










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAfrican Natural History
On-line version ISSN 2305-7963
Print version ISSN 1816-8396
Afr. nat. history (Online) vol.8 Cape Town Jan. 2012
Revision of the crown crabs, genus Hymenosoma (Crustacea: Hymenosomatidae), of South Africa
Jessica DawsonI; Charles L. GriffithsII
IDepartment of Zoology, University of Cape Town, Private Bag, Rondebosch, Cape Town, 7701 South Africa. E-mail: jessica.dawson@uct.ac.za
IIDepartment of Zoology, University of Cape Town, Private Bag, Rondebosch, Cape Town, 7701 South Africa. E-mail: charles.griffiths@uct.ac.za
ABSTRACT
Recent genetic studies have shown that the South African crown crab, Hymenosoma orbiculare, in fact represents five distinct species, three of which are new to science and one of which needs to be resurrected. Three of these species occur in the southwestern Cape, the remaining two being found in estuaries on the South East and East coasts of South Africa. Based on genetic evidence and morphological analyses, H. geometricum Stimpson, 1858 is no longer considered a junior synonym of H. orbiculare, and is resurrected as a full species. A deeper-water species from False Bay, H. trilobatum sp. nov. as well as two East coast species, H. longicrure sp. nov. and H. projectum sp. nov., are described. The true H. orbiculare ranges from Walvis Bay in Namibia to the Gqunube Estuary near East London. H. geometricum and H. trilobatum sp. nov. are both found in deeper water and are currently known only from False Bay. H. longicrure sp. nov. is found from the Qolora Estuary to the Mngazi Estuary in the Eastern Cape and H. projectum sp. nov. along the east coast from the Mzimvubu Estuary to Lake Sibaya and Kosi Bay. The size and shape of the rostrum and ornamentation of the carapace are the main features distinguishing species. H. orbiculare has a smooth, simple rostrum and only two pairs of plain teeth on the carapace; H. geometricum has a very complex, long, trilobate rostrum and three pairs of complex, granulate teeth on the carapace; H. trilobatum sp. nov. has a less complex, extended, trilobate rostrum with smooth margins and granulate surface, with only two pairs of teeth on the carapace; H. longicrure sp. nov. and H. projectum sp. nov. both have a simple rostrum, little ornamentation on the carapace and unusual abdominal projections.
Keywords: Hymenosoma, new species, crown crab, South Africa, taxonomy
INTRODUCTION
Members of the family Hymenosomatidae, commonly known as false spider crabs or crown crabs, occur mainly in tropical and subtropical, shallow coastal waters and low-salinity habitats in the Indo-West Pacific (Lucas 1980; Chuang & Ng 1994). Hymenosomatids can occur in high densities and are common in the gut contents of fish, playing a significant role in food webs (Lucas 1980). They are among the smallest members in the infraorder Brachyura, with a maximum carapace width of only 3-26 mm (Lucas 1980). This, combined with their dull and cryptic colouration (Chuang & Ng 1994), makes them inconspicuous (Lucas 1980) and partially explains why they are so poorly known and studied.
Studies of the taxonomy of the family have been published for Australia (Lucas 1980), New Zealand (Melrose 1975), Southeast Asia (Ng & Chuang 1994) and more recently, South Africa (Edkins et al. 2007; Teske et al. 2009). The recorded number of genera and species is constantly increasing, Lucas (1980) reported 10 genera and 64 species, Edkins et al. (2007) 16 genera and 90 species and Ng et al. (2008) 17 genera and 109 described species. Until recently, it was considered that only three species of Hymenosomatidae occurred in South Africa - Hymenosoma orbiculare, Neorhynchoplax bovis (Stimpson 1858) (cf. Lucas 1980) and Elamena mathoei (Desmarest 1823) (cf. Barnard 1950) (see Ng et al. 2008, p. 109, note {2}). H. orbiculare is distinguishable by the absence of an epistome, well-defined carapace grooves, slender third maxilliped and smooth dactyls. N. bovis has a well-defined epistome and carapace grooves; narrow third maxilliped not completely covering the buccal cavity; denticles (teeth) on the dactyls and a tridentate rostrum. E. mathoei is characterized by presence of an epistome; carapace not with defined grooves, smooth; broad third maxilliped covering the buccal cavity; sub-apical denticles (teeth) on the dactyls and a broadly triangular rostrum (Barnard 1950).
On a global basis the genus Hymenosoma currently comprises just three species:
1) H. orbiculare, as currently defined, which has a distribution in southern Africa and Zanzibar and is characterized by absence of an epistome, well-defined grooves on the dorsal carapace and having the infero-external orbital angle bifed.
2) H. depressum, occurring in New Zealand and distinguishable by presence of antennal spines, epistome and narrow third maxilliped.
3) H. hodgkini, occurring in eastern Australia and distinguishable by absence of epistome, elongated eyestalks, broad third maxilliped and teeth on dactyls of the walking legs (Lucas 1980).
Melrose (1975) treated H. depressum as a separate genus, Cyclohombronia. Lucas (1980) agreed that, should these three species be divided into two groups, H. depressum would be the outlier but argued that, because these three species are much more closely related to each other than to any other hymenosomatid, they should be retained within one genus.
Hymenosoma orbiculare Desmarest, 1825, as currently defined, is found all along the coast of southern Africa (Barnard 1950). Although restricted to calmer areas where fine sands and mud accumulate (Broekhuysen 1955), it is common in fully marine habitats to depths up to 80 m (Barnard 1950), as well as in estuaries, with a small population even occurring in the freshwater (but relictual estuarine) Lake Sibaya (Allanson et al. 1966). Although H. orbiculare is currently the only species in the genus recognized from South Africa, several authors have noted morphological variants. In 1858 Stimpson described a smaller, more granulose, deeper-water form as H. geometricum (cf. Stebbing 1905), but this was subsequently treated as a junior synonym of H. orbiculare (Stebbing 1914, Barnard 1950). Barnard (1950) and Broekhuysen (1955) both mentioned a variety of differences between deep-water specimens and coastal H. orbiculare, but neither found it necessary to consider these as separate species. Barnard (1955), however, suggested that the deep-water species be fully investigated and Lucas (1980) suggested that the large range of morphological and habitat variation of H. orbiculare indicated that several species exist under that one name.
