










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAfrican Natural History
On-line version ISSN 2305-7963
Print version ISSN 1816-8396
Afr. nat. history (Online) vol.8 Cape Town Jan. 2012
Cretaceous faunas from Zululand and Natal, South Africa. The Santonian-Campanian ammonite genus Eulophoceras Hyatt, 1903
William James KennedyI; Herbert Christian KlingerII
IOxford University Museum of Natural History, Parks Road, Oxford OX1 3PR, U.K., and Department of Earth Sciences, South Parks Road, Oxford OX1 3AN, U.K. E-mail: jim.kennedy@oum.ox.ac.uk
IINatural History Collections Department, Iziko South African Museum, P.O. Box 61, Cape Town, 8000 South Africa. E-mail: hklinger@iziko.org.za
ABSTRACT
The distinctive Santonian-Campanian ammonite genus Eulophoceras Hyatt, 1903, is revised and assigned to the subfamily Lenticeratinae. Pelecodiscus Van Hoepen, 1921, and Spheniscoceras Spath, 1921, are junior synonyms. Praelibycoceras H. Douvillé, 1912, regarded as a junior synonym by previous authors, is regarded as a separate taxon. All seven species previously recorded from the Mzamba Formation of the Eastern Cape Province are assigned to a variable, dimorphic Eulophoceras natalense Hyatt, 1903, which is also recorded from the St Lucia Formation of KwaZulu-Natal, as is a second species, Eulophoceras bererense Hourcq, 1949. The Madagascan representatives of the genus are described and illustrated for comparison, and the status of other species referred to the genus reviewed.
Key words: Ammonites, Eulophoceras, Santonian, Campanian, Cretaceous, KwaZulu-Natal, Eastern Cape Province, South Africa.
INTRODUCTION
Alpheus Hyatt (1903, p. 86, pl. 11, figs 2-6) introduced his new genus and species Eulophoceras natalense, which he referred to his new family Eulophoceratidae Hyatt, 1903 (p. 83), on the basis of a single specimen from 'Port Natal, South Africa' in the collections of the Peabody Museum of Yale University (Figs 1-3). Port Natal was renamed Durban on the latter's foundation in 1835. There are no outcrops of Cretaceous sediments in the environs of Durban (although they are present in the subsurface: Kennedy et al., 1973). The Yale collections also include a series of other ammonites labelled 'Port Natal': Plesiotexanites stangeri (Baily, 1855) (YPM6250), Hauericeras gardeni (Baily, 1855) (YPM1000, 1026), and Kossmaticeras (Karapadites) planissimus Collignon, 1966 (YPM1071, figured by Kennedy & Klinger, 1985, figs 25, 26). The preservation of the material indicates it all to be from the Mzamba Formation, and from outcrops at the mouth of the Mzamba Estuary in the Eastern Cape Province (locality 1 of Kennedy & Klinger, 1975; see also Klinger & Kennedy, 1980).
REPOSITORIES OF SPECIMENS
OUM: Geological Collections, Oxford University Museum of Natural History, Oxford. SAM: Natural History Collections Department, Iziko South African Museum, Cape Town. BMNH: The Natural History Museum, London. YPM: Peabody Museum of Yale University, Yale.
FIELD LOCALITIES
Details of field localities are given by Kennedy & Klinger (1975); further descriptions of these localities are deposited in the Geological Collections, Oxford University Museum of Natural History, The Natural History Museum, London, and the Natural History Collections Department, Iziko, South African Museum, Cape Town.
CONVENTIONS
Dimensions are given in millimetres: D = diameter; Wb = whorl breadth; Wh = whorl height; U = umbilicus; c = costal dimension; ic = intercostal dimension. Figures in brackets are dimensions as a percentage of the diameter. The suture terminology is that of Korn et al. (2003): E = external lobe; A = adventive lobe (= lateral lobe, L, of Kullmann & Wiedmann 1970); U = umbilical lobe; I = internal lobe.
SYSTEMATIC PALAEONTOLOGY
Superfamily ACANTHOCERATOIDEA de Grossouvre, 1894 Family SPHENODISCIDAE Hyatt, 1900
(= Libycoceratinae Zaborski, 1982, p. 306) Subfamily LENTICERATINAE Hyatt, 1900
(= Eulophoceratinae Hyatt, 1903, p. 83)
Discussion
Hyatt introduced his family Lenticeratidae validly in 1900 (p. 585), of which Eulophoceratidae Hyatt, 1903 is a junior synonym, notwithstanding the comment in the posthumously published Pseudoceratites of the Cretaceous (Hyatt, 1903) by the editor, T.W. Stanton (footnote b on p. 84) that 'In Zittel's Text-book of Palaeontology, Vol. 1, p. 590, Professor Hyatt referred this genus [Lenticeras Gerhardt, 1897], together with Paralenticeras and Platylenticeras, to the family Lenticeratidae. Although the statement is not directlymade that the family is abandoned, it may be inferred from the reference of Platylenticeras to Coilopoceratidae, the remark under Paralenticeras about its affinity for Eulophoceras, and the fact that the manuscript was arranged as now published with Lenticeras and Paralenticeras between Tegoceras and Eulophoceras'.
Genus Eulophoceras Hyatt, 1903
(= Spheniscoceras Spath, 1921, p. 242; Pelecodiscus Van Hoepen 1921, p. 30).
Diagnosis
Oxycone, whorl section lanceolate or with fastigiate venter separated from flanks by change in slope. Near-smooth, or with feeble bullae at the umbilical shoulder; ribs arise singlyorin pairs at the umbilical shoulder and strengthen progressively across the flanks, straight on the inner flank and flexing back on the outer, where they may efface or strengthen, be feebly concave or feebly convex, and accentuated into an elongate outer lateral bulla. Intercalated ribs, where present, arise around mid-flank. Few, feeble, spiral ridges in some. Suture highly variable, with asymmetrically trifid to subtrifid E/A, and several auxiliary elements; folioles on saddles may be highly elongated.
Discussion
Praelibycoceras H. Douvillé, 1912 (p. 315), type species Lenticeras jullieni Pervinquière, 1910, p. 69, pl. 12 (3), fig. 24, text-fig. 34 was regarded as a synonym of Eulophoceras by Wright (1957, p. L436; 1996, p. 204); Spath (1922, p. 143) had previously regarded it as belonging to the Coniacian Tissotiidae Hyatt, 1900. Lenticeras jullieni is known from the holotype of the type species only, an unregistered specimen in the Sorbonne collections, now housed in the Muséum National d'Histoire Naturelle, Paris. It is a composite mould of a battered, worn, and corroded phragmocone 91 mm in diameter. An oxycone, the original proportions cannot be established because of the damage. The tiny shallow umbilicus comprises an estimated 5.5% of the diameter, the umbilical wall inclined outward to form a conical pit. An estimated 12 primary ribs arise at the umbilical seam, and are straight and prorsiradiate on the flanks, across which they initially broaden and strengthen progressively, before declining and effacing on the outer flank and not extending to the venter. Most ribs bifurcate around mid-flank. The adapical secondary rib of each pair is markedly rursiradiate, so that the primary and secondary ribs have the form of a reversed letter Y. Some strengthen on the outer flank into a low rounded swelling or incipient bulla before effacing. The suture, as depicted by Pervinquière (1910, text-fig. 34) has broad, only moderately incised bifid saddles, with narrower, little-incised lobes. The specimen is from Les Tamarins, southeast of Batna, Algeria, and was said to be associated with Tissotia robini Thiollière (Pervinquière, 1910, p. 71), a junior synonym of the Middle Coniacian Metatissotia ewaldi (Von Buch, 1848) (Kennedy, 1984, p. 127). If correctly dated, there is a substantial gap between Middle Coniacian Praelibycoceras and Upper Santonian-Lower Campanian Eulophoceras, while the suture of the former lacks the distinctive features of the suture of the latter. In fact, the oxycone shell form is the main common feature they share. Accordingly, we see no strong evidence to link Praelibycoceras and Eulophoceras; the familial position of the former remains uncertain in our view.
