










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAfrican Natural History
On-line version ISSN 2305-7963
Print version ISSN 1816-8396
Afr. nat. history (Online) vol.8 Cape Town Jan. 2012
Cretaceous faunas from Zululand and Natal, South Africa. The ammonite genera Mojsisovicsia Steinmann, 1881, Dipoloceroides Breistroffer, 1947, and Falloticeras Parona & Bonarelli, 1897
William James KennedyI; Herbert Christian KlingerII
IOxford University Museum of Natural History, Parks Road, Oxford OX1 3PW, U.K., and Department of Earth Sciences, South Parks Road, Oxford OX1 3AN, U.K. E-mail: jim.kennedy@oum.ox.ac.uk
IINatural History Collections Department, Iziko South African Museum, P.O. Box 61, Cape Town, 8000 South Africa. Email: hklinger@iziko.org.za
ABSTRACT
Mojsisovicsia ventanillensis (Gabb, 1877), Dipoloceroides delaruei (d'Orbigny, 1841), D. semicornutum (Spath, 1931), D. cornutum (Pictet, 1847) and Falloticeras proteus (d'Orbigny, 1842) are described from the Middle Albian Mzinene Formation of northern KwaZulu-Natal. The interpretation of micromorph species associated with these taxa is reviewed, and duerfeldi of Steinmann, 1881, is interpreted as the microconch of ventanillensis as a working hypothesis. The status of proteus as either microconch or micromorph is unresolved.
Keywords: ammonites, Albian, micromorphs, KwaZulu-Natal, South Africa.
INTRODUCTION
The Middle Albian part of the Mzinene Formation of northern KwaZulu-Natal has yielded representatives of one species referred to the genus Mojsisovicsia Steinmann, 1881, three species referred to Dipoloceroides Breistroffer, 1947, and one referred to Falloticeras Parona & Bonarelli, 1897. In describing these taxa, and establishing their generic assignation, a wider problem arises, that of the relationship between 'normal-sized' and diminutive species of Mojsisovicsiinae. These have been regarded, in the case of the latter, as either the microconchs (m) of larger, macro-conch (M) individuals, forming a dimorphic pair, or as dwarf, paedomorphic offshoots of 'normal-sized' species. The key taxa in this area were reviewed by Breistroffer (1947), Kennedy & Cooper (1977), Gebhard (1983), and Wright (1996).
The historical background to the problem was carefully reviewed by Gebhard, and is updated here.
1841 d'Orbigny described and illustrated his Ammonites delaruei from the condensed Middle Albian of southeast France, based on a series of individuals up to 53 mm diameter (p. 296, p. 87, figs 6-8) (Fig. 4A-E).
1842 d'Orbigny described, but did not figure his Ammonites proteus from the condensed Middle Albian of southeast France (p. 624) (Fig. 8F,G)
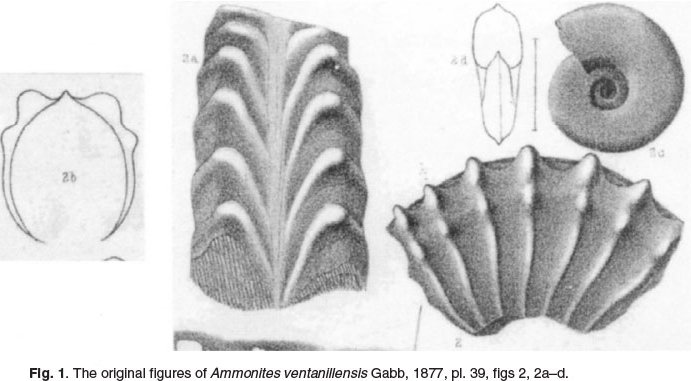
1877 Gabb described and illustrated his Ammonites ventanillensis from the Middle Albian of Parietambo, Peru (p. 273, pl. 39, figs 2, 2a-d; Fig. 1 herein).
1881 Steinmann described and illustrated his new genus Mojsisovicsia and the type species Mojsisovicsia duerfeldi from Parietambo, Peru (p. 144, pl. 6, fig. 2).
1882 Steinmann concluded that his species duerfeldi was a junior synonym of ventanillensis.
1897 Parona & Bonarelli introduced the genus Falloticeras, with Ammonites proteus d'Orbigny, 1842, as type species, and provided the first illustrations of the species (p. 89, pl. 12, fig. 1).
1921 Douglas accepted Mojsisovicsia duerfeldi as a junior synonym of Ammonites ventanillensis (p. 270, pl. 15, fig. 8; pl. 17, fig. 4).
1942 Spath referred to both Mojsisovicsia duerfeldi and M. ventanillesis, but equivocated on their separateness (p. 709).
1947 Breistroffer introduced the subgenus Dipoloceras (Dipoloceroides) with Ammonites delaruei as type species (p. 90 (74)), and noted: 'Enfin, il y a des liaisons certaines entre D.(D.) delaruei d'Orb. Sp. et Falloticeras Proteus d'ORB. sp. in Par. Et Bon., de même qu'entre D.(D). ventanillense GABB sp. et Mojsisovicsia Dürfeldi STEINM. sp. Ce sous-genre et ses deux dérivés nains à cloisons simplifiées précèdent les Dipoloceras typiques.' (p. 90 (75).
1957 Wright regarded duerfeldi of Steinmann as a junior synonym of ventanillensis of Gabb, and Dipoloceroides Breistroffer, 1947, as a junior synonym of Mojsisoviczia (sic) Steinmann, 1881.
1977 Kennedy & Cooper revised Falloticeras, and regard it as possibly a neotenous offshoot of Mojsisovicsia.
1979 Gebhard regarded Falloticeras as a junior synonym of Mojsisovicsia (p. 98).
1983 Gebhard revised and illustrated the type material of Mojsisovicsia duerfeldi and concluded that it was not a junior synonym of ventanillensis. Falloticeras was interpreted as a junior synonym of Mojsisovicsia. Dipoloceroides Breistroffer, 1947, was regarded as a valid taxon, and treated as a subgenus of Dipoloceras Hyatt, 1900. It was further suggested that Mojsisovicsia (= Falloticeras) and Dipoloceras (Dipoloceroides) are a dimorphic pair.
1996 Wright (p. 137) regarded Mojsisovizsia (sic) duerfeldi as a junior synonym of Ammonites ventanillensis, the former the microconch, the latter the macroconch. He also considered Dipoloceroides to be a junior synonym of Mojsisoviczia (sic). Falloticeras was retained as a separate genus.
We have re-examined the type material of the type species of Falloticeras and Dipoloceroides as well as topotype material of Mojsisovicsia duerfeldi and Ammonites ventanillensis, and reached the following conclusions.
1. Mojsisovicsia duerfeldi is characterized by an ornament of sinuous growth lines and constrictions, the aperture preceded by a constriction, the mouth border flared. Adults are up to 27 mm in diameter. It co-occurs with Ammonites ventanillensis, which has an initial smooth stage, then develops a keel, then ribs and tubercles, as clearly described by Douglas (1921, p. 271). Common early development is a necessary feature for ventanillensis and duerfeldi to be dimorphs, and this view is adopted here as a working hypothesis, following Wright (1996, p. 137). Recognition of dimorphism in larger ventanillensis at the ornamented stage, or in the diminutive duerfeldi would confirm that they are not a dimorphic pair, and that duerfeldi is a genuinely diminutive taxon, and a paedomorphic off shoot of ventanillensis.
2. Ammonites delaruei differs from Ammonites ventanillensis in having ribs from a much earlier stage, and thus lacks the distinctive unribbed but carinate stage of the latter. Dipoloceroides and Mojsisovicsia (of which these are the type species) can be differentiated on this criterion, but whether this is a difference of species, sub-genera, or genera, is debatable. Recognition of dimorphism in Dipoloceroides delaruei would help resolve this uncertainty.
