










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkAfrican Natural History
versión On-line ISSN 2305-7963
versión impresa ISSN 1816-8396
Afr. nat. history (Online) vol.7 Cape Town ene. 2011
Hoplitoplacenticeras (H.) howarthi Collignon, 1970 (Cephalopoda: Ammonoidea) from KwaZulu-Natal, South Africa and Madagascar; intraspecific variation, dimorphism and affinities
Herbert Christian KlingerI; William James KennedyII
INatural History Collections Department, Iziko South African Museum, P.O. Box 61, Cape Town. 8000 South Africa E-mail: hklinger@iziko.org.za
IIDepartment of Earth Sciences, Oxford University, South Parks Road, Oxford 0X1 3AN, U.K. E-mail: jim.kennedy@oum.ox.ac.uk
ABSTRACT
Additional specimens of the Campanian ammonite Hoplitoplacenticeras (H.) howarthi Collignon, 1970, as well as examination of Collignon's (1970) Madagascar! specimens of Hoplitoplacenticeras allow us to discuss the intraspecific variation, probable dimorphism and affinities of the species.
Key words: ammonites, Hoplitoplacenticeras; Upper Campanian, intraspecific variation, dimorphism, affinities, Madagascar, KwaZulu-Natal, South Africa.
INTRODUCTION
In September 2007 one of us (H.C.K.) collected several specimens of the Campanian ammonite Hoplitoplacenticeras (H.) howarthi Collignon, 1970 from the Nibela Peninsula in KwaZulu-Natal, South Africa. These specimens add to the previous description of the species from the same locality by Klinger & Kennedy (1989), subsurface material from Durban (Kennedy & Klinger 1973) and Madagascar (Collignon 1970). This species is extremely variable as far as ornamentation and relative proportions are concerned. We were able to examine the holotype and other representatives of the subgenus from Madagascar and compare these with some of the better-known representatives of the subgenus from other geographic regions. In addition we can recognize the probable nature of dimorphism in Hoplitoplacenticeras (H-) howarthi.
LOCALITY DATA, REPOSITORIES AND CONVENTIONS
Details of localities in KwaZulu-Natal, South Africa are given in Kennedy & Klinger (1975).
The following abbreviations are used to indicate the repository of specimens:
SAM: Natural History Collections Department, Iziko South African Museum, Cape Town.
GPIB: Institut für Paleontologie der Rheinischen Priedrich- Wilhelms-Universitat, Bonn.
GPIG: Geologisch-Paláontologisches Institut und Museum der Georg-August Universitat, Göttingen
NMB: National Museum, Bloemfontein (on permanent loan to SAM)
UBGD: UFR Sciences de la Terre, Université de Bourgogne, Dijon (Collignon Collection) SP: Sorbonne Collections, Paris.
EMP: Ecole National Supérieur des Mines Collections, now housed in the Université de Lyon, Villeurbanne.
OUM: Oxford University Museum of Natural History, Oxford.
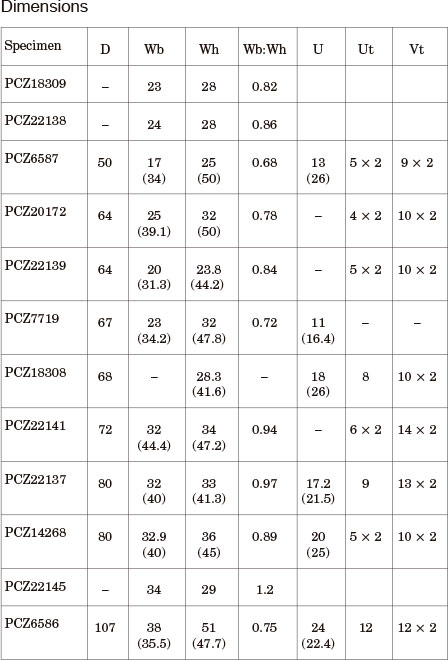
Dimensions are given in millimetres; numbers in brackets indicate a percentage of the measured diameter. D = diameter; Wb = whorl breadth, Wh = whorl height, Wb:Wh = ratio of whorl breadth to whorl height; U = umbilical diameter. Ut and Vt refer to the number of umbilical and ventrolateral tubercles respectively. Numbers followed by 'x' indicate tubercles per half whorl.
SYSTEMATIC PALAEONTOLOGY
Order AMMONOIDEA Zittel, 1884
Suborder AMMONITINA Hyatt, 1889
Superfamily HOPLITOIDEA Douvillé, 1890
Family PLACENTICERATIDAE Meek, 1876
Genus Hoplitoplacenticeras Paulcke, 1907 (ICZN name No. 1348)
(= Dechenoceras Kayser, 1924)
Subgenus Hoplitoplacenticeras (Hoplitoplacenticeras) Paulcke, 1907.