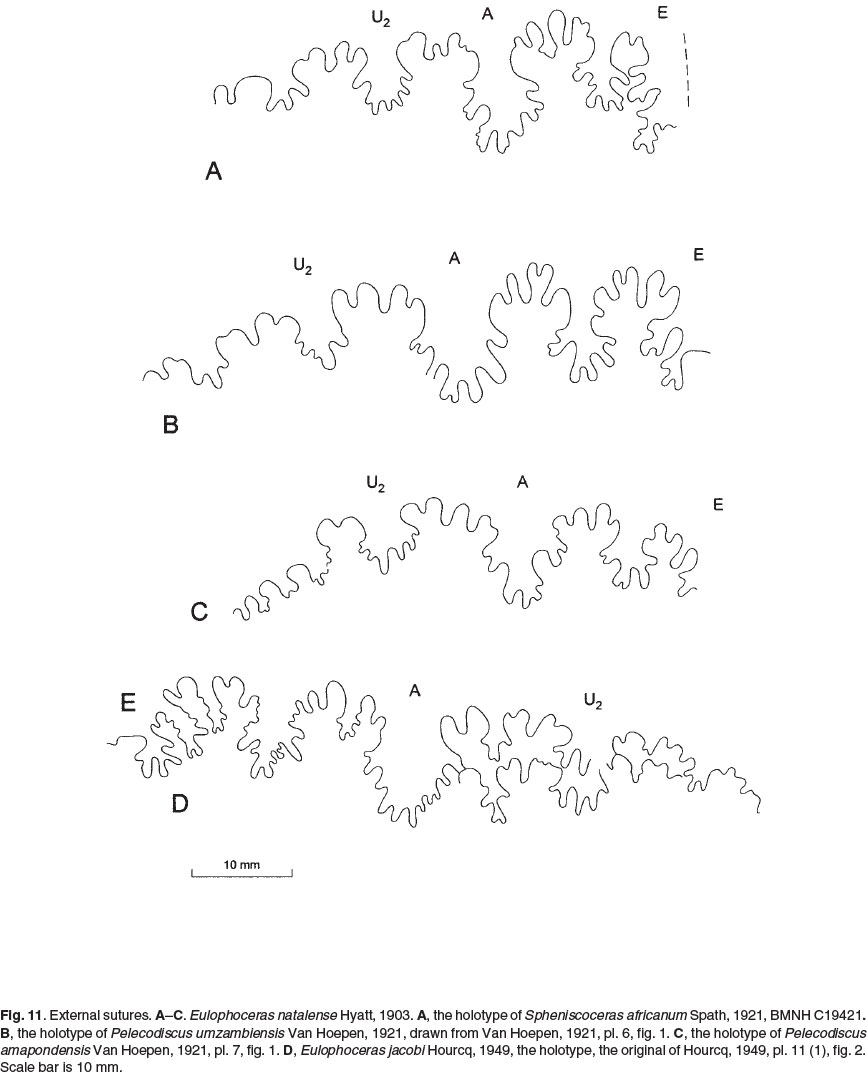
Spheniscoceras Spath, 1921, p. 242, was introduced as Spheniscoceras Crick MS, but must be attributed to Spath. His comments (footnote on pp. 242-243) on Crick's manuscript are less than positive 'In fact the writer is not convinced that Crick was right in separating the forms he described [Spheniscoceras africanum, minor, and tenue, G.C. Crick MS] from Eulophoceras.' In spite of this doubt, Spath separated Spheniscoceras from Eulophoceras and designated S. africanum (Crick MS) as type species (explanation of text-fig. C, 1921, p. 243) based on a figure of a suture line only (Spath, 1921, p. 243, text-fig. C1a). The holotype of africanum (Figs 6, 11A) is from the Mzamba Formation at the mouth of the Mzamba River in the Eastern Cape Province, as is the holotype of Eulophoceras natalense. There are no significant differences between the two, as described below, and Spheniscoceras is a junior subjective synonym of Eulophoceras (Spath continued to use Spheniscoceras in his 1922 revision of the ammonite fauna of Pondoland, but it is very much out of respect for Crick '...Hyatt's form [Eulophoceras natalense] may only be one and an uncommon type of the development separated by Crick and again by Dr Van Hoepen on account of minute differences in what appears to be a very variable suture-line' (p. 142). No type species was designated for Pelecodiscus Van Hoepen, 1921, p. 30; P. umzambiensis Van Hoepen, 1921, p. 30, pl. 5, fig. 10, pl. 6, fig. 1, is here designated type species. Van Hoepen separated his new genus from Eulophoceras on the basis of details of the suture line, but as described below, sutures are variable both within and between individuals referred to E. natalense, and we follow previous authors in regarding Pelecodiscus as a synonym of Eulophoceras.
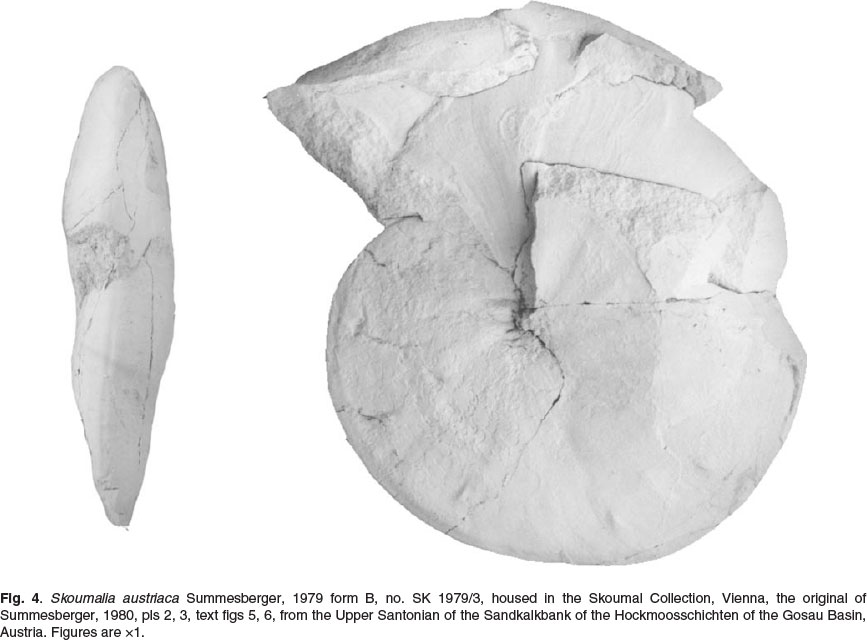
Kennedy (1987, p. 776; in Kennedy et al., 1995, p. 425) and Wright (1996, p. 204) regarded Skoumalia Summesberger, 1979, p. 141, type species Skoumalia austriaca Summesberger, 1979 (p. 141, pl. 9, figs 37-41, text-figs 26-30), from the Upper Santonian of the Gosau Basin, Austria, as a synonym of Eulophoceras. Summesberger recognized two morphotypes in Skoumalia austriaca. His Form A included the holotype (Summesberger, 1979, pl. 9, figs 37, 38; text-fig. 26-28) an oxycone phragmocone 75 mm in diameter with fastigiate venter, strong umbilical bullae that give rise to pairs of straight prorsiradiate ribs that terminate in small ventral bullae. The suture has a broad E/A with a deep median incision. The suture of the holotype (Summesberger, 1979, text-fig. 27) lacks the ventral part, but the second specimen has a bifid E/A (Summesberger, 1979, fig. 28). A is deep and narrow, A/U2 small and bifid. The ornament is that of Diaziceras Spath, 1921, while E/A is asymmetrically bifid, rather than trifid as in Eulophoceras. Form B of Summesberger (1979, p. 143, pl. 9, figs 39-41; text figs 29, 30) was based on a phragmocone 58.7 mm in diameter; according to Summesberger (1979, p. 143) it differs from form A in its flat, discoidal shell, narrow umbilicus, and lack of umbilical tubercles; also, the lateral ornament is less conspicuous. They were regarded as probable sexual dimorphs. The suture has a broad, subtrifid E/A (Summesberger, 1979, fig. 30). Summesberger (1980, p. 280, pl. 2, figs 5-6; pl. 3, figs 7-8; text figs 5,6) subsequently described and figured what he believed to be the adult of his Form B, 122 mm in diameter ornamented by delicate umbilical bullae on the phragmocone. The outer whorl bears delicate feebly flexuous growth lines and striae, with ten distant outer lateral bullae (Fig. 4). The suture has a broad subtrifid E/A, deep, narrow A, bifid A/U2, and simplifying auxiliary elements.