Using morphological and genetic methods to compare specimens from all along the South African coast Edkins et al. (2007) identified five monophyletic clusters within the species, each with distinct distribution ranges. With the support of morphological data, they proposed that South African H. orbiculare in fact represent five distinct species. These are:
1) The true H. orbiculare, characterized by larger size and absence of characteristic ornamentations of other morphotypes and found in estuaries, lagoons and shallow, sheltered marine habitats all along the west and south coasts of South Africa.
2) Two False Bay deep-water species; the one previously described as H. geometricum, and distinguished by a long rostrum, a raised setaceous gastric region on the carapace and post-branchial projections, and the second a new, undescribed species.
3) Two new species occurring in estuaries along the southeast and east coasts respectively and distinguished by their small size, relatively large eyes, long walking legs, projections on the abdomen and small anterior spikes on coxae of the legs. Southeastern specimens can be differentiated from east coast specimens primarily by darker colour and longer second walking legs (Edkins et al. 2007).
The aims of this paper are thus to formalize the taxonomic status and to illustrate each of these species. H. geometricum is officially resurrected and the new False Bay deep-water species and two new east coast estuarine species are named and described.
METHODS AND MATERIALS
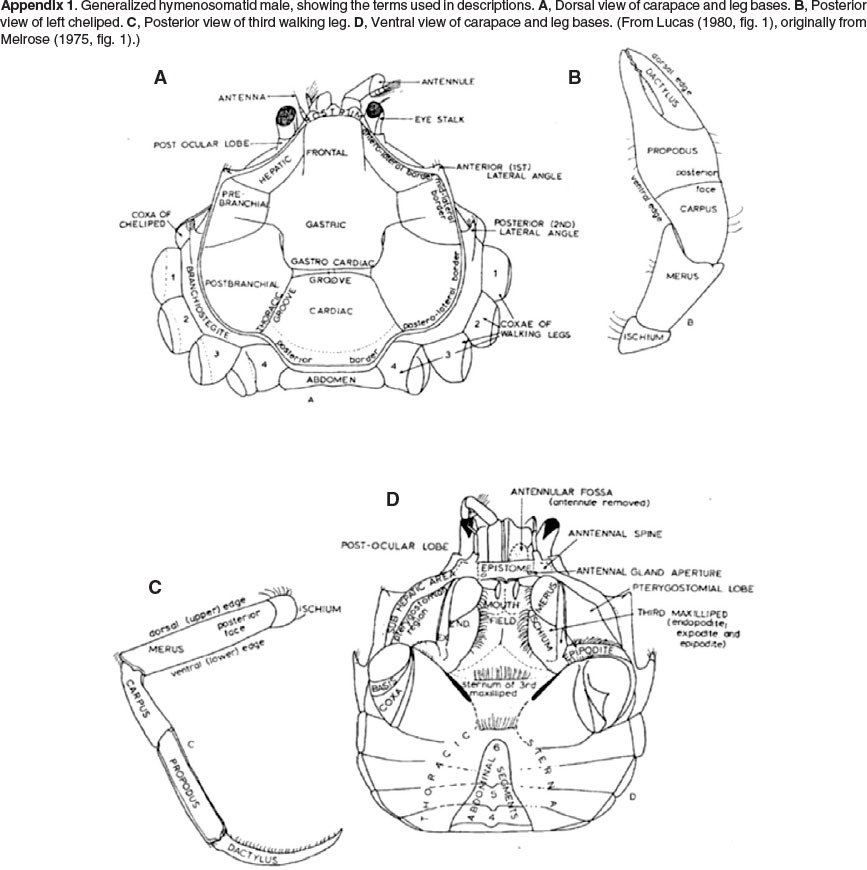
This study is based on the same specimens collected for the genetic study by Edkins et al. (2007). Deeper water specimens from False Bay were collected using two methods; by hand while on SCUBA (for samples at 5 m depth), or with a small dredge towed behind a speedboat (for samples at 20 m depth). The remaining estuarine and coastal material was collected by wading into the water and sieving bottom sediments with a D-frame hand-net. Drawings were made using a Wild Camera Lucida (type 300) mounted on a Wild binocular microscope (M5 48418) and whole specimens photographed with a Nikon D50 digital SLR camera fitted with a Nikon 105 mm macro lens. A generalized hymenosomatid male, showing the terms used in descriptions in the text, is illustrated in Appendix 1.
All specimens of Hymenosoma in the collections of the South African Iziko Museum were also re-examined, separated into the five newly-recognized species, relabelled and used here to provide additional distributional data.
TAXONOMIC SECTION
Family HYMENOSOMATIDAE MacLeay, 1838
Description
(based on Barnard (1950) and Lucas (1980))
Carapace thin, flat, not heavily calcified, lacking hooked setae; dorsal carapace surface often with well-defined gastro-cardiac groove and neighbouring grooves, usually with horizontal rostrum. No orbits, or very incomplete orbits, eyes exposed and little retractable; antennular sockets shallow and ill-defined; antennae joined to epistome (when present); ischium of third maxillipeds well-developed, palp articulating from antero-lateral corner of merus; chelipeds not long or particularly mobile. Male and female abdomens not more than five segments, excluding telson; male genital openings on last thoracic sternite (hidden beneath abdomen), female genital openings on sternite of second (first walking) legs; female pleopods on abdominal segments 2-5.
