










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950Print version ISSN 0378-4738
Water SA vol.51 n.3 Pretoria Jul. 2025
https://doi.org/10.17159/wsa/2025.v51.i3.4114
RESEARCH PAPER
Spatio-temporal changes in the riparian vegetation associated with the Grassland Biome section of the Olifants River, Mpumalanga, South Africa
PN MashabaI, II; WJ MyburghI; MD PanagosI
IDepartment of Nature Conservation, Tshwane University of Technology, Pretoria, South Africa
IILapalala Wilderness Foundation, P.O. Box 348, Vaalwater 0530, South Africa
ABSTRACT
The spatio-temporal changes over a 25-year period in the plant communities associated with the macro-channel of the Olifants River in the Mesic Highveld Grassland in the Mpumalanga Province, South Africa, were investigated. An area-based variable belt transect was used to resurvey selected sites which were previously surveyed roughly 25 years ago. Species richness increased from upstream to downstream driven by high influxes from specifically herbaceous species. Trends in vegetation cover varied from an increase in canopy cover in the upstream plant communities to a decrease in mid-to-downstream plant communities. The woody component increased in upstream communities, altering the riparian vegetation structure within the Grassland Biome section and resulting in spatial changes. The observed human-induced disturbances, such as livestock grazing, fire and river regulation, in conjunction with natural disturbances such as floods, contributed to the observed spatio-temporal changes. This study establishes a foundation for examining various disturbances, ranging from local to landscape levels, to develop research-based management guidelines tailored to different land uses.
Keywords: macro-channel, riparian vegetation, Olifants River, disturbances, plant communities
INTRODUCTION
Macro-channels of river systems are often associated with relatively narrow strips of vegetation on either side of the edge of the active channel. This vegetation is referred to as riparian vegetation and is defined by Kemper (2001) as vegetation which is found adjacent to rivers and perennial streams in a defined riparian zone which is functionally dependent on the watercourse. It exhibits structural, compositional and functional traits which are distinct from the adjacent terrestrial vegetation and it is distributed according to the lateral inundation gradient (Gregory et al., 1991; Kemper, 2001). The distinction between terrestrial and riparian vegetation is not always clearly defined. Myburgh and Bredenkamp (2004) found that the riparian vegetation in the Grassland Biome section of the Olifants River is not distinguishable from the surrounding vegetation as in many cases it presents terrestrial plant species. The type of vegetation found within the riparian zone is determined by multiple factors, such as climate, species composition and most importantly, the hydrogeomorphology of the area in question (Tabacchi et al., 1998; Richardson et al., 2007). This vegetation is also influenced by natural disturbances such as floods and variable river flows as well as human-induced disturbances such as land use, river regulation, pollution and climate change (Bendix, 1998; Naiman et al., 2010; Stromberg et al., 2010). In terrestrial ecosystems where climate is complementary to woody plants, such as in the Savanna Biome, riparian vegetation is structurally dominated by woody plants (Van Coller, 1997; Myburgh and Bredenkamp, 2004) and is classified and described based on dominant floristic parameters with reference to the structural organization of the vegetation (see Edwards, 1983). The herbaceous layer dominates in areas where climate, hydrogeomorphology and disturbances are non-complementary for woody species (Richardson et al., 2007). This herbaceous dominance is typically true for the stretch of the Olifants River system transecting the higher altitude Highveld Grasslands of Mpumalanga in South Africa (Myburgh and Bredenkamp, 2004).
Physical processes are the ultimate external control basins through rivers that control the structure and function of riparian vegetation (Charlton, 2008; Naiman et al., 2010). Although physical processes, through hydrological and geomorphological processes, are the primary drivers of change in riparian vegetation (Bendix, 1998; Stromberg et al., 2010), anthropogenic influence on riparian areas is well documented and dates back from the Gallo-Roman periods to the present time, where more pervasive anthropogenic-induced changes are prevalent (Naiman and Decamps, 1997; Naiman et al., 2010).
Multiple human-induced disturbances resulting from different land uses have been studied by various researchers in the Olifants River system (Balance et al., 2001; Myburgh, 2001; De Villiers and Mkwelo, 2009; Dabrowski and De Klerk, 2013). Myburgh (2001) found that different land-use activities such as mining, agriculture and land development, together with natural disturbances such as floods, have impacted the species composition and structure of the plant communities of the Olifants River.
A total of 8 plant communities were identified and described in the Grassland Biome section of the Olifants River (Myburgh and Bredenkamp, 2004). Recognizing that macro-channels and their associated riparian vegetation are exposed to multiple disturbance regimes and to obtain an insight into the possible changes in plant communities over temporal and spatial scales, this study aimed to (i) investigate changes in plant community structure, canopy cover and species composition over 25 years, and (ii) identify the disturbance regimes mostly associated with changes in plant communities.
STUDY AREA
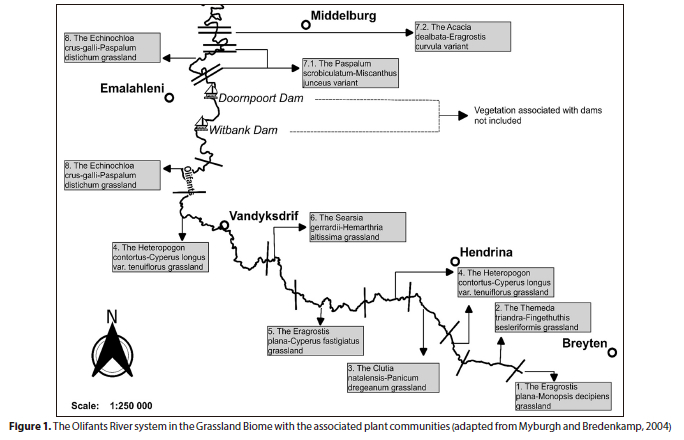
This study specifically focuses on the stretch of river between the towns of Breyten and Emalahleni in the Mpumalanga Province, South Africa (Fig. 1). The geology within this stretch of the river consists of granite rock types and intrusions of dolerite dykes and sills associated with the Post-African Planation Surfaces (McCartney et al., 2004). The vegetation is broadly described as belonging to the Grassland Biome within the Mesic Highveld Grassland bioregion (Mucina and Rutherford, 2006). The macro-channel width varies from < 5 m wide, with no clear riparian zone, to as wide as 32 m with a clearly defined riparian zone towards the Savanna Biome (Myburgh and Bredenkamp, 2004).
METHODS
Seven out of eight originally described plant communities by Myburgh (2001) were resurveyed during this study. The initial floristic data were originally collected in 1995 (Myburgh, 2001), while the data for this study were collected at a selected number of the exact same original sites in 2020. Plant community floristic data were collected in an opportunistic manner, i.e., those sampling sites that could be accessed or for which landowners, tenants or residents had granted permission. At each of the 13 sampling sites, both river banks were sampled, accounting for a total of 26 sampling plots. Sampling plots per plant community ranged from a minimum of 2 to a total of 6 plots (Myburgh and Bredenkamp, 2004).
An area-based vegetation sampling method was employed to survey the macro-channel of the Olifants River. A variable belt transect with a minimum size of 200 m2 was placed on a GPS-referenced sampling site on the macro-channel bank of the Olifants River (Myburgh and Bredenkamp, 2004).