3. Falloticeras differs from microconch Mojsisovicsia in having a ribbed early growth stage (Fig. 8), whereas microconch Mojsisovicsia lacks ribs throughout ontogeny. They can be separated on this basis.
4. The early developmental stages of Dipoloceroides semicornutum (Spath, 1931) and Falloticeras proteus are similar, suggesting a relationship between the two. F. proteus may be a paedomorphic offshoot of D. semicornutum, or the microconch, as suggested by Gebhard. The former of these alternatives can only be confirmed if dimorphism is demonstrated in either or both species.
These issues can only be fully addressed by a revision of the material from the classic Albian of Escragnolles in France (the type locality of Falloticeras and Dipoloceroides) and Parietambo in Peru. Accordingly, we adopt a conservative position with respect to the present material from northern KwaZulu-Natal, noting that the main record has what are interpreted as macroconchs of Mojsisovicsia ventanillensis co-occuring with the microconch/micromorph Falloticeras proteus.
REPOSITORIES OF SPECIMENS
BMNH: The Natural History Museum, London.
MNHP: The Muséum National d'Histoire Naturelle, Paris.
OUM: Geological Collections, Oxford University Museum of Natural History, Oxford.
SAM: Natural History Collections Department, Iziko South African Museum, Cape Town.
FIELD LOCALITIES
Details of field localities are given by Kennedy & Klinger (1975); further descriptions of these localities are deposited in the Geological Collections, Oxford University Museum of Natural History, The South African Museum, Cape Town, and the Department of Palaeontology, The Natural History Museum, London.
CONVENTIONS
Dimensions are given in millimetres: D = diameter; Wb = whorl breadth; Wh = whorl height; U = umbilicus; c = costal dimension; ic = intercostal dimension. Figures in brackets are dimensions as a percentage of the diameter. The suture terminology is that of Korn et al. (2003): E = external lobe; A = adventive lobe (= lateral lobe, L, of Kullmann & Wiedmann, 1970); U = umbilical lobe; I = internal lobe.
SYSTEMATIC PALAEONTOLOGY
Suborder AMMONITINA Hyatt, 1889
Superfamily ACANTHOCERATOIDEA de Grossouvre, 1894
Family BRANCOCERATIDAE Spath, 1934
Subfamily MOJSISOVICSIINAE Hyatt, 1900
Genus Mojsisovicsia Steinmann, 1881 Type species
Mojsisovicsia duerfeldi Steinmann, 1881, p. 144, pl. 6, fig. 2, by original designation = Ammonites ventanillensis Gabb, 1877, p. 273, pl. 39, figs 2, 2a-d.
Diagnosis
Strongly dimorphic, macroconchs up to 100 mm diameter, microconchs 25-35 mm in diameter. Earliest growth stage of macroconch smooth but for growth lines, followed by the development of a keeled stage with growth lines only, then the abrupt appearance of flexuous primary ribs, flared on the outer flank, with a well developed inner ventrolateral bulla on some ribs, and transverse to oblique outer ventrolateral clavi on all ribs. Inner ventrolateral bullae efface on the adapertural part of the adult body chamber. Microconchs with rounded whorl section, ornamented by growth lines and occasional constrictions; adult aperture preceded by broad shallow constriction with flared aperture, produced into a blunt ventral lappet.
Occurrence
Middle Albian, Peru, northern KwaZulu-Natal.
Mojsisovicsia ventanillensis (Gabb, 1877) Figs 1-3
1877 Ammonites ventanillensis Gabb, p. 273, pl. 39, figs 2, 2a-d.
1881 Mojsisovicsia Dürfeldi Steinmann, p. 144, pl. 6, fig. 2.
1903 Mojsisovicsia durfeldi Steinmann; Hyatt, p. 25, pl. 1, figs 1-3.
1908 Schloenbachia juv. Ventanillensis Gabb (?Mojsisovicsia Dürfeldi Steinmann); Lisson, p. 15, pl. 15, figs 1-3.
1908 Schloenbachia ventanillensis Gabb; Lisson, p. 16a-c, pl 16a, figs 16, 1-4.
1911 ?Mojsisovicsia Dürfeldi Steinmann; Lisson, p. 168.
1921 Schloenbachia ventanillensis (Gabb); Douglas, p. 270, pl. 15, fig. 8; pl. 17, fig. 4.
1922 Mojsisovicsia dürfeldi (Steinmann); Spath, p. 97.
1924 Mortoniceras Ventanillensis Gabb juv.; Lisson, pp. 71, 146.
1938 Mojsisovicsia Dürfeldi Stein.; Roman, p. 37, figs 352, 352a, 352b.
1942 Mojsisovicsia dürfeldi Steinmann; Spath, p.708.
1942 Mojsisovicsia ventanillensis (Gabb); Spath, p. 708.
1947 Mojsisovicsia Dürfeldi Steinmann; Breistroffer, p. 90 (75).
1957 Mojsisoviczia (sic) ventanillensis (Gabb); Wright, p.L 404, text-fig. 522, 4a,b.
1977 Mojsisovicsia aff. ventanillensis (Gabb); Kennedy & Cooper, p. 800.
1977 Mojsisovicsia sp. juv. Kennedy & Cooper, pl. 105, figs 4, 12.
1977 Mojsisovicsia ventanillensis (Gabb); Kennedy & Cooper, pl. 105, figs 11, 13, 18.
1977 Falloticeras sp. Kennedy & Cooper, pl. 105, fig. 14.
1982 Mojsisovicsia cf. ventanillensis (Gabb); Cooper, text-fig. 20.
1983 Mojsisovicsia dürfeldi Steinmannn; Gebhard, p. 114, text-figs 1a, 3.3, 3.4.
1996 Mojsisoviczia [sic] ventanillensis (Gabb); Wright, p. 137, text-fig. 105, 1a-g.
?2002 Mojsisovicsia cf. ventanillensis (Gabb,1869); Robert, p. 180, pl. 32, figs 1-3.
Types
The lectotype, here designated is the original of Gabb, 1877, p. 273, pl. 39, fig. 2, 2a, 2b, from the' limestones of the Cerro del Ventanillo, between Pachanchaca and Jauja, Height 5000 metres'. The paralectotype is the original of Gabb, 1877 p. 273, fig. 2a,b, from the 'coal mine of Paraiatambo, 5 leagues from Morococho.' The original illustrations are reproduced here as Fig. 1. A second, unfigured paralectotype is from the 'neighbourhood of Fingo, Prov. of Huari, Dept. of Huaraz, from a schist containing coal'. The specimens were collected by Dr Antonio Raimondi (1826-1890), who considered them to be of Jurassic age. We have not seen these specimens, which we presume survive in the Raimondi Museum in Lima.
Material
SAM-PCZ022404 (ex H207/1/1), SAM-PCZ022405-409 (ex H201/1/4-8), BMNH C78865, C78867, all from the Middle Albian fauna of bed 1 of the Mzinene Formation at locality 51 of Kennedy & Klinger (1975, p. 288, text-fig. 4), stream bed and bank exposures in the Mzinene River north of Hluhluwe, northern KwaZulu-Natal, coordinates 27°53'43"S, 32°19'22"E.
Dimensions
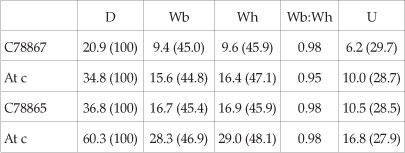
All the available specimens are macroconchs. BMNH C78872 (Fig. 3D-F), a wholly septate internal mould lacking the innermost whorls, preserves parts of two successive growth stages. Coiling is moderately evolute, with 45.2% of the previous whorl covered at the largest preserved diameter. The umbilicus comprises 27.8% of the diameter, and is shallow, with a low, convex umbilical wall that inclines outwards and merges with the broadly rounded umbilical shoulder. There is no flank ornament to a diameter of 27.5 mm, and the flanks are very feebly convex, subparallel, the ventrolateral shoulder relatively narrowly rounded, the venter broad, and very feebly convex. There is a well-developed siphonal ridge that strengthens progressively as size increases. Ribs appear abruptly at a diameter of 27.5 mm, and there are nine in the 90° sector between 27.5 and 34.8 mm diameter. They arise at the umbilical seam or on the umbilical wall, and strengthen across the umbilical shoulder without developing a bulla. They are narrow, straight and prorsiradiate on the flanks, across which they strengthen progressively. They sweep forwards over the ventrolateral shoulders and venter, and terminate against a progressively strengthening siphonal keel. The adapical two ribs bear a weak outer ventrolateral clavus; these strengthen progressively on the succeeding ribs. Inner ventrolateral bullae are present on two ribs, with three intervening ribs that lack such bullae. SAM-PCZ022404 (ex H207/1/1) (Fig. 3G) shows an identical ontogenetic change from smooth to ribbed stages.
