, and the age of the animal. Gram-negative bacteria are sensitive to acids with carbon chains with fewer than eight carbon atoms, while gram-positive bacteria are more sensitive to acids with longer carbon chains (Partanen & Mroz, 1999). The use of organic acids varies among countries, depending on differences in legislation and other factors. In some countries, acids are used to decontaminate microorganisms in feed ingredients prior to heat treatment, while in others acids are added primarily to finished feed or drinking water (Koyuncu et al, 2013).
3.1. Modes of action of organic acids
The successful use of organic acids in the diets of pigs requires understanding of their mechanisms of action. Although some hypotheses have been proposed, their modes of action have not been clarified. First, it is considered that organic acids and their salts reduce gastric pH, which is essential to limit pathogen survival in the stomach, thus preventing access to the intestine (Partanen & Mroz, 1999). However, the factor that limits the efficacy of these acids that they must reach the intestine to exert their antibacterial activity, without being absorbed too rapidly on leaving the stomach (Grilli et al., 2010).
This reduction in gastric pH results in shortened intestinal transit by increasing gastric retention and enhances the activity of proteolytic enzymes. Owing to the lower rate of gastric depuration, protein molecules can be hydrolyzed more efficiently, which results in a beneficial effect on digestion, absorption and retention of amino acids (Gabert & Sauer, 1994). Some authors (Thomlinson & Lawrence, 1981; Bolduan et al., 1988a; b; Risley et al, 1992) found a lower gastric pH in piglets when these animals received organic acids in their diet, leading to increased activity of pepsin and consequently higher protein digestion. However, it is hard to compare studies that aim to determine the reduction in gastric pH of animals that receive organic acids as feed additives, because of the wide variation in methods and the difficulty in measuring pH in the stomach.
Studies have produced strong evidence that organic acids per se stimulate endocrine and exocrine pancreatic secretion. Intestinal acidification increases the contents of serum secretion, which stimulates exocrine pancreatic secretion and bile secretion (Partanen & Mroz, 1999). Thaela et al. (1998) showed that supplementation of 2.5% lactic acid in the diets of weaning pigs increased the volume and protein content of pancreatic juice, as well as the secretion of trypsin and chymotrypsin. However, the amount of bicarbonate produced in the animal intestine was not affected by the supplementation with lactic acid. For this reason it is unlikely that the stimulation of pancreatic secretion was caused by decreased dietary and gastric pH, and probably by the direct action of the acid.
In addition to lowering gastric pH and stimulating pancreatic secretion, supplementation with organic acids and their salts reduces the buffering capacity of the diet, which may lead to inhibition proliferation and colonization of undesirable microorganisms in raw materials, feeds and in the gastrointestinal tract of animals. However, diets may be resistant to changes in pH because of their high level of protein and minerals, which reduces the effect of acidifiers on animal growth promotion (Partanen & Mroz, 1999).
Organic acids may act on the intestinal microbiota and reduce the incidence of diarrhoea in weanling pigs, but not all acids are effective, and studies have found contradictory results. Knarreborg et al. (2002) studied the antimicrobial activity of six organic acids on the population of coliforms in the proximal part of the gastrointestinal tract of piglets and found significant differences among them, where the activity was benzoic acid > fumaric acid > lactic acid > butyric acid > formic acid > propionic acid.
These acids acted against the population of pathogenic and beneficial bacteria, reducing microbial fermentation and the consequent production of VFAs. On the other hand, Partanen & Mroz (1999) showed that acidic conditions favour the growth of lactobacilli in the stomach, which inhibits the proliferation of E.
coli by blocking adhesion sites or by producing lactic acid and other metabolites that decrease pH and affect the growth of this bacterium. In other experiments investigating the population of microorganisms in the intestine of weanling pigs fed with organic acids, Partanen & Mroz (1999), Piva et al. (2002) and Namkung et al. (2004) observed a modulation of the microbiota, in which beneficial microorganisms grew at a much faster pace than pathogenic ones.
Organic acids have been shown to improve the diet digestibility and nutrient retention in pigs. In general, the acids exert a small but significant effect on the apparent digestibility in the whole digestive tract and retention of protein (amino acids) and energy. However, when the buffering capacity of the diet is high, the efficiency of organic acids in improving the digestibility of amino acids is reduced, requiring high levels to express its effect. The level of dietary fibre influences the apparent ileal digestibility of amino acids, masking the effect of organic acids, reducing their effects on digestibility (Mroz et al., 1998).