Genus Hymenosoma Desmarest, 1825
Description
(based on Barnard (1950) and Lucas (1980))
Carapace sub-circular or elongate oval, with anterior and posterior lateral angles not evident; carapace flat, with well-defined or fine dorsal grooves, triangular rostrum, usually with groove separating rostrum from dorsal carapace, concave laterally, usually shorter than eyes. Epistome absent or very short and narrow. Plain pterygostonial region. Third maxilliped usually with narrow third and fourth joint and not meeting centrally, underlying appendages partially visible, ischium subequal to or shorter than merus, palp large and conspicuous, epipodites conspicuous. Chelipeds heftier than walking legs, particularly in male specimens, not greatly overstated; walking legs slender or moderately stout, two to three times longer than carapace width, dactyli with row of small teeth, or without spines or denticles on inner margin, usually with dense long setae. Female abdomen with no fused segments, triangular or trapezoidal plate formed by medial portions of segments 1-3, visible dorsally -segments 1, 2 and part of 3, weak convex laterally making broad, shallow, abdominal cavity (brood cavity); male abdomen with no fused segments, short and narrow and with sutures of all segments distinct.
The following three species are recognized from the Western Cape region of South Africa.
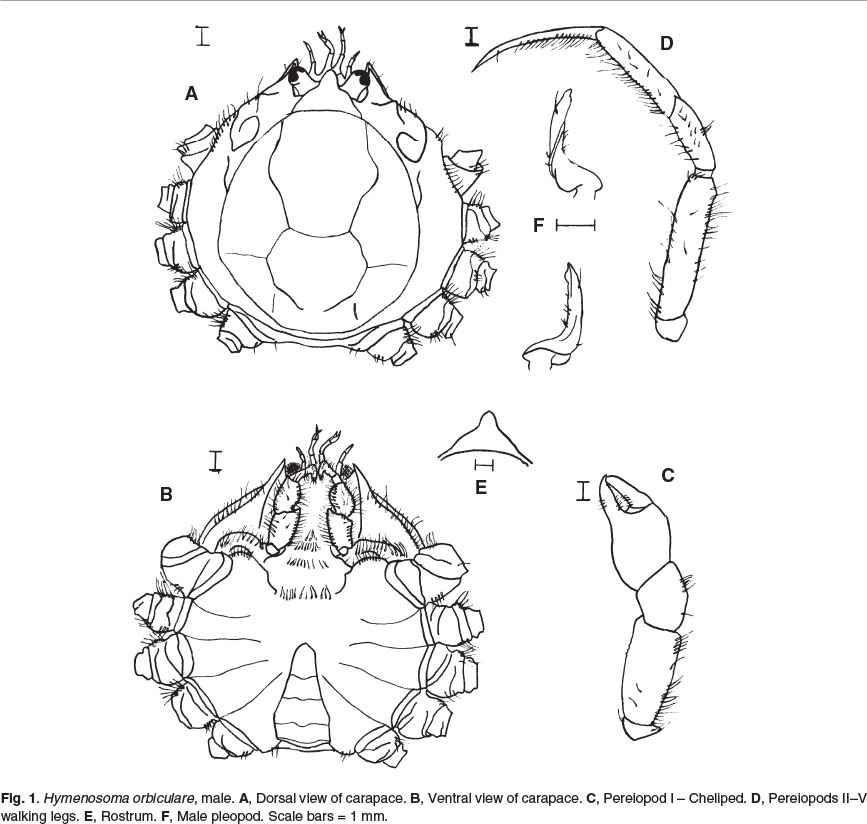
Hymenosoma orbiculare Desmarest, 1825 (Crown crab) Figs 1A-F, 4A
1825 Hymenosoma orbiculare Desmarest, Considerations Generales sur la Classe des Crustaces, p. 163, pl. 26. fig. 1, 1a-e.
1914 Hymenosoma orbiculare ( partim), Stebbing, Transactions of the Royal Society of Edinburgh, p. 270, pl. 25, fig. A.
1950 Hymenosoma orbiculare (partim), Barnard, Annals of the South African Museum 38 p. 68.
1955 Hymenosoma orbiculare, Broekhuysen, Annals of the South African Museum 41, p. 313-343. figs 1-13.
1955 Hymenosoma orbiculare (non), Barnard, Annals of the South African Museum 43 p. 19. fig. 5.
1980 Hymenosoma orbiculare, Lucas, Records of the Australian Museum 33 pp. 166-169.
2007 Hymenosoma orbiculare (partim), Edkins et al., Crustaceana 80(6), p. 667-683.
Material examined
South Africa: SAM A 45731, Kromme River Estuary, July 2005, 7 ♀,13♂; SAM A 45732, Gonubie River Estuary, 2004, 1 ♀9♂; SAM A 45733, Kowie River Estuary, 2004, 2♀, 38; SAM A 45734, Milnerton Lagoon, 2005, 29,♂; SAM A 45735, Knysna Lagoon, March 2008, 1♂; SAM A 45736, Langebaan Lagoon, March 2005, 49,1♂; SAM A1425, Salt River Mouth, 3 ind; SAM A 46795, Walvis Lagoon Namibia, 50 ind; SAM A 45070, Berg River Estuary, 70 ind; SAM A 3137, Port Elizabeth, 2♂; SAM A 42077, St. Helena Bay, 2♂, 1♀ SAM A 13378, Kenton-on-Sea, 1♂; SAM A 44007, Berg River Estuary, 7 ind; SAM A 39589, Lambert's Bay, 1♂,1♀ SAM A 47808, Berg River Estuary, 30 ind; SAM A 459, East London, 18; SAM A 11022, Woodstock Power Station, 1♂ SAM A 6882, Kleinmond Lagoon, 9 ind; SAM A 15361, Port Alfred, 8 ind; SAM A 12142, Luderitz, 1♀ SAM A 12629, Luderitz, 4 ind; SAM A 12771, Luderitz, 2♂; Knysna, 3♂,1 9; Sandwich Harbour Namibia, 7 ind; Leisure Isle Knysna, 1 ♂; Sandwich Harbour Namibia, 3♂,1♀ Sandwich Harbour Namibia, 8 ind (these last samples without museum accession numbers).