BMNH C78865 (Fig. 3A-C, J-L) is a near-complete adult individual broken into three pieces. The smooth stage described in the previous specimen extends to an estimated diameter of 21 mm, at which size feeble, irregularly spaced flank ribs appear (Fig. 3A-C). Regular ornament appears at a diameter of 25 mm. There are 14 ribs on the adapertural half whorl at a diameter of 36.8 mm. They arise at the umbilical seam, or on the umbilical wall, and are narrower than the interspaces, straight and prorsiradiate on the flanks, across which they strengthen progressively, sweeping forwards and strengthening further on the ventrolateral shoulder and venter, where they decline, forming an obtuse chevron with a progressively strengthening keel at the apex. Every third or fourth rib bears a strong inner ventrolateral bulla, and all ribs bear an oblique outer ventrolateral clavus, strong on the ribs with an inner ventrolateral bulla and weak on those without. The second fragment (Fig. 3K,L) extends to the end of the phragmocone at an estimated diameter of 70 mm. The adapertural half of the outer whorl bears 13 primary ribs. These are prorsiradiate, narrow, high and strong, convex on the umbilical wall, shoulder and inner flank, and feebly concave on the ventrolateral shoulders. Strong to weak umbilical bullae are present on the adapical 3 or 4 ribs. The remaining ribs lack any trace of a bulla. All ribs bear strong oblique outer ventrolateral clavi, from which the ribs project forwards and decline, forming a chevron with the very strong, high siphonal keel at the apex. A 120° sector of body chamber is present, extending to a maximum preserved diameter of 98 mm (Fig. 3J). Parts of seven ribs are preserved. They are very narrow, and widely spaced. They arise on the umbilical wall and shoulder and are initially weak, strengthening progressively across the flanks, where they are markedly prorsiradiate and flexuous, concave on the umbilical wall, convex across the inner and middle flanks, and concave on the outermost flank and ventrolateral shoulder, where they strengthen into strong, near-transverse outer ventrolateral bullae. A broad smooth zone separates the bullae from the very high, strong siphonal keel.
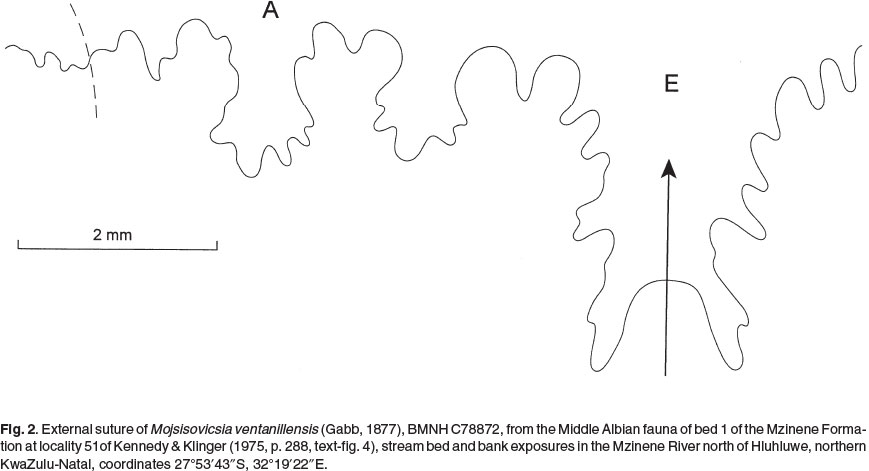
The suture (Fig. 2) is moderately incised, with a very broad, asymmetrically bifid E/A with a small median element, and small U2.
Discussion
Gabb's original figures (Fig. 1) illustrate two individuals that differ in no significant respects from the present specimens: the juvenile, smooth but for the prominent siphonal keel, and the adult fragment with comparable flexuous ribs, inner and outer ventrolateral tubercles. The costal whorl breadth to height ratio, measured from his figure, is 0.87, and if accurate suggests a slightly more compressed individual, but not significantly so. Lisson (1908) provides photographic illustrations of specimens that match well with the present material.
Douglas (1921, p. 270, pl. 15, fig. 8; pl. 17, fig. 4) described and figured material from what he regarded as the same stratigraphic unit as that described by Gabb and Lisson, the associated faunas indicating a lower Middle Albian horizon. His material matches well with the present specimens
Occurrence
Middle Albian of Saco, Peru, and northern KwaZulu-Natal.
Genus Dipoloceroides Breistroffer, 1947
Type species
Ammonites delaruei d'Orbigny, 1841, p. 296, pl. 87, figs 6-8, by the original designation of Breistroffer, 1947, p. 90 (74).
Diagnosis
Moderately evolute, compressed to depressed, with persistent siphonal keel; inner whorls with ribs on outer flank and ventrolateral shoulders, thereafter extending to the umbilical seam, some or all ribs flared on outer flank with a lateral bulla and outer ventrolateral tubercle, the latter either transverse or forming part of a ventral chevron. In forms with differentiated ribbing, the minor ribs lack a lateral flared bulla. Lateral bullae lost on adapertural part of adult body chamber.
Occurrence
Middle Albian, southern England, France, Switzerland, Poland, Pakistan, and northern KwaZulu-Natal.
Dipoloceroides delaruei (d'Orbigny, 1841) Fig. 4
1841 Ammonites Delaruei d'Orbigny, p. 296, pl. 87, figs 6-8.
1850 Ammonites Delaruei d'Orbigny; d'Orbigny, p. 124.
1897 Schloenbachia delaruei d'Orbigny, 1841; Parona & Bonarelli, p. 88, pl. 2, figs 9, 10.
1920 Oxytropidoceras delaruei (d'Orbigny); Stieler, p. 350.
1930 Mojsisovicsia aff. delaruei (d'Orbigny); Spath, p. 61, pl. 9, figs 13, 16.
1930 Mortoniceras delaruei d'Orb.; Passendorfer p. 657.
1931 Dipoloceras delaruei (d'Orbigny); Spath, p. 355, text-fig. 116 (copy of d'Orbigny).
1931 Dipoloceras delaruei compressa Spath, p. 355, text-fig. 115a; table on p. 355.
1947 Dipoloceras (Dipoloceroides) Delaruei (d'Orbigny); Breistroffer, p. 90 (74).
1963 Mojsisovicsia delaruei (d'Orbigny); Milbourne, table 1.
1971 Dipoloceras delaruei (d'Orbigny); Owen, table 3.
1979 Dipoloceras (Mojsisovicsia) delaruei (d'Orbigny); Gebhard, p. 99, pl. 7, figs 4, 5; text-figs 65, 66.
1983 Dipoloceras (Dipoloceroides) delaruei (d'Orbigny); Gebhard, p. 112.
1990 Dipoloceras (Dipoloceroides) delaruei (d'Orbigny); Marcinowski & Wiedmann, p. 84, pl. 8, fig. 6.
2006 Mojsisovicsia delaruei (d'Orbigny, 1841); Kennedy & Juignet in Gauthier, p. 109, pl. 41, figs 6, 7.
Type
The neotype, designated by Kennedy & Juignet in Gauthier, 2006, p. 109, is MNHP-B46122, ex d'Orbigny Collection 5671-1, illustrated here as Fig. 4C-E, from the condensed Albian of Escragnolles, Alpes-Maritimes, France.