A Garmin Etrex 30 GPS was used to locate the sampling sites, which were surveyed in 1995. The original sampling sites were located and the same sampling strategy and methodology were applied as in the previous study (Myburgh 2001; Myburgh and Bredenkamp, 2004). This enabled direct comparisons of floristic data per sampling plot and plant community.
The length and width of the variable belt transect, placed parallel and perpendicular to the river, respectively, varied on different sampling plots based on the width of the macro-channel bank and floristic parameters of the riparian vegetation (Myburgh and Bredenkamp, 2004). In each GPS-located sampling plot across the Grassland Biome section of the Olifants River, all rooted plants were recorded and canopy cover was assigned using the 'plant number scale' (Westfall and Panagos, 1988). Each recorded plant species was assigned a growth form following Westfall et al. (1996). Nomenclature followed Germishuizen and Meyer (2003).
A phytosociological classification approach, as described in Myburgh and Bredenkamp (2004), was used. However, in this study a variation in the form of a forced phytosociological classification (Panagos, 2019) was employed with the use of the PHYTOTAB-PC program (Westfall et al., 1997), to allow for comparison between sampling periods. A constancy value, an output of PHYTOTAB-PC program, was assigned per plant species, which refers to the proportional presence of the plant species in the plant community as a function of the number of sampling plots in which a specific plant species was recorded in relation to the total number of sampling plots in that plant community. Floristic parameter results from the PHYTOTAB-PC program were tested statistically using the IBM SPSS statistics software. Pearson's chi-squared test of association with the p-value set at 0.05 was used. The use of Pearson's chi-squared test was due to the asymmetrical distribution of the count data collected using the Plant Number Scale (Westfall and Panagos, 1988).
RESULTS
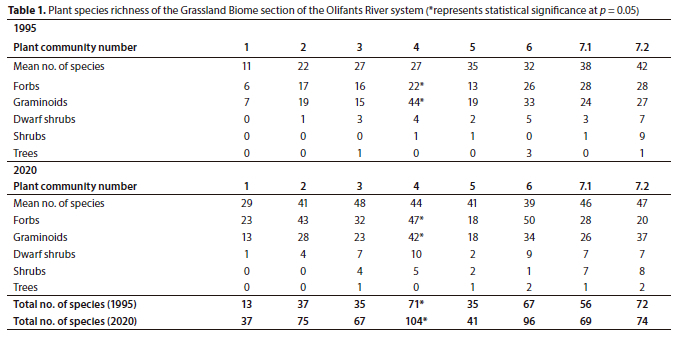
A total of 203 plant species were recorded during the 2020 sampling period, in comparison to 150 plant species recorded in 1995. The total species richness per plant community was higher in 2020 relative to 1995 (Table 1). However, only Plant Community 4 had a statistically significant (p < 0.05) increase in species richness between 1995 (71) and 2020 (104).
Plant Community 1: Eragrostis plana-Monopsis decipiens grassland
The Eragrostis plana-Monopsis decipiens grassland is located at the source of the Olifants River within the Soweto Highveld Grassland (Mucina and Rutherford, 2006) (Fig 1). The macro-channel is characterised by a single active channel and a flat herbaceous riparian bank. The total species number increased from 13 (1995) to 37 (2020) over 25 years. However, the increase was statistically insignificant (p > 0.05) (Table 1).
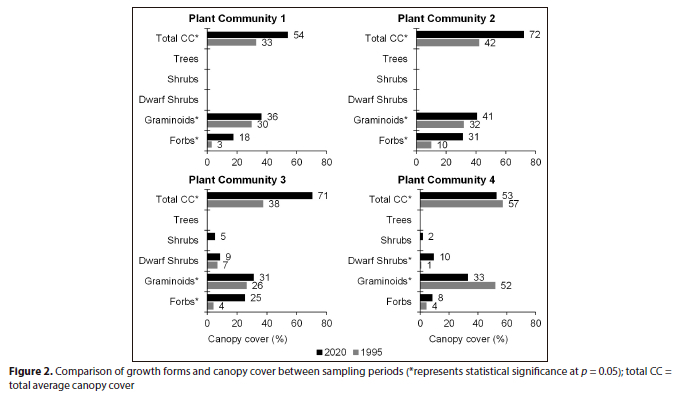
There was an increasing trend in canopy cover between 1995 and 2020. The forb layer increased from 3% to 18% cover while the graminoid layer increased from 30% to 36% between 1995 and 2020, respectively. All increases were statistically significant, resulting in a total canopy cover of 54% in 2020 in comparison to 33% in 1995 (p < 0.05) (Fig. 2).
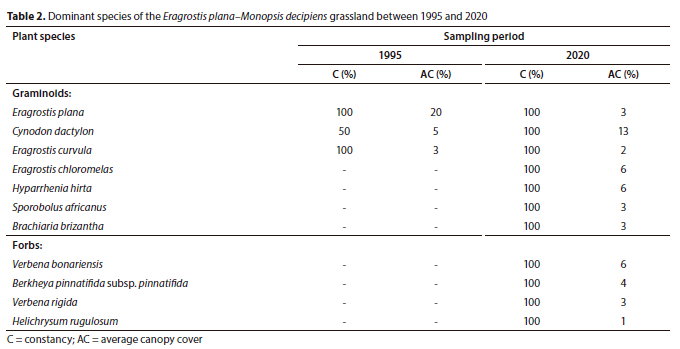
Three plant species were dominant during 1995, namely, Eragrostis plana, Eragrostis curvula and Cynodon dactylon (Table 2). Only Cynodon dactylon increased in canopy cover between 1995 (5%) and 2020 (13%). Additional dominant species recorded in 2020 include grasses Hyparrhenia hirta, Eragrostis chloromelas and the invasive alien forbs Verbena bonariensis and Verbena rigida, all exhibiting 100% constancies.
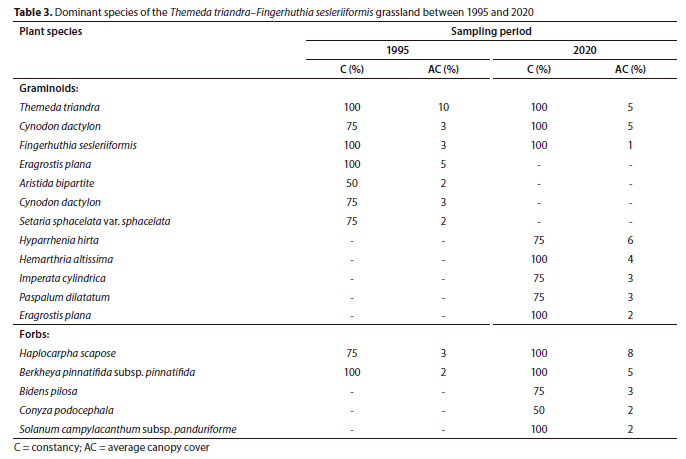
Plant Community 2: Themeda triandra–Fingerhuthia sesleriiformis grassland
Plant Community 2 occurs in the Soweto Highveld Grassland and is associated with steep macro-channel banks and random occurrences of exposed bedrock at the edges of the active channel. The total species richness increased from 37 in 1995 to 75 in 2020 (p > 0.05) (Table 1). The graminoid and forb layers characterised the plant community and displayed statistically significant (p < 0.05) increases in canopy cover ratios from 32%:10% to 41%:31% between the sampling periods, respectively. This resulted in an overall statistically significant increase in total canopy cover between 1995 (42) and 2020 (72) (p < 0.05) (Fig. 2). Only three graminoid species out of seven species maintained their dominance over 25 years, namely, Themeda triandra, Cynodon dactylon and Fingerhuthia sesleriiformis (Table 3). The newly established graminoid species such as Hyparrhenia hirta and Hemarthria altissima were amongst the most dominant species in 2020. The forb species Haplocarpha scaposa was the most dominant plant species in the plant community (Table 3).