The increase in energy digestibility is the result of a rise in the apparent digestibility of crude protein, fat and nitrogen. The acids act on the microorganisms by reducing their metabolic activities, which results in increased fat digestibility by the animal, and consequently influences th energy digestibility positively. These acids also reduce bacterial growth, decreasing the use of amino acids for bacterial protein formation, therefore promoting greater nitrogen absorption by the animal (Partanen & Mroz, 1999).
Supplementing the diet with minerals, calcium and phosphorus in particular can increase the buffering capacity of the digesta in the stomach, allowing pathogen growth and reducing the digestibility of the diet. Organic acids can lower the buffering capacity of the digesta and improve the absorption of calcium and phosphorus (Partanen & Mroz, 1999). Another factor that influences the absorption of minerals, particularly phosphorus, is the amount of intrinsic or microbial phytase present in the diet (Rutherfurd et al., 2012). This enzyme is able to degrade phytic acid, which releases inorganic phosphorus, and consequently enhances its bioavailability. Organic acids have been shown to improve the performance of phytase, promoting better absorption of phosphorus, thus providing lower excretion and lower environmental pollution (Partanen & Mroz, 1999).
Organic acids may influence the physiology of the intestinal mucosa. They act on the villi, maintaining their integrity, promoting an increase in the number of cells and preventing its flattening, as well as serving as a substrate in the intermediary metabolism of the citric acid-cycle (Partanen & Mroz, 1999).
Several studies reported the performance of piglets that received organic acids in their diet (Omogbenigun et al., 2003; Namkung et al., 2004; Biagi et al., 2006; 2007; Manzanilla et al., 2006), but the results are inconsistent and contradictory. Organic acids are not equal in their efficacy to combat infection and enhance growth when administered to pigs via feed or water (Walsh et al., 2012). The main actions of growth promotion derived from the use of organic acids as feed additives are summarized on Table 2.
Organic acid supplementation appears to be more effective during the first two to four weeks after weaning, when the animal's immune and digestive systems are still immature, whereas its effect on pigs was shown to decline in the growing and finishing phases (Giesting et al., 1991). The type and concentration of acid determine their efficacy. Ravindran & Kornegay (1993) reported that a better response in performance can be obtained in conditions that are less favourable to the growth of piglets, such as in high-challenge and less complex diets.
4. Conclusion
Animal welfare and growth performance are the main reasons for the widespread use of animal feed additives. Antibiotics, chemotherapeutic agents and, more recently, organic acids, herbals and their extracts are the main additives that have been used to stabilize the digestive function in pig breeding worldwide.
Many traditional antibiotics have been banned from animal feed owing to the trend of incorporating more "natural" animal production systems. Therefore, the livestock industry is searching for environmentally friendly supplements with higher acceptance by the consumer. Herbal extracts and organic acids are claimed to exert antioxidative and antimicrobial activities, reduce the production of fermentation products, which may reflect in improved overall gut health, and stimulate nutrient absorption.
The current experience in using these natural compounds as additives in pig feed seems to justify the assumption that they would improve growth performance and animal productivity, and thus could be added to the set of non-antibiotic/chemotherapeutic growth promoters. However, studies do not show consistent results and this may be explained by the differences in the quality of plant materials, selection of particular herbs, extraction and concentration methods, and forms of administration.
Similar to herbal extracts, not all organic acids have shown constant results in the literature, and the methodology used in these studies varies widely. The concentration of acid, its type and combinations are important in improving animal performance. To date, formic, citric and sorbic acids have shown the best results in pig performance and production of the organic acids that are used as feed additives.
