










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
On-line version ISSN 2311-9284
Print version ISSN 0006-8241
Bothalia (Online) vol.53 n.1 Pretoria 2023
http://dx.doi.org/10.38201/btha.abc.v53.i1.11
ORIGINAL RESEARCH
The relationship between mammalian burrow abundance and bankrupt bush (Seriphium plumosum) encroachment
Morné OosthuysenI; W. Maartin StraussII; Michael J. SomersIII
IMammal Research Institute, University of Pretoria, Department of Zoology and Entomology, Pretoria, South Africa
IIABEERU, Department of Environmental Science, University of South Africa, Johannesburg, South Africa
IIIEugène Marais Chair of Wildlife Management, Mammal Research Institute, Centre for Invasion Biology, Department of Zoology and Entomology, University of Pretoria, Pretoria, South Africa
ABSTRACT
BACKGROUND: Much of the Grassland Biome in South Africa is prone to shrub encroachment, leading to loss of ecosystem services, habitat heterogeneity and species diversity. Burrowing mammals are an important component of grasslands as these animals create microhabitats for other taxa, including smaller mammal species, birds, reptiles and invertebrates. However, our understanding of how shrub encroachment affects burrowing mammals is poor
OBJECTIVES: Here we assessed the relationship between burrow abundance and bankrupt bush, Seriphium plumosum, encroachment as well as burrowing mammal diversity in bankrupt bush encroached and non-encroached grasslands
METHOD: Shrub density, medium and large mammal burrow abundance and density were measured in 24 encroached and 24 non-encroached areas randomly selected in the Telperion Nature Reserve, Mpumalanga, South Africa. In addition, burrowing mammal diversity was assessed using camera traps in a subset of six encroached and six non-encroached areas
RESULTS: Our results show that the abundance and density of medium and large burrows were significantly lower in encroached areas than in non-encroached areas (p = 0.011 and p < 0.001, respectively). The relationship between burrow abundance and bankrupt bush encroachment was negative (rho = -0.456, p = 0.001). However, burrowing mammal diversity had no significant difference between encroached and non-encroached areas
CONCLUSION: Our data, therefore, suggest that with increasing bankrupt bush encroachment and a decreased abundance in burrowing mammal ecosystem services, a negative effect will occur on burrowing mammal communities, leading to the reduction in species-specific habitat heterogeneity and possibly animal biodiversity
Keywords: burrowing mammals, burrow density, Seriphium plumosum, shrub density, Telperion Nature Reserve, Grassland Biome.
Introduction
The Grassland Biome is the second largest biome in South Africa, encompassing 28% of the land area, and supports a number of ecosystems inhabited by diverse vertebrate and invertebrate communities (Mucina & Rutherford 2006; Carbutt et al. 2011). For example, the small, medium and large burrowing mammals that live in grasslands are ecosystem engineers that play a vital role in sustaining the open habitats characterising grassland areas (Davidson et al. 2012; Jayadevan et al. 2018). Grasslands are of agricultural, ecological and conservation management importance; however, due to an increase in CO2, inadequate fire regimes and poor management, grasslands are threatened by an increase in shrub or bush encroachment (Ward 2005; Buitenwerf et al. 2011; Carbutt et al. 2011; Soto-Shoender et al. 2018; Graham et al. 2020). Shrub or bush encroachment is a phenomenon observed through an increase in woody biomass and cover, which in turn leads to a detriment in herbaceous layers (O'Connor et al. 2014). Shrub encroachment can thus lead to changes in ecological succession and ultimately influence biodiversity (O'Connor et al. 2014). Bankrupt bush, Seriphium plumosum, is a native, encroaching woody plant originating from the fynbos region in the Western Cape of South Africa and belonging to the Asteraceae family (Jordaan 2009; Snyman 2012; Van Zyl & Avenant 2018; Graham et al. 2020). The ability of S. plumosum, to encroach grasslands has interested researchers since the 1930s, as it reduces grazing potential of rangelands (Roux 1969; Avenant 2015). Nevertheless, little work has been done to examine the effect of shrub encroachment generally, or S. plumosum encroachment specifically, on mammals and mammalian ecosystem engineers (Avenant 2015; Rodgers et al. 2017; Jayadevan et al. 2018). This is despite S. plumosum being regarded as an aggressive encroacher of the Grassland Biome of South Africa (Mucina & Rutherford 2006). The encroaching aetiology of S. plumosum is well documented (Jordaan 2009; Buitenwerf et al. 2011; Snyman 2012; Van Zyl & Avenant 2018), with previous research indicating that S. plumosum follows a pioneer plant growth strategy by mainly encroaching grasslands after soil disturbance through overgrazing and trampling (Roux 1969; Jordaan 2009; Snyman 2012; Avenant 2015). With a root system ensconcing 1 m2 of the soil surrounding the bush, and reaching depths of 1.8 m (Jordaan 2009; Snyman 2012; Van Zyl & Avenant 2018), this shrub allows for tight soil binding, which may limit the burrowing services of ecosystem engineers in densely encroached areas (Vahrmeijer 2017; Uys 2018).