Form A closely resembles Diaziceras guillantoni Hourcq, 1949, p. 108 (22), pl. 12 (2), fig. 1, from the Upper Santonian of Madagascar according to Hourcq, but actually Lower Campanian (after Collignon, 1969, p. 212) in whorl section, ornament, and may even be conspecific, so that Skoumalia should be regarded as a synonym of Diaziceras, an hypothesis to be developed elsewhere. In contrast, Form B of Summesberger, with compressed whorl, fastigiate venter, near smooth outer whorl, tiny outer lateral bullae in the adult, is close to Eulophoceras jacobi Hourcq, 1949, p. 95 (9), pl. 11 (1), fig. 2, text-fig. 7 (Fig. 11D, 12A-C), from the Lower Campanian of Madgascar. They differ in that the outer lateral bullae are prorsiradiate in austriaca, but swept back and markedly rursiradiate in jacobi. The following species are assigned to Eulophoceras:
1. E. natalense Hyatt, 1903, of which E. africanum (Spath, 1921), E. minor (Spath, 1921), E. tenue (Spath, 1921), E. umzambiensis (Van Hoepen, 1921), E. capensis (Van Hoepen, 1921), and E. amapondensis (Van Hoepen, 1921), are synonyms.
2. E. bererensis Hourcq, 1949, of which E. miloni Hourcq, 1949, E. besairiei Hourcq, 1949, E. manambolense Hourcq, 1949, and E. hourcqi Collignon, 1969, are synonyms.
3. E. jacobi Hourcq, 1949.
4. Eulophoceras vautrini Basse, 1954
5. Eulophoceras sp. of Reyment (1955, p. 83, pl. 22, fig. 5; text-fig. 42).
6. Eulophoceras wollmanae Young, 1963.
7. Eulophoceras sp. (= Eulophoceras jacobi Renz, 1982, non Hourcq, p. 111, pl. 36, figs 10, 11; pl. 37, figs 2, 3; text-fig. 86a-b).
8. Eulophoceras losaense Santamaria Zabala, 1995.
Eulophoceras berryi Knechtel, 1947 (p. 125, pl. 43, text-fig. 21) is based upon a single specimen from Rio Cuxiabatay, Loreto, in the Peruvian Andes. It occurs in association with three species of Tissotia introduced by Knechtel, and is presumably of Coniacian age. It has a distinctive ornament of convex growth lines only, and a suture with an asymmetrically bifid E/A, and very narrow A. We are uncertain as to its generic position.
The generic assignation of Eulophoceras phansalkari Ghare, 1969 (p. 165, pl. 4, figs 1-4; text-fig. 1) is problematic in our view. It is based on the holotype only, a half whorl of worn and corroded phragmocone 140 mm in diameter, from the Upper Trichinopoly Group of Iluppakudi in the Trichinopoly District of Tamil Nadu, South India. It was described as having very feeble ornamentation of fold-like ribs of which five or six are just visible round the umbilicus, and differing from E. natalense in '...not having the ornamentation of weak ribs, crowded sutures with lobes of at least three consecutive sutures telescoping into one another ... the folioles of E. natalense are twice longer than those of the present species.'
The Eulophoceras sp. of Reyment (1955, p. 83, pl. 22, fig. 5; text-fig. 42) was based on a fragment of phragmocone only, with faint low undulations on the flank, and a suture with a very broad asymmetric E/A with large median element. The poor preservation makes generic assignation questionable. It is from the upper part of the Awgu-Ndeaboh Shale of Onitscha province, Nigeria, and dated as Coniacian by Reyment.
The Eulophoceras jacobi Hourcq of Renz (1982, p. 111, pl. 36, figs 10, 11; pl. 37, figs 2, 3; text-fig. 86a-b) from the Coniacian of Venezuela, differs from Hourcq's species in having radial ribs that flex forwards and are concave on the outermost flank, whereas the ribs flex back and are concave on the outer flank in jacobi. The specimens are between 25 and 55 mm in diameter, much smaller than the Madagascan material; they are best referred to as Eulophoceras sp. juv. Pattaroyo & Dueñas (2006, p. 504, pl. 1, figs 1-3) described and figured a specimen with a diameter of ca. 150 mm with weak ventrolateral ribs and striae on the inner, lateral flanks as Eulophoceras jacobi from the Lower Coniacian of Quebrada Bambucá, Aipe-Huila Colombia, which, judging by their synonymy, they regarded as the same as Renz's (1982) species and questionably that of Reyment (1955). This Coniacian specimen certainly does not belong to E. jacobi, a lower Campanian species. This may belong to a new species, but with no further material, we provisionally regard it as Eulophoceras sp.
Eulophoceras grossouvrei Collignon 1983 (p. 205, pl. 7, fig. 5; see Kennedy, Bilotte & Melchior, 1995, p. 426, pl. 25, figs 1-2) is from the Upper Santonian of the Corbières in southern France. It is a worn juvenile 55 mm in diameter, a nomen dubium in our view. Referred to as Eulophoceras sp. juv. by Kennedy et al. (1995) it is without ornament and shows no diagnostic features.
Occurrence
Coniacian of Colombia, and possibly Nigeria. Santonian of KwaZulu-Natal, South Africa, Austria, southern France, and northern Spain, and Syria? Lower Campanian of KwaZulu-Natal, Eastern Cape Province, Madagascar, and Angola.
Eulophoceras natalense Hyatt, 1903
1903 Eulophoceras natalense Hyatt, p. 86, pl. 11, figs 2-6.
1906 Eulophoceras natalense Hyatt; Woods, p. 337, pl. 42, fig. 3.
1921 Eulophoceras natalense Hyatt; Spath, footnote # on p. 242; text-fig. C-2.
1921 Spheniscoceras africanum Crick MS; Spath, p. 242, footnote # on p. 242, text-fig. C-1a.
1921 Spheniscoceras minor Crick MS; Spath, p. 242, footnote # on p. 242, text-fig. C-1b.
1921 Spheniscoceras tenue Crick MS; Spath, p. 242, footnote # on p. 242, text-fig. C-1c.
1921 Eulophoceras natalense Hyatt; Van Hoepen, p. 30, pl. 6, figs 2, 3.
1921 Pelecodiscus umzambiensis Van Hoepen, p. 30, pl. 5, fig. 10;pl. 6, fig. 1.
1921 Pelecodiscus capensis Van Hoepen, p. 32, pl. 5, fig. 11.
1921 Pelecodiscus amapondensis Van Hoepen, p. 33, pl. 7, fig. 1, 2.
1921 Eulophoceras natalense Hyatt; Van Hoepen, pl. 6, fig. 2 only.
1922 Eulophoceras natalense Hyatt; Spath, p. 142
1922 Spheniscoceras africanum (Crick MS); Spath, p. 143, pl. 6, fig. 1.
1922 Spheniscoceras tenue (Crick MS); Spath, p. 144, pl. 7, fig. 3; pl. 8, fig. 3.
1922 Spheniscoceras minor (Crick MS); Spath, p. 144, pl. 6, fig. 2.
1922 Spheniscoceras amapondense v. Hoepen sp.; Spath, p. 144, pl. 7, fig. 2.
1922 Spheniscoceras umzambiense v. Hoepen; Spath, p. 145.
1982 Eulophoceras natalense Hyatt, 1903; Immel, Klinger & Wiedmann, p. 24, pl. 8, fig. 5.
1987 Eulophoceras natalense Hyatt, 1903; Immel, p. 113.
1988 Eulophoceras natalense Hyatt; Cooper, p. 209, figs 1a, b. Non 1995 Eulophoceras cf. natalense Hyatt, 1903; Kennedy in Kennedy, Bilotte & Melchior, p. 235, pl. 26, figs 3, 4, 7; text-fig. 32 (=Eulophoceras losaense Santamaria Zabala, 1995).
