










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkAfrican Natural History
versión On-line ISSN 2305-7963
versión impresa ISSN 1816-8396
Afr. nat. history (Online) vol.8 Cape Town ene. 2012
Cretaceous faunas from Zululand and Natal, South Africa. The desmoceratoid ammonite genera Moretella Collignon, 1963, Beudanticeras Hitzel, 1902, and Aioloceras Whitehouse, 1926
William James KennedyI; Herbert Christian KlingerII
IOxford University Museum of Natural History, Parks Road, Oxford OX1 3PR, U.K., and Department of Earth Sciences, South Parks Road, Oxford OX1 3AN, U.K. E-mail: jim.kennedy@oum.ox.ac.uk
IINatural History Collections Department, Iziko South African Museum, P.O. Box 61, Cape Town, 8000 South Africa. E-mail: hklinger@iziko.org.za
ABSTRACT
Three taxa of desmoceratoid ammonites are recognized in the Lower Albian of northern KwaZulu-Natal, all of them previously known only from Madagascar: Moretella sp., Beudanticeras komihevitraense Collignon, 1950, and Aioloceras besairiei (Collignon, 1949). Madagascan type and figured material, much of it difficult to interpret from the original figures is re-illustrated, and the intraspecific variation and dimorphism in Moretella and Aioloceras are documented.
Keywords: ammonites, Desmoceratoidea, Albian, Cretaceous, KwaZulu-Natal, Madagascar.
INTRODUCTION
Desmoceratoid ammonites from the Lower Albian part of the Mzinene Formation of northern KwaZulu-Natal were recorded as species of 'Beudanticeras' and 'Cleoniceras' from division Albian II in our preliminary account of the ammonite faunas of the region (Kennedy & Klinger, 1975, p. 276)). We subsequently recognized Aioloceras as a key element of a Tegoceras mosense/Aioloceras fauna (Kennedy & Klinger, 2008, p. 79). Study of the classic Lower Albian faunas of Madagascar (Collignon, 1949, 1950, 1963) has enabled us to recognize three taxa in KwaZulu-Natal, Moretella sp., Beudanticeras komihevitraense Collignon, 1950, and Aioloceras besairiei (Collignon, 1949). The quality of reproduction of the plates in the Albian volume (1963) of Collignon's Atlas des fossiles caractéristiques de Madagascar (Ammonites) did not do justice to the specimens, or the original photographs, and many taxa are difficult if not impossible to interpret without an examination of the actual specimens, housed in the collections of the Université de Bourgogne, Dijon. Accordingly, we re-illustrate here key type and figured specimens relevant to the interpretation of the KwaZulu material.
REPOSITORIES OF SPECIMENS
EMP: École National Supérieur des Mines, Paris, now housed in the Université de Lyon-Villeurbanne.
OUM: Geological Collections, Oxford University Museum of Natural History, Oxford.
SAM: Natural History Collections Department, Iziko South African Museum, Cape Town.
UB: Département des Sciences de la Terre, Université de Bourgogne, Dijon.
FIELD LOCALITIES
Details of field localities are given by Kennedy & Klinger (1975); further descriptions of these localities are deposited in the Geological Collections, Oxford University Museum of Natural History, The Natural History Museum, London, and the Natural History Collections Department, Iziko South African Museum, Cape Town. Specimens with prefixes EM or LJE were presumably collected from the Albian localities on the western side of the Pongola River, north of the Mfongozi Spruit, described by Haughton (1936, pp. 290-292, fig. 3), (see Kennedy & Klinger 2011, fig. 24)
CONVENTIONS
Dimensions are given in millimetres: D = diameter; Wb = whorl breadth; Wh = whorl height; U = umbilicus; c = costal dimension; ic = intercostal dimension. Figures in brackets are dimensions as a percentage of the diameter. The suture terminology is that of Korn et al. (2003): E = external lobe; A = adventive lobe (= lateral lobe, L, of Kullmann & Wiedmann 1970); U = umbilical lobe; I = internal lobe.
SYSTEMATIC PALAEONTOLOGY
Superfamily DESMOCERATOIDEA Zittel, 1895
Family CLEONICERATIDAE Whitehouse, 1926
(= Vnigriceratinae Saveliev, 1973; Lemuroceratinae Owen, 1988)
Genus Moretella Collignon, 1963
Type species
Pseudosonneratia madagascariensis Collignon, 1949, p. 81, pl. 16, figs 13, 14, by the original designation of Collignon, 1963, p. 101.
Diagnosis