Type species
Hoplites-Placenticeras plasticus Paulcke, 1907, p. 186, ICZN Opinion 554, 1959, Name No. 1629).
For an extensive review and discussion of the genus Hoplitoplacenticeras see Kennedy (1986, p. 63) and description of the European, specifically the German representatives of the genus see Kaplan et al. (1996,2005) and Kennedy & Kaplan (1997). Hoplitoplacenticeras differs from other genera referred to the family Placenticeratidae Meek, 1876 (see e.g. Kennedy & Wright 1983 and Klinger & Kennedy 1989) mainly by the ontogenetic change in ornament from the phragmocone to the body chamber, especially on the later part of the latter towards the aperture. The ontogenetic changes include weakening of tubercu-lation and ribbing, leaving ultimately a body chamber covered in dense lirae and striae only or with near-effaced tubercles.
Hoplitoplacenticeras (Hoplitoplacenticeras) howarthi
Collignon, 1970 Figs 1-6
1931 Hoplites Vari Schlüter; Basse p. 35, pl. 5, figs 1-3; pl. 12, fig. 2; pl. 13, fig. 1.
1970 Hoplitoplacenticeras marroti Coquand (= H. vari Schlüter); Collignon, p. 75, pl. 638, figs 2342-2343.
1970 Hoplitoplacenticeras sp. aff. marroti Coquand; Collignon, p. 80, pl. 639, fig. 2353.
1970 Hoplitoplacenticeras sp. aff. plasticum-crassum Paulcke; Collignon, p. 75, pl. 638, fig. 2344; p. 80, pl. 639, fig. 2352.
1970 Hoplitoplacenticeras cf. dolbergense Schlüter; Collignon, p. 75, pl. 638, fig. 2345.
1970 Hoplitoplacenticeras cf. costulosum Schlüter; Collignon, p. 76, pl. 638, fig. 2346.
?1970 Hoplitoplacenticeras trangahyense Collignon, p. 76, pl. 638, fig. 2347.
1970 Hoplitoplacenticeras antokazoense Collignon, p. 76, pl. 638, fig. 2348; p. 80, pl. 639, fig. 2354.
1970 Hoplitoplacenticeras besairiei Collignon, p. 77, pl. 638, figs 2349-2350.
1970 Hoplitoplacenticeras howarthi Collignon, p. 80, pl. 639, fig. 2351.
1973 Hoplitoplacenticeras plasticum plasticum Paulcke; Kennedy & Klinger, p. 102, pl. 5, figs 4a-e.
1989 Hoplitoplacenticeras howarthi Collignon; Klinger & Kennedy, p. 358, figs 14b, 108-109.
Name of the species
As first revising authors we select howarthi as the name of the species
Type
Holotype by monotypy is the original of Collignon (1970, p. 80, pl. 639, fig. 2351 UBGD12351 (Fig. 8G-I) housed in the collections of the UFR Sciences de la Terre, Université de Bourgogne, Dijon, from the Upper Campanian of Gisement 227-2, Mokotibe (Antsalova), Madagascar.
Material
SAM-PCZ6586-6587, PCZ7421, 7711, 7719, PCZ18308 (ex NMB-D1314) PCZ18309, PCZ14268, PCZ22138-22145, OUM KX8284-6, all from locality 110, cliff and foreshore sections at the southwestern tip of the Nibela Peninsula, and SAM-PCZ20172, from locality 109F, to the west of the former locality in foreshore exposures; both St Lucia Formation, Campanian III. A single specimen is from excavations at Somtseu Road, Durban in the collections of the Geology Department, University of Natal, Mzamba Formation, Campanian II?
Description
As is the case in the family Placenticeratidae in general, (see e.g. Klinger & Kennedy 1989), but specifically in the genus Hoplitoplacenticeras, the material is extremely variable, both in terms of strength and density of lateral ornament and relative proportions of the shell. This is further complicated by the extreme changes in ornament in the transition from the phragmocone to the body chamber, as well as the effects of dimorphism.
Despite this wide variation in morphology, the following general diagnosis applies to H. (H.) howarthi from KwaZulu-Natal:
Narrowly umbilicate, umbilical wall on phragmocone near-vertical, whorl section trapezoidal, generally higher than wide, ornament consists ofprominent, conical umbilical tubercles, connected to bifurcating, prorsiradiate sinusoidal ribs with inner and forwardly displaced outer ventro-lateral tubercles. On body chamber ornament changes, with ribbing becoming closer spaced, less prominent, and weaker. The whorl section becomes more rounded, and the umbilical wall tends to slope outwards. Adult size varies considerably, probable microconchs and macroconchs can be distinguished, but relative proportions between these overlap considerably. Complete apertures are unknown.