Material
SAM-PCZ022410 (ex EM 175), presumably from the Pongola River, near localities Z16 to 20 of Haughton (1936, p. 290-291, fig. 3).
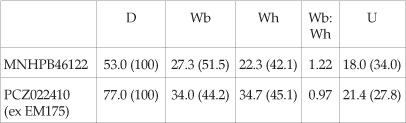
Dimensions
Description
The neotype of Dipoloceroides delaruei (Fig. 4C-E) is a phosphatized individual 53 mm in diameter, retaining part of the body chamber. Coiling is moderately evolute, the umbilicus comprising 34% of the diameter, deep, with a broadly rounded umbilical wall and shoulder. The whorl section is depressed, oval in intercostal section and polygonal in costal section, with the greatest breadth at mid-flank. There are 29-30 ribs per whorl (not 'par demi-tour' as in Kennedy & Juignet in Gauthier, 2006, p. 109), all primaries that arise at the umbilical seam, strengthening and sweeping back across the umbilical wall and shoulder. They are high and sharp on the flanks, narrower than the interspaces, prorsiradiate and straight on the umbilical shoulder, then flexing back and feebly convex on the inner flank and feebly concave on the outer flank. On the adapical parts of the outer whorl, some ribs are strengthened into a mid-lateral flare and sharp bulla, with up to three non-bullate ribs between. All ribs terminate in coarse, blunt transverse ventral clavi, separated by a smooth zone from a high, sharp siphonal keel.
PCZ022410 (exEM175) (Fig. 4F,G) is a beautifully preserved specimen that retains traces of the original aragonitic shell material. It has suffered non-lethal damage in life, and shows pathological modification of the ornament beyond a diameter of 70 mm. On the normal part of the shell, coiling is relatively evolute, with an umbilicus of moderate depth that comprises 27% of the diameter. The umbilical wall is feebly convex, the umbilical shoulder broadly rounded. The whorls are as broad as high, and trapezoidal in costal section, the flanks convergent, the venter flattened, with a strong, high, entire siphonal keel. At the smallest diameter visible there are 27 ribs per whorl, and 27-28 at 78 mm diameter, beyond which the pathological ornament appears. The ribs are prorsiradiate, strong, sharp, high, and crowded. They arise at the umbilical seam, sweep forwards across the umbilical wall, strengthen markedly and are feebly convex on the umbilical shoulder. They are straight and prorsiradiate on the inner flank, where they are produced into a high, narrow incipiently bullate flare. They flex backwards from the flare, broaden, decline in elevation, and terminate in a strong, blunt ventrolateral tubercle. On the venter, these tubercles give rise to a low, blunt, coarse rib, transverse on the outer part of the venter, but projected forwards and declining in strength, ultimately sweeping forwards as mere growth striae on the siphonal keel. Beyond 77 mm diameter the ribs change markedly, becoming convex, crowded, and narrow, without a flared section on the inner flank. At the largest preserved diameter, the keel is replaced by an irregular mass of shell material, indicating substantial non-lethal damage to the mantle in this region.
Discussion
The present specimen is much larger than the neotype and topotype material before us, one of the largest of which is the basis of var. compressa of Spath, BMNH C37612a, 65 mm in diameter, with ornament very close to that of the present specimen at the same diameter.
Dipoloceroides delaruei differs from Dipoloceras semicornutum, described below (Fig. 5) in that the latter has a very depressed whorl section in middle and later growth, far fewer ribs - 14 to 15 per whorl, the stronger with massive flared ventrolateral horns that give rise to a wedge-shaped ventral rib. The weaker ribs, two between successive stronger ribs on the penultimate whorl and adapical part of the outer whorl, one beyond this, bear a weak outer ventrolateral tubercle only.
Mojsisovicsia ventanillensis (Gabb, 1877) (p. 273, pl. 39, figs 2, 2a-d) (Figs 1-3) is discussed fully above. It has an early developmental stage that extends to a diameter of up to 27.5 mm where the shell lacks ornament other than growth lines and a strong siphonal keel, thereafter the ribs are weaker on the inner flank, more widely spaced and more markedly flexuous, the tubercles are an inner ventrolateral rather than lateral flared rib/bulla on some ribs and an oblique to near-transverse outer ventrolateral clavus on all ribs.
Occurrence
The geographic range of the species extends from southern England to the Pas-de-Calais, Aube, and Alpes-Maritimes in France, the Samana Range in the Kohat District of the North-West Frontier Province, Pakistan, and northern KwaZulu-Natal. The species is well dated as Lower Middle Albian (Hoplites spathi Subzone of the Hoplites dentatus Zone) in southern England (Owen, 1971, table on p. 155).
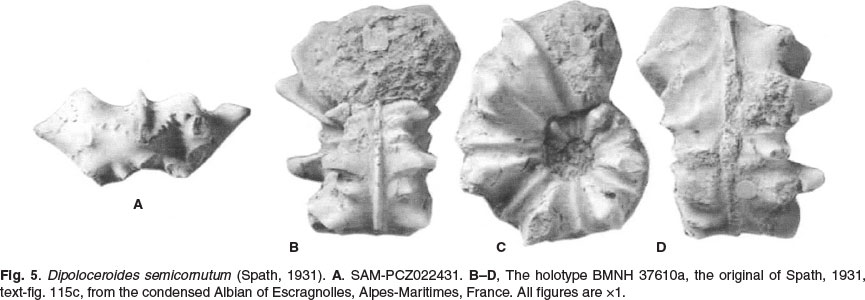
Dipoloceroides semicornutum (Spath, 1931) Fig. 5
1931 Dipoloceras semicornutum Spath, p. 349, text-fig. 115c.
1947 Dipoloceras (Dipoloceroides) semicornutum Spath; Breistroffer, p. 90 (74).
1977 Mojsisovicsia sp. juv. Kennedy & Cooper, pl. 105, figs 1, 2; text-fig. 2a.
1979 Dipoloceras (Mojsisovicsia) semicornutum Spath; Gebhard, p. 100, pl. 7, fig. 6; text-figs 67, 68.
1983 Dipoloceras (Dipoloceroides) semicornutum Spath; Gebhard, p. 116, text-figs 1d, 2. 5a,b, 3.
Type
The holotype is BMNH 3760a, the original of Spath, 1931, text-fig. 115c, from the condensed Albian of Escragnolles, Alpes-Maritimes, France (ex Astier Collection). It was referred to by Gebhardt (1983, p. 117) as C12493. It is re-illustrated here as Fig. 5B-D.
Material
SAM-PCZ022411 (ex H201/1) from Bed 1, at locality 51 of Kennedy & Klinger (1975, p. 288, text-fig. 4), stream bed and bank exposures in the Mzinene River north of Hluhluwe, northern KwaZulu-Natal, coordinates 27°53'43"S, 32°19'22"E.
Dimensions
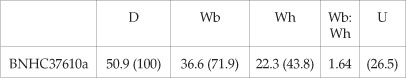
Description
The holotype was previously known only from a line drawing of the venter. It is a phosphatised internal mould with a maximum preserved diameter of 50.9 mm. The adapertural 180° sector of the outer whorl is body chamber. Coiling is evolute, the umbilical seam notched to accommodate the inner ventrolateral horns of the previous whorl. The umbilicus comprises 26.5% of the diameter and is deep, with a high convex wall. The intercostal whorl section is depressed oval; the costal section is very depressed, with the greatest breadth at the prominent inner ventrolateral horns. There are 14-15 ribs on the right flank of the outer whorl. Very strong, narrow primary ribs arise at the umbilical seam, and are straight, strong, high, and prorsiradiate on the flanks. They develop into a massive flare on the ventrolateral shoulder that is produced into a large inner ventrolateral horn and a much smaller outer ventrolateral tubercle, which gives rise to a wedge-shaped ventral rib that declines and effaces before reaching the coarse, prominent siphonal keel. Two weak primary ribs, each bearing a weak outer ventrolateral tubercle, separate the massive flared primaries at the adapical end of the outer whorl, to be replaced by a single weak intercalated rib as size increases. These weak intercalated ribs decline and efface at the adapertural end of the outer whorl. Sixteen ribs are preserved on the left flank of the outer whorl. The ornament corresponds to that on the right flank on the adapical 120° sector of the outer whorl; beyond this, the major ribs lack the inner ventrolateral horn (possibly but not necessarily a pathological condition, the result of damage to the shell in life); it only reappears on the penultimate rib, where symmetry is restored.