Type
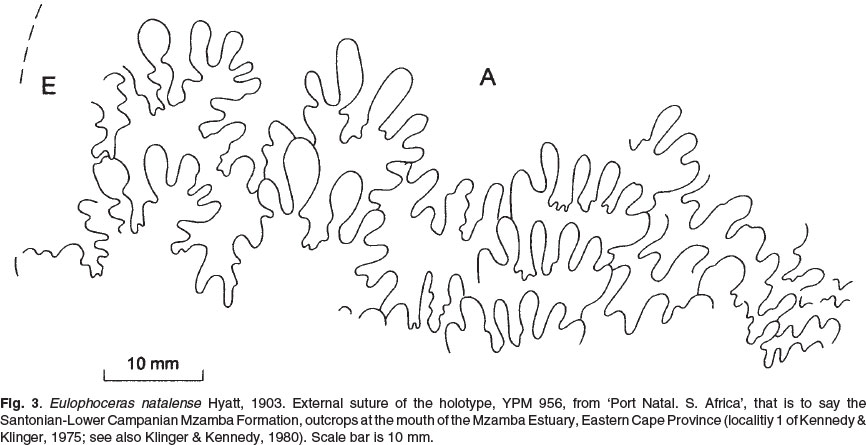
The holotype is YPM 956 (Figs 1-3), the original of Hyatt (1903, pl. 11, fig. 2), from 'Port Natal. S. Africa.'
Material
From unspecified horizons in the Mzamba Formation at the mouth of the Mzamba Estuary, Eastern Cape Province: BMNH C19421, the holotype of Spheniscoceras africanum Spath, 1921, p. 242, footnote # on p. 242, text-fig. C-1a; BMNH C19423, the holotype of Spheniscoceras minor Spath, 1921, p. 242, footnote # on p. 242, text-fig. C-1b; BMNH C19423, the holotype of Spheniscoceras tenue Spath, 1921, p. 242, footnote # on p. 242, text-fig. C-1c; BMNH C19424, the original of Spheniscoceras amapondense of Spath (1922, p. 144, pl. 7, fig. 2); the holotype of Pelecodiscus umzambiensis Van Hoepen, 1921, p. 30, pl. 5, fig. 10; pl. 6, fig. 1, and two additional paratype specimens referred to the species by Van Hoepen, the holotype of Pelecodiscus capensis Van Hoepen 1921, p. 32, pl. 5, fig. 11, the holotype of Pelecodiscus amapondensis Van Hoepen, 1921, p. 33, pl. 7, fig. 1, 2, all in the collections of the Ditsong National Museum of Natural History, (formerly the Transvaal Museum; Northern flagship Institution), Pretoria., SAM-PCP019070, PCP006727, 019752, 019749, 019753019764, 0201244, from bed 7 or 8, PCP20144, 19733, 19750, 19754, 19784 presumably from the same horizon, of the Mzamba Formation, Campanian I at locality 1 of Kennedy and Klinger (1975, p. 281), at the mouth of the Mzamba Estuary, Eastern Cape Province. OUM KX5348 from the St Lucia Formation, Santonian III, in the environs of locality 17 of Kennedy & Klinger (1975, p. 283), southeast of Mtubatuba, KwaZulu - Natal. OUM KX12925, 12926, SAM-PCZ007346, 019750-019752, 019785, 27346 from the St Lucia Formation, Campanian I-II, locality74 of Kennedy & Klinger (1975, p. 292), Die Rooiwalle, western False Bay, Lake St Lucia, KwaZulu-Natal.
Description
The holotype (Figs 1-3) is a phragmocone retaining most of the original shell on one side (Fig. 1). The other side (Fig. 2), that illustrated by Hyatt (1903, pl. 11, fig. 2) with only traces of shell remaining is labelled 'Port Natal. S. Africa.' The specimen has been sawn, and the section polished (Fig. 2). It is interpreted as a macroconch phragmocone, with an estimated original diameter of 164 mm. It is oxycone, with a tiny conical umbilicus 4 mm deep, that comprises approximately 2% of the diameter. The shell of the succeeding whorls completely infills the umbilicus of the preceding whorls. The minimum preserved whorl height is 62 mm, where the whorl breadth to height ratio is 0.37; the ratio is 0.38 at a whorl height of 91.3 mm, beyond which the venter is damaged, rendering accurate measurement impossible. The umbilical shoulder is broadly rounded, the whorl section lanceolate. There are eight low, broad, prorsiradiate fold-like ribs on the half whorl preserved. They arise on the umbilical shoulder, broaden and strengthen progressively across the inner three quarters of the flanks, thereafter effacing, leaving the outermost flanks smooth. On the basis of the size ratio of phragmocone to adult shell in other Eulophoceras, the complete diameter of this specimen may have been well over 200 mm. None of the specimens has the aperture preserved, but PCP019762 retains the apertural margin, which is concave across the dorsal 75% of the flank.
The sutures (Fig. 3) interfere throughout. E/A is deeply incised and asymmetrically bifid, with elongated folioles; A is broad; U2 is deeply incised with very elongate narrow-necked folioles.
Woods' specimen of Eulophoceras natalense (1906, p. 337 pl. 42, fig. 3; Fig. 5A herein) is embedded in a block of matrix, has a maximum preserved diameter of 131 mm, and retains extensive areas of original shell material. It is almost wholly septate, and is interpreted as the macroconch phragmocone of a feebly ornamented variant of the species, with low, broad, prorsiradiate ribs detectable under oblique light to a diameter of 112 mm.
BMNH C19421, the holotype of Spheniscoceras africanum Spath, 1921 (p. 242, text-fig. C-1a; 1922, p. 143, pl. 6, fig. 1; Figs 5B, 6, 11A herein), is an incomplete macroconch just over 142 mm in diameter, with a 180° sector of the incomplete body chamber preserved. The dimensions are as follows:
D:142.0 (100) Wb: 33.7 (23.7) Wh: 84.5 (59.5) Wb:Wh: 0.40 U: 6.4 (4.5)
The specimen retains extensive areas of variably exfoliated shell, and is oxycone, with the greatest breadth low on the flanks, and a whorl breadth to height ratio of 0.40. On the phragmocone, broad distant primary ribs arise on the umbilical shoulder, some of them incipiently bullate. They give rise to low, broad, feebly prorsiradiate ribs that bifurcate low on the flanks, with additional ribs intercalating, the secondary and intercalated ribs low and broad, and strengthening at their termination on the outermost flank: they do not extend to the ventral keel. The ornament is relatively strong on adapical 120° sector of the outer whorl, thereafter declining, so that the adapertural part of the body chamber is near-smooth. The suture (Fig. 11A) has a broad, strongly asymmetric, incipiently trifid E/A with a deep, narrow median incision and moderately elongated folioles. A is narrower, and asymmetrically bifid, U2 plump, bifid, with minor incisions.
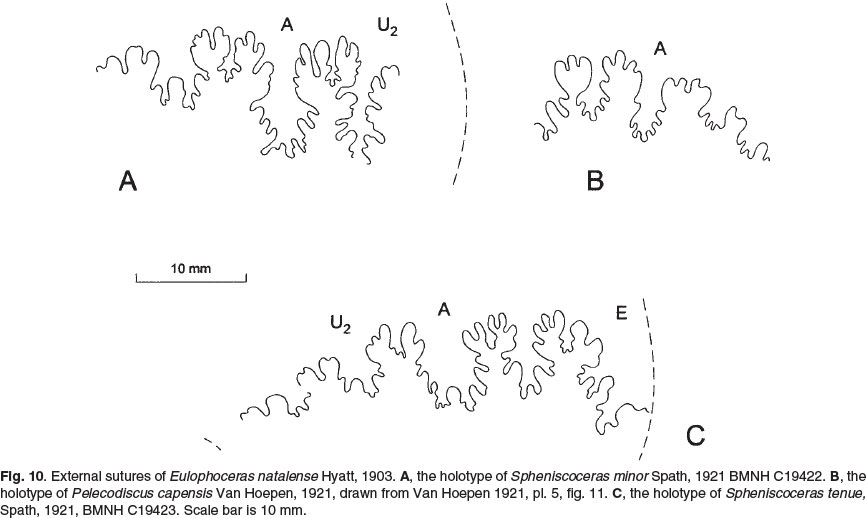
BMNH C19422, the holotype of Spheniscoceras minor Spath, 1921(p. 242, text-fig. C-1b; 1922, p. 144, pl. 6, fig. 2; Figs 8A,B, 10A), retains extensive areas of original aragonitic shell, and is septate to a diameter of 86 mm, with indications of the original presence of a 240° sector of body chamber. The last-formed sutures are crowded, indicating the specimen to be adult. The shell is oxycone, with a tinyumbilicus. The whorl breadth to height ratio is 0.44, the greatest breadth is low on the flanks. The dimensions are as follows:
At: D: 97.2 (100) Wb: 27.8 (28.6) Wh: 58.5 (60.2) Wb:Wh: 0.48 U:-(-).