Small, mature adults, presumed to be microconchs, are as little as 40 mm in diameter. Coiling moderately evolute, with around 50% of the previous whorl covered. Whorl section rounded, oval or subquadrate. Coarse primary ribs, bullate or not, arise at the umbilical seam and strengthen across the umbilical wall and shoulder. On the flanks ribs arise in pairs at the shoulder or on the innermost flank, and are prorsiradiate and feebly flexuous; additional short ribs intercalate. All ribs strengthen on the ventrolateral shoulders and venter, which they cross in a shallow convexity. Suture moderately incised, with bifid E/A and A/U2
Discussion
Collignon (1963) referred the following species to his new genus Moretella:
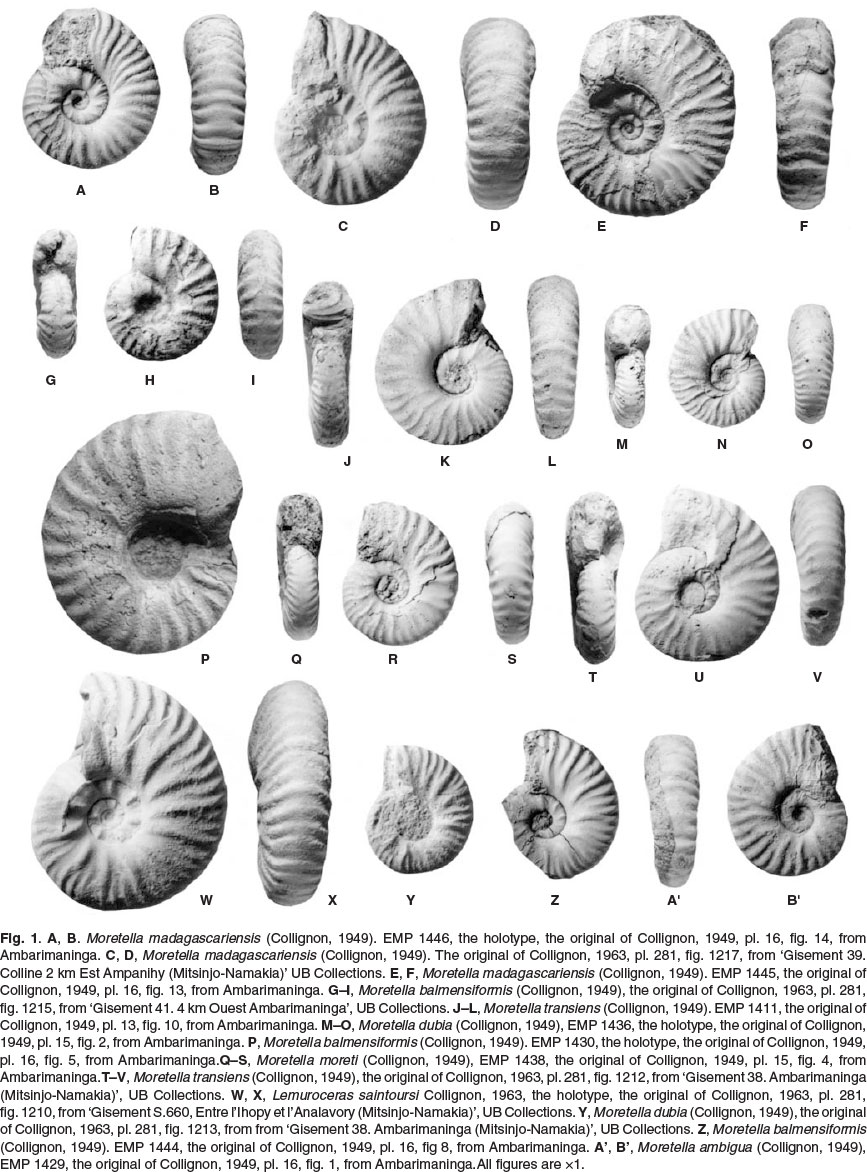
Moretella ambigua (Collignon, 1949): p. 79, pl. 16, figs 1-4; 1963, p. 102, pl. 281, fig. 1216 (Figs 1A',B', 2C,D).
Moretella balmensiformis (Collignon, 1949): Collignon, 1949, p. 80, pl. 16, figs 5-8; 1963, p. 102, pl. 281, fig. 1215 (Fig. 1G-I, P, Z).
Moretella dubia (Collignon, 1949): Collignon, 1949, p. 74, pl. 15, fig. 2; 1963, p. 102, pl. 281, fig. 1213 (Fig. 1M-O).
Moretella madagascariensis (Collignon, 1949): Collignon, 1949, p. 81, pl. 16, figs 13,14; 1963, p. 102, pl. 281, fig. 1217 (Fig. 1A-F).
Moretella moreti (Collignon, 1949): Collignon, 1949, p. 75, pl. 15, fig. 4; 1963, p. 102, pl. 281, fig. 1214 (Fig. 1Q-S).
Moretella subquadrata Collignon, 1963, p. 103, pl. 281, figs 1218, 1219 (Fig. 2H-J, M-O).
Moretella transiens (Collignon, 1949): Collignon, 1949, p. 74, pl. 13, fig. 10; 1963, p. 101, pl. 281, fig. 1212 (Fig. 1J-L).
Moretella undata Collignon, 1963, p. 103, pl. 281, fig. 1220 (Fig. 2E-G).
The co-occurrence of seven species of Moretella within the Zone of Lemuroceras spathi and Brancoceras besairiei in Madagascar is unlikely. Where Collignon saw differences, we see dimorphism and intraspecific variation: the morphological differences between his species are trivial. Already in 1949 Collignon regarded moreti (Figs 1Q-S, 2K-L) as a passage form between ambigua (Figs 1A',B', 2C,D) and madagascariensis (Fig. 1A-F) and we regard them as synonyms. It is likely that further analysis of the assemblage will lead to the conclusion that most of his species are synonyms.
We are unable to see any significant differences between several of the 'Pseudosonneratia' not included in Moretella by Collignon, in particular dondeyi Collignon, 1963, p. 105, pl. 282, fig. 1230, and ihopensis Collignon, 1963, p. 106, pl. 282, fig. 1231 (Fig. 2PQ), which we include in Moretella here.
Collignon suggested that the origin of Moretella lay in Lemuroceras Spath, 1942. The resemblance between a specimen referred to Moretella dubia by Collignon (Fig. 1Y), and the much larger (47 mm diameter), but still septate holotype of Lemuroceras saintoursi Collignon, 1963 (Fig. 1W,X) is striking; they may even be dimorphs. A full revision of the numerous co-occuring Lemuroceras, Moretella aud 'Pseudosonneratia' from the Lower Albian of Madagascar is required to resolve their actual relationships.
Occurrence
Lower Albian of Madagascar and northern KwaZulu-Natal.
Moretella sp.
Material
SAM-PCZ22239, from the Lower Albian Mzinene Formation in the environs of locality 153 of Kennedy & Klinger (1975) on the side of the road leading up to Mantuma Rest Camp in the Mkuze Game Reserve, northern KwaZulu-Natal.
Dimensions

Description
The specimen is wholly septate, and retains recrystallized shell. Coiling is moderately evolute, with 50% of the previous whorl covered. The umbilicus comprises 31% of the diameter, and is of moderate depth, with a convex wall and broadly rounded umbilical shoulder that merges imperceptibly with the feebly convex, subparallel flanks. The ventrolateral shoulders are broadly rounded, the venter relatively broad, and feebly convex. There are 25-26 ribs on the outer whorl.
Twelve primaries arise at the umbilical seam, and strengthen into strong bullae, perched on the umbilical shoulder. These give rise to pairs of strong, feebly flexuous coarse ribs, feebly convex on the inner to middle flank and feebly concave on the outer flank, sweeping forwards, coarsening on the ventrolateral shoulders and venter, which they cross in a feeble convexity. Occasional intercalated ribs arise around mid-flank, and strengthen to match the primaries on the outer flank, ventrolateral shoulders and venter.