Description
(based on Stebbing (1914), Barnard (1950) and Edkins et al. (2007))
Carapace surface sub-circular, dorsal surface flat, well-defined gastro-cardiac groove and neighbouring grooves, with single, broad tooth on anterior sub-hepatic region and low rounded prominence on posterior hepatic region; infero-external orbital angle produced in a bifid tooth, the upper tooth reaching to, or slightly beyond, apex of rostrum (Figs 1A & 4A); rostrum triangular, with groove separating rostrum from dorsal carapace, concave above, with slightly up-turned, rounded apex with point at very tip, shorter than eyes, with lateral sections protruding over eyestalks, no serrations (Fig. 1E). Epistome absent. Plain pterygostonial region. Maxilliped 3 with many setae, not meeting medially, with underlying appendages partially visible, third and fourth joints twice as long as broad, ischium subequal to merus, palp large and conspicuous (Fig. 1B). Elongated eyestalks, very prominent post-ocular lobes. Antennules slightly longer than, or subequal to, antennae. Chelipeds slightly stouter than walking legs in females, almost twice as stout as walking legs in males, with fine teeth on dactyli, no teeth on merus, moderately setose (Fig. 1C); walking legs long and slender, almost twice carapace width, heavily fringed on inner margins of fourth to seventh joints, dactyli longer than propodus, with no teeth (Fig. 1D). Abdomen smooth, with no setae, female abdomen circular to ovate, male abdomen triangular, expands to maximum width mid- way between segments 3-4 and then narrows to segment 5, segment 5 narrows to base of telson which is semi-ovate (Fig. 1B); no abdominal projections; male first pleopod slightly setose, distinct proximal curve, stout at base, tapering to thin, twisted shaft, slightly curved tip (Fig. 1F). Maximum carapace width 28 mm; carapace dark: reddish, yellowish, brownish or greenish, sometimes carapace mottled and legs banded.
Remarks
Hymenosoma orbiculare is distinguishable from the other two Western Cape (False Bay) species described here by its larger size and lack of the characteristic ornamentations present in the other species. It has a simple, smooth, rostrum with rounded apex. The rostrum is only slightly trilobate, with the medial lobe reaching the apex of the eyes.
Occurrence
The most common Hymenosoma species along the entire west and south coasts of South Africa, found in estuaries and lagoons and extending up to 5 m depth in shallow-water marine environments. Range extends from Walvis Bay in Namibia to the Gqunube Estuary, near East London in South Africa (Edkins et al. 2007). Also reported from Zanzibar by Barnard (1950), citing Lenz (1905), although in the light of this study it seems highly likely that the Zanzibar material in fact represents either one of the east coast species described here, or yet another as yet undescribed species.
Hymenosoma geometricum Stimpson, 1858 (reinstated)
1858 Hymenosoma geometricum Stimpson, Proceedings of the Academy of Natural Sciences of Philadelphia 10, p. 108.
1905 Hymenosoma geometricum, Stebbing, Marine Investigations of South Africa, South African Crustacea Part 3, p. 50.
1914 Hymenosoma orbiculare (partim). Stebbing, Transactions of the Royal Society of Edinburgh, p. 270, pl. 25, fig. A.
1950 Hymenosoma orbiculare,(partim). Barnard, Annals of the South African Museum 38 p. 68. 1955 Hymenosoma orbiculare, Barnard, Annals of the South African Museum, 43 p. 19. fig. 5.
2007 Hymenosoma orbiculare (partim), Edkins et al., Crustaceana, 80(6), pp. 667-683.
Material examined
SAM A 45737, False Bay, South Africa, 2005, 39,28, 20 m; SAM A 12399, False Bay, 19; SAM A 12400, False Bay, 19.
Description
(based on SAM A 12398, mature male, False Bay, South Africa, 20 m)
Carapace surface circular, with slight elongation at posterior margin, dorsal carapace surface flat, well-defined gastro-cardiac and neighbouring grooves; gastric region raised into setaceous median boss; carapace surface slightly setose with small, conical granulations; elongate, granular tooth protruding from sub-hepatic area, adjacent to orbital angle, conical, granular tooth on hepatic margin and pre-branchial margin of carapace, bifid tooth at outer orbital angle visible ventrally, reaching below apex of eyestalk (Figs 2A & 4B). Rostrum distinctly trilobate, with groove separating rostrum from dorsal carapace, medial lobe narrow with rounded apex, curving slightly upwards and extending significantly beyond end of eyestalks, more than twice length of callosities at the base and serrated along margins, with conical granulations on surface; two basal callosities conical with granulations, extending almost to apex of eyestalks (Fig. 2E). Epistome absent. Pterygostonial region slightly setose. Maxilliped 3 narrow, not meeting medially with underlying appendages partially visible, ischium subequal to merus, highly setose, palp large and conspicuous (Fig. 2B). Elongated eyestalks, prominent post-ocular lobes. Antennules longer than antennae, extend slightly beyond elongate rostrum. Chelipeds slightly stouter than walking legs in females, almost twice as stout as walking legs in males, no fine teeth on dactyli, single large tooth on outer, distal margin of merus, with several smaller conical teeth along length of cheliped (Fig. 2C); walking legs slender, highly setose, 1.5 times longer than carapace width, dactyli shorter than propodus, with no teeth (Fig. 2D). Abdomen with highly setose margin, female abdomen circular, male abdomen narrow, club-shaped, maximum width at base of segment t4 and then narrows to segment t5 and telson which is semi-ovate (Fig. 2G); no abdominal projections; male first pleopod setose, distinct proximal curve straightening to tip, narrow at base, tapering to thin, pointed tip, twisted shaft (Fig. 2F). Maximum carapace width small, <10 mm; pale, slightly transparent carapace.