Ecosystem services provided directly or indirectly by ecosystem engineers include soil development, increased soil fertility, reduction in soil erosion, an increase in nutrient cycling and even food provision (Laundré & Reynolds 1993; Gabet et al. 2003; James & Eldridge 2007; Martin 2017; Rodgers et al. 2017). Landscape development by bioturbation (the movement or reworking of soil by burrowing organisms) creates and transforms habitats by physically altering the species and community specific distribution of resources within ecosystems (Gabet et al. 2003; Martin 2017). However, soil characteristics and the availability of suitable food sources influence the abundance, ecology and behaviour of ecosystem engineers such as semi-fossorial mammals. Medium (15-30 cm) and large (34-100 cm) sized burrows created by, for example, yellow mongoose, Cynictis penicillata, suricates, Suricata suricatta, aardvark, Orycteropus afer, and Cape porcupine, Hystrix africaeaustralis, create microhabitats within their burrows (Ewacha et al. 2016; Rodgers et al. 2017) providing refuge to many different species such as invertebrates, rodents, birds and reptiles (Davidson et al. 2012; Rodgers et al. 2017). However, it is speculated that the abundance of burrows and associated microhabitats decrease as shrub growth intensifies and soils become less productive in open grassland habitats (James & Eldridge 2007; Rodgers et al. 2017). With a decrease in burrow abundance, it is expected that burrowing mammal diversity would decrease as shrub encroachment leads to a reduction in available resources such as productive soils and food sources (Iribarren & Kotler 2012; Kgosikoma et al. 2012; Jayadevan et al. 2018).
The thickening of S. plumosum in grassland habitat and its relationship with elusive burrowing mammals are of interest to us. Here we investigate if there is a correlation between burrow abundance and S. plumosum density and if there is a difference between burrowing mammal diversity in encroached and non-encroached areas. We predict that a negative correlation will exist between burrow abundance and S. plumosum density and that encroached areas will have lower burrowing mammal diversity than non-encroached areas.
Study area
The Telperion Nature Reserve (25°38'S, 29°01'E), measuring some 7 350 ha, is located northeast of the town Bronkhorstspruit on the border of the Gauteng and Mpumalanga provinces of South Africa (Figure 1). The vegetation on the Telperion Nature Reserve is described as Mesic Highveld Grassland, comprising grass plains, wooded areas and vegetated mountainous or rocky areas (Figure 1; Mucina & Rutherford 2006). The area is characterised by having a mean annual precipitation of 726 mm and a mean annual temperature of 14.7°C (Mucina & Rutherford 2006). Telperion Nature Reserve contains a variety of large mammal species, including giraffe, Giraffa camelopardalis, eland, Tragelaphus oryx, plains zebra, Equus quagga, waterbuck, Kobus ellipsiprymnus and blue wildebeest, Connochaetes taurinus, and carnivores such as leopard, Panthera pardus, and brown hyaena, Parahyaena brunnea. Smaller mammal species on the reserve include Cape porcupine, yellow mongoose, South African springhare, Pedetes capensis, and black-backed jackal, Canis mesomelas (Fagir et al. 2015).