Discussion
The present specimen shows similarities to the holotype of Moretella ihopensis Collignon, 1963 (p. 106, pl. 282, fig. 1231: Fig. 2PQ herein). This is smaller, more compressed than the present specimen, with a whorl breadth to height ratio of 0.28, and weaker ornament of similar style. It shows weakening and modification of the last few ribs, and may be an adult. The ornament is closer to that of Moretella subquadrata Collignon, 1963 (p. 103, pl. 281, figs 1218, 1219: Fig. 2H-J, M-O), which differs chiefly in its smaller size and more compressed whorl section.
Occurrence As for material.
Family DESMOCERATIDAE Zittel, 1895
Subfamily BEUDANTICERATINAE Breistroffer 1953
Genus Beudanticeras Hitzel, 1905
(= Boliteceras Whitehouse, 1928; Rapidoplacenticeras Alabushev, 1988)
Discussion
Beaudanticeras is comprehensively reviewed by Riccardi & Medina (2002). These authors referred 12 species to the genus with confidence, and listed many more referred to that genus by previous authors. They referred many of these to other genera but there remained a number of 'Beudanticeras', of uncertain generic position. Among these is Beudanticeras komihevitraense Collignon, 1950, to which a number of specimens from northern KwaZulu-Natal are referred.
Beudanticeras komihevitraense Collignon, 1950
Figs 3A-I, L-O, 4B-D, G, 8D-F, K-N, 10E
1950 Beudanticeras komihevitraense Collignon, p. 41, pl. 6, fig. 3.
1963 Beudanticeras komihevitraense Collignon; Collignon, p. 80, pl. 271, fig. 1172.
Type
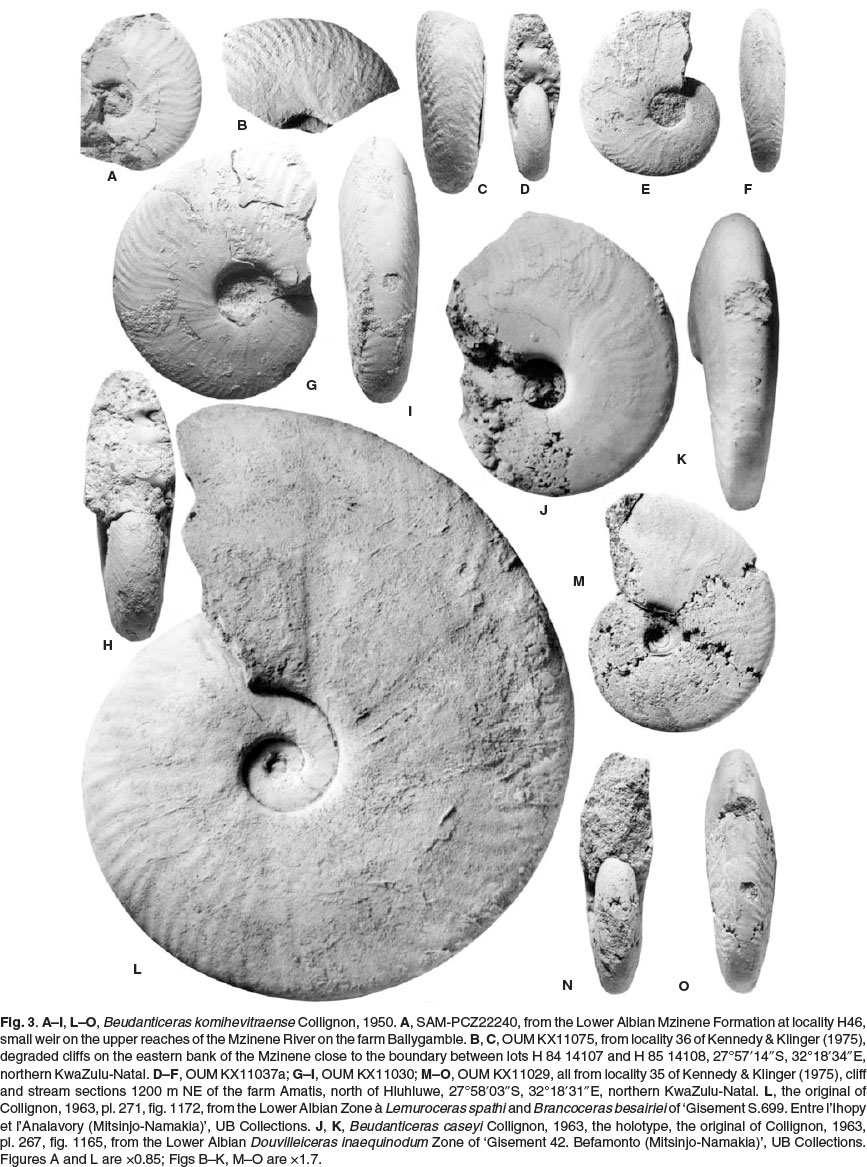
The holotype is EMP 1481, the original of Collignon, 1950, p . 41, pl. 6, fig. 3, from the Lower Albian of Komihevitra, Madagascar, re-illustrated here as Fig. 4E,F.
Material
OUM KX 11017, 11018, 11028, 11029, 11030, 11033,
11034, 11036, 11037a-g, from the Lower Albian Mzinene Formation at locality 35 of Kennedy & Klinger (1975), cliff and stream sections 1200 m NE of the farm Amatis, north of Hluhluwe, 27°58'03"S, 32°18'31"E, northern KwaZulu-
Natal. OUM KX 11070-11072, 11074-11076, are from the Lower Albian Mzinene Formation at locality 36 of Kennedy & Klinger (1975), degraded cliffs on the eastern bank of the Mzinene close to the boundary between lots H 84 14107 and H 85 14108, 27°57'14"S, 32°18'34"E, northern KwaZulu-Natal. SAM-PCZ22240, from the Lower Albian Mzinene
Formation at locality H45, from Klinger's field notes and not referred to in Kennedy & Klinger (1975), a small weir on the upper reaches of the Nyalazi River, on the farm Ballygamble, north of Mtubatuba, approximately 28°16'S, 32°15'E.