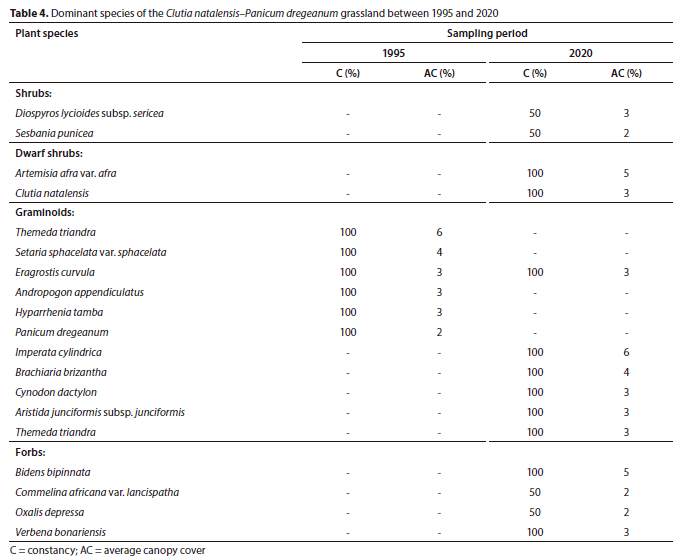
Plant Community 3: Clutia natalensis-Panicum dregeanum grassland
The Clutia natalensis-Panicum dregeanum grassland is found within the Eastern Highveld Grassland (Mucina and Rutherford, 2006). The macro-channel is characterised by the woody layer as well as alien and invasive species growing on steep riparian banks associated with alluvium deposits. The total species richness increased from 37 to 75 species, although this change was not statistically significant (p > 0.05) (Table 1).
The plant community was represented by three growth forms in 1995, namely, forbs, graminoids and dwarf shrubs. However, over 25 years, the shrub layer became established in the plant community with an average canopy cover of 5%, whereas the tree layer had a canopy cover of < 0.5%. The graminoid and forb layers showed statistically significant (p < 0.05) increases in canopy cover between the sampling periods (Fig. 2).
The shrubs Diospyros lycioides subsp. sericea and the invasive Sesbania punicea were established in the plant community between 1995 and 2020, with average canopy covers of 3% and 2%, respectively, in 2020. In the graminoid layer, only the grass Eragrostis curvula maintained its dominance over 25 years (Table 4). The recently established grass Imperata cylindrica (6% cover) and the largely absent Brachiaria brizantha (4% cover) dominated the graminoid layer in 2020. The naturalized Bidens bipinnata and the invasive Verbena bonariensis were the dominant forb species in 2020 (Table 4).
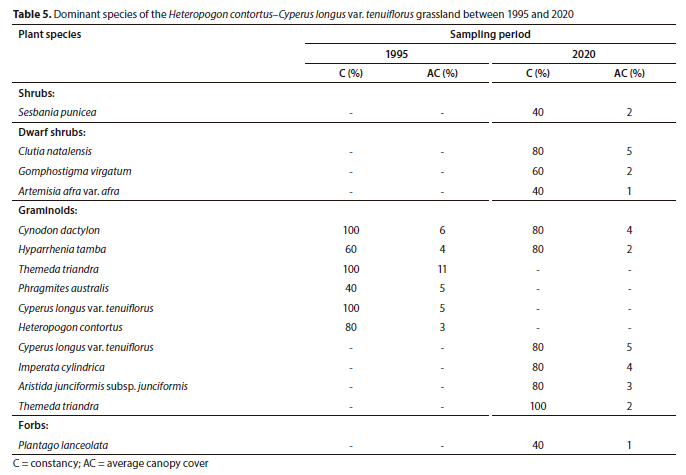
Plant Community 4: Heteropogon contortus-Cyperus longus var. tenuiflorus grassland
Similarly, the Heteropogon contortus-Cyperus longus var. tenuiflorus grassland is associated with the Eastern Highveld Grassland and occurs on steep macro-channel banks with isolated rocky areas. In some sections, the active channel is split into two channels as a result of alluvial deposits on the channel bed. This plant community recorded the highest species richness (104 species) in 2020, attributed to herbaceous species and to a lesser extent the dwarf shrubs (Table 1).
Contrasting trends in plant community structure were observed in this community. Graminoid average canopy cover declined between 1995 (52%) and 2020 (33%) while the dwarf shrub layer increased from 1% to 10% over the same period. As a result, the total average canopy cover declined between 1995 (57%) and 2020 (53%). All trends were statistically significant (p < 0.05) (Fig. 2).
No woody plant dominated the plant community in 1995, whereas the dwarf shrubs Clutia natalensis (5% cover), Gomphostigma virgatum (2% cover) and the invasive alien shrub Sesbania punicea (2% cover) dominated the community in 2020 (Table 5). The dominant grasses in 1995, namely, Themeda triandra, Cynodon dactylon and Hyparrhenia hirta, were replaced by the sedge associated with the edge of the active channel, Cyperus longus var. tenuiflorus, and the grass Imperata cylindrica as the dominant graminoid species (Table 5).
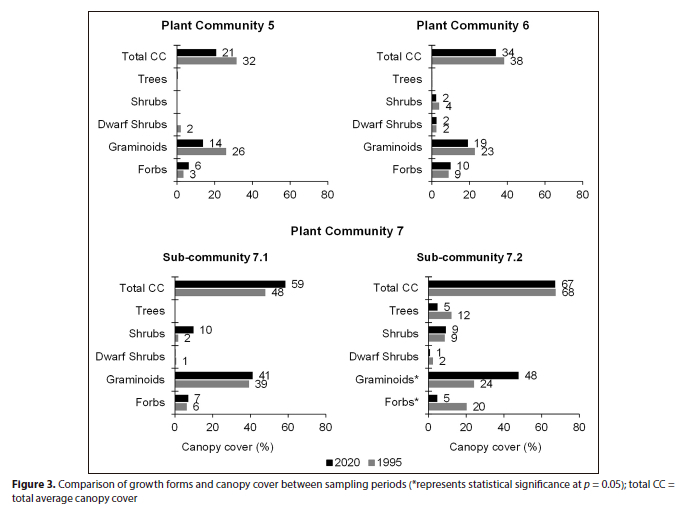
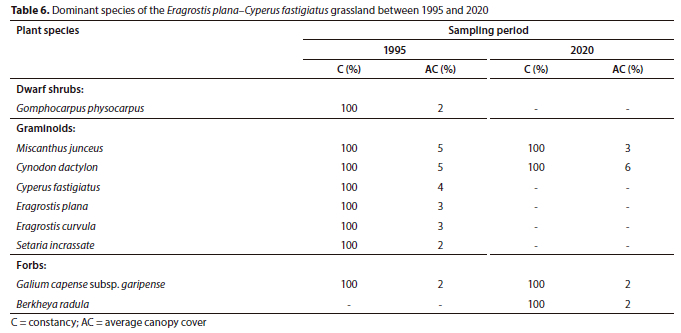
Plant Community 5: Eragrostis plana-Cyperus fastigiatus grassland
The Eragrostis plana-Cyperus fastigiatus grassland lies at an elevation of 1 600 m amsl within the Eastern Highveld Grassland (Mucina and Rutherford, 2006). The macro-channel has a single deep pool active channel and a well vegetated riparian bank with no visible signs of bank erosion.