Reference
Alçiçek, A., Bozkurt, M. & Çabuk, M., 2004. The effect of a mixture of herbal essential oils, an organic acid or a probiotic on broiler performance. S. Afr. J. Anim. Sci. 34, 217-222. [ Links ]
Araújo, C.A.C. & Leon, L.L., 2001. Biological activities of Curcuma longa L. Memórias do Instituto Oswaldo Cruz, Rio de Janeiro, 96 (5), 723-728. [ Links ]
Biagi, G., Piva, A., Moschini, M., Vezzali, E. & Roth, F.X., 2006. Effect of gluconic acid on piglet growth performance, intestinal microflora and intestinal wall morphology. J. Anim. Sci. 84, 370-378. [ Links ]
Biagi, G., Piva, A., Moschini, M., Vezzali, E. & Roth, F.X., 2007. Performance, intestinal microflora, and wall morphology of weanling pigs fed sodium butyrate. J. Anim. Sci. 85, 1184-1191. [ Links ]
Biswas, A.K., Chatli, M.K. & Sahoo, J., 2012. Antioxidant potential of curry (Murraya koenigii L.) and mint (Mentha spicata) leaf extracts and their effect on colour and oxidative stability of raw ground pork meat during refrigeration storage. Food Chem. 133, 467-472. [ Links ]
Bolduan, G., Jung, H., Schneider, R., Block, J. & Klenke, B., 1988a. Influence of propionic and formic acids on piglets. J. Anim. Physiol. Anim. Nutr. 59, 72-78. [ Links ]
Bolduan, G., Jung, H., Schneider, R., Block, J. & Klenke, B., 1988b. Influence of fumaric acid and propanediol formate on piglets. J. Anim. Physiol. Anim. Nutr. 59, 143-149. [ Links ]
Botsoglou, N.A., Christaki, E., Fletouris, D.J., Florou-Paneri, P. & Spais, A.B., 2002a. The effect of dietary oregano essential oil on lipid oxidation in raw and cooked chicken during refrigerated storage. Meat Sci. 62, 259-265. [ Links ]
Botsoglou, N.A., Florou-Paneri, P., Christaki, E., Fletouris, D.J. & Spais, A.B., 2002b. Effect of dietary oregano essential oil on performance of chickens and on iron-induced lipid oxidation of breast, thigh and abdominal fat tissues. Br. Poult. Sci. 43, 223-230. [ Links ]
Botsoglou, N.A., Fletouris, D.J., Florou-Paneri, P., Christaki, E. & Spais, A.B., 2003a. Inhibition of lipid oxidation in long-term frozen stored chicken meat by dietary oregano essential oil and α-tocopheryl acetate supplementation. Food Res. Int. 36, 207-213. [ Links ]
Botsoglou, N.A., Govaris, A., Botsoglou, E.N., Grigoropoulou, S.H. & Papageorgiou, G., 2003b. Antioxidant activity of dietary oregano essential oil and r-tocopheryl acetate in long-term frozen stored turkey meat. J. Agri. Food Chem. 51, 2930-2936. [ Links ]
Botsoglou, N.A., Florou-Paneri, P., Christaki, E., Giannenas, I. & Spais, A.B., 2004. Performance of rabbits and oxidative stability of muscle tissues as affected by dietary supplementation with oregano essential oil. Arch. Anim. Nutr. 58, 209-218. [ Links ]
Burt, S., 2004. Essential oils: their antibacterial properties and potential applications in foods - a review. Int. J. Food Microbiol. 94, 223-253. [ Links ]
Castillo, M., Martín-Orúe, S.M., Roca, M., Manzanilla, E.G., Badiola, I., Perez, J.F. & Gasa J., 2006. The response of gastrointestinal microbiota to avilamycin, butyrate, and herbal extracts in early-weaned pigs. J. Anim. Sci. 84, 2725-2734. [ Links ]
Cherrington, C.A., Hinton, M., Mead, G.C. & Chopra, I., 1991. Organic acids: chemistry, antibacterial activity and practical applications. Advant. Microb. Physiol. 32, 87-108. [ Links ]
Costa, L.B., Tse, M.L.P. & Miyada, V.S., 2007. Herbal extracts as alternatives to antimicrobial growth promoters for newly weaned piglets. Rev. Bras. de Zootec. 36 (3), 589-595. [ Links ]
Costa, L.B., Berenchtein, B., Almeida, V.V., Tse, M.L.P., Braz, D.B., Andrade, C., Mourao, G.B. & Miyada, V.S., 2011a. Phytogenic additives and sodium butyrate as growth promoters of weaned piglets. Arch. de Zootec. 60, 687-698. [ Links ]
Costa, L.B., Almeida, V.V., Berenchtein, B, Tse, M.L.P., Andrade, C. & Miyada, V.S., 2011b. Phytogenic additives and sodium butyrate as alternatives to antibiotics for weaned piglets. Arch. de Zootec. 60, 733-744. [ Links ]
Dorman, H.J.D. & Deans, S.G., 2000. Antimicrobial agents from herbals: antibacterial activity of herbal volatile oils. J. Appl. Microbiol. 88, 308-316. [ Links ]