Remarks
Hymenosoma geometricum has the most elaborate ornamentation of the described species. It has a characteristically very complex rostrum, with serrulate margins and granulate surface. The rostrum is distinctly trilobate, with the medial lobe reaching well beyond the apex of the eyes. The carapace is covered with small conical granulations and is slightly setose, with a raised, setose median boss on the gastric region. Based on both genetic work by Edkins et al. (2007) and this morphological analysis, H. geometricum of Stimpson 1858 should thus be resurrected as a valid species and no longer treated as a junior synonym of H. orbiculare.
Occurrence
Presently known only from deeper marine habitats (>20 m) in False Bay, South Africa. Edkins et al. (2007) found that H. geometricum was either picked off the large ascidian Pyura stolonifera, or found in dredges that contained P. stolonifera, thus suggesting that P. stolonifera may represent a habitat for this species.
Hymenosoma trilobatum sp. nov.
2007 Hymenosoma deep water species, C, Edkins et al., Crustaceana 80(6), pp. 667-683.
2009 Hymenosoma species 1, Teske et al., Molecular Phylogenetics and Evolution 53, pp. 23-33.
Etymology
This species is named for its triple lobed, triangular rostrum.
Type specimen
SAM A 45738 mature male, False Bay, South Africa, 20 mm.
Material examined
SAM A 45739, False Bay, South Africa, 20 March 2005, 69, 20 m; SAM A 45749, False Bay, South Africa, 19,28; SAM A 448, False Bay, South Africa, 209,128; SAM A 11994 (D430), Saldanha Bay, South Africa, 39.
Description
(based on the mature male in Fig. 3 and Edkins et al. (2007))
Carapace surface sub-circular, dorsal carapace surface flat with well-defined gastro-cardiac groove, but poorly-defined neighbouring grooves; posterior margin slightly concave; strong granulation over entire dorsal carapace surface, with prominent beaded and granulated lateral rim. Carapace, coxopodites and posterior margin all setose. Orbital angle produced in a bifid tooth, each of equal length, sharply pointed apex, protruding slightly beyond eyes; small, granular tooth protruding from sub-hepatic area, adjacent to orbital angle, pustulate or mammilate tooth of hepatic carapace margin (Figs 3A & 4C). Rostrum trilobate, with groove separating rostrum from dorsal carapace, medial lobe triangular with rounded apex and smooth margin, extending to tip of eyestalk, almost twice length of callosities at base. Two basal callosities rounded with circular granulations, extend- ing only to base of eyestalks (Fig. 3E). Epistome absent. Plain pterygostonial region with setose margin. Maxilliped 3 very narrow, highly setose, not meeting medially, with underlying appendages partially visible, palp reduced, heavily fringed margins of ischium and merus (Fig. 3B). Shortened eyestalks, prominent post-ocular lobes. Antennules almost twice length of antennae, protruding well beyond eyestalks. Chelipeds stouter than walking legs, male chelipeds more robust, covered in circular granulations, very fine teeth on dactyli, merus without tooth, heavily fringed on both fore and hind margins (Fig. 3C); walking legs slender, setose, 1.5 times longer than carapace width, heavily fringed on inner margins, dactyli subequal to propodus, with no teeth (Fig. 3D). Abdomen surface highly setose, female abdomen circular to ovate, male abdomen triangular, expands to maximum width at base of segment 2 and then tapers to segment 5 and telson which is slightly pointed (Fig. 3B); males have two flattened, highly setose, posterior abdominal projections, abdominal projections absent on females; male first pleopod slightly setose, broad at base, tapering to thin, slightly rounded tip, twisted shaft with slight proximal curve (Fig. 3F). Maximum carapace width small, <12 mm; pale, slightly creamy, transparent carapace.
Remarks
Hymenosoma trilobatum is highly granulate over its entire body surface, but the granulations are more circular than the conical granulations seen on H. geometricum. The rostrum is of moderate complexity, with smooth margins but a granulate surface. Like H. geometricum the rostrum is distinctly trilobate but the medial lobe is not as long, reaching only slightly past the apex of the eye stalks. Based on both genetic work by Edkins et al. (2007) and this morphological analysis, this form needs to be accepted as a separate species.
Occurrence
Currently known only from deeper marine habitats (20 m) in False Bay, South Africa.
The following two species are recognized from the Eastern Cape and KwaZulu-Natal regions of South Africa.
Hymenosoma longicrure sp. nov.
2007 Hymenosoma species D, Edkins et al., Crustaceana 80(6), pp. 667-683.
2009 Hymenosoma species 2, Teske et al., Molecular Phylogenetics and Evolution 53, pp. 23-33.
Etymology
This species is named for the extended length of its walking legs
Type specimen
SAM A 45740, mature male, Mngazi Estuary, Eastern Cape, South Africa, 2005.
Material examined SAM A 45741, Mbanyana Estuary, September 2005, 10♂, 5♀; SAM A 45742, Mngazi Estuary, 2005, 8♂,12♀; SAM A 45368, Nxaxo, 3 April 1993, 1♂, 1♀.
Description
(based on the mature male in Fig. 5 and Edkins et al. (2007))
Carapace surface sub-circular, dorsal surface flat, well-defined cardiac, gastro-cardiac groove and neighbouring grooves, with single, small bulbous tooth on carapace margin of hepatic region and raised rounded prominence over central hepatic region; external orbital angle produced in a bifid tooth, the upper tooth reaching to or slightly below eye (Figs 5A & 7A); rostrum triangular, with groove separating rostrum from dorsal carapace, indented along central axis -concave surface, with slightly up-turned, rounded apex, shorter than eyestalks, no serrations (Fig. 5E). Epistome absent. Plain pterygostonial region. Maxilliped 3 narrow with many setae, not meeting medially, with underlying appendages partially visible, with third and fourth joint twice as long as broad, ischium subequal to merus, palp large and conspicuous (Fig. 5B). Eyes large relative to carapace, elongated eyestalks, very prominent post-ocular lobes. Antennules almost twice length of antennae, extending well beyond eyes. Female chelipeds narrower, but no stouter than walking legs, male chilipeds equal or slightly stouter than walking legs with very swollen, bulbous propodus, fine teeth on dactyli and merus, moderately setose. In larger males, small rounded projection found on proximal, inner margin of cheliped dactylus (Fig. 5C); walking legs long and slender, more than twice carapace width, fringed on inner margins of fourth to seventh joints, dactyli subequal to propodus, small anterior spike on coxae of walking legs (Fig. 5D). Abdomen smooth, with setae, female abdomen circular with fringed margin, male abdomen triangular, expands to maximum width midway between segments 2-3 and then narrows to segment 5, segment 5 narrows to base of small ovate telson (Fig. 5B); abdominal projections present, females with slight point on third abdominal somite, males with two, short, flattened posterior projections, only slightly setose; male first pleopod minimal setae, hooked - broad, but straight at base with strong distal curve to pointed tip, slightly twisted shaft (Fig. 5F). Maximum carapace width small, <10 mm; carapace dark: yellowish, brown sometimes with mottled carapace.