Methods
Shrub density and burrow abundance measurements
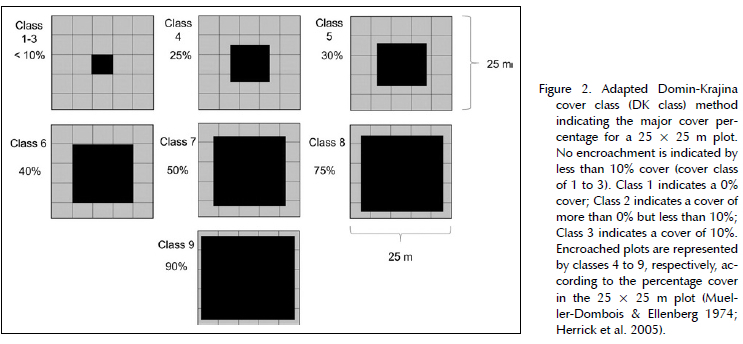
Sampling took place from April 2018 to July 2018. We measured S. plumosum abundance in the northern and southern grasslands of the Telperion Nature Reserve. Here we classified the two grassland areas into two categories based on the estimated cover of S. plumosum. In doing so, we used an adapted Domin-Krajina (DK) cover abundance scale to select ground cover classes of the area occupied by S. plumosum in each area (Mueller-Dombois & Ellenberg 1974; Herrick et al. 2005). We demarcated 24 sampling plots, 12 encroached and 12 non-encroached plots, respectively, each measuring 25 x 25 m (625 m2). All S. plumosum individuals in the plot were visualised into the centre of the plot and compared to the cover class (DK Class) (Figure 2). If the ground cover of S. plumosum was 10% or less, the area had little to no encroachment. The area was classified as densely encroached when the ground cover was estimated to exceed 10% (Westfall & Panagos 1984; Herrick 2005). We randomly allocated the centre position of each sampling plot within the encroached and non-encroached areas. All sampling plots were placed at least 50 m from the nearest road. To estimate the abundance of S. plumosum, we divided each quadrat into 25 blocks, 5 x 5 m in size, and counted the total number of adult shrubs in each sample plot. These data were expressed as the S. plumosum density (shrubs/m2) per sampling plot. We did not count any of the shrubs with less than 50% of their base within the quadrat (Elzinga et al. 1998; Van Zyl & Avenant 2018). Following the work by Avenant (2015), we considered adult shrubs to have a stem height taller than 45 cm.
We counted the medium (15-30 cm) and large (34100 cm) mammalian burrows in each sampling plot and calculated burrow abundance and density. We measured the width and length of each burrow entrance with a tape measure (model number 30-657, Stanley Black and Decker, USA) to identify medium and large mammalian burrows (Rodgers et al. 2017).
Mammalian diversity
We placed one camera trap (Browning Strike Force HD PRO, model BTC - 5HDP, Browning trail cameras, Alabama, USA) at the edge of each of the six plots in the encroached and non-encroached areas. Cameras were deployed from April 2018 to July 2018. Each camera trap was fixed onto a metal fence pole 50 cm above the surface of the ground and left to capture animal activity within the selected plot. Cameras were set to burst mode, with three pictures taken over 10 seconds when triggered. Pictures within 30 minutes of the same animals captured were not used in data analyses. The locations of all camera traps were recorded with a handheld GPS (Garmin eTrex® 10, Garmin Consumer Electronics, USA). We classified burrowing mammals into two categories based on their length and mass. We considered mammals with a reported total length < 90 cm and mass < 14 kg as medium sized, while those with a total length > 100 cm and mass > 15 kg were considered large burrowing mammals. We used Skinner and Chimimba (2005) as our reference work on burrowing mammal size.
Statistical analyses
Data were statistically analysed in R (version 3.4.0) and RStudio (version 1.0.143, RStudio: Integrated development environment for R, Boston, USA). We used paired t-tests to determine if the mean shrub density and burrow abundance differed between the encroached and non-encroached areas. We used Spearman's rank correlation to investigate the relationship between burrow abundance and S. plumosum encroachment, and the Shannon Wiener Index (H where H=Σ(pi)xln(pi)]) using the VEGAN package (version 2.4-3, RStudio (version 1.0.143) (Oksanen et al. 2013) to assess species diversity in the two areas. We also conducted an Analysis of Similarity (ANOSIM) (Jaccard) of the two areas using the VEGAN package (version 2.4-3, RStudio (version 1.0.143) (Oksanen et al. 2013).
Results
Shrub and burrow density
In the encroached and non-encroached areas, the S. plumosum shrub density ranged from 0.16 to 1.74 shrubs/m2 and 0-0.04 shrubs/m2, respectively (Figure 3). The recorded densities are equivalent to up to 17 400 S. plumosum shrubs in the encroached areas, while the non-encroached areas had up to 400 S. plumosum shrubs. The mean shrub density in the encroached areas (0.5 ± 0.4 shrubs/m2) was significantly higher than in the non-encroached areas (0.004 ± 0.01 shrubs/m2; t = 6.59, p < 0.001).