Essawi, T. & Srour, M., 2000. Screening of some Palestinian medicinal herbals for antibacterial activity. J. Ethnopharmacol. 70, 343-349. [ Links ]
Falkowski, J.F. & Aherne, F.X., 1984. Fumaric and citric acid as feed additives in starter pig nutrition. J. Anim. Sci. 58, 935-938. [ Links ]
Foegeding, P.M. & Busta, F.F., 1991. Chemical food preservatives. In: Disinfection, sterilization and preservation. Eds: Foegeding, P.M., Busta, F.F. & Block, S.S., Philadelphia, PA, USA. pp. 802-832. [ Links ]
Gabert, V.M. & Sauer, W.C., 1994. The effects of supplementing diets for weanling pigs with organic acids. J. Anim. Feed Sci. 3, 73-87. [ Links ]
Giesting, D.W., Roos, M.A. & Easter, R.A., 1991. Evaluation of the effect of fumaric acid and sodium bicarbonate addition of performance of starter pigs fed diets of different types. J. Anim. Sci. 69, 2489-2496. [ Links ]
Grilli, E., Messina, M.R., Tedeschi, M. & Piva, A., 2010. Feeding a microencapsulated blend of organic acids and nature identical compounds to weaning pigs improved growth performance and intestinal metabolism. Livest. Sci. 133, 173-175. [ Links ]
Guggenbuhl, P., Séon, A., Piñón Quintana, A. & Simões Nunes, C., 2007. Effects of dietary supplementation with benzoic acid (VevoVitall®) on the zootechnical performance, the gastrointestinal microflora and the ileal digestibility of the young pig. Livest. Sci. 108, 218-221. [ Links ]
Hagmüller, W., Jugl-Chizzola, M., Zitterl-Eglseer, K., Gabler, C., Spergser, J., Chizzola, R. & Franz, C., 2006. The use of Thymi herba as feed additive (0.1%, 0.5%, 1.0%) in weanling piglets with assessment of the shedding of haemolysing E. coli and the detection of thymol in the blood plasma. Berliner und Münchener tierärztliche Wochenschrift, Berlin. 119, 50-54. [ Links ]
Harris, J.C., Cottrel, S.L., Plummer, S. & Lloyd, D., 2001. Antimicrobial properties of Allium sativum (garlic). Appl. Microbiol. Biotechnol. 57, 282-286. [ Links ]
Jamroz, D., Wiliczkiewicz, A., Wertelecki, T., Orda, J. & Skorupinska, J., 2005. Use of active substances of herbal origin in chicken diets based on maize and domestic grains. Br. Poult. Sci. 46, 485-493. [ Links ]
Jamroz, D., Wertelecki, T., Houszka, M. & Kamel, C., 2006. Influence of diet type on the inclusion of herbal origin active substances on morphological and histochemical characteristics of the stomach and jejunum walls in chicken. J. Anim. Physiol. Anim. Nutr. 90, 255-268. [ Links ]
Jongbloed, A.W., Mroz, Z., Van der Weij-Jongbloed, R. & Kemme, P.A., 2000. The effects of microbial phytase, organic acids and their interaction in diets for growing pigs. Livest. Prod. Sci. 67, 113-122. [ Links ]
Juntachote, T., Berghofer, E., Siebenhandl, S. & Bauer, F., 2006. The antioxidative properties of Holy basil and Galangal in cooked ground pork. Meat Sci. 72, 446-456. [ Links ]
Kamel, C., 2000. A novel look at a classic approach of herbal extracts. Feed Mix - The Int. J. Feed Nutr. Technol. 9 (6), 19-24. [ Links ]
Karanika, M.S., Komaitis, M. & Aggelis, G., 2001. Effect of aqueous extract of some herbals of Lamiaceae family on the growth of Yarrowia lipolytica. Int. J. Food Microbiol. 64, 175-181. [ Links ]
Knarreborg, A., Miquel, N., Granli, T. & Jensen, B.B., 2002. Establishment and application in vitro methodology to study the effects of organic acids on coliform and lactic acid bacteria in the proximal part of the gastrointestinal tract of piglets. Anim. Feed Sci. Technol. 99, 131-140. [ Links ]
Koyuncu, S., Andersson, M.G., Charlotta L., Skandamis P.N., Gounadaki, A., Zentek, J. & Hággblom, P., 2013. Organic acids for control of salmonella in different feed materials. BM Vet. Res. 9, 81-89. [ Links ]
Lambert, R.J.W., Skandamis, P.N., Coote, P.J. & Nychas, G.J.E., 2001. A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J. Appl. Microbiol. 91, 453-462. [ Links ]
Lauridsen, C., Buckley, D.J. & Morrissey, P.A., 1997. Influence of dietary fat and vitamin E supplementation on α-tocopherol levels and fatty acid profiles in chicken muscle membranal fractions and on susceptibility to lipid peroxidation. Meat Sci. 46, 9-22. [ Links ]