Early ontogeny
The early ontogenetic stage is partially visible on the inner whorls of PCZ22145 (Fig. 5A-C) and preserved as an impression on PCZ22141 (Fig. 2A-C). It is best seen on the juvenile specimen from Somtseu Road in Durban figured by Kennedy & Klinger (1973, pl. 5, fig. 4a-c) where the dominant ornament consists of numerous, minute, spirally elongated outer ventrolateral, and larger, less numerous inner ventrolateral tubercles. At a diameter of about 20 mm in the small figured specimen, the prominent umbilical tubercles are already visible, but the lateral ribbing is poorly developed.
Variation
In our material, extreme forms in ornament, size and whorl section in the later part of the phragmocone can be distinguished, but these features overlap to such an extent that they have to be considered as part of the intraspecific variation.
Specimens with fine, dense ribbing include PCZ22140 (Fig. 4F) and PCZ7719 (Fig. 4A-C), and those with extremely coarse ribbing, e.g. PCZ20172 (Fig. 3D,G), PCZ7711 (Fig. 3C,H) andPCZ18308 (Fig. 6A-B). Specimens with compressed whorl sections include PCZ22141 (Fig. 2A-C), PCZ22143 (Fig. 3E-F) while PCZ22145 (Fig. 5A-C) has an extremely wide whorl section. Specimens like PCZ22137 (Fig. 1A-E) are intermediate with a more rounded whorl section. Small specimens with distinct body chamber modifications such as PCZ7719 (Fig. 4A-C) maybe regarded as microconchs, and large specimens, e.g. PCZ6586 (Fig. 6C-D) macroconchs, but specimens with dimensions which fall between these, though apparentlynear-adult, e.g. PCZ22141 (Fig. 2A-C), PCZ22140 (Fig. 4F) and PCZ18308 (Fig. 6A-B) cannot unequivocably be designated as either macro- or microconchs As to be discussed below, we suspect that there may be a correlation between absolute size and whorl section; those specimens with compressed whorl sections becoming adult at smaller diameters than those with wider, more inflated whorl sections.
Discussion
Aperture
Despite the abundance of specimens available, complete apertures in the genus (and in the family Placenticeratidae) are a rarity. A complete body chamber and aperture were recorded in H. (H.)preyi (Kennedy & Summesberger 1999, p. 25, pl. 3, figs 1, 4) and another possibly in Hoplito-placenticeras (Lemfoerdiceras) lemfoerdense from Poland by Machalski et al. (2004, p. 458, pl. 5, fig. 2). Numerous specimens of Hoplitoplacenticeras from Germany described by Kaplan et al. (1996) appear to have parts ofthe aperture preserved in which the shape of the aperture seems to follow that of the growth lines on the body chamber. PCZ22137 (Fig. 1A-E) appears to have lateral sinuses and a ventral rostrum. The former, however, are asymmetric on either side, and truncate, rather than follow the general direction of the lateral ornament (Fig. 1A,B); the latter (Fig. 1E) also appears to truncate the ventral ribbing. Because of this, we have to interpret the apparent lateral sinuses and ventral rostrum in PCZ22137 (Fig. 1A-E) as probably due to biological (arthropod?) or mechanical damage to the bodychamber rather than as indications of a complete aperture.
Dimorphism
In discussions on the family Placenticeratidae, and specifically the genus Hoplitoplacenticeras several references have been made to dimorphism. The earliest reference to the presence of possible dimorphism in the genus was already made by Paulcke (1907 p. 52[218] in his discussion of H. (H.) plasticus. He suggested that the crassus-types, i.e. the inflated forms were females (macroconchs) and the compressed laevis-forms, male (i.e. microconchs). Subsequent references to dimorphism made mainly on differences in size (in Hoplitoplacenticeras) are found in e.g. Kennedy & Wright (1983, p. 868); Kennedy 1986, p. 63); Klinger & Kennedy (1989); Cobban & Kennedy (1993, p. 73); Kaplan et al. 1996) and Kaplan et al. (2005)
We suggest that compressed forms mature at smaller diameters than forms with quadrate whorl sections, and are consequently tentatively referred to as micro- and macro-conchs respectively.
Comparison with other species
To date, the following species in chronological order have been referred to the genus Hoplitoplacenticeras s.l.: Hoplitoplacenticeras (H.) lafresnayanum (d'Orbigny, 1841)
H. (H.) marroti (Coquand, 1859)
H. (H.) vancouverense (Meek, 1862) H. (H.) coesfeldiense (Schlüter, 1867) H. (H.) costulosum (Schlüter, 1867)
H. (L.) lemfoerdense (Schlüter, 1872)
(= Ammonites scaphitoides Schlüter, 1872)
H. (H.) vari (Schlüter, 1872)
(= Ammonites striatocostatus Schlüter, 1872)