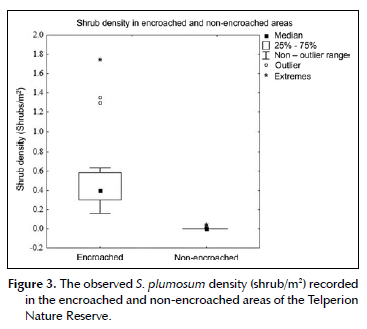
We found three burrows (two medium and one large-sized) spread across two (8.3%) of the 24 encroached sampling sites. We found 57 burrows distributed across 15 (62.5%) of the non-encroached sampling areas. Of these, 22 burrows were medium-sized and 35 were large-sized. Overall, the mean burrow density in the encroached areas (0.0002 ± 0.0007/m2) was significantly lower than in the non-encroached areas (0.004 ± 0.005/m2; t = -3.48, p = 0.002).
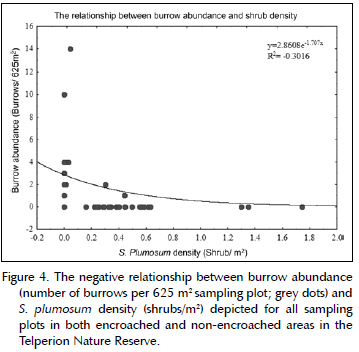
The mean abundance of medium sized burrows was significantly lower in the encroached areas (0.083 ± 0.408 per 625 m2) than in non-encroached areas (0.92 ± 2.02 per 625 m2; t = -2.03, p = 0.05). Similarly, the mean abundance of large burrows was lower in the encroached (0.041 ± 0.204 per 625 m2) than in the non-encroached areas (1.46 ± 1.95 per 625 m2; t = -3.47, p = 0.002). There was a significant moderate negative correlation between total burrow abundance and shrub density (Figure 4) (Spearman rank correlation test: rho = -0.456, p = 0.001).
Diversity of burrowing mammals
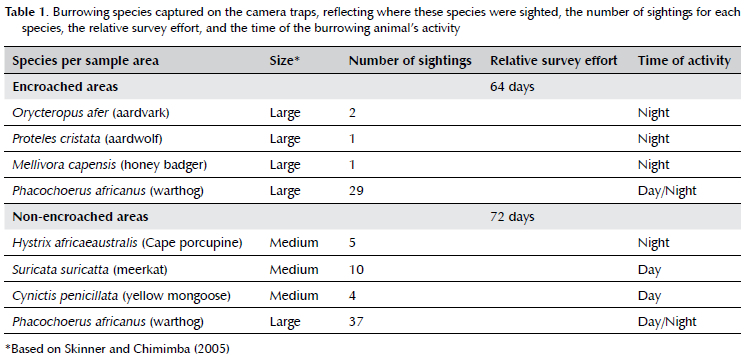
The total number of camera trap days equalled 64 in the encroached and 72 in the non-encroached areas. We obtained images of burrowing mammals from seven of 12 camera traps, i.e., from three of six cameras deployed in the encroached areas and from four of six cameras deployed in the non-encroached areas. The camera traps captured seven burrowing species, including three species of medium sized burrowing mammal and four species of large burrowing mammals (Table 1). The common warthog, Phacochoerus africanus, was the most sighted and the only burrowing species recorded in both the encroached and non-encroached areas. Burrowing mammal species richness was even for both areas, as four species were captured in non-encroached areas and four in encroached areas. The mean Shannon Wiener diversity index (H) for encroached and non-encroached areas was 0.45 ± 0.3 and 0.12 ± 0.22, respectively. Our ANOSIM indicated that the diversity of burrowing mammals had no significant difference (R = -0.231, p = 0.933) between the encroached and the non-encroached areas.
Discussion
Burrow and shrub relationship
Although absent in some of our sample plots, we found a mean S. plumosum density ranging from 40 plants per hectare in the non-encroached areas to 5 000 plants per hectare in the encroached areas. Our results on S. plumosum density align with those of Graham et al. (2020), who reported densities of up to 9 500 S. plumosum individuals per hectare on the Telperion Nature Reserve. Both medium and large mammalian burrows occurred at lower densities in the encroached than non-encroached areas, and we found a negative relationship between burrow abundance and shrub density. Despite our limited sampling across the Telperion Nature Reserve, these results support our first prediction. Moreover, our results correspond with earlier studies that reported a higher prevalence of medium and large sized burrows in open pastures and grasslands compared to woody, bushy areas (Butynski & Mattingly 1979; Melton & Daniels 1986; Augustine et al. 1995; Whittington-Jones 2006; Whittington-Jones et al. 2011; Rodgers et al. 2017). Many burrowing mammals prefer open grasslands to burrow in rather than dense shrublands, presumably because open grasslands increase predator detection and tend to have higher food and spatial availability (Melton & Daniels 1986; Davidson et al. 2012; Jayadevan et al. 2018).