There are five low, broad ribs on the adapertural half whorl of the phragmocone. They arise on the umbilical shoulder and sweep forwards on the inner flanks, weakening at mid-flank and branching into two or three secondary ribs, with additional ribs intercalating between the groups of secondaries to give as many as 20 low, broad ribs that extend to the sharp venter. This ornament declines rapidly and effaces on the body chamber, which is near-smoooth.
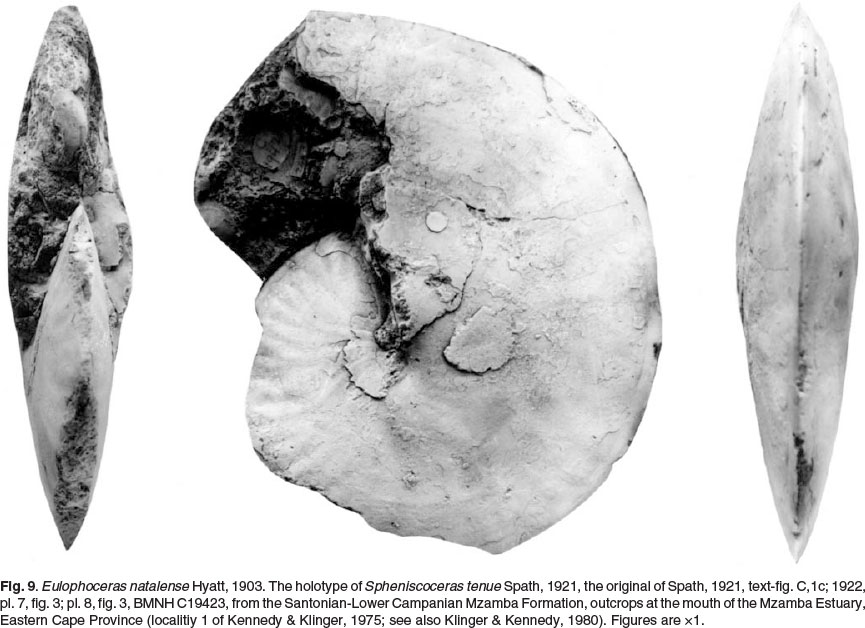
BMNH C19423, the holotype of Spheniscoceras tenue Spath, 1921 (p. 242, text-fig. C-1c; 1922, p. 144, pl. 7, fig. 3; pl. 8, fig. 3; Figs 9, 10C herein), is interpreted as a micro-conch. It consists of a phragmocone 79 mm in diameter, with a 240° sector of body chamber preserved. The dimensions are as follows:
At: D: 122.6 (100) Wb: 29.0 (23.7) Wh: 71.2 (58.1) Wb:Wh: 0.41 U: -(-)
The shell is oxycone, more compressed than the holotype of Spheniscoceras minor, the greatest breadth low on the flanks, with a whorl breadth to height ratio of 0.41 and, with a comparable style of flank ornament, but much weaker. The suture (Fig. 10C) has abroad, deeply incised E/A with a deep, narrow median element and moderately elongated folioles; A is narrow and asymmetrically bifid.
BMNH C19424 (Fig. 8C) was referred to Spheniscoceras amapondense Van Hoepen, 1921, by Spath (1922, p. 144, pl. 7, fig. 2), and is a fragment of the outer flank of a macro-conch phragmocone.
The holotype of Pelecodiscus umzambiensis Van Hoepen, 1921, is the original of his pl. 5, fig. 10; pl. 6, fig. 1, in the collections of the Ditsong National Museum of Natural History (formerly the Transvaal Museum, Northern flagship Institution). It is 124 mm approximately in diameter, and retains a short section of body chamber. It is a moderately robustly ornamented individual, corresponding in this respect to the holotypes of natalense and tenue of Spath (Figs 8, 9A,B). The suture (Fig. 11B) also differs in no significant respects.
The holotype of Pelecodiscus capensis Van Hoepen, 1921 (p. 32, pl. 5, fig. 11), housed in the collections of the Ditsong National Museum of Natural History (formerly the Transvaal Museum, Northern Flagship Institution), Pretoria is a battered individual just over 50 mm in diameter. Van Hoepen differentiated it on the basis of details of the suture (Fig. 10B), specifically the 'long and narrow saddles and lobes external to the third lateral saddle. The internal portion runs much stronger backwards' (p. 33).
The holotype of Pelecodiscus amapondensis Van Hoepen (1921, p. 33, pl. 7, figs 1, 2) is based on a fragment of phragmocone with a whorl height of over 70 mm. It was separated from umzambiensis on the basis of details of the suture, but this differs in no significant respects from that of africanum of Spath (compare Figs 11A and C).
Discussion
The specimens referred here to Eulophoceras natalense all share a common style of weak to near effaced ribbing (Fig. 7), and in this respect clearly belong to a single dimorphic species. The species recognized by Spath (1921, 1922) and Van Hoepen (1921) were all differentiated on details of the external suture line. But as can be seen from Figs 2, 8C, 10, and 11A-C, no two specimens have identical suture lines, and sutures are not a basis for differentiating a series of species within the present material. Even Spath (1922) who recognized one species of Eulophoceras and five of Spheniscoceras from the Mzamba Formation clearly had considerable doubts on their distinctiveness based on minute differences of suture line: '.the four 'species' of Crick are very close to Eulophoceras natalense Hyatt, and it might be held that the development of what may be a thinner keel, and the slight differences in suture line and ornamentation, are not sufficient to justify the creation of even different species.' (1922, p. 142).
His comments on BMNH C19424 (Fig. 8C) a fragment that he referred to Spheniscoceras cf. amapondense of Van Hoepen are even more telling:
'...A fragment ... agrees with the smooth outer whorl of the type-species, S. africanum, but its suture-line differs from those of the known 'species' of this genus, notably in the external saddle. The suture line of S. amapondense is probably nearer than any other to that of the present fragment, but the half of the suture line of S. africanum, opposite to that figured [Spath, 1922, pl. 7, fig. 2 is the figure referred to here] is also similar, so that by splitting one of these specimens in two a further 'species' could be established.' In other words, the differences between the sutures on the left and right flanks of this fragment are as great as that between the 'species' Spath recognized.
Eulophoceras jacobi Hourcq, 1949 ( p. 95, pl. 11 (1), fig. 2, text-fig. 7) was based on a number of specimens from south of Berere, Madagascar, which Hourcq assigned to the top of the Santonian. Collignon (1969, p. 204, pl. 600, fig. 2253) figured a specimen from the Hourcquiella bererensis Subzone of his Lower Campanian Zone à Anapachydiscus wittekindi et Eulophoceras jacobi. The original of Hourcq's pl. 11 (1), fig. 2, text-fig. 7, an unregistered specimen in the École des Mines Collections, now housed in the Université Claude Bernard, Lyon, is here designated lectotype, and illustrated here as Fig. 12A-C. Its dimensions are as follows:
D: 119 (100) Wb: 25.0 (0.21), Wh: 70.0 (0.59), Wb:Wh: 0.36, U: 2.7 (2.3)
The specimen is an internal mould with traces of recrystallized shell, retaining a 60° sector of body chamber, an oxycone, with a whorl breadth to height ratio of 0.36, the greatest breadth well below mid-flank. Ornament is greatly reduced. Spiral ridges and depressions are prominent on the outer flank of the mould, and where shell is preserved on the phragmocone, delicate growth lines and striae arise in bunches at the umbilical shoulder and sweep forwards across the flanks, together with feeble low, broad, prorsiradiate ribs. These strengthen and flex back on the outer flank, where theyare markedly rursiradiate and feebly crescentic, extending across the outer third of the flank, but failing to reach the venter. On the adapical half of the outer whorl the ribs terminate in a feeble bulla. The ribs decline towards the adapertural end of the phragmocone, where the ornament consists of delicate falcoid growth lines and striae, straight and prorsiradiate to mid-flank, thereafter flexed back on the outer two thirds of the flank. The sutures are approximated, and the shell margin is flared at the greatest preserved diameter. If this is an indication of maturity, the body chamber was extraordinarily short. The suture (Fig. 11D) has a very wide subtrifid E/A with a deep median incision, and folioles of moderate length; A is broad and bifid, with numerous minor incisions; U2 is asymmetrically bifid. The species is immediately distinguished from E. natalense on the basis of the course of the ribs, which are markedly rursiradiate on the outer flank, and terminate in distinctive bullae.