H. (H.) dolbergense (Schlüter, 1876)
H. (H.) rejaudryi (de Grossouvre, 1894)
H. (H.) gosseleti (de Grossouvre, 1894)
H. (H.) marroti praematura (Imkeller, 1901)
H. (H.) plasticus Paulcke, 1907.
H. (H.) coesfeldiense var. schlueteri Mikhailov, 1951 H. (H.) vari var. nov. ind. Mikhailov, 1951 H. (H.) trangahyense Collignon, 1970 H. (H.) antokazoense Collignon, 1970
H.(H.) besairiei Collignon, 1970 H. (H.) howarthi Collignon, 1970
H. (H.) rarecostatum Khakimov, 1976
H. (H.) monju Matsumoto, 1982
H. (H.) fugen Matsumoto, 1984
H. (H.) minor Cobban & Kennedy, 1993
H. (H.) preyi Kennedy & Summesberger, 1999
Doubtful contenders that have been referred to the genus include:
(?) Scaphites kambysis Zittel in: Quaas, 1902 Hoplitoplacenticeras awadi Hassan,1971.
According to Kennedy (1986, pp. 68-70) these latter two are possibly scaphitine homoeomorphs of Hoplitoplacenticeras, and may, for the present, be disregarded in the discussion. Also, H. lemfoerdense and H. lafresnayanum (the latter from the Maastrichtian), differ from other Hoplitoplacenticeras by the presence of an additional set of ventral tubercles, and are referred to the subgenus H. (Lemfoerdiceras) Kennedy, 1986.
Because of the extreme variation, not only in our material, but in the subgenus Hoplitoplacenticeras in general, it is difficult to give precise differences between all these species that have been referred to the subgenus.
Our first record of Hoplitoplacenticeras from subsurface exposures at Somtseu Road, Durban, was identified as H. plasticum plasticum (Kennedy & Klinger 1973, p. 102).
When we first discovered specimens of H. (Hoplito-placenticeras) at locality 110 on the Nibela Peninsula, H. (H.) howarthi Collignon (1970, 80, pl. 639, fig. 2351) from the Upper Campanian of Madagascar appeared to be the closest match, and the present material seems to confirm this, especially as far as the whorl section and prominent umbilical tubercles are concerned. The affinities ofthe other species of Hoplitoplacenticeras described by Collignon from only two Upper Campanian localities are more difficult to resolve. Collignon described specimens from the Hoplito-placenticeras marroti Zone at Gisement 240 'de la 2° Butte-témoin de la piste Trangahy-Antokazo (Antsalova)' as H. marroti Coquand (= H. vari Schlüter) (Collignon 1970, p. 75, pl. 638, figs 2342-3) (Fig. 7A-B, C-D); H. sp. aff. plasticum-crassum Paulcke (Collignon 1970, p. 75, pl. 638, fig. 2344) (Fig. 7E-F); H. cf. dolbergense Schlüter (Collignon 1970, p 75, pl. 638, fig. 2345) (Fig. 7G-H); H. cf. costulosum Schlüter (Collignon 1970, p. 76, pl. 638, fig. 2346) (Fig. 7I-K); H. trangahyense Collignon (1970, p. 76, pl. 638, fig. 2347) (Fig. 8A-C); H. antokazoense Collignon (1970, p. 76, pl. 638, fig. 2348) (Fig. 8D-F) and H. besairiei Collignon (1970, p. 77, pl. 638, figs 2349-50) (Fig. 7L-N, 9H-I) and from the same zone, at gisement 227-2 Mokotibe (Antsalova) H. howarthi (Collignon 1970, p. 80, pl. 639, fig. 2351) (Fig. 8G-I); H. sp. aff. plasticum-crassum Paulcke (Collignon 1970, p. 80, pl. 639, fig. 2352) (Fig. 9C-D); H. sp. aff. marroti Coquand (Collignon 1970, p. 80, pl. 639, fig. 2353) (Fig. 9E-G) and H. antokazoense Collignon (1970, p. 80, pl. 639, fig. 2354) (Fig. 9A-B).
We suspect that all these 'species' fall within the variation of H. (H.) howarthi (see Klinger & Kennedy 1989, p. 360). H. marrotti, H. cf. costulosum and H. besairiei of Collignon can be compared with the KwaZulu specimens with a compressed whorl section; H. cf. dolbergense, H. trangahyense and H. antokazoense of Collignon correspond to the specimens with a quadrate whorl section, whereas the remaining Madagascan 'species' can be referred to those specimens with 'intermediate', slightly rounded whorl sections. As first revising authors, we choose the name H. (H.) howarthi Collignon for this variable Afro-Malagassy species.