SAM-PCZ022411 (Fig. 5A) is a cameral fragment only, but it shows the utterly distinctive whorl section of Dipoloceroides semicornutum. The maximum preserved whorl height is 16 mm The intercostal whorl section is depressed reniform, with intercostal whorl breadth to height ratio of 1.6, and a strong siphonal keel. The costal whorl section is even more depressed, with a whorl breadth to height ratio 2.3, the rib enormously flared on the ventrolateral shoulder and expanded into an inner ventrolateral horn, with a much smaller outer ventrolateral tubercle, exactly as in the holotype.
Discussion
The massively expanded, flared ventrolateral rib and inner ventrolateral horn and very feeble minor ribs immediately distinguish the species from Dipoloceroides delaruei (compare Figs 4C-E and 5B-D). It differs from Mojsisovicsia ventanillensis in the same respects, and in being ribbed at a size where juveniles of the latter are not. There are close similarities to Dipoloceroides cornutum (Pictet, 1847) (p. 93, pl. 8, fig. 6), a cast ofthe holotype ofwhich is illustrated here as Fig. 6C-E, I), and described below. The major ribs of cornutum also develop into a massive ventrolateral flare.
But this is directed adapically, with parallel edges, and broad, evenly rounded termination, and there is no outer ventrolateral tubercle. The weaker ribs between the flared ribs have thickened ends on the venter, but no clearly differentiated tubercle.
Occurrence
Condensed Middle Albian of southeastern France, Middle Albian of northern KwaZulu-Natal.
Dipoloceroides cornutum (Pictet, 1847)
1847 Ammonites cornutus Pictet, p. 93, pl. 8, fig 6.
1850 Ammonites cornutus Pictet; d'Orbigny, p. 124.
1860 Ammonites cornutus Pictet & Campiche, p. 310.
1875a Ammonites cornutus Pictet: Neumayr, p. 21.
1875b Ammonites cornutus Pictet; Neumayr, p. 888.
1920 Inflaticeras cornutum (Pictet); Stieler, p. 399.
1921 Dipoloceras cornutum (Pictet); Spath, p. 278.
1921 Dipoloceras cornutum (Pictet); Douglas, p. 270.
1922 Inflaticeras cornutum (Pictet); Stieler, p. 309, text-figs 2, 3, 9.
1923 Dipoloceras cornutum (Pictet); Spath, p. 75.
1926 Dipoloceras cornutum (Pictet); Spath, p. 421.
1931 Dipoloceras cornutum (Pictet); Spath, p. 358 (pars), pl. 35, figs 1-4; text-figs 117a-c, 118d,e,f only; non var. corbulata, non. var. multispinosa (with additional synonymy).
1941 Dipoloceras cornutum Pictet sp.; Van Hoepen, p. 58, text-fig. 3.
1947 D. (D.) cornutum (Pictet); Breistroffer, p. 90 (74).
Type
The lectotype, designated by Spath, 1931, p. 360, is the original of Pictet, 1847, pl. 8, fig. 6, no. 18904 in the collections ofthe Muséum d'Histoire Naturelle, Geneva, from the condensed Albian ofla Perte du Rhone, Ain, France. A cast (kindly supplied by Dr Christian Meister) is illustrated here as Fig. 6C-E, I.
Material
SAM-PCZ19170 (ex D2355), the original of Van Hoepen, 1941, p. 58, text-fig. 3, from the Middle Albian Mzinene Formation, according to Van Hoepen's catalogue entry on the banks of the Munywana, below the causeway on the Somkele-Mkuze road; this seems to be in the general region of Kennedy & Klinger's (1975) localities 68-70.
Dimensions
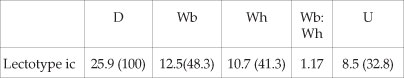
Description
The lectotype is an internal mould 25.9 mm in maximum preserved diameter, the adapical 90° is body chamber. Coiling is moderately evolute, the umbilicus comprising 32.8% of the diameter, deep, with a feebly convex, outward-inclined umbilical wall, and broadly rounded umbilical shoulder. The intecostal whorl section is depressed oval, with a whorl breadth to height ratio of 1.17. There is a strong continuous siphonal keel. The inner to mid-flank region of the penultimate whorl is smooth; delicate prorsiradiate ribs appear on the outer flank. On the adapical 90° sector of the outer whorl ten delicate primary ribs arise on the umbilical wall, sweep forwards, strengthen, and are convex across the umbilical shoulder, straight and prorsiradiate on the flanks, strengthening further on the outermost flanks and ventrolateral shoulder, where they are feebly convex. The ribs sweep forwards on the venter, forming an obtuse chevron, but efface before they reach the keel. There are three major ribs on the adapical sector of the outer whorl. They are narrow and prorsiradiate on the inner to middle flank, strengthening into a massive spatulate ventrolateral horn. This is directed upwards and backwards, with adapertural and adapical edges parallel, and a broadly and evenly rounded termination. The horns are slightly offset on either side of the venter, the best-preserved pair, at the adapertural end of the specimen, are unequally developed (Fig. 6I), the one smaller than the other. A smooth zone separates the horns from the siphonal keel. The major ribs are separated by two, three and, adaperturally, a single minor rib. These are narrower than the interspaces, very feebly convex across the umbilical shoulder, prorsiradiate on the inner flank, sweeping forwards and concave on the ventrolateral shoulder, to form an obtuse chevron on the venter. They efface before reaching the siphonal keel; some are incipiently bullate.
SAM-PCZ 19170 (ex D2335) (Fig. 6A,B) is 18.5 mm in diameter. Coiling is moderately evolute, the small, deep umbilicus comprising 24.3% of the diameter, the umbilical wall feebly convex, the umbilical shoulder relatively broadly rounded. The intercostal whorl section is depressed oval, the costal section polygonal. Primary ribs arise at the umbilical seam, flex back and are convex on the umbilical shoulder, rursiradiate and feebly flexuous across the flanks, convex on the inner flank, concave on the outer flank, projected forwards on the ventrolateral shoulder, where they terminate in feeble oblique elongated ventrolateral bullae, separated by a smooth zone from a blunt siphonal keel. The specimen bears periodic massive, blunt, outward-directed ventro-lateral horns, borne on a single rib. There are two on the adapertural half of the outer whorl.
Discussion
Ammonites cornutus has generally been referred to Dipoloceras by previous authors. It is however, significantly older than the type species, which is the marker fossil for the base of the Upper Albian, whereas cornutum, where well dated (Owen, 1971, p. 155) is upper Middle Albian. The presence of massive ventrolateral horns, not seen in Dipoloceras, links it to Dipoloceroides semicornutum, and this in turn links to the type species, Dipoloceroides delaruei. We illustrate here well-preserved individuals from the English Gault Clay (Fig. 6F-H, J-L), which further confirm the identity of the KwaZulu specimen.