The total species richness increased by 6 species between 1995 (35) and 2020 (41), but the increase was statiscally not significant (p > 0.05) (Table 1). The total average canopy cover declined from 32% (1995) to 21% (2020), but was also statistically not significant (p > 0.05). The decline was attributed to the graminoid layer, which declined from 26% to 14% cover over 25 years (Fig. 3). The woody component was largely absent in this community in 1995 and remained so, as seen from the 2020 results (Table 6). Only the grasses Miscanthus junceus and Cynodon dactylon remained dominant in the plant community between the sampling periods, each obtaining 100% constancy, whereas the graminoid species such as the sedge Cyperus fastigiatus and the grasses Eragrostis plana, Eragrostis curvula and Setaria incrassata were largely absent in 2020 (Table 6).
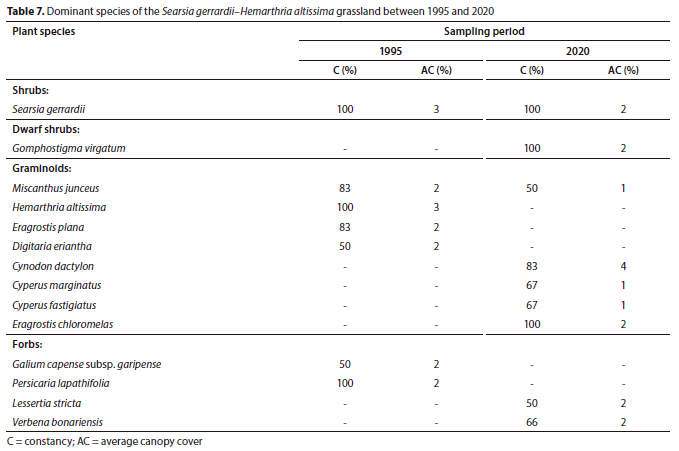
Plant Community 6: Searsia gerrardii-Hemarthria altissima grassland
The Searsia gerrardii-Hemarthria altissima grassland occurs within the Eastern Highveld Grassland at an elevation range of 1 630 m to 1 550 m amsl. The macro-channel is associated with rock cover of varying sizes, from small stones to boulders, resulting in rapids and pools. This plant community has the highest species richness of the forb layer across the Grassland Biome section of the Olifants River, with a total of 50 plant species in 2020. The overall species richness increased from 67 to 96 species between 1995 and 2020, respectively, but this change was not statistically significant (p > 0.05) (Table 1).
In terms of plant community structure, the herbaceous growth forms showed contrasting yet comparable trends over 25 years. The graminoid layer's average canopy cover declined from 23% (1995) to 19% (2020), whereas the forb layer only increased by 1% (Fig. 3). Similarly, the shrub layer declined from 4% (1995) to 2% cover in 2020. As a result of mixed trends, the total average canopy cover decreased between sampling periods, although this change was not statistically significant (p > 0.05) (Fig. 3).
There were no changes in terms of species dominance in the woody component. The shrub Searsia gerrardii maintained dominance with 100% constancy, while the previously largely absent dwarf shrub Gomphostigma virgatum was recorded with an average canopy cover of 2% in 2020 (Table 7). The herbaceous layer exhibited changes in plant species dominance. The grasses Cynodon dactylon and Eragrostis chloromelas replaced Miscanthus junceus and Hemarthria altissima as the most dominant graminoid species between 1995 and 2020. The invasive alien forb Verbena bonariensis characterised the forb layer in 2020 with 68% constancy (Table 7).
Plant Community 7: Salix mucronata subsp. woodii-Eragrostis curvula grassland
The Salix mucronata subsp. woodii-Eragrostis curvula grassland is located downstream of the Doornpoort Dam along the Olifants River. The plant community occurs through a narrow to wide macro-channel bank within the Rand Highveld Grassland underlain by diabase intrusions (Myburgh and Bredenkamp, 2004; Mucina and Rutherford, 2006).
Myburgh and Bredenkamp (2004) identified and described two sub-communities in this plant community, the Paspalum scrobiculatum-Miscanthus junceus sub-community (7.1) and the Acacia dealbata-Eragrostis curvula sub-community (7.2). The plant community's total average canopy cover between 1995 (61%) and 2020 (64%) was comparable at p = 0.05.
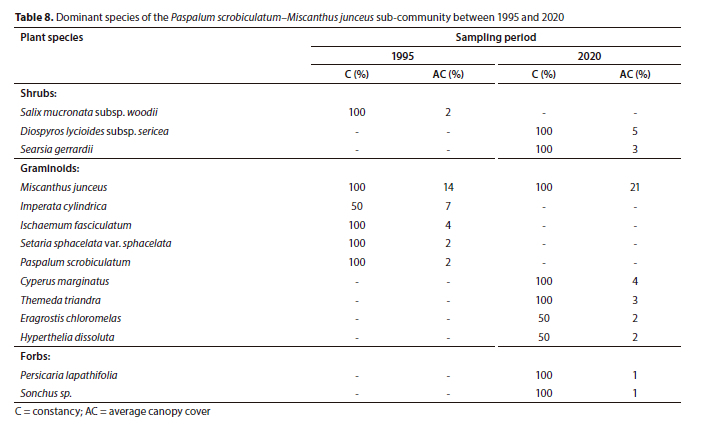
Plant Sub-Community 7.1: Paspalum scrobiculatum-Miscanthus junceus sub-community
The Paspalum scrobiculatum-Miscanthus junceus sub-community is located downstream of the Doornpoort Dam. The sub-community's macro-channel bank is characterised by dense stands of the grass Miscanthus junceus and the sedge Cyperus marginatus on the alluvium-dominated edge of the active channel.
The shrubs Diospyros lycioides subsp. sericea and Searsia gerrardii were associated with the hilly sections of the macro-channel. The total species richness increased from 56 to 69 species over 25 years, largely attributed to the herbaceous layer (Table 1). However, the increase was statistically insignificant (p > 0.05).
Forbs, graminoids and shrubs exhibited statistically insignificant (p > 0.05) increases in average canopy cover over 25 years. The notable upward trend was for the shrub layer, which increased from 2% (1995) to 10% (2020) cover. However, the overall total canopy cover increase was not statistically significant (p > 0.05) (Fig. 3).