Remarks
Hymenosom longicrure has the longest legs relative to carapace size of these five species, with the second walking legs always more than 1.5 times length of the first. Males have a highly differentiated first pleopod when compared to the remaining four species, being broad at the base with a strong distal, not proximal, curve. Males also show a bulbous swelling of the propodus of the cheliped. Based on both genetic work by Edkins et al. (2007) and this morphological analysis, this form needs to be accepted as a separate species.
Occurrence
Currently known range from Qolora to Mngazi Estuaries on the southeast coast of South Africa.
Hymenosoma projectum sp. nov.
2007 Hymenosoma species E (partim), Edkins et al., Crustaceana 80(6), pp. 667-683.
2009 Hymenosoma species 3, Teske et al., Molecular Phylogenetics and Evolution 53, pp. 23-33.
Etymology
This species is named for the distinct abdominal projections seen on males and immature females.
Type specimen
SAM A 45743, mature male, Kosi Bay third Lake, South Africa, August 2005.
Material examined
SAM A 45744, Mzingazi Estuary, June 2005, 1 ♂,5♀; SAM A 45745, Kosi Bay Lakes, August 2005, 5♂; SAM A 45746, Kosi Bay third Lake, August 2005, 16♂,7♀; SAM A 45747, Mpenjati Estuary, September 2005, 5♂,3♀; SAM A 45748, Mbotyi Estuary, October 2005, 1♀.
Description
(based on the mature male in Fig. 6 and Edkins et al. (2007))
Carapace surface sub-circular/ovate, dorsal carapace surface flat with well-defined gastro-cardiac groove, but poorly-defined neighbouring grooves; raised rounded prominence over central hepatic region; external orbital angle produced in a bifid tooth, upper tooth reaching slightly beyond eye (Figs 6A & 7B); rostrum triangular, with groove separating rostrum from dorsal carapace, slightly indented along central axis - concave surface, with slightly up-turned, rounded apex, shorter than eyes, no serrations (Fig. 6E). Epistome absent. Plain pterygostonial region with fringed margin. Maxilliped 3 with many setae, not meeting medially, with underlying appendages partially visible, third and fourth joints twice as long as broad, ischium subequal to merus, palp large and conspicuous (Fig. 6B). Large eyes relative to carapace size, eyestalks elongated, prominent post-ocular lobes. Antennules longer than antennae. Female chelipeds, narrower, or no stouter, than walking legs, male chelipeds equal or slightly stouter than walking legs, with swollen, bulbous propodus, moderately setose. In larger males, small rounded projection found on proximal, inner margin of cheliped dactylus (Fig. 6C); walking legs long and slender, fringed on inner margins of fourth to sixth joints, dactyli subequal to propodus, small anterior spike on coxae of walking legs (Fig. 6D). Abdomen smooth, slightly setose, female abdomen circular, male abdomen triangular, expands to maximum width midway between segments 3-4 and then narrows to segment 5, segment 5 narrows to base of small semi-ovate telson (Fig. 6B); abdominal projections present, females with slight point on third abdominal somite, males with two, extended, flattened posterior projections, only slightly setose; male first pleopod moderately setose, broad at base, slight proximal curve to pointed tip, slightly twisted shaft (Fig. 6F). Maximum carapace width small, <10 mm; carapace light: yellowing orange, to clear and translucent.
Remarks
Hymenosoma projectum is distinguishable by its very light, translucent colour and the presence of abdominal projections. Males show long posterior, flattened abdominal projections also present in immature females.
Occurrence
Currently known to inhabit South African east coast north of Mzimvubu Estuary to Lake Sibaya and Kosi Bay. Edkins et al. (2007) suggest the range probably extends further north, possibly even to Zanzibar (Barnard, 1950) and Madagascar (Ng & Chuang, 1996). Small individuals noted at Inhaca Island, Mozambique (Macnae & Kalk, 1958; Dornelas et al., 2003), probably also belong to this species.
DISCUSSION
Edkins et al. (2007) used genetic and morphological data to show that the South African crown crab, Hymenosoma orbiculare, in fact represents five distinct species. Three of these can be found in the Western Cape Province (one extending into Namibia) and the remaining two along the east coast of South Africa.
Aside from the DNA evidence (Edkins et al. 2007) the separation of the rostrum from the carapace by a groove and/or a ridge, and the absence of an epistome, clearly identify all these species as belonging to the genus Hymenosoma.
Barnard (1950) reported that specimens of H. orbiculare ranged not only around the entire coastline of southern Africa, but from the sublittoral down to 80 m, into estuaries, and in one case into a coastal freshwater lake (Allenson et al. 1966). Lucas (1980) called this 'a surprising distribution for a euryhaline species.' Thus it is not unexpected to discover that this distribution is not that of a single species, but rather of five different Hymenosoma species. The true coastal H. orbiculare is found in estuaries and lagoons along the west and south coasts; H. geometricum Stimpson, 1858, resurrected in this paper, and H. trilobatum sp. nov. occur in deeper-water in False Bay; while estuaries on the southeast and east coasts harbour H. longicrure sp. nov. and H. projectum, respectively.