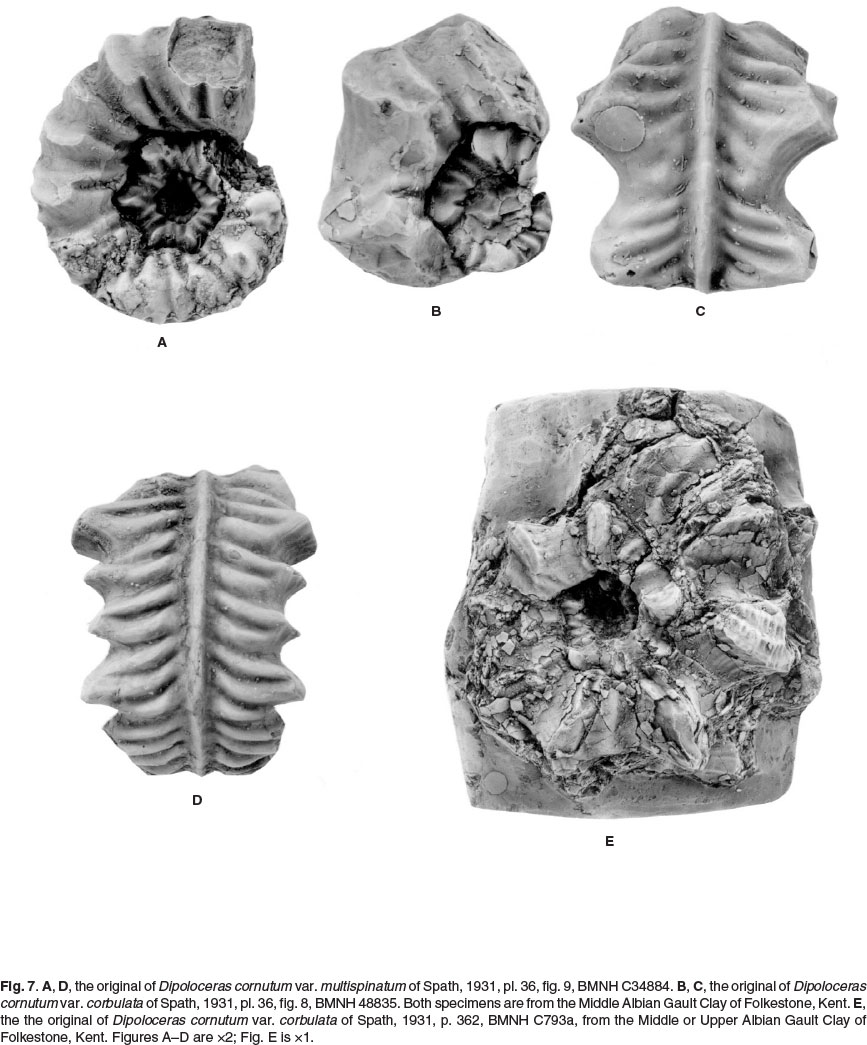
Spath (1931) introduced two varieties of this species, which we exclude from cornutum. The specimens described in Spath's text on Dipoloceras cornutum var. corbulata Spath, 1931 (p. 362) were never figured. One of the specimens referred to by Spath, BMNH C793a, is illustrated here (Fig. 7E). It is a large individual, over 80 mm in diameter, characterized by the development of flared ribs and strong spiral ridges; its affinities lie with lower Upper Albian forms of the type referred to as Cechenoceras by Van Hoepen (1941). The smaller specimen figured (Spath, 1931,pl. 36, fig. 8) but not mentioned by Spath in his text, is BMNH 48835 (Fig. 7B,C), which more closely resembles his variety multispinosa. The type of this form is BMNH C34884, the original of Spath, 1931, p. 363, pl. 38, fig. 9 (Fig. 7A,D). Spath noted transitions to Dipoloceras cristatum (1931, p. 308, pl. 32, fig. 3), and the affinities of multispinosa lie with Dipoloceras rather than Dipoloceroides cornutum in our view.
Occurrence
Where well-dated, this is an upper Middle Albian species. The geographic distribution is Southern England, northwest and southeast France, and northern KwaZulu-Natal.
Genus Falloticeras Parona & Bonarelli, 1897
Type species
Ammonites proteus d'Orbigny, 1842, p. 624, by the original designation of Parona & Bonarelli, 1897, p. 89.
Falloticeras proteus (d'Orbigny, 1842)
1842 Ammonites Proteus d'Orbigny, p. 624.
1850 Ammonites Proteus d'Orbigny; d'Orbigny, p. 124.
1860 Ammonites proteus d'Orbigny; Pictet & Campiche, p. 306.
1897 Falloticeras proteus d'Orb.; Parona & Bonarelli, p. 89, pl. 12, fig. 1.
1922 Falloticeras Spath, p. 97.
1931 Falloticeras Spath, pp. 346, 352.
1938 Falloticeras Proteus d'Orb.; Roman, p. 370, pl. 37, fig. 353, 353a, 353b.
1942 Falloticeras Spath, p. 708.
1947 Falloticeras Proteus d'Orb. sp.; Breistroffer, p. 30.
1949 Falloticeras proteus d'Orbigny; Collignon, p. 122.
1957 Falloticeras proteus (d'Orbigny); Wright, p. L304.
1959 Falloticeras cf. proteus (d'Orbigny); Casey, p. 207.
1963 Falloticeras proteus (d'Orbigny); Milbourne, table 1.
1971 Falloticeras proteum (d'Orbigny); Owen, p. 155.
1977 Falloticeras proteus (d'Orbigny); Kennedy & Cooper, p. 800, pl. 104, figs 1-19; pl. 105, figs 3, 6, 7, 10, 15; text-figs 1-3.
1983 Mojsisovicsia proteus (d'Orbigny); Gebhard, p. 116, figs 1b,c, 2.1, 2.2.
1996 Fallloticeras proteus (Orbigny); Wright, p. 137, fig. 105, 2a-c.
2006 Falloticeras proteus (d'Orbigny, 1842); Kennedy & Juignet in Gauthier, p. 176, pl. 41, figs 3, 4.
Type
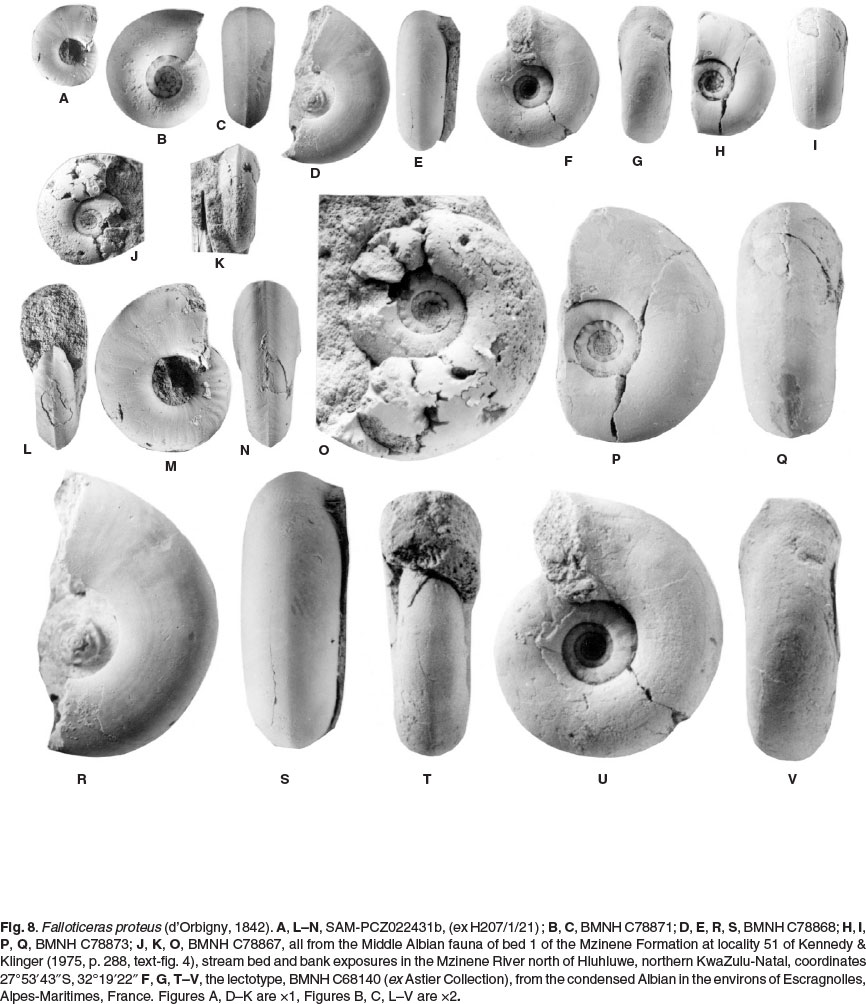
The lectotype, by the subsequent designation of Kennedy & Cooper (1977, p. 800) is BMNH C68140 (ex Astier Collection), from the condensed Albian in the environs of Escragnolles, Alpes-Maritimes, France, re-illustated here as Figure 8F,G, T-V. There are numerous paralectotypes, listed by Kennedy & Juignet (2006, p. 176).