There was a complete change in terms of species dominance in the woody component. The shrub Diospyros lycioides subsp. sericea (5% cover) and Searsia gerrardii (3% cover) replaced Salix mucronata subsp. woodii as the dominant woody plants in the sub-community in 2020 (Table 8). In contrast, within graminoids, the grass Miscanthus junceus increased from 14% (1995) to 21% (2020) cover to remain as the most dominant plant species in the sub-community (Table 8). However, there were changes in species composition, as the previously largely absent graminoid species, such as the sedge Cyperus marginatus and the grasses Themeda triandra, Hyperthelia dissoluta and Eragrostis chloromelas, were recorded in the sub-community in 2020 (Table 8).
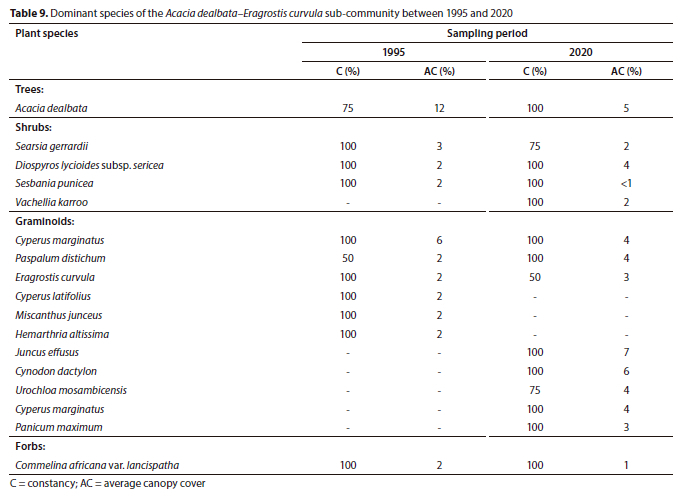
Plant Sub-Community 7.2: Acacia dealbata-Eragrostis curvula sub-community
The Acacia dealbata-Eragrostis curvula sub-community is located on the farm Mooifontein 285 JS, north of the Emalahleni-Middelburg Road (R555). The active channel changes from a single channel to multiple channels of variable widths aided by islands colonized by woody species. The sub-community occurs just before the ecotone of the Grassland and Savanna Biomes, hence the presence of the conspicuous woody component as compared to the preceding sub-community and plant communities. The total species richness between 1995 (72) and 2020 (74) was comparable (p > 0.05) (Table 1).
The herbaceous layer exhibited mixed trends in average canopy cover. The graminoid layer showed a statistically significant increase from 24% to 48% cover between 1995 and 2020, respectively, whereas the forb layer declined from 20% to 5% over the same period (p < 0.05) (Fig. 3). In the woody component, the tree layer declined from 12% (1995) to 5% (2020) cover; however, the decrease was not statistically significant at p = 0.05. The difference in the total average canopy cover of this sub-community was comparable between 1995 (68%) and 2020 (67%) (Fig. 3).
The invasive alien tree Acacia dealbata dominated the sub-community, notwithstanding the decline in average canopy cover between 1995 (12%) and 2020 (5%). Within the shrub layer, the invasive alien shrub Sesbania punicea displayed a decline in average canopy cover over 25 years while the previously largely absent Vachellia karroo was among the dominant species in 2020 (Table 9). In the graminoid layer, the sedge Cyperus marginatus, the naturalized alien grass Paspalum distichum and the grass Eragrostis curvula retained their dominance over 25 years. However, these species were surpassed by the rush Juncus effusus (7% cover) and the grass Cynodon dactylon (6% cover) as the most dominant graminoids in 2020 (Table 9).
DISCUSSION
The Olifants River system is associated with different land uses (Myburgh, 2001; Balance et al., 2001) which coexist with natural disturbances such as floods (Myburgh, 2001; Parsons et al., 2006) and human-induced disturbances (Patten, 1998; Shafroth et al., 2016). Myburgh and Bredenkamp (2004) identified and described a total of 8 plant communities in the Grassland Biome section of the macro-channel of the Olifants River. During Myburgh's 2001 study, a 1:100-year flood occurred in the Olifants River system which impacted the macro-channel riparian vegetation and the geomorphic template of the river system (Myburgh, 2001).
The flood impacts have been shown to have an influence on riparian vegetation dynamics (Bendix, 1998; Parsons et al., 2006). The plant communities of the Grassland Biome section of the Olifants River were dominated by the herbaceous layer during 1995. The woody component increased along the downstream longitudinal gradient from Plant Communities 3 to 7 (Myburgh and Bredenkamp, 2004).
The total species richness increased across all plant communities between 1995 and 2020. In Plant Communities 1, 2, 4 and 6, the forb layer replaced graminoids as the growth form with the highest species richness over 25 years. Increases in the shrub and dwarf shrub layers were also observed in Plant Communities 2, 3, 4, 6 and Sub-Community 7.1. Dwarf shrubs have been reported to show upstream species richness increases in river systems (Jansson et al., 2000), as was the case in Plant Community 2.
The increases in species richness were predominantly driven by the high number of forb plant species phasing into the macro-channel. This could be a result of integrated disturbance regimes such as livestock grazing and fire (Patten, 1998), evidence of which was observed during this study, as well as other factors such as variable river flows providing ideal conditions for the establishment of new plant species and thus increasing the species richness over 25 years. Variable river flows provide ideal conditions and substrate for the establishment of early plant colonizers (Gregory et al., 1991).
Plant Community 1 had an increase in total canopy cover over 25 years. The increase was driven by grasses Cynodon dactylon and Eragrostis chloromelas. Commercial livestock farming is the primary land use in the plant community which can be linked to the increase of the above Increaser 2 grass species as a result of heavy grazing. Livestock grazing has been shown to cause species compositional changes in riparian plant communities of the ephemeral rivers of South Africa (Allsopp et al., 2007), in Tanzania (Mligo et al., 2016) as well as in the USA (Kauffman et al., 1983). However, from a riparian-zone ecological perspective, the lawn-forming character of Cynodon dactylon can be viewed as beneficial in terms of riparian bank stabilization (Rowntree, 1991; Chen et al., 2015).
In Plant Community 2, the forb Haplocarpha scaposa and the grass Hyparrhenia hirta contributed largely to the overall increase in the total canopy cover. The macro-channel of this community has cut into the landscape making it prone to flash floods which caused bare patches open to colonization by the forb Haplocarpha scaposa. The changes in species dominance within the graminoid layer can be attributed to the decline in the canopy cover of the grass Themeda triandra.
There was a higher grass biomass visually observed in higher-lying riparian banks. Themeda triandra is a Decreaser species; lack of defoliation, fire and grazing, leads to a decrease in the abundance this grass. This provided ideal conditions for the unpalatable Increaser 1 grass Hyparrhenia hirta to establish and increase canopy cover and dominate the graminoid layer's species composition. Various researchers have found a lack of defoliation to reduce the abundance, cover and productivity of Themeda triandra in various habitats across South Africa (Fynn et al., 2005; Raitt et al., 2005; Snyman et al., 2013).