Lee, K.G. & Shibamoto, T., 2002. Determination of antioxidant potential of volatile extracts isolated from various herbs and spices. J. Agric. Food Chem. 50, 4947-4952. [ Links ]
Li, P., Piao, X., Ru Y., Han Xu., Xue L. & Zhang H., 2012. Effects of adding essential oil to the diet of weaned pigs on performance, nutrient utilization, immune response and intestinal health. Asian-Aust. J. Anim. Sci. 25, 1617-1626. [ Links ]
Lopez-Bote, C.J., Gray, J.I., Gomaa, E.A. & Flegal, C.J., 1998. Effect of dietary administration of oil extracts from rosemary and sage on lipid oxidation in broiler meat. Br. Poult. Sci. 39, 235-240. [ Links ]
Luciano, F.B. & Holley, R.A., 2009. Enzymatic inhibition by allyl isothiocyanate and factors affecting its antimicrobial action against Escherichia coli O157:H7. Int. J. Food Microbiol. 131, 240-245. [ Links ]
Madsen, H.L., Bertelsen, G. & Skibsted, L.H., 1997. Antioxidative activity of spice extracts. In: Spices, flavor, chemistry and antioxidant properties. Eds: Risch, S.J. & Ho, S.C.T., American Chemical Society, Washington, D.C., USA. pp. 176-187. [ Links ]
Manzanilla, E.G., Perez, J.F., Martin, M., Kamel, C., Baucells, F. & Gasa, J., 2004. Effect of herbal extracts and formic acid on intestinal equilibrium of early-weaned pigs. J. Anim. Sci. 82, 3210-3218. [ Links ]
Manzanilla, E.G., Nofrarías, N., Anguita, M., Castilho, M., Perez, J.F., Martín-Orúe, S.M., Kamel, C. & Gasa, J., 2006. Effects of butyrate, avilamycin, and a herbal extract combination on the intestinal equilibrium of early-weaned pigs. J. Anim. Sci. 84, 2743-2751. [ Links ]
Marianne, N.L., Marchen, S. & Leif, H.S., 2007. The combined effect of antioxidants and modified atmosphere packaging on protein and lipid oxidation in beef patties during chill storage. Meat Sci. 76, 226-233. [ Links ]
Matysiak, B., Jacyno, E., Kawecka, M., Kolodziej-Skalska, A. & Pietruszka, A., 2012. The effect of plant extracts fed before farrowing and during lactation on sow and piglet performance. S. Afr. J. Anim. Sci. 42, 15-21. [ Links ]
Mellor, S., 2000. Alternatives to antibiotic. Pig Prog. 16, 18-21. [ Links ]
Mroz, Z., Grela, E.R., Krasucki, W., Kies, A.K. & Schoner, F.J., 1998. Microbial phytase in combination with formic acid for reproductive sows. J. Anim. Sci. 76, Suppl. 1, Abstract 177. [ Links ]
Namkung, H., Li, M., Gong, J., Yu, H., Cottrill, M. & De Lange, C.F.M., 2004. Impact of feeding blends of organic acids and herbal extracts on growth performance, gut microbiota and digestive function in newly weaned pigs. Can. J. Anim. Sci. 84, 697-704. [ Links ]
Nascimento, G.G.F., Locatelli, J., Freitas, P.C. & Silva, G.L., 2000. Antibacterial activity of herbal extracts and phytochemicals on antibiotic-resistant bacteria. Braz. J. Microbiol. 31, 247-256. [ Links ]
Nofrarías, M., Manzanilla, E.G., Pujols, J., Gibert, X., Majó, N., Segalés, J. & Gasa, J., 2006. Effects of spray-dried porcine plasma and herbal extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J. Anim. Sci. 84, 2735-2742. [ Links ]
Oetting, L.L., Utiyama, C.E., Giani, P.A., Ruiz, U.S. & Miyada, V.S., 2006a. Effects of herbal extracts and antimicrobials on apparent digestibility, performance, organs morphometry and intestinal histology of weanling pigs. Rev. Bras. de Zootec. 35, 1389-1397. [ Links ]
Oetting, L.L., Utiyama, C.E., Giani, P.A., Ruiz, U.S. & Miyada, V.S., 2006b. Effects of antimicrobials and herbal extracts on intestinal microbiology and diahrrea incidence in weanling pigs. Rev. Bras. de Zootec. 35, 2013-2017. [ Links ]
Omogbenigum, F.O., Nyachoti, C.M. & Slominski, B.A., 2003. The effect of supplementing microbial phytase and organic acids to a corn-soybean based diet fed to early-weaned pigs. J. Anim. Sci. 81, 1806-1813. [ Links ]
Overland, M., Kjos, N.P., Borg, M. & Sorum, H., 2007. Organic acids in diets for entire male pigs. Livest. Sci. 109, 170-173. [ Links ]
Ozer, H.M., Shokmen, M., Gulluce, M., Adiguzel, A., Sahin, F., Sokmen, A., Kilic, H. & Baris, O., 2007.