Eulophoceras bererense Hourcq, 1949, of which E. miloni Hourcq, 1949, E. besairiei Hourcq, 1949 E. manambolense Hourcq, 1949, and E. hourcqi Collignon, 1969, are synonyms, is described below. It differs from E. natalense in its stouter shell, the body chamber of which broadens markedly with a rounded venter at the aperture. The primary ribs are broader and coarser than those of natalense, flex back, coarsen, and are markedly concave on the outer flank.
Eulophoceras vautrini Basse, 1954 (p. 869, pl. 27, figs 1a-c) is based on a single specimen from the Upper Santonian or Lower Campanian of 'Forlos', southeast of Palmyra in Syria. The species is based on a single battered individual 73 mm in diameter, the adapertural 180° sector body chamber. The flanks are ornamented by strong, concave ribs on the outer flank, and the suture has a broad, subtrifid E/A, and an A/U2 with a pair of distinctive narrow-necked bulbous folioles.
Eulophoceras wollmanae Young, 1963 (p. 126, pl. 72, fig. 5; pl. 74, figs 1-3, 6; text figs 11c,g,m,s) from the Lower Campanian of Texas has a less compressed whorl section, and is described by its author as being smooth throughout its ontogeny, while the poorly preserved sutures are rather different.
Eulophoceras losaense Santamaria Zabala, 1995 (p. 98, pl. 5, figs 3-4; text-fig. 1f), from the Santonian of northern Spain, is distinguished by having ribs that flex back and are markedly rursiradiate and concave on the middle and outer flank.
The Eulophoceras natalense of Immel et al. (1982, p. 24, pl. 8, fig. 5) from the Gosau Group of Brandenberg Tirol, Austria, was originally described as being from the Lower Santonian, but Immel (1987, p. 113) subsequently suggested it might range into the Lower Campanian. It is a composite mould 77 mm in diameter, with low broad ribs well-developed on the adapertural half whorl, as in the holotype of Spath's tenue (Fig. 9) at a comparable diameter. Specimens referred to as Eulophoceras cf. natalense from the Santonian of the Corbières in southern France (Kennedy in Kennedy et al., 1995, p. 425, pl. 26, figs 3, 4, 7; text-fig. 32) have ribs that flex back and are markedly concave on the outer flank, and are better referred to E. losaense, discussed above.
Occurrence
Eulophoceras natalense occurs at the contact of beds A7 and A8 and in bed A8 of the Mzamba Formation at Mzamba Cliff in Eastern Cape Province (Klinger & Kennedy, 1980, p.216, text-fig. 4), where it is associated with Submortoniceras condamyi (Collignon), and probably Hauericeras gardeni (Baily) and Pseudoschloenbachia umbulazi (Baily), indicating Campanian I, the lowest division of the Campanian of Kennedy & Klinger (1975). It occurs in the St Lucia Formation, Santonian III, in the environs of locality 17 of Kennedy & Klinger (1975), southeast of Mtubatuba, associated with Hauericeras gardeni, Pseudoschloenbachia umbulazi (Baily, 1855), and Anagaudryceras politissimum (Kossmat, 1895) At Die Rooiwalle, western False Bay, Lake St Lucia, KwaZulu-Natal, locality 74 of Kennedy & Klinger (1975), it ranges from Santonian III to Campanian I. Records from Austria are Lower Santonian to Lower Campanian (Immel, 1987, p. 113). Lower Campanian of Angola.
Eulophoceras bererense Hourcq, 1949
1949 Eulophoceras Bererense Hourcq, p. 91 (5), pl. 11 (1), text figs 1, 2.
1949 Eulophoceras Miloni Hourcq, p. 93 (7), pl. 11 (1), fig. 3; text-fig. 3.
1949 Eulophoceras Besairiei Hourcq, p. 94 (8), pl. 11 (1), fig. 4; text figs 4, 5.
1949 Eulophoceras Manambolense Hourcq, p. 96, pl. 13(3), fig. 2; text-fig. 8.
1969 Eulophoceras bererense Hourcq; Collignon, p. 204, pl. 600, fig. 2254.
1969 Eulophoceras hourcqi Collignon, p. 206, pl. 601, fig. 2255.
1969 Eulophoceras miloni Hourcq; Collignon, p. 206, pl. 601, fig. 2256.
1969 Eulophoceras besairiei Hourcq; Collignon, p. 208, pl. 602, fig. 2257.
Name of the species
As discussed below, we believe that Eulophoceras bererense Hourcq, 1949, Eulophoceras miloni Hourcq, 1949, E. besairiei Hourcq, 1949, and E. manambolense Hourcq, 1949, are conspecific. As first revising authors, we select the name bererense for the species.
Types
Hourcq mentioned 12 specimens in his account of E. bererense (1949, p. 91 (5)), all of which ranked as syntypes. We here designate the original of Hourcq, 1949, pl. 11 (1), fig. 1, lectotype. It is re-illustrated here as Figs 13A, B, 17B, and is an unregistered specimen in the collections of the École des Mines, Paris, now housed in the collections of the Université Claude Bernard, Lyon. The types are from Berere, Madagascar, and were referred to the top of the Santonian by Hourcq.
Material
OUM KX12924, from the St Lucia Formation, Campanian I-II, at locality 74 of Kennedy & Klinger (1975), Die Rooiwalle, False Bay, western Lake St Lucia, KwaZulu-Natal.
Description
The lectotype is an internal mould of an adult microconch with a 240° sector of body chamber preserved. The dimensions are as follows:
D:125 (100) Wb:33.0(26.4) Wh: 66.5 (53.2) Wb:Wh: 0.5 U: 7.9 (6.1)
The phragmocone is oxcone; the body chamber broadens markedly, and develops an obtusely fastigiate, rounding venter. On the adapical half of the outer whorl six low broad ribs arise at the umbilical seam, strengthen across the flanks, and bifurcate around mid-flank, while occasional shorter ribs intercalate. The ribs sweep back and are concave and rursiradiate on the outer flank, thicken, and terminate before reaching the venter, to give a total of 13 ribs at the ventrolateral shoulder. Similar but more subdued ornament is present on the adapical half of the body chamber; the adoral half is smooth. Suture (Fig. 17B) moderately incised with short folioles; E/A verywide, asymmetrically bifid, with a large median element; A narrow; A/U2 broad, asymmetrically bifid.
Hourcq mentioned four specimens in his account of Eulophoceras miloni, which rank as syntypes. We designate the original of his pl. 11 (1) fig. 3 lectotype, and illustrate it here as Figs 13D, 17A. It is an unregistered specimen in the collections of the École des Mines, Paris, now housed in the collections of the Université Claude Bernard, Lyon. The types are from Berere, Madagascar, and were referred to the top of the Santonian by Hourcq.
The lectotype is an internal mould of a microconch of a feebly ornamented variant. The dimensions are as follows:
D: 107 (100) Wb: 126.5 (24.8) Wh:61.5 (57.5) Wb;Wh 0.43U: 5.5 (5.1)
Oxycone, with a 120° sector of somewhat stouter incomplete body chamber preserved. Ornament is conspicuous on the phragmocone only. Delicate prorsiradiate primary ribs, riblets and striae arise on the umbilical shoulder and sweep forwards and are prorsiradiate and feebly concave on the inner 60% of the flanks, thereafter sweeping back, and broadening into concave blunt ribs, 12 per half whorl, that decline before reaching the venter. In some cases two inner flank ribs link to a single outer flank rib. On the body chamber the course of the growth lines remains falcoid, but the outer flank ribs are reduced to mere feeble undulations. Suture (Fig. 17A) with very broad asymmetrically bifid E/A, more deeply incised than in the lectotype of bererense.