Dimensions
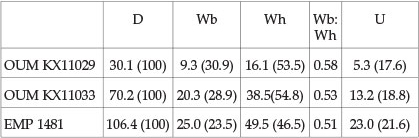
Description
OUM KX11029 (Fig. 3M-O), 11030 (Fig. 3G-I), and 11037a (Fig. 3D-F) and 11075 (Fig. 3B,C) are part of a series of nuclei, 13-35 mm in diameter. Coiling is involute, the umbilicus small, comprising17-19% of the diameter, shallow with a flattened, outward-inclined umbilical wall and narrowly rounded umbilical shoulder. The whorl section is compressed, the greatest breadth just outside the umbilical shoulder, the inner flanks very feebly convex, the middle and outer flanks flattened, converging to broadly rounded ventrolateral shoulders, the venter very feebly convex. Ornament is of closely spaced, crowded ribs. Primaries arise singly or in pairs at the umbilical shoulder, sometimes as mere lirae, and are straight and prorsiradiate on the inner flank. They bifurcate around mid-flank and on the outer flank, while additional ribs intercalate, the ribs convex at mid-flank, then flexing forwards and concave on the outer flank, strengthening progressively before weakening over the venter, which they cross in a very shallow convexity. There are thus many more ribs at the ventrolateral shoulder than at the umbilical shoulder. SAM-PCZ22240 (Fig. 3A) with an estimated original diameter of 40 mm shows the same pattern of branching and intercalating ribs, now well developed across the whole of the flanks. This specimen links to individuals such as OUM KX11033 (Fig. 4B-D, G), OUM KX11018 (Fig. 8D,E), OUM 11028 (Fig. 8F,K) and OUM 11070 (Fig. 8L-N) these specimens range up to 70 mm in diameter, and show variable inner flank ornament, from near-effaced (Fig. 8L-N) to well developed prorsiradiate falcoid primaries, straight on the inner flank, feebly convex at mid-flank, and concave on the outer flank. They bifurcate around mid-flank and on the outer flank, where they are at their maximum strength, declining and effacing over the venter. These specimens overlap in size with the phragmocone fragment preserved on the holotype (Fig. 4E,F). This is interpreted as an adult microconch. It is 106.4 mm in diameter, with a 240° sector of body chamber preserved. Coiling is very involute, the small umbilicus comprising 21.6% of the diameter, the umbilical wall of the body chamber concave, the umbilical shoulder very narrowly rounded. The whorl section is very compressed, with a whorl breadth to height ratio of 0.51, the greatest breadth below mid-flank. The inner to middle flanks are very feebly convex, the outer flanks flattened, and converging to the narrowly rounded ventrolateral shoulders, the venter very feebly convex. There are 11-12 primary ribs on the adapertural half whorl of body chamber. They arise at the umbilical seam, strengthen across the umbilical wall, and are fully differentiated on the umbilical shoulder, low, straight and prorsiradiate to mid-flank, where they increase by branching and intercalation, then sweeping back and concave on the outer flank, where they strengthening progressively. There are 32 ribs at the ventrolateral shoulder of the adapertural half whorl. Larger specimens have phragmocone whorl heights of up to 37 mm (OUM KX11072: Fig. 10E). They show the same pattern of falcoid ribs that increase in strength on the outer flanks and increase in number by branching and intercalation. The 163 mm diameter phragmocone figured by Collignon in 1963 (pl. 271, fig. 1172) is interpreted as part of a macroconch of the species. The suture of the holotype is deeply incised, with an asymmetrically trifid E/A, large trifid A, and bifid A/U2.
Discussion
Riccardi & Medina (2002, p. 301) suggested this species might not belong to Beudanticeras, as they did Beudanticeras caseyi Collignon, 1963 (p. 72, pl. pl. 267, fig. 1165), and other Madagascan species with well developed ribs. B. caseyi is uninterpretable from the original figures, and is illustrated here as Fig. 3J,K. It has much more delicate ornament than B. komihevitraense of the same size (compare Figs 3J and 3G) but the style is identical. The holotype of caseyi is barely distinguishable from a specimen referred to Beudanticeras revoli (Pervinquière, 1907) figured by Riccardi & Medina (2002, pl. 1, fig. 1), and the affinities of the Madagascan material with Beudanticeras is clear.
Occurrence
Lower Albian of Madagascar and northern KwaZulu-Natal.
Genus Aioloceras Whitehouse, 1926
(= Paracleoniceras Collignon, 1963)
Type species
Cleoniceras argentinum Bonarelli in Bonarelli & Nagera, 1921, p. 24, pl. 4, figs 3, 6, from the Lower Albian of Lago San Martin, Santa Cruz, Argentina, by the original designation of Whitehouse, 1926, p. 206.
Diagnosis
Coiling involute, umbilicus small, shallow, umbilical wall flattened and outward-inclined, umbilical shoulder narrowly rounded. Whorl section compressed, with feebly convex inner flanks, outer flanks converging to narrow rounded venter. Primary ribs feebly bullate or not, arise singly or in pairs from umbilical shoulder and strengthen across the flank; falcoid, straight on inner flank, concave on outer flank, increasing by branching and intercalation; effaced on venter. Ornament weakens progressively and effaces on body chamber of macroconchs. Constrictions develop on late phragmocone and body chamber of both dimorphs. Suture with deeply incised bifid E/A and trifid L.
Discussion
See Riccardi & Medina (2002, pp. 312, 340) for a comprehensive review of the genus and its constituent species. Their conclusions are followed here, and four species referred to the genus with confidence: Aioloceras argentinum (Bonarelli, 1921), A. besairiei (Collignon, 1949), A. ptychitiforme (Collignon, 1949), and A. rollieri (Leanza, 1970).
Occurrence
Upper Lower Albian, Argentina, Antarctica, Madagascar, and northern KwaZulu-Natal.
Aioloceras besairiei (Collignon, 1949)
Figs 5-7, 8A-C, G-J, O,P, 9, 10A-D, F-G, 11-14
1949 Cleoniceras (Aioloceras) Besairiei Collignon, p. 86, pl. 18, figs 1-3; pl. 21, fig. 7; text-fig. 24.
1963 Cleoniceras (Paracleoniceras) besairiei Collignon, p. 88, pl. 275, figs 1186, 1187; pl. 276, figs 1188, 1189.
1963 Cleoniceras (Paracleoniceras) inaequale Collignon, p. 86, pl. 274, fig. 1185.
1963 Cleoniceras (Paracleoniceras) cleoniforme Collignon, p. 89, pl. 276, figs 1190-1191.
1963 Cleoniceras (Paracleoniceras) morganiforme Collignon, p. 89, pl. 276, fig. 1192.
1963 Cleoniceras (Paracleoniceras) tenuicostatum Collignon, p. 92, pl. 277, figs 1193-1194.
1963 Cleoniceras (Paracleoniceras) crassefalcatumCollignon, p. 94, pl. 278, fig. 1195.
?1963 Cleoniceras (Paracleoniceras) ambiguum Collignon, p. 94, pl. 278, fig. 1196.