Material
SAM-PCZ022431 (ex H207/1/21), SAM-PCZ17089, BMNH C78867, C78868, C78870, C78871, C78873, all from the Middle Albian fauna of bed 1 of the Mzinene Formation at locality 51 of Kennedy & Klinger (1975, p. 288, text-fig. 4), stream bed and bank exposures in the Mzinene River north of Hluhluwe, northern KwaZulu-Natal, coordinates 27°53'43"S, 32°19'22"E.
Dimension
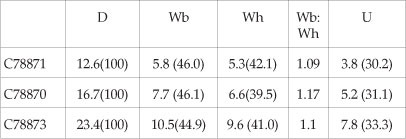
Description
BMNH C78871 (Fig. 8B,C) is an internal mould of a juvenile phragmocone 12.6 mm in diameter. Coiling is moderately involute, with a convex, outward-inclined wall and broadly rounded umbilical shoulder. The whorl section is depressed oval, with a whorl breadth to height ratio of 1.1. The flanks, ventrolateral shoulders, and venter are broadly convex. There is a distinct keel from the smallest diameter visible. The umbilical seam of the outer whorl is notched to acommodate the estimated 10 umbilical bullae per whorl of the penultimate whorl. There are no such bullae on the outer whorl of this specimen.
BMNH C78870 is 16.7 mm in diameter, and broken into two pieces. The adapertural 90° whorl sector of his specimen is body chamber. As in the previous specimen, there are coarse umbilical bullae, 10-12 on the penultimate whorl, but disappearing at the beginning of the outer whorl. The siphonal keel effaces at the adapertural end of the body chamber.
BMNH C78867 (Fig. 8J,K,O) is the phragmocone of a more compressed individual, 20.4 mm in diameter, the shallow umbilicus comprising 34% of the diameter. The siphonal keel is strong throughout. There are well-developed umbilical bullae on the adapical 90° sector of the penultimate whorl, and all of the antepenultimate whorl. SAM-PCZ022431 (ex H207/1/21) (Fig. 8A, L-N) is a similarly compressed individual, with well-developed ribs on the adapical part of the outer whorl.
BMNH 78873 (Fig. 8H,I,PQ) is 23.4 mm in diameter, retains well-preserved aragonitic shell, and is part body chamber. Coiling is moderately involute, the umbilicus of moderate depth, comprising 33% of the diameter. The umbilical wall is broadly convex and outward-inclined, the umbilical shoulder broadly rounded. The whorl section is only slightly compressed, with a whorl breadth to height ratio of 1.1, the flanks feebly convex, the ventrolateral shoulders and venter broadly rounded. There are 12 coarse umbilical bullae on the 90° adapertural sector of the penultimate whorl, and the adapical 270° sector of the outer whorl. The surface of the shell is ornamented by delicate growth lines and striae, convex across the umbilical wall and shoulder, prorsiradiate across the flanks, concave on the ventrolateral shoulders, and projected forwards to produce an obtuse ventral chevron, with the blunt siphonal keel at the apex. The keel declines and effaces on the adapertural 90° sector of the outer whorl.
BMNH C78868 (Fig. 8D,E,R,S) is a beautifully preserved internal mould of a 180° sector of body chamber. The whorl breadth to height ratio is 1.05 at the adapical end ofthe body chamber. The external mould of the antepenultimate whorl reveals the presence of crenulations in the umbilical seam, indicating the presence of of strong umbilical bullae on the inner whorls, as in the previous specimens. The ventral keel is very subdued at the adapical end of the body chamber, and effaces progressively, leaving only a blunt mid-ventral angulation at the greatest preserved diameter.
Discussion
The KwaZulu specimens match well with the lectotype: compare Figs 8P-S and 8T-V, leaving no doubt as to the affinities of the present material.
Occurrence
Lower Middle Albian of southeast France, southern England, and northern KwaZulu-Natal. Gebhard (1983, p. 116) records the species from Austria, but the specimen was not illustrated.
ACKNOWLEDGEMENTS
Kennedy acknowledges the support of the staff of the Geological Collections, Oxford University Museum of Natural History, and the Department of Earth Sciences, Oxford, and the financial assistance of the Oppenheimer Fund (Oxford). Klinger acknowledges support from the staff of the Iziko South African Museum, Natural History Collections Department and financial assistance from the NRF, South Africa. The originals of Figs 5B-D, 6F-H, J-L, were supplied by the photographic unit of the Department of Palaeontology, The Natural History Museum, London. Thanks are due to drs F. Amédro (Calais) and J.-L. Latil (Lazer) for their reviews of the manuscript.
REFERENCES
BREISTROFFER, M. 1947. Sur les zones d'ammonites dans l'Albien de France et d'Angleterre. Travaux du Laboratoire de Géologie de la Faculté des Sciences de l'Université de Grenoble 26: 17-104 (1-88 in separates). [ Links ]
CASEY, R. 1959. Field meeting at Wrotham and the Maidstone by-pass. Proceedings of the Geologists' Association 70: 206-209. [ Links ]
COLLIGNON, M. 1949. Recherches sur les faunes albiennes de Madagascar I. L'Albien d'Ambarimaninga. Annales Géologiques du Service des Mines, Madagascar 14: 1-128. [ Links ]
COOPER, M.R. 1982. Lower Cretaceous (Middle Albian) ammonites from Dombe Grande, Angola. Annals ofthe South African Museum 89, 265-314. [ Links ]
DOUGLAS, J.A. 1921. Geological sections through the Andes of Peru and Bolivia: III From the Port of Callao to the River Perene. Quarterly Journal of the Geological Society 77: 246-284. [ Links ]
GABB, W.M. 1877. Description of a collection of fossils made by Dr. Antonio Raimondi in Peru. Journal of the Academy of Natural Sciences of Philadelphia (new series) 8: 263-336. [ Links ]
GAUTHIER, H. 2006. Révision Critique de la Paléontologie Française d'Alcide d'Orbigny, 6, Céphalopodes Crétacés. 292 + 662 + 28 pp., 65 + 146 + 9 pls. Leiden: Backhuys. [ Links ]
GEBHARD, G. 1979. Glauconitische Kondensation im Alb der sub-alpinen Ketten (Clars, Escragnolles, SE-Frankreich), deren Ammonitenfauna und Kartierung in der Umgebung von Escragnolles. Unpublished thesis, University of Tübingen, 1-152. [ Links ]
GEBHARD, G. 1983. Taxonomische Konsequenzen aus der Synonymie von Mojsisovicsia Steinmann und Falloticeras Parona & Bonarelli (Ammonidea, Alb). Paläontologische Zeitschrift 57: 111-119. [ Links ]
GROSSOUVRE, A. de 1894. Recherches sur la craie supérieure, 2, Paléontologie. Les ammonites de la craie supérieure. Mémoires du Service de la Carte Géologique détaillée de la France: 1-264. [ Links ]
HAUGHTON, S.H. 1936. Account of the geology of the Cretaceous beds and a preliminary analysis of the associated ammonite fauna. In: RENNIE, J.L.V. Lower Cretaceous Lamellibranchia from Northern Zululand. Annals of the South African Museum 31: 283-297. [ Links ]
HOEPEN, E.C.N. VAN. 1941. 3. Die gekielde ammoniete van die Suid-Afrikaanse Gault. Dipoloceratidae, Cechenoceratidae en Drepanoceratidae. Paleontologiese Navorsing van die Nasionale Museum, Bloemfontein 1: 55-90. [ Links ]
HYATT, A. 1889. Genesis of the Arietidae. Smithsonian Contributions to Knowledge 673: xi + 1-239. [ Links ]
HYATT, A. 1900. Cephalopoda. In: ZITTEL, K.A. VON, 1896-1900, Textbook of Palaeontology (transl. Eastman, C.R.), pp. 502-604. London and New York: Macmillan. [ Links ]
HYATT, A. 1903. Pseudoceratites of the Cretaceous. United States Geological Survey Monograph 44: 1-351. [ Links ]
KENNEDY, W J. & COOPER, M.R. 1977. The micromorph Albian ammonite Falloticeras Parona and Bonarelli. Palaeontology 20: 793-804. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 1975. Cretaceous faunas from Zululand and Natal, South Africa. Introduction, stratigraphy. Bulletin of the British Museum (Natural History) Geology 25: 263-315. [ Links ]
KORN, D., EBBIGHAUSEN, V., BOCKWINKEl, J. & KLUG, C. 2003. The A-mode ontogeny in prolecanitid ammonites. Palaeontology 46: 1123-1132. [ Links ]
KULLMANN, J. & WIEDMANN, J. 1970. Significance of sutures in phylogeny of Ammonoidea. University of Kansas, Paleontological Contributions 42: 1-32. [ Links ]
LISSON, C.I. 1908. Contribución al conocimiento sobre algunos ammonites del Perú: 1-64. Lima: Tipografia 'El Perú [ Links ]'.