Hourcq mentioned two specimens in his account of Eulophoceras besairiei, which rank as syntypes. We designate the original of his pl. 11 (1) fig. 4 lectotype, and illustrate it here as Fig. 12D. It is an unregistered specimen in the collections of the École des Mines, Paris, now housed in the collections of the Université Claude Bernard, Lyon. The types are from Berere, Madagascar, and were referred to the top of the Santonian by Hourcq.
The lectotype is an internal mould of a microconch with a 240° sector of body chamber preserved. The dimensions are as follows:
D: 134 (100) Wb:28.5 (21.3) Wh: 70.5 (52.6) Wb:Wh: 0.4 U: -(-)
The phragmocone is an oxycone, the body chamber somewhat stouter, with an obtusely fastigiate venter that rounds towards the adapertural end. The specimen is closely similar to the lectotype of Eulophoceras bererense, but with more delicate ornament of an estimated fourteen primary ribs on the inner flank. The ribs increase by bifurcation and intercalation to give an estimated total of 28 ribs per whorl at the ventrolateral shoulder. The suture (Hourcq, 1949, text-fig. 5) is essential the same as that of the lectotype of E. bererense.
The holotype, by monotypy, of Eulophoceras manambolense Hourcq, 1949, is the original of his p. 96, pl. 13 (3), fig. 2; text-fig. 8. It is an unregistered specimen in the collections of the École des Mines, Paris, now housed in the collections of the Université Claude Bernard, Lyon. It is from Berere, Madagascar, and was referred to the top of the Santonian by Hourcq.
The specimen is a small corroded phragmocone, with the following dimensions:
D: 81.0 (100) Wb: 21.5 (26.5) Wh: 47.0 (58.0) Wb:Wh: 0.54 U: 5.3 (6.5)
It is a relatively stout oxycone with a tiny umbilicus. The greatest breadth is just outside the umbilical shoulder, the inner to mid-flank region broadly convex, the outer flanks convergent, and feebly concave on either side of the sharp venter. Seven low, broad, straight prorsiradiate ribs arise at the umbilical shoulder of the outer whorl and bifurcate below mid-flank to give rise to pairs of feebly concave rursiradiate ribs that efface before reaching the venter. Additional ribs intercalate at the level that the primary ribs bifurcate, to give a total of 26-28 ribs on the outer flank of the outer whorl. The suture (Hourcq, 1949, text-fig. 8) differs in no significant respects from that of E. bererense.
The holotype of Eulophoceras hourcqi Collignon, 1969 (p. 206, pl. 601, fig. 2255) is from the Lower Campanian of Ampamba-Antsirasira (Belo-sur-Tsiribihina), Madagascar (Fig. 15); it is housed in the Collignon Collection in the Université de Bourgogne, Dijon. It is an internal mould of an adult microconch 119 mm in diameter, with a 200° sector of body chamber preserved. As in previous specimens, the phragmocone is oxycone, the body chamber broadening, with a fastigiate venter that rounds at the adapertural end. The ribs are low and broad on the phragmocone, and weaken and efface progressively on the body chamber. The specimen differs in no significant respects from the lectotype of E. bererense.
All of the Madagascan specimens described above are interpreted as microconchs. Collignon (1969, p. 208, pl. 602, fig. 2257) described and illustrated a complete adult individual 175 mm in diameter that he referred to Eulophoceras besairiei Hourcq, 1949. The specimen (Figs 18, 19) has a maximum phragmocone height of 70 mm. The outer whorl has lost virtually all ornament. The phragmocone is oxycone, while the body chamber broadens markedly, developing a fastigiate venter, that broadens and rounds at the adapertural end. We interpret this specimen as an adult macroconch. On this basis, the single KwaZulu specimen, OUM KX12924 (Figs 13C, 14, 17C), with a maximum measurable phragmocone whorl height of 70 mm, and an estimated maximum phragmocone whorl height of 75-80 mm is also a macroconch, adult on the basis of the crowded final sutures, but lacking virtually all of the body chamber. The specimen is an internal mould; the phragmocone has the following dimensions:
At D: 109 (100) Wb: 31.7 (29.1) Wh: 67.8 (62.1) Wb:Wh: 0.47 U: 7.3 (6.7)
A stout oxycone, with a whorl breadth to height ratio of 0.47, the greatest breadth is just outside the umbilical shoulder. Ten primary ribs arise at the umbilical seam of the outer whorl, and strengthen across the umbilical shoulder, where some develop into low bullae, best developed at the adapical end of the outer whorl. Ribs arise either singly or in pairs at the umbilical shoulder. They are low, broad, straight and prorsirdiate on the inner flank, across which they strengthen progressively. The ribs flex back around mid-flank, and may bifurcate, sweeping back and concave on the outer flank, strengthening progressively before effacing, and not extending to the venter. Little but the inner flank of the adapical end of the body chamber survives (Fig. 14), but this shows a marked weakening of the ornament. A suture from some way adapical of the end of the phragmocone (Fig. 17C) has a broad, asymmetrically bifid E/A, with a narrow median incision; A is narrow and subtrifid; U2 is bifid. Both lobes and saddles have only minor, shallow incisions. The sutures crowd markedly towards the end of the phragmocone, with simplifying elements on the inner flank region.
Discussion
The suite of Madagascan specimens, and the single KwaZulu-Natal example referred to the species share a common basic style of ornament of low, falcoid weak to coarse ribs, a whorl section in which the body chamber changes from the lanceolate of the phragmocone to fastigiate on the adapical part of the body chamber to rounded at the adapertural end. These features separate bererense from natalense, as noted above. The sutures are also distinctive. Those of natalense, albeit variable, are characterized by elongate folioles, seen in extreme form in the lectotype (Figs 2, 3), but also present in others (Figs (10A-C; 11A-C). Those of bererense are, by contrast, characterized by minor incisions and, commonly, short, blunt folioles.
As noted above, the Eulophoceras jacobi Hourcq of Renz (1982, p. 111, pl. 36, figs 10, 11; pl. 37, figs 2, 3; text-fig. 86a-b) from the Coniacian of Venezuela, has quite different ornament, and is best referred to as Eulophoceras sp. juv.; the specimen described by Pattaroya & Duenas (2006) as E. jacobi also is best referred to Eulophoceras sp.
Occurrence
Hourcq (1949) referred his species to the top of the Santonian. Collignon (1969) recorded 227 Eulophoceras from Madagascar, all from his lowest Lower Campanian Anapachydiscus wittekindi and Eulophoceras jacobi Zone. All those figured came from the upper, Hourcquiella bererensis Subzone. The KwaZulu-Natal specimen comes from Campanian I-II of Kennedy & Klinger (1975).
ACKNOWLEDGEMENTS
Kennedy acknowledges the support of the staff of the Geological Collections, Oxford University Museum of Natural History, and the Department of Earth Sciences, Oxford, and the financial assistance of the Oppenheimer Fund (Oxford). Klinger acknowledges financial support from the NRF (South Africa) and logistic support from the staff of the Natural History Collections Department, Iziko, Museums of Cape Town. Dr H. Summesberger (Vienna) kindly provided the originals of Fig. 4. We also thank him and Dr Marcin Machalski (Warsaw) for their constructive comments.