Type
The holotype, by original designation, is EMP 1455, the original of Collignon, 1949, pl. 18, fig. 2, from the A. besairiei Zone of Ambarimaninga, Madagascar, re-illustrated here as Fig. 5B-D.
Material
All our specimens are from the upper Lower Albian Mzinene Formation.
OUM KX 10990, 10992, 10993, 11026, 11031, 11032, 11035, 10990, are from locality 35 of Kennedy & Klinger (1975), cliff and stream sections 1200 m NE of the farm Amatis, north of Hluhluwe, 27°58'03"S, 32°18'31"E, northern KwaZulu-Natal. OUM KX 11068, 11069 and 11073, are from locality 36 of Kennedy & Klinger (1975), degraded cliffs on the eastern bank of the Mzinene close to the boundary between lots H 84 14107 and H 85 14108, 27°57'14"S, 32°18'34"E, northern KwaZulu-Natal. OUM KX4791, 4796, 4800, 4802, 4804, and 4806 are from locality 53 of Kennedy & Klinger (1975), derelict dam site on Indabana 13162, 2.2 km south of the farm Izwehelia, north of Hluhluwe, 27°52'24"S, 32°19'02". SAM-PCZ 022403 (ex LJE191), PCZ022402 (ex EM115) presumably from Haughton's (1936) locality, or in the vicinity of, Z18 on the Pongola River.
Dimensions
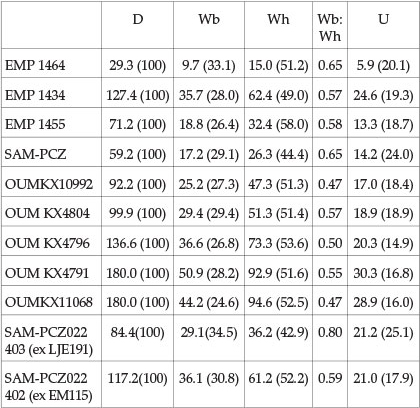
Description
Collignon based this species on a suite of 12 specimens. The holotype, EMP 1455 (Collignon, 1949, pl. 18, fig. 2; Fig. 5B-D herein) is an internal mould just over 71 mm in diameter that retains half a whorl of body chamber; it is interpreted as a microconch. Coiling is involute, the umbilicus comprising 18.7% of the diameter, shallow, the umbilical wall flattened and outward-inclined, with a narrowly rounded umbilical shoulder. The whorl section is compressed, with a whorl breadth to height ratio of 0.58, the greatest breadth low on the flanks. The inner and middle flanks are feebly convex, the outer flanks convergent, the ventrolateral shoulders broadly rounded, the venter flattened and feebly convex. A total of 35 ribs are present on the well-preserved 300° adapertural sector of the outer whorl. Primary ribs arise at the umbilical shoulder, numerous long intercalated ribs arise low on the flanks, and shorter intercalated ribs arise around mid-flank. Some ribs bifurcate on the inner flank. The ribs are weak, straight and prorsiradiate on the innermost flank, strengthening across the middle of the flanks, as do the long intercalatories. Primary and long intercalatories flex back and are convex across the mid-flank region, strengthen, flex forwards and are markedly concave on the outer flank, broaden, and cross the venter in a broad convex curve. Periodic interspaces on the adapertural part of the body chamber are deepened into incipient constrictions on the outer flank. The specimen appears to retain the adult aperture, preceded by a constriction on the inner flank. The suture (Collignon, 1949, text-fig. 24) is deeply and intricately incised, with asymmetrically bifid E/A, and deep asymmetrically bifid to subtrifid A and four auxiliary lobes. Collignon also figured (1949, pl. 18, fig. 1, re-illustrated here as Fig. 5A) a juvenile, dissected out from a larger individual. It is an internal mould 29.3 mm in diameter. Coiling is very involute with a small umbilicus that comprises 20% of the diameter. The umbilical wall is convex and outward-inclined, the umbilical shoulder narrowly rounded. The inner flanks are feebly convex, the outer flanks convergent, the ventrolateral shoulders broadly rounded, the venter feebly convex. Ornament consists of falcoid ribs, 17 on the adapertural half whorl. Primary ribs arise either singly or in pairs, and are weak, straight and prorsiradiate on the inner flank. They strengthen around mid-flank, flex back, and are concave on the outer flank, strengthening progressively on the ventrolateral shoulder, weakening somewhat over the venter, which they cross in a broad convexity. Intercalated ribs arise both low and high on the flank, and have an outer flank and ventral development like that of the primary ribs. The third figured specimen (Collignon, 1949, pl. 18, fig. 3; pl. 21, fig. 7; illustrated here as Fig. 6) is interpreted as an incomplete macroconch. It is just over 127 mm in diameter, wholly septate, and retains extensive areas of partly altered shell material. Coiling is involute, with a small shallow umbilicus that comprises 19.3% of the diameter, the umbilical wall flattened, the umbilical shoulder narrowly rounded. The whorl section is very compressed, with a whorl breadth to height ratio of 0.59, the greatest breadth low on the flanks, the inner to mid-flank region broadly convex, the outer flanks convergent, the ventrolateral shoulders broadly rounded, the venter feebly