Basse (1931, p. 35, pl. 5, figs 1-3; pl. 12, fig. 2; pl. 13, fig. 1) had previously described Hoplites Vari Schlüter from '8 km. au Nord de Trangahy, sur la route d'Antsalova, dans les calcaires subcrayeux du Niveau 10' . These specimens have the same general ornament as compressed varieties of H. (H.) howarthi, and undoubtedly belong to this species. The other species described by Basse (1931, p. 36, pl. 4, figs 5-6; pl. 13, fig. 3 as Hoplites (Hoplitoplacenticeras) plasticus from 'Entre Mokotibe et la forêt située á l'Ouest de ce village Niveau 10', was referred by Collignon (1970, p. 76, pl. 638, fig. 2346) to Hoplitoplacenticeras cf. costulosum Schlüter. This is a very compressed form with hardly any lateral ornament, and probably is only an extremely compressed and smooth variety of H. (H.) howarthi.
As far as the strong ornamentation is concerned, H. (H.) howarthi bears some similarityto H. (H.) plasticus (Paulcke, 1907) and its 'varieties' (Untergruppen) Hauthali, crassus, costatus, semicostatus and laevis (see Paulcke 1907, p. 20[186]-54[220]. Pls 10(1) -15(6), figs 1-3; Riccardi 1988, pl. 16, figs 4-6 and herein Fig. 10A-G). Paulcke (1907) listed 48 specimens which he referred to five morphological groups ('Untergruppen'), Hauthali, crassus, costatus, semicostatus and laevis, based more or less on decreasing whorl breadth and strength of ornamentation, though realizing he was probably dealing with a single, variable species. Despite the extreme variation in that species, the umbilical tubercles are never as spinose as in H. (H.) howarthi, but are elongated obliquely radial, and the umbilical wall slants outwards rather than being vertical as in the latter. (see Fig. 10A-G) Also, none of our specimens has as rounded and inflated a whorl section as the 'Hauthali' variety (Fig. 10D-E) of the latter species. Thus as here interpreted, none of the Madagascan specimens referred in open nomenclature to H. plasticus by Collignon belong to that species.
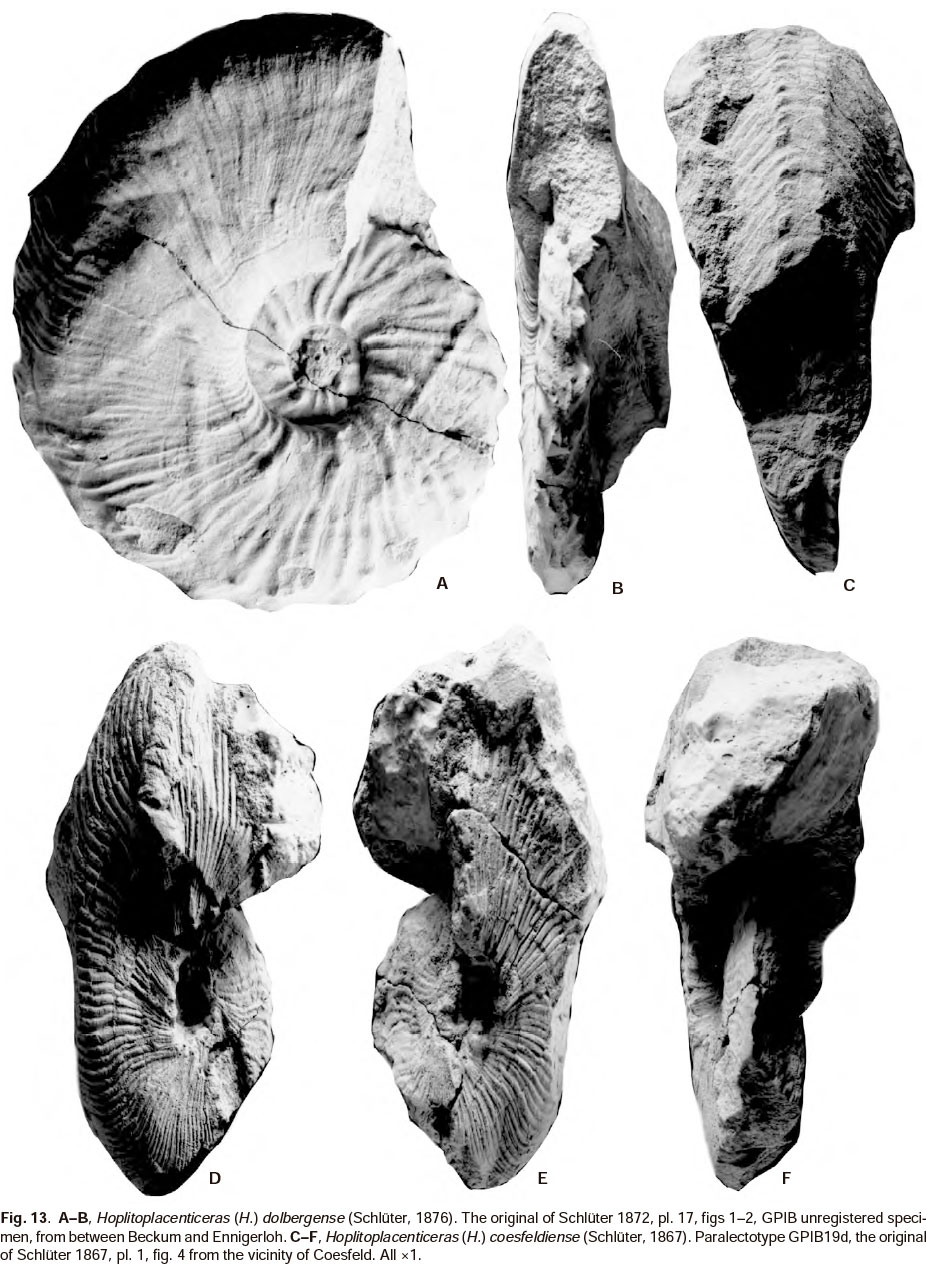
Of the predominantly European species, only H. ( H. ) dolbergense (Schlüter, 1876) (1876, p. 159, pl. 44, figs 1-4), comprehensively revised by Kaplan et al. (1996, p. 39, pl 25, figs 3-4; pl. 26, fig. 5; pl. 27, figs 3-4; pl. 28, figs 1-5; pl. 29, figs 1-4; pl. 30, figs 1-5; pl. 31, figs 1-9; pl. 32, figs 4-5) and mentioned subsequently (Kaplan et al. 2005, p. 100) (see also Figs 11G-K, 13A-B) is close to H. (H.) howarthi as far as strength of ornament is concerned. It lacks the strong umbilical tubercles of H. (H.) howarthi, and tends to form looped ribbing on the flanks; a feature not present in the latter. Only the holotype (by monotypy) of H. (H.) trangahyense (Fig. 8A-C) has weakly looped ribbing, similar to that of H. (H.) dolbergense, as noted by Collignon (1970, p. 76) and Kennedy (1986, p. 66) and may either be regarded as an atypical variant of H. (H.) howarthi, or as belonging to a different species; on the basis of a single specimen we cannot be more definite. The specimen referred to H. cf. dolbergense by Collignon (1970, p. 75, pl. 638, fig. 2345) (here Fig. 7G,H) lacks the looped ribbing of typical H. (H.) dolbergense and may best be regarded as an inflated form of H. (H.) howarthi with a rounded whorl section.
All the other European species revised by Mikhailov (1951), Kennedy (1986), Kaplan et al. (1996, 2005, 2006), Kennedy & Kaplan (1997), e.g. H. (H.) vari (Schlüter, 1876), (here Figs 12, 14A), H. (H.) costulosum (Schlüter, 1867) (here Figs 11B-F; 14B-E) and H. (H.) coesfeldiense (Schlüter, 1867) (here Figs 11A, 13C-F) have much finer and denser ribbing than H. (H.) howarthi and lack the prominent umbilical tubercles.
Ornament on the body chamber of the macroconch of H. (H.) howarthi (Fig. 6C-D) from KwaZulu superficially resembles that of H. (H.) coesfeldiense, but ornament on the inner, phragmocone whorls is totally different.
The specimen referred to H. cf. costulosum by Collignon (1970, p. 76, pl. 638, fig. 2345) (here Fig. 7I-K) has the strong umbilical tubercles typical of H. (H.) howarthi; it is here merely regarded as a compressed and weakly ornamented form of the latter species.
H. (H.) marroti (Coquand, 1859) (see Kennedy 1986, p. 70, pl. 2, figs 3-4; pl. 9, figs 1-8, 11-12; pl. 10, figs 1-12; pl. 12, figs 1-2) (here Fig. 15) has similar, though apparently denser lateral ornament than H. (H.) howarthi and differences between the two species are difficult to formulate. None of the known specimens of H. (H.) marroti are known to have as coarse ornament as some H. (H.) howarthi, (e.g. Figs 3D,G; 6A-B), or to grow to such large size, but some of the Madagascan specimens, e.g. Collignon's (1970, p. 75, pl. 638, fig. 2342) (here Fig. 7A-B) or H. (H.) besairiei (Collignon 1970, p. 77, pl. 638, fig. 2349) (lectotype here designated; Fig. 7L-N) are impossible to distinguish from the French specimens ofH. (H.) marroti figured by Kennedy (1986). It is possible that, given more material, H. (H.) howarthi may eventually be merely regarded as a larger, more coarsely ornamented, South African-Malagassy subspecies of H. (H.) marroti.
H. (H.) rejaudryi (de Grossouvre, 1894) (see Kennedy 1986, p. 78, pl. 7, figs 1-5; pl. 14, figs 6-7) is consistently much finer ribbed than H. (H.) howarthi.
H. (H.) preyi Kennedy & Summesberger (1999, p. 25, pl. 3, figs 1, 4) from the Late Campanian of the Gschliefgraben, Austria, is monotypical and crushed. It differs from H. (H.) howarthi in its finer ribbing and complete absence of conspicuous umbilical tubercles ofthe latter species. Differences with other species are summarized by Kennedy & Summesberger (1999, p. 25-26).