Chemical composition and antimicrobial and antioxidant activities of the essential oil and methanol extracts of Hippomarathun microcarpum (Bieb.) from Turkey. J. Agr. Food Chem. 55, 937-942.
Papageorgiou, G., Botsoglou N., Govaris, A., Giannenas, I., Iliadis, S. & Botsoglou, E., 2003. Effect of dietary oregano oil and a-tocopheryl acetate supplementation on iron-induced lipid oxidation of turkey breast, thigh, liver and heart tissues. J. Anim. Physiol. Anim. Nutr. 87, 324-335. [ Links ]
Partanen, K.H. & Mroz, Z., 1999. Organic acids for performance enhancement in pig diets. Nut. Res. Rev. 12, 117-145. [ Links ]
Partanen, K., Siljander-Rasi, H., Pentikáinen, J., Pelkonen, S. & Fossi, M., 2007. Effects of weaning age and formic acid-based feed additives on pigs from weaning to slaughter. Arch. Anim. Nutr. 61, 336-356. [ Links ]
Piva, A., Casadei, G. & Biagi, G., 2002. An organic acid blend can modulate swine intestinal fermentation and reduce microbial proteolysis. Can. J. Anim. Sci. 82, 527-532. [ Links ]
Platel, K. & Srinivasan, K., 1996. Influence of dietary spices or their active principles on digestive enzymes of small intestinal mucosa in rats. Int. J. Food Sci. Nutr. 47, 55-59. [ Links ]
Platel, K. & Srinivasan, K., 2004. Digestive stimulant action of spices: A myth or reality? Indian J. Med. Res. 119, 167-179. [ Links ]
Racanicci, A.M.C., Danielsen, B, Menten, J.F.M., Regitano-D'arce, M.A.B. & Skibsted, L.K., 2004. Antioxidant effect of dittany (Origanum dictamnus) in pre-cooked chicken meat balls during chill-storage in comparison to rosemary (Rosmarinus officinalis). Eur. Food Res. Technol. 218, 521-524. [ Links ]
Racanicci, A.M.C., Danielsen, B. & Skibsted, L.H., 2008. Mate (Ilex paraguariensis) as a source of water extractable antioxidant for use in chicken meat. Eur. Food Res. Technol. 227, 255-260. [ Links ]
Ravindran, V. & Kornegay, E.T., 1993. Acidification of weaner pig diets: A review. J. Sci. Food Agric. 62, 313-322. [ Links ]
Risley, C.R., Kornegay, E.T., Lindemann, M.D., Wood, C.M. & Eigel, W.N., 1992. Effect of feeding organic acids on selected intestinal content measurements at varying times postweaning in pigs. J. Anim. Sci. 70, 196-206. [ Links ]
Rodtjer, A., Skibsted, L.H. & Andersen, M.L., 2006. Antioxidative and prooxidative effects of extracts made from cherry liqueur pomace. Food Chem. 99, 6-14. [ Links ]
Rutherfurd, S.M., Chung, T.K, Thomas, D.V., Zou, M.L. & Moughan, P.J., 2012. Effect of a novel phytase on growth performance, apparent metabolizable energy, and the availability of minerals and amino acids in a low-phosphorus corn-soybean meal diet for broilers. Poult. Sci. 91, 1118-1127. [ Links ]
Salvat, A., Antonnacci, L., Fortunato, R.H., Suarez, E.Y. & Godoy, H.M., 2001. Screening of some herbals from northern Argentina for their antimicrobial activity. Lett. Appl. Microbiol. 32, 293-297. [ Links ]
Schöne, F., Vetter, A., Hartung, H., Bergmann, H., Biertümpfel, A., Richter, G., Müller, S. & Breitschuh, G., 2006. Effects of essential oils from fennel (Foeniculi aetheroleum) and caraway (Carvi aetheroleum) in pigs. J. Anim. Physiol. Anim. Nutr. 90, 500-510. [ Links ]