The Clutia natalensis-Panicum dregeanum grassland's (Plant Community 3) canopy cover nearly doubled in 25 years. The macro-channel of this community is characterised by alluvium deposits; this constitutes a disturbance providing a suitable substrate for pioneer species such as forbs, which had the greatest increase in canopy cover over 25 years, in contrast to other growth forms. Highly disturbed riparian substrates provide suitable conditions for early colonizer plant species (Gregory et al., 1991). The deposited alluvium could further be linked to the establishment of the grasses Imperata cylindrica and Brachiaria brizantha through seed dispersal, which dominated the graminoid layer in 2020.
Similarly to Plant Community 2, the Decreaser grasses Themeda triandra and Setaria sphacelata var. sphacelata declined in canopy cover post-1995. This could be attributed to the land use of livestock farming, where cattle were observed grazing within the macro-channel. Several studies have reported the negative impacts of livestock grazing within riparian systems (Allsopp et al., 2007; Hoffman and Rohde, 2010; Jones et al., 2022).
The species composition of Plant Community 3 was largely altered by the establishment and increase of the shrub Diospyros lycioides subsp. sericea and the invasive alien shrub Sesbania punicea, both of which were absent in 1995. This shows evidence of spatial changes in the upstream section of the Olifants River in the Grassland Biome section. Russell and Ward (2014) reported a consistent increase in the woody component in the Sub-Escarpment Grasslands of KwaZulu-Natal. This signifies the overall increasing trend of the woody component across different habitats in the Grassland Biome.
The total average canopy cover of Plant Community 4 significantly declined over 25 years. The declines were largely attributed to the grasses Themeda triandra, Heteropogon contortus and the reed Phragmites australis, which were largely phased-out post-1995. This plant community is associated with both commercial and communal livestock farming which might explain the decrease of relatively palatable grasses such as Themeda triandra and Heteropogon contortus. The stability of macro-channel banks were variable between these land uses with evidence of eroded bare riparian banks. This might be linked to the decline of Phragmites australis as a result of intermediate disturbances such as flash floods, enhanced by the trampling effect of livestock. Floods have been shown to reset riparian plant communities (Bendix, 1998; Parsons et al., 2006) while trampling has been shown to impact the macro-channels and their associated riparian vegetation (Green and Kauffman, 1995; Ruto et al., 2023).
The woody component modified the species composition and structure of Plant Community 4. The dwarf shrub Clutia natalensis, Gomphostigma virgatum and the invasive alien shrub Sesbania punicea dominated the community post-1995. The increase of dwarf shrub species can be attributed to patches opened up by heavy grazing and flood disturbances. Disturbances from heavy grazing have been related to the increase in the woody component in Grassland Biome habitats (O'Connor et al., 2014), whereas the post-flood disturbance period provides substrate for the recovery of the woody layer (Gregory et al., 1991; Parsons et al., 2006).
The decline in the total canopy cover continued from Plant Community 4 through Plant Community 5. The decrease was attributed to the dwarf shrub Gomphocarpus physocarpus, grasses Eragrostis plana, Eragrostis curvula, Setaria incrassata and the sedge Cyperus fastigiatus, all of which were predominantly phased-out in 2020. Disturbances associated with livestock farming, such as fire, grazing and trampling within riparian vegetation, together with increased intensity of variable river flows, have been shown to cause a decline in the cover of the herbaceous layer of riparian vegetation (Bendix, 1999; Pettit and Naiman, 2007; Douglas et al., 2015). Evidence of these factors was observed in this community and these could therefore be linked to the decrease in canopy cover of these species.
The Searsia gerrardii-Hemarthria altissima grassland followed the downward trend in total canopy cover between 1995 and 2020. Changes in species composition and dominance in the graminoid layer can be linked to the overall decline in canopy cover. The palatable Decreaser grasses, Hemarthria altissima, Themeda triandra and the Increaser 1 grass Brachiaria brizantha were largely absent in the community post-1995. This led to an increase of the Increaser 2 grass Cynodon dactylon on the macro-channel banks. The indiscriminate use of fire, follow-up grazing, as well as natural disturbances such as floods, have been found to cause a decline in cover and changes in species composition (Dwire and Kauffman, 2003; Parsons et al., 2006; Jones et al., 2022). This could explain the changes observed as livestock grazing and associated activities (fire) are a common practice in this community, while floods are a common occurrence.
Plant Community 7, at the downstream portion of the Grassland Biome section of the Olifants River, had a comparable total canopy cover. However, increases were observed in Sub-Communities 7.1 and 7.2, largely attributed to the shrub layer in 7.1 and the graminoid layer in Sub-Community 7.2.
In the Paspalum scrobiculatum-Miscanthus junceus sub-community (7.1), the shrubs Diospyros lycioides subsp. sericea and Searsia gerrardii were responsible for the increase in canopy cover and replaced the previously dominant shrub Salix mucronata subsp. woodii. Salix mucronata subsp. woodii is associated with the edge of the active channel and alluvial deposits. Sub-community 7.1 is below both the Witbank and Doornpoort Dams and sediment deposition may indicate increased flooding following long periods of base flow caused by river regulation (Jansson et al., 2000; Mallik and Richardson, 2009; Shafroth, et al., 2016), which might explain the decline of Salix mucronata subsp. woodii. Hoffman and Rohde (2010) have attributed the increase in the cover of terrestrial woody species in riparian areas in the semi-arid winter rainfall region of South Africa to river regulation.
The Acacia dealbata-Eragrostis curvula sub-community (7.2) occurs just before the river transitions from the Grassland Biome to the Savanna Biome. The invasive alien tree Acacia dealbata was found to dominate the woody component of this sub-community (Myburgh and Bredenkamp, 2004). However, over 25 years there was a noticeable decline in the canopy cover of the tree layer. The decline can be linked to the intensive mechanical and chemical control of the invasive alien tree Acacia dealbata observed within the macro-channel banks of this sub-community.
The decline of the invasive alien tree Acacia dealbata favoured the establishment of the previously absent shrub Vachellia karroo. Furthermore, the shade effect imposed by Acacia dealbata on the graminoid layer, as noted by Myburgh and Bredenkamp (2004), was reduced, as the graminoid layer significantly increased in canopy cover and dominated the herbaceous layer. However, the constancy of Acacia dealbata increased to 100% over 25 years. Beater (2006) found that clearing alien plants does not always decrease alien plant species abundance in favour of indigenous riparian plant species along the Sabie River. Clearing does, however, reduce the canopy cover of large invasive alien plants (Beater, 2006), as was the case in this sub-community.
Climate is one of the major external basin controls that influences that development of fluvial systems and associated riparian areas (Charlton, 2008). However, climate change, not attributed thus far as one ofthe factors causing the observed spatio-temporal changes in the plant communities, is worth consideration in the broader context. Naiman et al. (2010) recognises climate change as one of the four broad types of human-induced disturbance occurring in riparian systems, alongside river regulation, pollution and land use, some which have been discussed.
The accelerated increase of greenhouse gas emissions is one of the factors contributing to climate change. With cognisance of spatial changes observed in this study as a result of an increase in the woody layer, the increase in the woody component in the Grassland Biome section of the Olifants River could be linked to the accelerated increase in atmospheric CO2 and warmer temperatures. O'Connor et al. (2014) reviewed the causes of bush encroachment in southern Africa and concluded that increased atmospheric CO2 and climate change to be important drivers in recent history.