REFERENCES
BAILY, W.H. 1855. Description of some Cretaceous fossils from South Africa. Quarterly Journal of the Geological Society of London 11: 454-465. [ Links ]
BASSE, E, 1954. Sur la presence du genre Coahuilites Böse 1927 dans le sud Tunisien et du genre Eulophoceras Hyatt 1903 dans le proche Orient. Bulletin de la Société Géologique de France (6)3: 865-871. [ Links ]
BUCH, L. VON. 1848. Über Ceratiten, besonders von denen, die in Kreidebildung sich finden. Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Kóniglich Preussischen Akademie der Wissenschaften zu Berlin für 1847, 214-223. [ Links ]
COLLIGNON, M. 1966. Atlas des fossiles caractéristiques de Madagascar (Ammonites). XIV, Santonien. x + 1-134. Tananarive: Service Géologique. [ Links ]
COLLIGNON, M. 1969. Atlas des fossiles caractéristiques de Madagascar (Ammonites). XV, (Campanien inférieur), xi + 1-216. Tananarive: Service Géologique. [ Links ]
COLLIGNON, M. 1983. Les faunes d'Ammonites du Santonien. In: BILOTTE, M. & COLLIGNON, M. Biostratigraphie et Paléontologie des ammonites du Sénonien Inférieur de Rennes-Les Bains-Sougraigne (Aude) [zone sous Pyrénéenne Orientale]. Documents de Laboratoire de Géologie de Lyon, H.S.6: 175-223. [ Links ]
COOPER, M.R. 1988. Lower Campanian (Cretaceous) ammonites from Angola. South African Journal of Science 91: 207-211. [ Links ]
DOUVILLÉ, H. 1912. Evolution et classification des Pulchelliidés. Bulletin de la Société Géologique de France (4)11: 285-320. [ Links ]
GERHARDT, K. 1897. Beiträge zur Kenntniss der Kreideformation in Columbien. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilage Band 11: 118-208. [ Links ]
GHARE, M.A. 1979. Lenticeratid ammonoids from South Indian Cretaceous rocks of Trichinopoly District, Tamil Nadu. Biovigyanam 5: 165-169. [ Links ]
GROSSOUVRE, A. de 1894. Recherches sur la craie supérieure, 2, Paléontologie. Les ammonites de la craie supérieure. Mémoires du Service de la Carte Géologique détaillée de la France. 1-264 (misdated 1893). [ Links ]
HOEPEN, E.C.N. VAN, 1921. Cretaceous Cephalopoda from Pondoland. Annals of the Transvaal Museum 7: 142-147. [ Links ]
HOURCQ, V. 1949. Paléontologie de Madagascar. XXVIII. Sur quelques ammonites du Sénonien. Annales de Paléontologie 35: 10 (87) - 31 (117). [ Links ]
HYATT, A. 1900. Cephalopoda. In: ZITTEL, K.A. VON 1896-1900, Textbook of Palaeontology (transl. Eastman, C.R.) pp. 502-604. London and New York: Macmillan. [ Links ]
HYATT, A. 1903. Pseudoceratites of the Cretaceous. United States Geological Survey Monograph 44: 351 p. [ Links ]
IMMEL, L. 1987. Die Kreideammoniten der nördlichen Kalkalpen. Zitteliana 15: 3-163. [ Links ]
IMMEL, H.., KLINGER, H.C. & WIEDMANN, J. 1982. Die Cephalopoden des Unteren Santon der Gosau von Brandenberg/Tirol, Osterreich. Zitteliana 8: 3-32. [ Links ]
KENNEDY, W.J. 1984. Systematic palaeontology and stratigraphic distribution of the ammonite faunas of the French Coniacian. Special Papers in Palaeontology 31: 160 pp. [ Links ]
KENNEDY, W.J. 1987. Ammonites from the type Santonian and adjacent parts of northern Aquitaine, western France. Palaeontology 30: 765-782. [ Links ]
KENNEDY, W. J., BILOTTE, M. & MELCHIOR, P. 1995. Ammonite faunas, biostratigraphy and sequence stratigraphy of the Coniacian-Santonian of the Corbières. Bulletin des Centres de Recherche Exploration et Production Elf-Aquitaine 19: 377-499. [ Links ]
KENNEDY, W.J., KAUFFMAN, E.G. & KLINGER, H.C. 1973. Upper Cretaceous invertebrate faunas from Durban, South Africa. Transactions of the Geological Society of South Africa 77: 95-111. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 1975. Cretaceous faunas from Zululand and Natal, South Africa. Introduction, stratigraphy. Bulletin of the British Museum (Natural History) Geology 25: 263-315. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 1985. Cretaceous faunas from Zululand and Natal, South Africa: the ammonite family Kossmaticeratidae Spath, 1922. Annals of the South African Museum 95: 165-231. [ Links ]
KLINGER H.C. & KENNEDY, W.J. 1980. The Mzamba Formation at its type locality, Mzamba Estuary (Pondoland, Transkei), ammonite content and paleogeographical distribution. Annals of the South African Museum 80: 207-222. [ Links ]
KNECHTEL, M.M. 1947. In: KNECHTEL, M.M., RICHARDS, E.F. & RATHBURN, V. Mesozoic fossils of the Peruvian Andes. Johns Hopkins University Studies in Geology 15: 1-150. [ Links ]
KORN, D., EBBIGHAUSEN, V., BOCKWINKEL, J. & KLUG, C. 2003. The A-mode ontogeny in prolecanitid ammonites. Palaeontology 46: 1123-1132. [ Links ]
KOSSMAT, F. 1895-1898. Untersuchungen über die Südindische Kreideformation. Beiträge zur Paläontologie Österreich-Ungarns und des Orients. 9 (1895), 97-203 (1-107); 11 (1897), 1-46 (108-153); 11(1898), 89-152 (154-217). [ Links ]
KULLMANN, J. & WIEDMANN, J. 1970. Significance of sutures in phylogeny of Ammonoidea. University of Kansas, Paleontological Contributions 42: 1-32. [ Links ]
PERVINQUIÈRE, L.. 1910. Sur quelques ammonites du Crétacé algérien. Mémoires de la Société Géologique de France. Paléontologie 17(2-3), 86 pp. [ Links ]
PATARROYO, P. & DUEÑAS, H. 2006. Eulophoceras jacobi Hourcq y palinomorphos asociados del Coniaciano Temprano, Formación Loma Gorda, Quebrada Bambucá (Aipe-Huila-Colombia, S.A. Revista Academia Colombia Ciencias 30: 503-510. [ Links ]
RENZ, O. 1982. The Cretaceous Ammonites of Venezuela. Basel: Maraven. [ Links ]
REYMENT, R.A. 1955. The Cretaceous Ammonoidea of Nigeria and the southern Cameroons. Bulletin of the Geological Survey of Nigeria 25: 112 pp. [ Links ]
SANTAMARIA ZABALA, R. 1995. Los Ammonoideos del Cenomaniense superior al Santoniense de la plataforma nord-castellana y la cuenca navarro-cántabra. Parte II Sistemática: Acanthocerataceae. Treballs del Museu de Geologia de Barcelona 4: 15-131. [ Links ]
SPATH, L.F. 1921. On Cretaceous Cephalopoda from Zululand. Annals of the South African Museum 12: 217-321. [ Links ]
SPATH, L.F. 1922. On the Senonian ammonite fauna of Pondoland. Transactions of the Royal Society of South Africa 10: 113-147. [ Links ]
SUMMESBERGER, H. 1979. Eine obersantone Ammonitenfauna aus dem Becken von Gosau (Oberõsterreich). Annalen des Natur-historischen Museums, Wien 83: 275-283. [ Links ]
SUMMESBERGER, H. 1980. Neue Ammoniten aus der Sandkalk-bank der Hochmoosschichten (Obersanton; Gosau, Austria). Annalen des naturhistorischen Museums, Wien 83: 275-383. [ Links ]
WOODS, H. 1906. The Cretaceous fauna of Pondoland. Annals of the South African Museum 4: 275-350 [ Links ]
WRIGHT, C.W. 1957. [Cretaceous Ammonoidea]. In: MOORE, R.C. (ed.) Treatise on Invertebrate Paleontology. Part L, Mollusca 4, Revised. Cephalopoda Ammonoidea. New York and Lawrence: Geological Society of America and University of Kansas Press. [ Links ]
WRIGHT, C.W. 1996. In: Treatise on Invertebrate Paleontology. Part L, Mollusca 4: Cretaceous Ammonoidea, (with contributions by J.H. Calloman (sic) and M.K. Howarth). Boulder, Colorado and Lawrence, Kansas: Geological Society of America and University of Kansas. [ Links ]
YOUNG, K. 1963. Upper Cretaceous ammonites from the Gulf Coast of the United States. University of Texas Bulletin 6304: ix + 373 pp. [ Links ]
ZABORSKI, P.M.P. 1982. Campanian and Maastrichtian sphenodiscid ammonites from southern Nigeria. Bulletin of the British Museum (Natural History) Geology 36: 303-332. [ Links ]
Received 3 May 2012
Accepted 18 July 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}