In this paper we describe and illustrate all five of the South African Hymenosoma species. The species fall into two groups, based on geographical range, thus in the account below we will concentrate on variations between species with overlapping or adjacent distribution ranges.
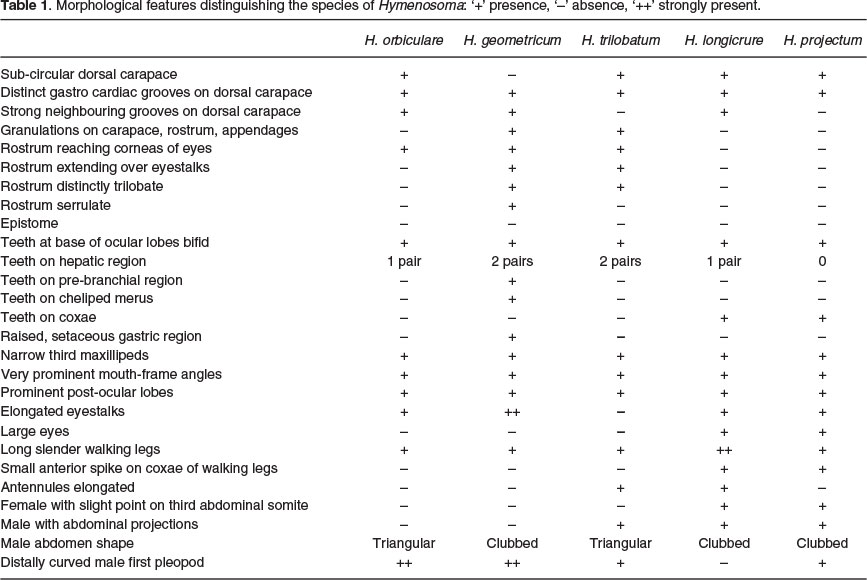
Although at first glance the three west and south coast species may appear similar, each has distinct characteristics that differentiate it from the others. Summarizing the data from Table 1, H. orbiculare and H. geometricum differ in 10 features, H. orbiculare and H. trilobatum in eight features and H. geometricum and H. trilobatum in ten features. H. trilobatum appears to be an intermediate between the other two species, it shares traits with each and a number of its features are not as extreme as H. geometricum, or as simple as those of H. orbiculare.
The simplest feature distinguishing the three southwestern species is the rostrum. H. orbiculare has a simple rostrum, with smooth margin and a flat surface, not extending past the apex of the eyestalk; H. geometricum has a complex, triangular, trilobate rostrum, the medial lobe, with serrulate margins, extending well beyond the eyestalks, and the two basal lobes and rostrum surface covered with conical granulations. H. trilobatum has a less complex, trilobate rostrum, with smooth margins, the medial lobe extends just beyond the apex of the eyestalks and the rostrum surface is covered in circular granulations.
The ornamentation on the carapace can also be used to distinguish these species. H. orbiculare has an obtuse tooth and low, rounded prominence on the hepatic regions; H. geometricum has a complex, elongate, granular tooth protruding from sub-hepatic area; a conical, granular tooth on the hepatic and pre-branchial regions; the former with a much smaller, inferior anterior tooth. Also unique to this species is the raised, setose, median boss protruding from the centre of the gastric region and the presence of one large and several smaller teeth on the merus of the cheliped. H. trilobatum has a small granular, tooth of the sub-hepatic region and a pustulate or mammilate tooth on the pre-branchial region and a continuation of the carapace granulations onto the cheliped. Uniquely, in terms of west and south coast species, H. trilobatum has two highly setose, flattened projections on the abdominal region of the male carapace.
The deeper-water species, H. geometricum and H. trilobatum also mature at a smaller size (Broekhuysen 1955, Lucas 1980), are more setose and more granulose, and are generally paler in colour than H. orbiculare.
The two east coast species, H. longicrure sp. nov. and H. projectum sp. nov., are very similar to one another and most similar to H. orbiculare amongst the west and south coast species. The geographical boundary between the southeast H. longicrure and southern H. orbiculare corresponds to the zoogeographic boundary separating the subtropical East Coast and the Warm Temperate South Coast provinces (Emanuel et al. 1992; Edkins et al. 2007).
Summarizing the data from Table 1 H. orbiculare differs from both H. longicrure and H. projectum in nine separate features, while H. longicrure and H. projectum only differ in four features. The east coast species are smaller, <10 mm carapace width, and paler in colour than H. orbiculare, have relatively larger eyes and are less setose than any of the west and south coast species. Although simple and similar in shape to H. orbiculare the lateral sections of the rostrum of H. longicrure and H. projectum do not protrude over the eyestalks and the apex never reaches past the eyes. East coast species have small anterior spikes on the coxae of the walking legs, and abdominal projections are present, females have a slight point on the third abdominal somite, males having two, slightly setose, flattened posterior projections, a trait only the males of H. trilobatum share.
Hymenosoma longicrure and H. projectum are not easily distinguishable, the simplest distinguishing feature being colour. H. longicrure is darker and often a mottled brown colour, while H. projectum is much paler and a light orange to translucent colour. Anatomically H. longicrure has longer walking legs, with the second walking leg always more than 1.5 times as long as the first (Edkins et al. 2007), more defined carapace grooves, with a small tooth on the hepatic carapace margin and longer antennules; H. projectum has poorly defined carapace grooves but more prominent, longer abdominal projections, distinctly visible in both dorsal and ventral view of carapace.
The greatest distinction between the east coast species is only seen in the males; H. longicrure has a uniquely shaped first pleopod, easily distinguishable from the other four species, it is hook-shaped with a broad base and a strong distal curve that is only slightly twisted. H. projectum has a broad base with a proximal curve that never fully straightens. The first pleopods of east coast species are similar in shape, only differing in the degree of distal curve and basal width, these species have a twisted shaft, distally curved, straightening and tapering to a pointed tip.