CONCLUSION
The macro-channel riparian vegetation of the Grassland Biome section of the Olifants River is exposed to multiple human-induced and natural disturbances. The plant communities exhibited temporal and spatial changes along the longitudinal gradient of the macro-channel. Upstream plant communities increased in canopy cover. The herbaceous layer was responsible for the increases bringing about temporal changes. As the river flows downstream, the midstream to downstream plant communities showed a decline in canopy cover. Overall changes in canopy cover were spatially variable across the Grassland Biome section of the Olifants River.
The high influx of new plant species across the longitudinal extent of the macro-channel, especially the establishment and increase of the woody component and alien and invasive species in the upstream plant communities, largely contributed to both temporal and spatial changes in the upstream section of the Olifants River macro-channel. The invasive alien woody plants Acacia dealbata and Sesbania punicea, which were already noted as species of concern in the macro-channel (Myburgh and Bredenkamp, 2004), as well as the invasive alien forbs Verbena bonariensis and Verbena rigida, increased spatially across the Grassland Biome extent of the Olifants River system.
The changes observed in the plant communities ofthe Olifant River within the Grassland Biome section are a manifestation of different disturbance regimes, such as cumulative effects of increased and variable river flow downstream, flood frequency and intensity, and land use activities (developments, grazing, trampling effect and fire) all acting synergistically across the system.
Such a wide array of factors, including climate change, will interact with each other, thus increasing the uncertainty of vegetation response and difficulty in quantifying the main disturbance factors. This poses major implications for the management of riparian vegetation in South Africa, amplified by the lack of empirical research on the impacts that different disturbance regimes have on the country's riparian systems. This study provides a basis upon which different disturbances can be further investigated at a local to landscape scale to formulate local research-based management guidelines across different land uses.
ACKNOWLEDGEMENTS
This research was made possible by the funds sponsored by Lapalala Wilderness Foundation, Tshwane University of Technology and Prof WJ Myburgh. Thanks to go Dawn Lear of Rapula Farming and Lapalala Wilderness for facilitating the finances.
AUTHOR CONTRIBUTIONS
PNM co-conceptualized the study, conducted data collection, led data analysis and interpretation of the results, led the writing and performed revisions of the manuscript. WJM conceptualized the study, collected the 1995 data, assisted with data analysis, interpretation of the results and performed critical revisions of the manuscript. MDP assisted with data and statistical analysis, interpretation of the results and performed critical revisions of the manuscript.
ORCIDS
PN Mashaba: https://orcid.org/0000-0002-4342-9486
WJ Myburgh: https://orcid.org/0000-0002-1020-1864
REFERENCES
ALLSOPP N, GAIKA L, KNIGHT R, MONAKISI C and HOFFMAN M (2007) The impact of heavy grazing on an ephemeral river system in the succulent karoo, South Africa. J. Arid Environ. 71 82-96. https://doi.org/10.1016/j.jaridenv.2007.03.001 [ Links ]
BALANCE A, HILL L, ROUX D, SILBERBAUER M and STRYDOM W (2001) State of the Rivers Report: Crocodile, Sabie, Sand and Olifants River Systems. WRC Report No. TT 147/01. Water Research Commission, Pretoria. [ Links ]
BEATER MMT (2006) Impacts of invasive alien plant clearing on riparian vegetation recovery along riverine corridors in Mpumalanga, South Africa. MSc dissertation, University of the Witwatersrand. [ Links ]
BENDIX J (1998) Impact of a flood on southern California riparian vegetation. Phys. Geogr. 19 162-174. https://doi.org/10.1080/02723646.1998.10642646 [ Links ]
BENDIX J (1999) Stream power influence on southern Californian riparian vegetation. J. Veg. Sci. 10 243-252. https://doi.org/10.2307/3237145 [ Links ]
CHARLTON R (2008) Fundamentals of Fluvial Geomorphology. Routledge, Milton Park. [ Links ]
CHEN F, ZHANG J, ZHANG, M and WANG J (2015) Effect of Cynodon dactylon community on the conservation and reinforcement of riparian shallow soil in the Three Gorges Reservoir area. Ecol. Process. 4 3. https://doi.org/10.1186/s13717-014-0029-2 [ Links ]
DABROWSKI JM and DE KLERK L (2013) An assessment of the impact of different land use activities on water quality in the upper Olifants River catchment. Water SA 39 231-244. https://doi.org/10.4314/wsa.v39i2.6 [ Links ]
DE VILLIERS S and MKWELO S (2009) Has monitoring failed the Olifants River, Mpumalanga? Water SA 35. https://doi.org/10.4314/wsa.v35i5.49193 [ Links ]
DOUGLAS MM, SETTERFIELD SA, MCGUINNESS K and LAKE PS (2015) The impact of fire on riparian vegetation in Australia's tropical savanna. Freshwater Sci. 34 1351-1365. https://doi.org/10.1086/684074 [ Links ]
DWIRE KA and KAUFFMAN JB (2003) Fire and riparian ecosystems in landscapes of the western USA. For. Ecol. Manage. 178 61-74. https://doi.org/10.1016/s0378-1127(03)00053-7 [ Links ]
EDWARDS E (1983) A broad-scale structural classification of vegetation for practical purposes. Bothalia 14 705-712. https://doi.org/10.4102/abc.v14i3/4.1231 [ Links ]
FYNN RW, MORRIS CD and EDWARDS TJ (2005) Long-term compositional responses of a South African mesic grassland to burning and mowing. Appl. Veg. Sci. 8 5-12. https://doi.org/10.1111/j.1654-109X.2005.tb00623.x [ Links ]
GERMISHUIZEN G and MEYER N (2003) Plants of Southern Africa: An Annotated Checklist. National Botanical Institute, Pretoria. [ Links ]
GREEN DM and KAUFFMAN JB (1995) Succession and livestock grazing in a northeastern Oregon riparian ecosystem. Rangeland Ecol. Manage./J. Range Manage. Arch. 48 307-313. https://doi.org/10.2307/4002482 [ Links ]
GREGORY SV, SWANSON FJ, MCKEE WA and CUMMINS KW (1991) An ecosystem perspective of riparian zones. BioScience 41 540-551. https://doi.org/10.2307/1311607 [ Links ]
HOFFMAN MT and ROHDE RF (2011) Rivers through time: historical changes in the riparian vegetation of the semi-arid, winter rainfall region of South Africa in response to climate and land use. J. Hist. Biol. 44 59-80. https://doi.org/10.1007/s10739-010-9246-4 [ Links ]