Our camera trapping results revealed no significant similarity in burrowing mammal diversity between the encroached and non-encroached areas. Therefore, we cannot accept our second prediction that an increase in S. plumosum density would result in decreased burrowing mammal diversity. Nevertheless, Rodgers et al. (2017) recently reported higher burrowing mammal diversity on a non-encroached Namibian game reserve compared to an encroached livestock farm. While shrub and bush encroachment does not necessarily lead to the loss of habitat heterogeneity, and the encroachment effects are likely species, scale and environment specific (Eldridge & Soliveres 2014), some burrowing species have been affected negatively by bush encroachment. For example, shrub thickening in semi-arid parts of North America has contributed to population declines in prairie dogs Cynomys spp. (Weltzin et al. 1997); shrub thickening in semi-arid regions of Australia has had similar effects on burrowing bettongs, Bettongia lesueur (Noble et al. 2007). In our study, predators and burrowing mammals were only captured together on one camera in the non-encroached area. In the encroached areas, no camera trap yielded a picture of a predator and a burrowing mammal in the same plot. Even though we did not calculate for predators in the vicinity of the burrowing mammal diversity, it was noted that black-backed jackals were sighted more in the non-encroached areas than in the encroached areas. Predators play an important role in the distribution of prey species such as ecosystem engineers, and we must investigate how predators can influence the distribution of ecosystem engineers in the Telperion Nature Reserve (Melton & Daniels 1986; Davidson et al. 2012; Jayadevan et al. 2018).
On the Telperion Nature Reserve, where S. plumosum is spread across an estimated 30% of the reserve (Brown, unpublished data), it was interesting to note that the less common burrowing species, such as aardvark and aardwolf (Table 1), which prefer feeding in open areas with termite mounds (Melton & Daniels 1986; Williams & Richardson 1997; Whittington-Jones 2006; Stuart 2015; Rodgers et al. 2017), were only recorded in our encroached sampling areas during the current study. Although we have not consistently quantified termite mound availability as part of this study, termite mounds were prevalent in the encroached areas that we sampled as part of this study. The presence of aardvark and aardwolf in only the encroached areas may, therefore, be an artefact of our sample site selection. Moreover, the extent to which the S. plumosum encroachment, which has not been quantified across the Telperion Nature Reserve, has affected these less-common species is not well-understood at present. However, elsewhere in South Africa, simulations have predicted likely negative effects of continued encroachment on local mammal diversity and abundance (Soto-Shoender et al. 2018). These predicted effects are in line with earlier work reporting that shrub thickening negatively influenced the abundance of medium sized burrowing carnivores (Blaum et al. 2007). It is not inconceivable, therefore, that increasing shrub cover and a decreased abundance in burrowing mammal populations could lead to a decrease in the ecosystem services that burrowing mammals provide (Carbutt et al. 2011; Davidson et al. 2012).
These services include changes in soil structure and nutrients through the creation of burrow mounds, burrow networks, and the presence of latrine sites close to burrow entrances (Melton & Daniels 1986; Davidson et al. 2012; Martin 2017; Rodgers et al. 2017), which could allow for improved water infiltration, seed germination, increased nutrients and landscape heterogeneity to occur (James 2009; Whittington-Jones et al. 2011; Davidson et al. 2012; Haussmann 2017; Louw et al. 2017). Burrowing mammals are vital in creating microhabitats (Blanco-Perez et al. in prep.) and thermal refuges for a range of other vertebrates (Weyer et al. 2020). Therefore, the disappearance of keystone species, including the burrowing ecosystem engineers, may result in the structural change of grassland habitats and can lead to cascading effects on burrowing mammal ecosystem services, other taxa and possibly biodiversity (Davidson & Lightfoot 2008; Martinez-Estevez et al. 2013; Meysman et al. 2006). In grassland areas it may, therefore, be beneficial to monitor burrowing mammal density and burrow occupancy to highlight potential changes in the ecosystem services provided. Furthermore, this will help elucidate how grassland structural changes occur and may facilitate the development of new management approaches to reduce the risk of grassland habitat change (Davidson & Lightfoot 2008; Martinez-Estevez et al. 2013; Meysman et al. 2006). Furthermore, quantifying the encroachment of S. plumosum, as well as the effects thereof on the burrowing mammal community, is therefore essential in the effective management of S. plumosum in the Telperion Nature Reserve.