convex. Where shell material is preserved, ornament consists of crowded falcoid ribs: primaries, long and short intercalatories that may reduce to mere lirae on the inner flank, where they are straight and prorsiradiate. The ribs strengthen progressively across the flanks, are feebly convex at mid flank, and feebly concave on the outer flank, where they broaden and crowd, weakening and crossing the venter in a broad convexity. Periodic very broad, shallow constrictions first appear on the internal mould at the beginning of the outer half whorl and become increasingly prominent as size increases, although largely concealed where shell is present. Collignon subsequently illustrated additional juvenile specimens that he referred to A. besairiei (1963, p. 88, pl. 275, figs 1186, 1187; pl. 276, figs 1188, 1189), and introduced six further species of 'Paracleoniceras' (= Aioloceras herein). Riccardi & Medina (2002, p. 340) suggested these might be synonyms of A. besairiei, a view supported here; they illustrate well the variability of the species, from weakly to more strongly ornamented. The holotype of Aioloceras cleoniforme (Collignon, 1963) (p. 89, pl. 276, figs 1190-1191; Fig. 7A herein) is the form with the weakest ornament, which is reduced to mere lirae, striae and growth lines on the inner part of the flanks, but retains the basic style of ribbing of the largest of Collignon's original specimens of besairiei (1949, pl. 18, fig. 3; pl. 21, fig. 7; Fig. 6 herein). It appears to retain part of the aperture close to the umbilical margin. The second specimen referred to cleoniforme by Collignon (1963, pl. 276, fig. 1191; Fig. 7C herein) has, in contrast, slightly stronger ribs than the holotype of A. besairiei at the same diameter, although the style is the same. Aioloceras tenuicostatum Collignon, 1963) (p. 92, pl. 277, figs 1193-1194; Fig. 7B herein) has well-differentiated crowded slightly stronger ribs than the holotype of cleoniforme. The ribs are weaker and more numerous than in the holotype of A. besairiei at the same diameter, but the ornament of the adapical 90° of the outer whorl is identical in style to that of a specimen referred to besairiei by Collignon (1963, pl. 276, fig. 1188) at the same diameter, while the adapertural 120° of the outer whorl differs in no significant respects from the same sector of Collignon's larger original specimen (compare Fig. 6 and Fig. 7B). Aioloceras morganiforme (Collignon, 1963) (p. 89, pl. 276, fig. 1192; Fig. 7D herein) has slightly stronger ribs than the holotype of A. besairiei at the same diameter; the style is the same. Stronger again at the same size is the ornament of the adapical 240° sector of the outer whorl of Aioloceras inaequale (Collignon, 1963) (p. 86, pl. 274, fig. 1185; Fig. 7E herein), whereas the adapertural 120° sector shows an abrupt change to narrower, crowded ribs. The holotype of Aioloceras crassefalcatum (Collignon, 1963) (p. 94, pl. 278, fig. 1195; Fig. 7F herein) is an incomplete macroconch, complete to 130mm in diameter, with indications of a further 120° sector of outer whorl. There are 20 ribs on the ventrolateral shoulder of the adapertural half of the outer whorl compared with 30 in the comparably sized specimen referred to Aioloceras besairiei by Collignon (1963, pl. 275, fig. 1186), the greater density reflecting the presence of fewer bifurcations of ribs on the outermost flanks of crassefalcatum. We have not seen the holotype and only specimen referred to Aioloceras ambiguum (Collignon, 1963) (p. 94, pl. 278, fig. 1196). According to its author, the ornament on the adapical half of the outer whorl is as in Aioloceras besairiei, but on the adapertural half the ribs are arranged in groups of 4-6, separated by prominent constrictions that form grooves in the umbilical wall, and E/A is distinctly asymmetric. So poor is the quality of reproduction of the figures of this species that the distinctive features of the ornament are unrecognizable. In conclusion, the differences between the various Madagascan specimens are to us those of individuals, of a variable population, rather than species.