Teske et al. (2009) showed that in southern Africa Hymenosoma range expansion occurred from the Tropical Western Indian Ocean southwards, with species moving south during warmer climate periods, then adapting once cooler climatic conditions returned, resulting in speciation. Given the presence of three distinct species of Hymenosoma in the South Western Cape of South Africa and two others along the eastern coast, it is highly unlikely that the specimens reported from Zanzibar (2500 km further north) by Barnard (1950), based on an original record by Lenz (1905), represent any of these species, certainly not the temperate H. orbiculare. We have been unable to locate these original specimens, but collection and examination of new material from Zanzibar should be carried out to determine the true identity of that population.
ACKNOWLEDGEMENTS
We thank Max Edkins, Isabelle Papadopoulos and the Iziko South African Museum for use of specimens from their collections. Financial support for this project was provided by a grant to C.L. Griffiths from the National Research Foundation SEAChange Programme. Jessica Dawson was supported by University of Cape Town Honours Merit Council and National Research Foundation Scholarships.
APPENDIX 1
REFERENCES
ALLANSON, B.R., HILL B.J., BOLTT, R.E. & SCHULTZ, V. 1966. An estuarine fauna in a freshwater lake in South Africa. Nature, London 209: 532-533. [ Links ]
BARNARD, K.H. 1950. Descriptive catalogue of South African decapod Crustacea. Annals of the South African Museum 38: 1-836. [ Links ]
BARNARD, K.H. 1955. Additions to the fauna-list of South Africa Crustacea and Pycnogonida. Annals of the South African Museum 43: 1-107. [ Links ]
BROEKHUYSEN, G.J. 1955. The breeding and growth of Hymenosoma orbiculare, Desm. (Crustacea, Brachyura). Annals of the South African Museum 41: 313-343. [ Links ]
CHUANG, C.T.N. & NG, P.K.L. 1994. The ecology and biology of Southeast Asian false spider crabs (Crustacea: Brachyura: Hymenosomatidae). Hydrobiologia 285: 85-92. [ Links ]
DESMAREST, A.J. 1825. Considérations Générales sur la des Crustacés et Description des Espèces de ces Animaux, qui vivant dans la Mer, sur les Côtes, on dans les Eaux Douces de la France. Paris & Strasbourg. 446 pp. [ Links ]
DORNELAS, M., PAULA, J. & MACIA, A. 2003. The larval development of Hymenosoma orbiculare Desmarest, 1825. (Crustacea: Brachyura: Hymenosomatidae). Journal of Natural History, London 37: 2579-2597. [ Links ]
EDKINS, M.T., TESKE, P.R., PAPADOPOULOS, I. & GRIFFITHS, C.L. 2007. Morphological and genetic analyses suggest that Southern African crown crabs, Hymenosoma orbiculare, represents five distinct species. Crustaceana 80: 667-683. [ Links ]
EMANUEL, B.P., BUSTAMANTE, R.H., BRANCH, G.M., EEKHOUT, S. & ODENDAAL, F.J. 1992. Zoogeographic and functional approach to the selection of marine reserves on the west coast of South Africa. South African Journal of Marine Science 12: 341-354. [ Links ]
LENZ, H. 1905. Ostafrikanische Dekapoden und Stomatopoden, gesammelt von Herrn Prof. Dr. A. Voeltzkow. In: VOELTZKOW, A., Wissenschaftliche Ergebnisse der Reisen in Madagaskar und Ostafrika in den Jahren 1889-1895. Vol. 3. Abhandlungen der Senckenbergischen naturforschenden Gesellschaft 27(4): 341-392, pls 47-48. [ Links ]
LUCAS, J.S. 1980. Spider crabs of the family Hymenosomatidae (Crustacea: Brachyura) with particular reference to Australian species: systematics and biology. Records of the Australian Museum 33: 148-247. [ Links ]
MACLEAY, W.S.1838. Invertebrates. Vol. 2 In: SMITH, A. Illustrations of the Zoology of South Africa. Smith, Elder: London [ Links ]
MACNAE, W. & KALK, 1958. A Natural History of Inhaca Island, Mozambique. Witwatersrand University Press: Johannesburg. 163 pp. [ Links ]
MELROSE, M.J. 1975. The marine fauna of New Zealand: family Hymenosomatidae (Crustacea, Decapoda, Brachyura). New Zealand Oceanographic Institute Memoir 34: 1-123. [ Links ]
NG, P.K.L., GUINOT, D. & DAVIE, P.J.F. 2008. Systema Brachyurorum: Part I. An annotated checklist of the extant Brachyuran crabs of the world. The Raffles Bulletin of Zoology 17: 1-286. [ Links ]
STEBBING, T.R.R. 1905. South African Crustacea. Part 3. Marine Investigations in South Africa 4: 21-123. [ Links ]
STEBBING, T.R.R. 1914. Stalk-eyed Crustacea Malacostraca of the Scottish National Antarctic Expedition. Transactions of the Royal Society of Edinburgh 50: 253-307. [ Links ]
STIMPSON, W. 1858. Prodromus descriptionis animalium evertebratorum, quae in Expeditione as Oceanum Pacificum Septentrionalem, a Republica Federate missa, Cadwaladaro Ringgold et Johanne Rodgers Ducibus, observatit et descripsit. V. Crustacea Ocypodoidea. Proceedings of the Academy of Natural Sciences of Philadelphia 10: p. 9-110. [ Links ]
TESKE, P.R., McLAY, C.L., SANDOVAL-CASTILLO, J., PAPADOPOULOS, I., NEWMAN, B.K., GRIFFITHS, C.L., McQUAID, C.D., BARKER, N.P., BORGONIE, G. & BEHEREGARAY, L.B. 2009. Tri-locus sequence data reject a 'Gondwanan origin hypothesis' for the African/South Pacific crab genus Hymenosoma. Molecular Phylogenetics and Evolution 53: pp. 23-33 [ Links ]
Received 16 February 2012
Accepted 6 August 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}