A limiting factor of our study is that we did not consider burrowing networks, which may have resulted in us overestimating the number of burrows. Nevertheless, we have shown that S. plumosum encroachment, if not controlled in grassland areas, can likely lead to a loss in ecosystem services associated with burrowing mammals, and reduce the state of the landscape in encroached areas. However, further investigation regarding burrowing mammal populations, occupancy and behaviour on the Telperion Nature Reserve must be done to determine the influence of woody encroachment on ecosystem engineers.
Acknowledgements
We thank Duncan MacFadyen from Oppenheimer Generations for allowing us access to the Telperion Nature Reserve. Elsabé Bosch, the reserve manager, and her staff, as well as ABEERU, are thanked for their support. Jancke Loggenberg, Tertius Junior Bester and Samkelo Mcineka helped with fieldwork. Stephan Henning and Cheré Mostert made comments on earlier versions of the manuscript. MJS was funded by the DSI-NRF Centre of Excellence for Invasion Biology.
Competing interest
We have no competing interests concerning the writing of this article.
References
Augustine, D., Manzon, A., Klopp, C. & Elter, J., 1995, 'Habitat selection and group foraging of the springhare, Pedetes capensis, in East Africa', African Journal of Ecology, 33, 347-357. [ Links ]
Avenant, P., 2015, 'Report on the national bankrupt bush (Seriphium plumosum) survey (2010-2012)', Department of Agriculture, Forestry & Fisheries, Directorate Land Use & Soil Management, Sub-directorate: Natural Resources Inventories & Assessments, South Africa. [ Links ]
Blaum, N., Rossmanith, E., Popp, A. & Jeltsch, F., 2007, 'Shrub encroachment affects mammalian carnivore abundance and species richness in semiarid rangelands', Acta Oecologica, 31, 86-92, https://doi.org/10.1016/j.actao.2006.10.004. [ Links ]
Buitenwerf, R., Bond, W., Stevens, N. & Trollope, W., 2011, 'Increased tree densities in South African savannas: >50 years of data suggests CO2 as a driver', Global Change Biology 18, 675-684, https://doi.org/10.1111/j.1365-2486.2011.02561.x. [ Links ]
Butynski, T.M. & Mattingly, R., 1979, 'Burrow structure and fossorial ecology of the springhare Pedetes capensis in Botswana', African Journal of Ecology, 17 205-215. [ Links ]
Carbutt, C., Tau, M., Stephens, A. & Escott, B., 2011, 'The conservation status of temperate grasslands in southern Africa', Grassroots 11, 17-23. [ Links ]
Davidson, A.D. & Lightfoot, D.C., 2008, 'Burrowing rodents increase landscape heterogeneity in a desert grassland', Journal of Arid Environments, 72, 1133-1145, https://doi.org/10.1016/j.jaridenv.2007.12.015. [ Links ]
Davidson, A.D., Detling, J.K. & Brown, J.H., 2012, 'Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world's grasslands', Frontiers in Ecology and the Environment, 10, 477-486, https://doi.org/10.1890/110054. [ Links ]
Eldridge, D.J.A. & Soliveres, S.B., 2014, 'Are shrubs really a sign of declining ecosystem function? Disentangling the myths and truths of woody encroachment in Australia', Australian Journal of Botany, 62, 594-608, http://dx.doi.org/10.1071/BT14137. [ Links ]
Elzinga, C.L., Salzer, D.W. & Willoughby, J.W., 1998, Monitoring Plant Populations, Bureau of Land Management, Denver, Colorado, USA. [ Links ]
Ewacha, M., Kaapehi, C., Waterman, J. & Roth, J., 2016, 'Cape ground squirrels as ecosystem engineers: modifying habitat for plants, small mammals and beetles in Namib Desert grasslands', African Journal of Ecology, 54, 68-75, https://doi.org/10.1111/aje.12266. [ Links ]
Fagir, D.M., Horak, I.G., Ueckermann, E.A., Bennett, N.C. & Lutermann, H., 2015, 'Ectoparasite diversity in the eastern rock sengis (Elephantulus myurus): the effect of seasonality and host sex', African Zoology, 50, 109-117, https://doi.org/10.1080/15627020.2015.1021173. [ Links ]