Within the KwaZulu-Natal material, microconchs are represented by individuals that are complete at diameters of between 52 and 62 mm. Strongly ornamented body chambers such as OUM KX 11031 (Fig. 8A-C) have around 22 ribs on the adapertural half whorl: primaries, long and short intercalatories. The ribs are weak, straight, and prorsiradiate on the inner flank, strengthening, flexingback and convex at mid-flank, then flexing forwards, strengthening further, and concave on the outermost flank and ventrolateral shoulder. Occasional interspaces are accentuated into constrictions on the outermost flank, ventrolateral shoulders and venter, with a strengthened collar rib or ribs. The venter broadens markedly towards the adult aperture, which is marked by a weakening of the final rib and a prominent constriction. The apertural margin is slightly flared, most obviously on the ventrolateral shoulders and venter. OUM KX 11035 (Fig. 8O,P) is a 90° fragment of body chamber with much weaker ornament, and well-developed constrictions, that immediately preceding the adult aperture is particularly prominent. SAM-PCZ 22238 (Fig. 8G-J) is a well-preserved microconch lacking only the final part of the middle and outer flank and ventral region, and complete to 59 mm diameter. The body chamber extends to 240° of the outer whorl. Ornament is feeble, but there are four well-developed constrictions on the body chamber.
OUM KX 4800 (Fig. 10F,G) and KX 4796 retain recrystallized shell material; the well-preserved ornament matches that of cleoniforme of Collignon (Fig. 7A). SAM-PCZ022402 (ex. EM 115) (Fig. 5E) is a more robustly ornamented and inflated individual.
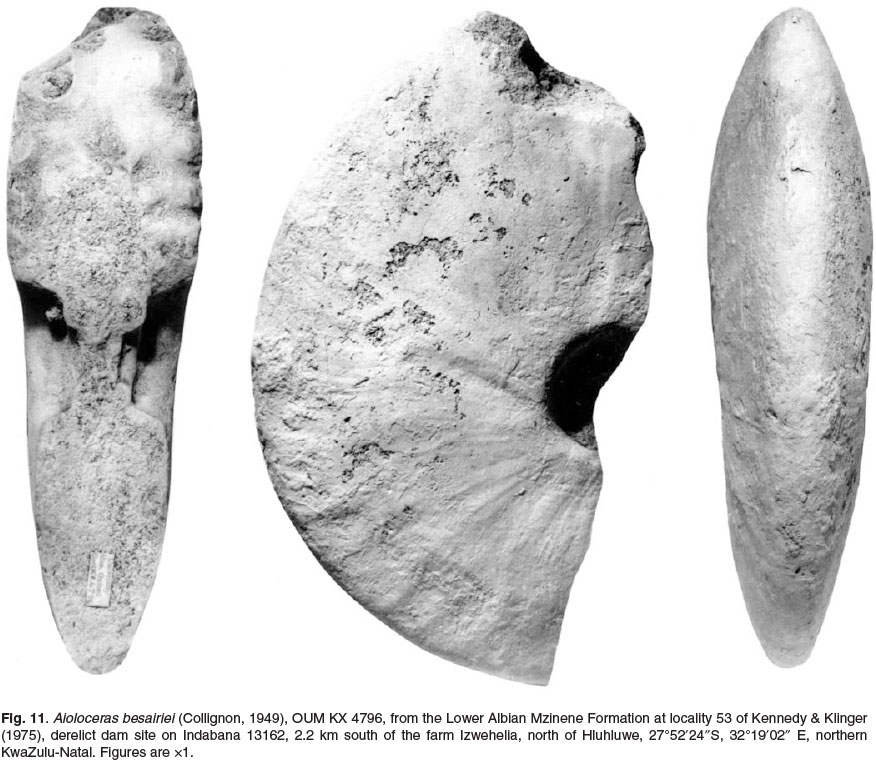
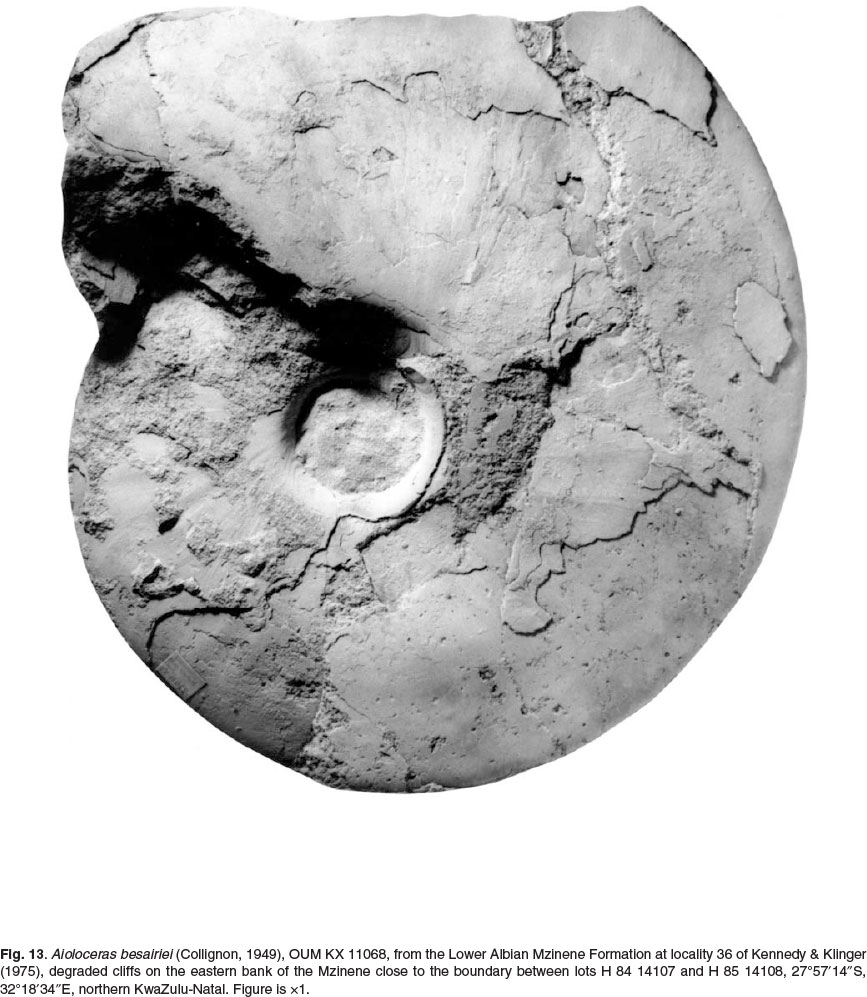
Also referred to Aioloceras besairiei are much larger individuals than those known from Madagascar. These are interpreted as macroconchs. OUM KX4796 (Fig. 11), a fragment of phragmocone 130 mm in diameter, retains low falcoid ribs like those of cleonifome, and links to OUM KX 4791 (Fig. 12), a phragmocone fragment 180 mm in diameter. The fractured section shows the penultimate whorl to have been coarsely ornamented. OUM KX11068 (Fig. 13) is complete to 180 mm diameter with a 240° sector of body chamber. OUM KX10990 (Fig. 14) is still septate to an estimated diameter of 190 mm and a maximum preserved whorl height of 95 mm. These large specimens are involute, with around 79% of the previous whorl covered, the umbilicus small (14.9-16.8% of the diameter), shallow, with a flattened, outward-inclined wall and very narrowly rounded umbilical shoulder. The whorl section is compressed, with a whorl breadth to height ratio of 0.47-0.57, the greatest breadth below mid-flank. The inner to mid-flank region is feebly convex, the outer flanks flattened and convergent, the ventrolateral shoulders broadly rounded, the venter very feebly convex (Fig. 12A). Ornament is reduced to low, broad, feeble flexuous ribs detectable on the inner and middle flank of phragmocones. On body chambers there are growth lines and striae only, straight and prorsiradiate on the inner flank, feebly convex at mid-flank, flexing forwards and feebly concave on the outer flank (Fig. 14).
Discussion
Aioloceras ptychitiforme (Collignon, 1949) (p. 88, pl. 18, fig. 4; pl. 21, fig. 6) is based on a single specimen, which comes from the same locality as the type material of A. besairiei, of which it may be no more than an aberrant individual. The original figures make interpretation difficult, and it is re-illustrated here as Fig. 4A. It differs in its whorl section, which is like that of the Triassic Ptychites, with the greatest whorl breadth just outside the umbilical shoulder, flattened, convergent flanks, and a broad feebly convex venter. The ornament is of 36-38 concave ribs on the outer whorl, confined to the outer part of the flanks, and effaced on the venter on the adapertural half of the outer whorl. Aioloceras argentinum (Bonarelli, 1921) (in Bonarelli & Nagera, 1921, p. 24, pl. 4, figs 3, 6, ; see revision in Riccardi & Medina, 2002, p. 315, pl. 4, figs 1-7; pl. 5, figs 1-6; text-figs 1-4, 8a-n, 9a-i; 10, 11) shows striking dimorphism, with macroconch phragmocones 58-74 mm in diameter and adults 80-110 mm in diameter. Microconch phragmocones are 35-55 mm in diameter, and adults 55-73 mm in diameter. As noted by Riccardi & Medina (2002, p. 319), Aioloceras besairiei has flatter and more convergent flanks, a broader and more rounded venter, coarser ornament, and reaches a larger size. Macroconchs of Aioloceras rollieri (Leanza, 1970) (p. 221, text-fig. 18.1-2) (see revision in Riccardi & Medina, 2002, p. 320, pl. 6, fig. 5; pl. 7, figs 1-4; pl. 8, figs 1-3; pl. 9, figs 1-3; pl. 10, figs 1,2; text-figs 11, 12a-c) are 192-320 mm in diameter, microconchs 70-109 mm in diameter. The variation in ornament is narrower than in Aioloceras besairiei, and that of the most robustly ornamented rollieri is weaker than that of the most robustly ornamented besairiei. The body chamber of both macro- and microconch rollieri lose the ribbing, and develop numerous strong falcoid constrictions.