H. (H.) rarecostatum Khakimov (Khakimov in: Atabekian & Khakimov 1976, p. 88, pl. 10, fig. 5) is difficult to interpret. The figured holotype is similar to inflated variants ofH. (H.) howarthi and has similar strong ornamentation; given more material more precise differences may be determined.
H. (H.) minor Cobban & Kennedy (1993, p. 73, figs 4.14.28) from the Middle Campanian Wolfe City Sand in northeastern Texas lacks the strong umbilical tubercles of H. (H.) howarthi and has very dense, thread-like ribbing on the body chamber.
H. (H.) monju Matsumoto, 1982 (1982, p. 244, figs 1-2; 1984, p. 24, pl. 7, fig. 5; pl. 8, fig. 6) was compared by Matsumoto (1984, p. 25) to H. (H.) trangahyense Collignon; we regard the latter as a synonym ofH.(H.) howarthi. They overlap in some morphological features, but H. (H.) monju lacks the strong umbilical tuberculation of typical H. (H.) howarthi, as does H. (H.) fugen Matsumoto (1984, p. 25, pl. 8, fig. 5) H. cf. marroti (Coquand) described from Angola by Howarth (1965, p. 391, pl. 12, fig. 3; pl. 13, fig. 3) has lateral ornament very similar if not identical to that of H. (H.) howarthi.
ACKNOWLEDGEMENTS
Financial assistance to Klinger from the National Research Foundation, South Africa, and logistical support by W.E. Grulke is gratefully acknowledged, as is the assistance of staff of the iSimangaliso Wetland Park Authority (formerly the Greater St Lucia Wetlands Park Authorities) with access and transport on the St Lucia-False Bay lakes region of KwaZulu-Natal. Schlüter's type material was examined by Klinger in Bonn, Göttingen and Tubingen during the tenure of an Alexander von Humboldt Foundation scholarship in 1980. Kennedy thanks the staff of the Oxford University Museum of Natural History and the Department ofEarth Sciences, Oxford for logistical support. We thank M.B. Aguirre Urreta (Buenos Aires) for the casts of Hoplitoplacenticeras (H.) plasticum deposited in the collections of the South African Museum.
REFERENCES
ATABEKIAN, A.A. & KHAKIMOV, F.K. 1976 [Campanian and Maastrichtian ammonites in Central Asia]. Trudy Institut geologichesko Dushanbe. 146 pp. [In Russian] [ Links ].
BASSE, E. 1931. Monographie paléontologique du Crétacé de la province de Maintirano. Mémoires du Service des Mines de Madagascar 1931: 1-86, pls 1-13. [ Links ]
COBBAN, W.A. & KENNEDY, W.J. 1993. Middle Campanian ammonites and inoceramids from the Wolfe City Sand in northeastern Texas. Journal of Paleontology 67: 71-82. [ Links ]
COLLIGNON, M. 1970. Atlas des fossiles caractéristiques de Madagascar (Ammonites) XVI (Campanien moyen) & (Campanien supérieur). Tananarive: Service Géologique. iv + 82 pp. [ Links ]
COQUAND, 1859. Synopsis des animaux et des végétaux fossiles observes dans la formation crétacée du sud-ouest de la France. Bulletin de la Société Géologique de France (Ser. 2) 16: 945-1023. [ Links ]
DOUVILLÉ, H. 1890. Sur la classification des Cératites de la Craie. Bulletin de la Société Géologique de France (Ser. 3) 18: 275-292. [ Links ]
GROSSOUVRE, A. de, 1894. Recherches sur la craie supérieure: 2. Paléontologie: Les ammonites de la craie supérieure. Mémoires pour servir a l'explication de la Carte Géologique détaillée de la France [for 1893] 264 pp. [ Links ]
HASSAN, M.Y. 1971. New Molluscan fauna from the Maastrichtian of Kharga Oasis, South Western Desert of Egypt. Proceedings of the Egyptian Academy of Science 23: 65-75. [ Links ]
HOWARTH, M.K. 1965. Cretaceous ammonites and nautiloids from Angola. Bulletin of the British Museum (Natural History) (Geology) 10: 335-412. [ Links ]
HYATT, A., 1889. Genesis ofthe Arietidae. Smithsonian Contributions to Knowledge 673: xi+ 238 pp. [ Links ]
IMKELLER, H. 1901. Die Kreidebildungen und ihre Fauna am Stallauer Eck und Enzenauer Kopf bei Tölz. Palaeontographica A48: 1-64. [ Links ]