LISSON, C.I. 1911. Terrenos reconocidas hasta hoy en el Peru. Boletín de Minas, Industrias, y Construcciones 3: 141-172. [ Links ]
LISSON, C.I. 1924. Edad de los fósiles peruanos y distribución de sus depósitos en toda la républica. Contribucion geologia del Peru 1924: 1-226. [ Links ]
MARCINOWSKI, R. & WIEDMANN, J. 1990. The Albian ammonites of Poland. Palaeontologica Polonica 50: 1-94. [ Links ]
MILBOURNE, R.A. 1963. The Gault at Ford Place, Wrotham, Kent. Proceedings of the Geologists' Association 74: 55-79. [ Links ]
NEUMAYR, M. 1875a. Die Ammoniten der Kreide und die Systematik der Ammonitiden. Zeitschrift der Deutschen Geologischen Gesellschaft 27: 854-942. [ Links ]
NEUMAYR, M. 1875b. ÜberKreideAmmonitiden. Sitzungsberichte der Kaiserlichen Kóniglichen Akademie der Wissenschaften in Wien, Mathematisch-Naturwissenschaftliche Klasse Abt. 1, 71: 639-693. [ Links ]
ORBIGNY, A. d'. 1840-1842. Paléontologie française: Terrains crétacés. 1. Céphalopodes. Masson: Paris. 1-120 (1840); 121-430 (1841); 431-662 (1842). [ Links ]
ORBIGNY, A. d'. 1850. Prodrome de Paléontologie stratigraphique universelle des animaux Mollusques et rayonnés faisant suite au cours élémentaire de Paléontologie et de Géologie stratigraphique 2: 1-427. Paris: Masson. [ Links ]
OWEN, H.G. 1971. Middle Albian stratigraphy in the Anglo-Paris Basin. Bulletin ofthe British Museum of Natural History (Geology), supplement 8: 1-164. [ Links ]
PARONA, C.F. & BONARELLI, G. 1897. Fossili Albiani d'Escragnolles, del Nizzardo e della Liguria occidentale. Palaeontographica Italica 2: 53-107 (1-55). [ Links ]
PASSENDORFER, E. 1930. Studjum stratygraficzne i paleontologiczne nad Kreda serji wierchowej w Tatrach. [Étude stratigraphique et paléontologique du Crétacé de la série hauttatrique dans les Tatras.] Prace Polskiego Instytutu Geologicznego 2 (for 1929): 439-677 [In Polish and French. [ Links ]]
PICTET, F.J. 1847. Description des mollusques fossiles qui se trouvent dans les Grès Verts des environs de Genève. Mémoires de la Société de Physique et d' Histoire Naturelle de Genève 11: 257-412. [ Links ]
PICTET, F.J. & CAMPICHE, G. 1860. Description des fossiles du terrain Crétacé des environs de Sainte-Croix. 2. Matériaux pour la Paléontologie Suisse (Séries 2): 209-380. [ Links ]
ROBERT, E. 2002. La transgression Albienne dans le Bassin Andin (Pérou: Biostratigraphie, Paléontologie (Ammonites) et Stratigraphie Séquentielle. Strata 38: 1-380. [ Links ]
ROMAN, F. 1938. Les ammonites jurassiques et crétacées. Essai de genera. 554 pp. Masson: Paris. [ Links ]
SPATH, L.F. 1921. On Cretaceous Cephalopoda from Zululand. Annals of the South African Museum 12: 217-321. [ Links ]
SPATH, L.F. 1922. On Cretaceous Ammonoidea from Angola, collected by Professor J.W. Gregory, D.Sc., F.R.S. Transactions of the Royal Society of South Africa 53: 91-160. [ Links ]
SPATH, L.F. 1923. Excursion to Folkestone, with notes on the zones of the Gault. Proceedings of the Geologists' Association 34, 70-76. [ Links ]
SPATH, L.F. 1926. On the zones of the Cenomanian and the upper-most Albian. Proceedings of the Geologists' Association 37: 420-432. [ Links ]
SPATH, L.F. 1930. The fossil fauna of the Samana Range and some neighbouring areas. V. The Lower Cretaceous Ammonoidea with notes on Albian Cephalopods from Hazara. Palaeontologica Indica (new series) 15: 51-66. [ Links ]
SPATH, L.F. 1931. A monograph of the Ammonoidea of the Gault. Part 8. Palaeontographical Society Monographs: 379-410. [ Links ]
SPATH, L.F. 1934. A monograph of the Ammonoidea of the Gault. Part 11. Palaeontographical Society Monographs: 443-496. [ Links ]
SPATH, L.F. 1942. A monograph of the Ammonoidea of the Gault. Part 14. Palaeontographical Society Monographs: 609-668. [ Links ]
SPATH, L. F. 1942. A monograph of the Ammonoidea of the Gault. Part 15. Palaeontographical Society Monographs: 669-720. [ Links ]
STEINMANN, G. 1881. Über Tithon und Kreide in den Peruanischen Anden. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 2: 130-153. [ Links ]
STEINMANN, G. 1882. Über Jura und Kreide in den Anden. Neues Jahrbuch far Geologie und Paläontologie 1881: 130-153. [ Links ]
STIELER, K. 1920. Über sogenannte Mortoniceraten des Gault. Zentralblatt far Mineralogie, Geologie und Paláontologie 1920: 345-352. [ Links ]
STIELER, K. (1922) Über Gault- und Cenoman-Ammoniten aus dem Cenoman des Cap Blanc Nez. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilage Band 47: 19-44. [ Links ]
WRIGHT, C.W. 1957. [Cretaceous Ammonoidea]. In: MOORE, R.C. (ed.) Treatise on Invertebrate Paleontology. Part L, Mollusca 4, Cephalopoda Ammonoidea. xxii + 1-490 pp. New York and Lawrence: Geological Society of America and University of Kansas Press. [ Links ]
WRIGHT, C.W. 1996. Treatise on Invertebrate Paleontology. Part L, Mollusca 4 Revised: Cretaceous Ammonoidea. xx + 1-362 (with contributions by J.H. Calloman (sic) and M.K. Howarth). Lawrence, Kansas and Boulder, Colorado: Geological Society of America and University of Kansas. [ Links ]
Received 2 July 2012
Accepted 11 August 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}