Gabet, E.J., Reichman, O.J. & Seabloom, E.W., 2003, 'The effects of bioturbation on soil processes and sediment transport', Annual Review of Earth and Planetary Sciences, 31, 249-273. [ Links ]
Graham, S.C., Barrett, A.S. & Brown, L.R., 2020, 'Impact of Seriphium plumosum densification on Mesic Highveld Grassland biodiversity in South Africa', Royal Society Open Science 7, 192025, https://doi.org/10.1098/rsos.192025. [ Links ]
Haussmann, N.S., 2017, 'Soil movement by burrowing mammals: A review comparing excavation size and rate to body mass of excavators', Progress in Physical Geography, 41, 29-45, https://doi.org/10.1177/0309133316662569. [ Links ]
Herrick, J.E., Van Zee, J.W., Havstad, K.M., Burkett, L.M. & Whitford, W.G. (eds), 2005, Monitoring manual for grassland, shrubland and savanna ecosystems. Volume I: Quick Start. Volume II: Design, supplementary methods and interpretation, USDA-ARS Jornada Experimental Range, Las Cruces, USA. [ Links ]
Iribarren, C. & Kotler, B., 2012, 'Foraging patterns of habitat use reveal landscape of fear of Nubian ibex Capra nubiana', Wildlife Biology, 18, 194-201, https://doi.org/10.2981/11-041. [ Links ]
James, A.I., 2009, 'Development of resource rich patches by soil disturbing animals in arid environments', Ph.D. thesis, School of Biological, Earth and Environmental Sciences, University of New South Wales. [ Links ]
James, A. & Eldridge, D.J., 2007, 'Reintroduction of fossorial native mammals and potential impacts on ecosystem processes in an Australian desert landscape', Biological Conservation, 138, 351-359, https://doi.org/10.1016/j.biocon.2007.04.029. [ Links ]
Jayadevan, A., Mukherjee, S. & Tamim, A.T., 2018, 'Bush encroachment in fluences nocturnal rodent community and behaviour in a semi-arid grassland in Gujarat, India', Journal of Arid Environments, 153, 32-38, https://doi.org/10.1016/j.jaridenv.2017.12.009. [ Links ]
Jordaan, D., 2009, 'Bankruptbush (slangbos) - A silent threat to grasslands?', Grassroots, 9, 40-42. [ Links ]
Kgosikoma O.E., Harvie, B.A. & Mojeremane, W., 2012, 'Bush encroachment in relation to rangeland management systems and environmental conditions in Kalahari ecosystem of Botswana', African Journal of Agricultural Research, 7, 2312-2319, https://doi.org/10.5897/AJAR11.2374. [ Links ]
Laundré, J. & Reynolds T., 1993, 'Effects of soil structure on burrow characteristics of five small mammal species', The Great Basin Naturalist, 53, 358-366. [ Links ]
Louw, M.A., Roux, P.C., Meyer-Milne, E. & Haussmann, N.S., 2017, 'Mammal burrowing in discrete landscape patches further increases soil and vegetation heterogeneity in an arid environment', Journal of Arid Environments, 141, 68-75, http://dx.doi.org/10.1016/j.jaridenv.2017.02.007. [ Links ]
Martin, S.A., 2017, 'The aardvark as an ecological engineer in the Eastern Karoo: dig patterns and emergent processes', M.Sc. thesis, Faculty of Science, Nelson Mandela Metropolitan University. [ Links ]
Martinez-Estevez, L., Balvanera, P., Pacheco, J. & Ceballos, G., 2013, 'Prairie dog decline reduces the supply of ecosystem services and leads to desertification of semiarid grasslands', PLoS ONE, 8, p.e75229, https://doi.org/10.1371/journal.pone.0075229. [ Links ]
Melton, D. & Daniels, C., 1986, 'A note on the ecology of the aardvark Orycteropus afer', South African Journal of Wildlife Research, 16, 4-6. [ Links ]
Meysman, F.J.R., Middelburg, J.J. & Heip, C.H.R., 2006, 'Bioturbation: a fresh look at Darwin' s last idea', Trends in Ecology and Evolution, 21, 688-695, https://doi.org/10.1016/j.tree.2006.08.002. [ Links ]
Mucina, L. & Rutherford, M.C., 2006, The vegetation of South Africa, Lesotho and Swaziland, Strelitzia 19, South African National Biodiversity Institute, Pretoria. [ Links ]