Si, W., Gong, J., Tsao, R., Zhou, T., Yu, H., Poppe, C., Johnson, R. & Du, Z., 2006. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 100, 296-305. [ Links ]
Srinivasan, D., Nathan, S., Suresh, T. & Perumalsamy, P.L., 2001. Antimicrobial activity of certain Indian medicinal herbals used in folkloric medicine. J. Ethnopharmacol. 74, 217-220. [ Links ]
Thaela, M.J., Jensen, M.S., Cornelissen, G., Halberg, F., Noddegaard, F., Jakobsen, K. & Pierzynowski, S.G., 1998. Circadian and ultradian variation in pancreatic secretion of meal-fed pigs after weaning. J. Anim. Sci. 76, 1131-1139. [ Links ]
Thomlinson, J.R. & Lawrence, T.L.J., 1981. Dietary manipulation of gastric pH in the prophylaxis of enteric disease in weaned pigs: Some field observations. Vet. Rec. 109, 120-122. [ Links ]
Utiyama, C.E., Oetting, L.L., Giani, P.A., Ruiz, U.S. & Miyada, V.S., 2006. Effects of antimicrobials, prebiotics, probiotics and herbal extracts on intestinal microbiology, diahrrea incidence and performance of weanling pigs. Rev. Bras. de Zootec. 36 (6), 2359-2367. [ Links ]
Vieira, R.H.S.F, Rodrigles, D.P., Goncalvez, F.A, Menezes, F.G.R., Aragao, J.S. & Sousa, O.V., 2001. Microbicidal effect of medicinal herbal extract (Psidium guajava Linn and Carica papaya Lin) upon bacteria isolated from fish muscle and known to induce diarrhea in children. Ver. Inst. de Med. Trop. de Sao Paulo 43 (3), 145-148. [ Links ]
Wang, J.P. & Kim, I.H., 2010. Effects of herbal extracts on growth performance, fecal microbiota, nutrient digestibility, and blood profiles in weaning pigs. J. Anim. Sci. 12, 49-52. [ Links ]
Walsh, M.C., Rostagno, M.H., Gardiner, G.E., Sutton, A.L., Richert, B.T. & Radcliffe, J.S., 2011. Controlling salmonella infection in weanling pigs through water delivery of direct-fed microbials or organic acids. Part I: Effects on growth performance, microbial populations, and immune status. J. Anim. Sci. 90, 261-271. [ Links ]
Windisch, W.M., Schedle, K., Plitzner, C. & Kroismayr, A., 2007. Use of herbal extracts as feed additives for swine and poultry. J. Anim. Sci. 86, 140-148. [ Links ]
Yan, L., Meng, Q.W. & Kim, I.H., 2011. The effects of dietary Houttuynia cordata and Taraxacum officinale extract powder on growth performance, nutrient digestibility, blood characteristics and meat quality in finishing pigs. Livest. Sci. 141, 188-193. [ Links ]
Yan, L., Meng, Q.W. & Kim, I.H., 2012. Effect of an herb extract mixture on growth performance, nutrient digestibility, blood characteristics, and fecal microbial shedding in weanling pigs. Livest. Sci. 145, 189-195. [ Links ]
Young, J.F., Stagsted, J., Jensen, S.K., Karlsson, A.H. & Henckel, P., 2003. Ascorbic acid, α-tocoferol and oregano supplements reduce stress-induced deterioration of chicken meat quality. Poult. Sci. 82, 1343-1351. [ Links ]
Received 23 April 2013
Accepted 28 June 2013
First published online 16 July 2013
# Corresponding author: leandro_esalq@yahoo.com.br


{kind=link}
{kind=link}