Occurrence
Lower Albian. In Madagascar, the Aioloceras besairiei Zone lies between that of 'Pseudosonneratia' sakalava below, and Douvilleiceras inaequinodum above. In northern KwaZulu-Natal, it co-occurs with Tegoceras mosense (d'Orbigny, 1841) (Kennedy & Klinger, 2008, p. 79) indicating a horizon in the mid-upper, but not uppermost Lower Albian (Cleoniceras floridum Subzone of the Sonneratia chalensis Zone to Protohoplites (Hemisonneratia) puzosianus Subzone of the Otohoplites auritiformis Zone of the northwest European sequence).
ACKNOWLEDGEMENTS
Kennedy acknowledges the support of the staff of the Geological Collections, Oxford University Museum of Natural History, and the Department of Earth Sciences, Oxford, and the financial assistance ofthe Oppenheimer Fund (Oxford). Klinger acknowledges the support of the staff of the Iziko South African Museum Natural History Collections Department and financial support of the NRF, South Africa. We thank Drs J.-L. Latil (Lazer) and F. Amedro (Calais) for their reviews of the manuscript.
REFERENCES
ALABUSHEV, A.I. 1988. [Rapidolacenticeras-novyi rod melovykh ammonoidei.] Paleontologicheskii Zhurnal 1988: 110-113. [in Russian. [ Links ]]
BONARELLI, G. 1921. In: BONARELLI, G. & NAGERA, J.J. Observaciones geologicas en las immediaciones del Lago San Martin (Territorio de Santa Cruz). Boletín, Ministereo de Agricultura, Buenos Aires, Series B (Geologia) 27: 1-39. [ Links ]
BREISTROFFER, M. 1953. Commentaires taxonomiques. In: BREISTROFFER, M. & VILLOUTREYS, O. de. Les ammonites albiennes de Peille (Alpes-Maritimes). Travaux du Laboratoire de Géologie de l' Université de Grenoble 30 (for 1952): 69-74. [ Links ]
COLLIGNON, M. 1949. Recherches sur les faunes albiennes de Madagascar. I. L'Albien d'Ambarimaninga. Annales Géologiques du Service des Mines, Madagascar 16: 1-128. [ Links ]
COLLIGNON, M. 1950. Recherches sur les faunes albiennes de Madagascar. III. L'Albien de Komihevitra. Annales Géologiques du Service des Mines, Madagascar 17: 19-54. [ Links ]
COLLIGNON, M. 1963. Atlas des fossiles caractéristiques de Madagascar (Ammonites). X. Albien. xv + 184 pp. Tananarive:, Service Géologique. [ Links ]
HITZEL, E. 1902. Sur les fossiles d'âge Albien recuillis par M.A. Guebhard dans la region d'Escragnolles (Alpes-Maritimes). Bulletin de la Société Géologique de France (4)2: 874-880. [ Links ]
HAUGHTON, S.H. 1936. Account of the geology of the Cretaceous beds and a preliminary analysis of the associated ammonite fauna. In: RENNIE, J.V.L. Lower Cretaceous Lamellibranchia from northern Zululand. Annals of the South African Museum 31: 283-294. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 1975. Cretaceous faunas from Zululand and Natal, South Africa. Introduction, stratigraphy. Bulletin of the British Museum (Natural History) Geology 25: 263-315. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 2008. Cretaceous faunas from Zululand and Natal, South Africa. The ammonite subfamily Lyelliceratinae Spath, 1921. African Natural History 4: 57-111. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 2011. Cretaceous faunas from Zululand and Natal, South Africa. The ammonite genus Oxytropidoceras Stieler, 1920. African Natural History 7: 69-102. [ Links ]
KORN, D., EBBIGHAUSEN, V., BOCKWINKEL, J. & KLUG, C. 2003. The A-mode ontogeny in prolecanitid ammonites. Palaeontology 46: 1123-1132. [ Links ]
KULLMANN, J. & WIEDMANN, J. 1970. Significance of sutures in phylogeny of Ammonoidea. University of Kansas, Paleontological Contributions 42: 1-32. [ Links ]
LEANZA, A.F. 1970. Ammonites nuevos o pocos conocidos de Aptiano, Albiano y Cenomaniano de los Andes Australes con notas acera de su posicion estratigráfica. Revista de la Asociacion Geologica, Argentina 25: 197-261. [ Links ]
ORBIGNY, A. d'. 1840-1842. Paléontologie française: Terrains Crétacés. 1. Céphalopodes. 1-120 (1840); 121-430 (1841); 431662 (1842). Paris: Masson. [ Links ]
OWEN, H.G. 1988. The ammonite zonal sequence and ammonite taxonomy in the Douvilleiceras mammilatum Superzone (Lower Albian). Bulletin of the British Museum of Natural History (Geology) 44: 177-231. [ Links ]
PERVINQUIÈRE, L. 1907. Études de paléontologie tunisienne. 1. Céphalopodes des terrains secondaires. Carte Géologique de la Tunisie. Paris: de Rudeval. [ Links ]
RICCARDI, A.C. & MEDINA, A. 2002. The Beudanticeratinae and Cleoniceratinae (Ammonitida) from the Lower Albian of Patagonia. Revue de Paléobiologie 21: 291-351. [ Links ]
SAVELIEV, A.A. 1973. [Stratigrafiya i Ammonity Nizhnego Al'ba Mangyschlaka.] Trudy Vsesoiuznogo Nauchno-Issledovatel'-skogo Geologicheskogo Neftegazovogo Instituta 323: 1-339. [In Russian. [ Links ]]
SPATH, L.F. 1942. A monograph of the Ammonoidea of the Gault. 15. Palaeontographical Society Monographs, 669-720. [ Links ]
WHITEHOUSE, F.W. 1926. The Cretaceous Ammonoidea of eastern Australia. Memoirs of the Queensland Museum 8: 195-242. [ Links ]
WHITEHOUSE, F.W. 1928. Additions to the Cretaceous ammonite fauna of eastern Australia. Part 2 (Desmoceratidae). Memoirs of the Queensland Museum 9: 200-206. [ Links ]
ZITTEL, K.A. VON 1895. Grundzüge der Palaeontologie (Palaeo-zoologie). vii + 972 pp. Munich & Leipzig: R. Oldenbourg. [ Links ]
Received 19 June 2012
Accepted 10 August 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}