Mueller-Dombois, D. & Ellenberg, H., 1974, Aims and methods of vegetation ecology, John Wiley and Sons, New York, USA. [ Links ]
Noble, J.C., Hik, D.S. & Sinclair, A.R.E., 2007, 'Landscape ecology of the burrowing bettong: fire and marsupial biocontrol of shrubs in semi-arid Australia', Rangeland Journal, 29, 107-119, https://doi.org/10.1071/RJ06041. [ Links ]
O'Connor, T., Puttick, J. & Hoffman, M., 2014, 'Bush encroachment in southern Africa: changes and causes', African Journal of Range & Forage Science, 31, 67-88, https://doi.org/10.2989/10220119.2014.939996. [ Links ]
Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P, Minchin, P.R., O'Hara, R.B., Simpson, G.L., Solymos, P., Stevens, M.H.H., Wagner, H. & Oksanen, M.J., 2013, Package 'vegan'. Community ecology package, version, 2. [ Links ]
Rodgers, M., Bilton, M.C. & Hauptfleisch, M.L., 2017, 'Responses and feedbacks of burrowing mammals under differently managed rangelands', Namibian Journal of Environment, 1, 40-51. [ Links ]
Roux, E. 1969, Grass, a story of Frankenwald, Oxford University Press, Cape Town, pp. 153-159. [ Links ]
Skinner, J.D. & Chimimba C.T., 2005, Mammals of the southern African subregion, 3rd edition, Cambridge University Press, Cambridge, United Kingdom. [ Links ]
Snyman, H.A., 2012, 'Habitat preferences of the encroacher shrub, Seriphium plumosum', South African Journal of Botany, 81, 34-39, https://doi.org/10.1016/j.sajb.2012.05.001. [ Links ]
Soto-Shoender, J.R., McCleery, R.A., Monadjem, A. & Gwinn, D.C., 2018, 'The importance of grass cover for mammalian diversity and habitat associations in a bush encroached savanna', Biological Conservation, 221, 127-136, https://doi.org/10.1016/j.biocon.2018.02.028. [ Links ]
Stuart, C., 2015, Stuarts' field guide to mammals of southern africa: including Angola, Zambia & Malawi, Penguin Random House, South Africa. [ Links ]
Uys, G., 2018, 'Running a small hunting operation', Farmer's Weekly,13, 34-36. [ Links ]
Vahrmeijer, B.H., 2017, 'Our unequal struggle with "problem" plants', FarmBiz, December, 28-31. [ Links ]
Van Zyl, E. & Avenant, F!, 2018, 'Bankrupt bush: A serious threat to South Africa's central grassland', FarmBiz, February, pp. 40-42. [ Links ]
Ward, D., 2005. 'Do we understand the causes of bush encroachment in African savannas?', African Journal of Range & Forage Science, 22, 101-105, https://doi.org/10.2989/10220110509485867. [ Links ]
Weltzin, J.F., Archer, S., & Heitschmidt, R.K., 1997, 'Small-mammal regulation of vegetation structure in a temperate savanna', Ecology, 78, 751-763. [ Links ]
Westfall, R. & Panagos, M., 1984, 'A cover meter for canopy and basal cover estimations', Bothalia, 15, 241-244. [ Links ]
Weyer, N.M., Fuller, A., Haw, A.J., Meyer, L.C.R., Mitchell, D., Picker, M., Rey, B. & Hetem, R.S., 2020, 'Increased diurnal activity is indicative of energy deficit in a nocturnal mammal, the aardvark', Frontiers in Physiology, 11, 637, https://doi.org/10.3389/fphys.2020.00637. [ Links ]
Whittington-Jones, G.M., 2006, 'The role of aardvarks (Orycteropus afer) as ecosystem engineers in arid and semi-arid landscapes of South Africa', M.Sc. thesis, Rhodes University. [ Links ]
Whittington-Jones, G.M., Bernard, R.T.F. & Parker, D.M., 2011, 'Aardvark burrows: a potential resource for animals in arid and semi-arid environments', African Zoology, 46, 362-370, https://doi.org/10.1080/15627020.2011.11407509. [ Links ]
Williams, J.B. & Richardson, PR.K., 1997, 'Seasonal differences in field metabolism, water requirements, and foraging behavior of free-living aardwolves', Ecology, 78, 2588-2602. [ Links ]
 Correspondence:
Correspondence:
W. Maartin Strauss
E-mail: strauwm@unisa.ac.za
Submitted: 10 December 2020.
Accepted: 11 January 2023
Published: 17 July 2023


{kind=link}
{kind=link}
{kind=link}