JANSSON R, NILSSON C, DYNESIUS M and ANDERSSON E (2000) Effects of river regulation on river-margin vegetation: a comparison of eight boreal rivers. Ecol. Appl. 10 203-224. https://doi.org/10.1890/1051-0761(2000)010[0203:eorror]2.0.co;2 [ Links ]
JONES CS, DUNCAN DH, RUMPFF L, ROBINSON D and VESK PA (2022) Permanent removal of livestock grazing in riparian systems benefits native vegetation. Glob. Ecol. Conserv. 33 e01959. https://doi.org/10.1016/j.gecco.2021.e01959 [ Links ]
KAUFFMAN JB, KRUEGER WC and VAVRA M (1983) Effects of late season cattle grazing on riparian plant communities. Rangeland Ecol. Manage./J. Range Manage. Arch. 36 685-691. https://doi.org/10.2307/3898185 [ Links ]
KEMPER N (2001) Riparian vegetation index. WRC Report No. 850/3/01. Water Research Commission, Pretoria. [ Links ]
MALLIK AU and RICHARDSON JS (2009) Riparian vegetation change in upstream and downstream reaches of three temperate rivers dammed for hydroelectric generation in British Columbia, Canada. Ecol. Eng. 35 810-819. https://doi.org/10.1016/j.ecoleng.2008.12.005 [ Links ]
MCCARTNEY MP, YAWSON DK, MAGAGULA TF and SESHOKA J (2004) Hydrology and water resources development in the Olifants River Catchment. IWMI, Pretoria. [ Links ]
MLIGO C (2016) Diversity and distribution pattern of riparian plant species in the Wami River system, Tanzania. J. Plant Ecol. 10 259-270. https://doi.org/10.1093/jpe/rtw021 [ Links ]
MUCINA L and RUTHERFORD M (2006) The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. Memoirs of the Botanical Survey of South Africa. South African National Biodiversity Institute, Pretoria. [ Links ]
MYBURGH WJ (2001) Die ekologie en plantegroeibestuur van die Olifantsriviersisteem. PhD thesis, University of Pretoria. [ Links ]
MYBURGH WJ and BREDENKAMP G (2004) Macro-channel riparian vegetation of the Olifants River System in the Grassland Biome, Mpumalanga. Koedoe 47 41-54. https://doi.org/10.4102/koedoe.v47i1.71 [ Links ]
MYBURGH WJ and BREDENKAMP G (2004) Macro channel riparian vegetation of the Olifants River System in the Savanna Biome, Mpumalanga. Koedoe 47 37-60. https://doi.org/10.4102/koedoe.v47i2.80 [ Links ]
NAIMAN RJ and DECAMPS H (1997) The ecology of interfaces: riparian zones. Ann. Rev. Ecol. Systemat. 28 621-658. https://doi.org/10.1146/annurev.ecolsys.28.1.621 [ Links ]
NAIMAN RJ, DECAMPS H and MCCLAIN ME (2010) Riparia: Ecology, Conservation, and Management of Streamside Communities. Elsevier Academic Press, Burlington MA. [ Links ]
O'CONNOR TG, PUTTICK JR and HOFFMAN MT (2014) Bush encroachment in southern Africa: changes and causes. Afr. J. Range Forage Sci. 31 67-88. https://doi.org/10.2989/10220119.2014.939996 [ Links ]
PANAGOS MD (2019) The cumulative effect of long-term fire trials on the phytodiversity of the savannas of Kruger National Park. PhD thesis, Tshwane University of Technology, Pretoria. [ Links ]
PARSONS M, MCLOUGHLIN CA, ROUNTREE MW and ROGERS KH (2006) The biotic and abiotic legacy of a large infrequent flood disturbance in the Sabie River, South Africa. River Res. Appl. 22 187-201. https://doi.org/10.1002/rra.905 [ Links ]
PATTEN DT (1998) Riparian ecosytems of semi-arid North America: Diversity and human impacts. Wetlands 18 498-512. http://dx.doi.org/10.1007/bf03161668 [ Links ]
PETTIT NE and NAIMAN RJ (2007) Fire in the riparian zone: characteristics and ecological consequences. Ecosystems 10 673-687. https://doi.org/10.1007/s10021-007-9048-5 [ Links ]
RAITT GR (2005) Themeda triandra renosterveld in the Heidelberg district. MSc thesis, Stellenbosch University. [ Links ]
RICHARDSON DM, HOLMES PM, ESLER KJ, GALATOWITSCH SM, STROMBERG JC, KIRKMAN SP, PYSEK P and HOBBS RJ (2007) Riparian vegetation: degradation, alien plant invasions, and restoration prospects. Diversity Distrib. 13 126-139. https://doi.org/10.1111/j.1366-9516.2006.00314.x [ Links ]
ROWNTREE K (1991) An assessment of the potential impact of alien invasive vegetation on the geomorphology of river channels in South Africa. South. Afr. J. of Aquat. Sci. 17 28-43. https://doi.org/10.1080/10183469.1991.9631311 [ Links ]
RUSSELL J and WARD D (2014) Vegetation change in northern KwaZulu-Natal since the Anglo-Zulu War of 1879: local or global drivers? Afr. J. Range Forage Sci. 31 89-105. https://doi.org/10.2989/10220119.2013.827740 [ Links ]
RUTO DK, MUSILA FM, LIMBUA PG, KINYANJUI JM and KAIGONGI MM (2023) Effects of land use on the riparian vegetation in Mau Forest Complex in Kenya. Glob. Ecol. Conserv. 46 e02624. https://doi.org/10.1016/j.gecco.2023.e02624 [ Links ]
SHAFROTH PB, PERRY LG, ROSE CA and BRAATNE JH (2016) Effects of dams and geomorphic context on riparian forests of the Elwha River, Washington. Ecosphere 7 e01621. https://doi.org/10.1002/ecs2.1621 [ Links ]
SNYMAN HA, INGRAM LJ and KIRKMAN KP (2013) Themeda triandra: a keystone grass species. Afr. J. Range Forage Sci. 30 99-125. https://doi.org/10.2989/10220119.2013.831375 [ Links ]
STROMBERG JC, LITE SJ and DIXON MD (2010) Effects of stream flow patterns on riparian vegetation of a semiarid river: implications for a changing climate. River Res. Appl. 26 712-729. https://doi.org/10.1002/rra.1272 [ Links ]
TABACCHI E, CORRELL DL, HAUER R, PINAY G, PLANTY-TABACCHI AM and WISSMAR RC (1998) Development, maintenance and role of riparian vegetation in the river landscape. Freshwater Biol. 40 497-516. https://doi.org/10.1046/j.1365-2427.1998.00381.x [ Links ]
VAN COLLER A, ROGERS K and HERITAGE G (1997) Linking riparian vegetation types and fluvial geomorphology along the Sabie River within the Kruger National Park, South Africa. Afr. J. Ecol. 35 194-212. https://doi.org/10.1111/j.1365-2028.1997.090-89090.x [ Links ]
WESTFALL R and PANAGOS M (1988) The Plant Number Scale -An Improved Method of Cover Estimation using Variable-Sized Belt Transects. Bothalia 18 289-291. https://doi.org/10.4102/abc.v18i1.1051 [ Links ]
WESTFALL RH, VAN STADEN JM, PANAGOS MD, BREYTENBACH PJJ and GREEF A (1996) Scale-related vegetation sampling, Private Bag X05 Lynn East, 0186, Pretoria., ARC-RFI publication. [ Links ]
WESTFALL RH, THERON GK and ROOYEN N (1997) Objective classification and analysis of vegetation data. Plant Ecol. 132 137-154. https://doi.org/10.1023/a:1009746217097 [ Links ]
 Correspondence:
Correspondence:
PN Mashaba
Email: peternyeko@gmail.com
Received: 14 December 2023
Accepted: 13 June 2025

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}