KAPLAN, U., KENNEDY, W.J. & ERNST, G. 1996. Stratigraphie und Ammonitenfaunen des Campan im südöstlichen Münsterland. Geologie und Paleontologie in Westfalen 43: 1-133. [ Links ]
KAPLAN, U., KENNEDY, W.J. & HISS, M. 2005. Stratigraphieund Ammonitenfaunen des Campan im nordwestlichen und zentralen Münsterland. Geologie und Paleontologie in Westfalen 64: 1-171. [ Links ]
KAPLAN, U., KENNEDY, W.J. & SCHEER, U. 2006. Ammoniten der Bottrop-Formation, Campanium, westliches Münsterland. Geologie und Paleontologie in Westfalen 67: 1-71. [ Links ]
KAYSER, E. 1924. Lehrbuch der Geologie. II. Geologische Formationskunde. 7th edn. viii + 675 pp. Ferdinand Enke: Stuttgart. [ Links ]
KENNEDY, W.J. 1986. Campanian and Maastrichtian ammonites from northern Aquitaine, France. Special Papers in Palaeontology 36: 5-145. [ Links ]
KENNEDY, W.J. & KAPLAN, U., 1997. Ammoniten aus dem Campan des Stemweder Berges, Dammer Oberkreidemulde, NW-Deutschland. Geologie und Paleontologie in Westfalen 50: 31-245. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 1973. In: KENNEDY, W.J., KAUFFMAN, E.G. & KLINGER, H.C. Upper Cretaceous invertebrate faunas from Durban, South Africa. Transactions of the Geological Society of South Africa 76: 95-111. [ Links ]
KENNEDY, W.J. & KLINGER, H.C. 1975. Cretaceous faunas from Zululand and Natal, South Africa. Introduction, stratigraphy. Bulletin of the British Museum (Natural History)(Geology) 25: 263-315. [ Links ]
KENNEDY, W.J. & SUMMESBERGER, H., 1999. New Late Campanian ammonites from the Gschliefgraben near Gmunden (Ultrahelvetic, Austria). Beitrage zur Paleontologie 24: 23-39. [ Links ]
KENNEDY, W.J. & WRIGHT, C.W. 1983. Ammonites polyopsis Dujardin, 1837 and the Cretaceous ammonite family Placenti-ceratidae Hyatt, 1900. Palaeontology 26: 855-873. [ Links ]
KHAKIMOV. F.K. In: ATABEKIAN, A.A. & KHAKIMOV, F.K. 1976 [Campanian and Maastrichtian ammonites in central Asia] Trudy Institut geologichesko Dushanbe 146 pp. [In Russian]
KLINGER, H.C. & KENNEDY, W.J. 1989. Cretaceous faunas from Zululand and Natal, South Africa. The ammonite family Placenticeratidae Hyatt, 1900; with comments on the systematic position of the genus Hypengonoceras Spath, 1924. Annals of the South African Museum 98: 241-08. [ Links ]
MACHALSKI, M., KENNEDY, W.J. & KIN, A. 2004. Early Late Campanian ammonite fauna from Busko Zdrój (Nida Trough, southern Poland). Acta Geologica Polonica 54: 447-471. [ Links ]
MATSUMOTO, T., 1982. Note on Hoplitoplacenticeras from Hokkaido. Proceedings of the Japanese Academy 58[B]: 249-252. [ Links ]
MATSUMOTO, T. 1984. Some ammonites from the Campanian (Upper Cretaceous) of northern Hokkaido. Palaeontological Society of Japan. Special Papers 27: v+1-32. [ Links ]
MEEK, F.B.1862. Description of new Cretaceous fossils collected by the North-Western Boundary Commission on Vancouver and Sucia islands. Proceedings ofthe Academy of Natural Sciences of Philadelphia 1861(10): 314-318. [ Links ]
MEEK, F.B. 1876. A report on the invertebrate Cretaceous and Tertiary fossils of the upper Missouri country. In: HAYDEN, F.V. Report of the United States Geological Survey of the Territories 9: lxiv + 1-629 pp. [ Links ]
MIKHAILOV, N.P. 1951. Verkhnemelovye ammonite yga evropeskoj chasti SSSR I ikh znachenie dlya zonal'noj stratigrafii. Trudy Instituta geologicheskikh nauk 129: 1-143. [ Links ]
ORBIGNY, A. D' 1840-1842 . Paleontologie frangaise: Terrains Crétacés. I. Céphalopodes. Paris: Victor Masson. [ Links ]
PAULCKE, W.W. 1907. Die Cephalopoden der oberen Kreide Südpatagoniens. Berichte der Naturforschenden Gesellschaft zu Freiburg i.B. 15: 167-248. [ Links ]
QUAAS, A. 1902. Beitrag zur Kenntnis der Fauna der obersten Kreidebildungen in der Libyschen Wüste (Overwegischichten und Blattertone). Palaeontographica 30: 153-336.
RICCARDI, A.C. 1988. The Cretaceous System of southern South America. The Geological Society of America Memoir 168: i-v+1-161. [ Links ]
SCHLÜTER, C. 1967. Beitrag zur Kenntniss der jüngsten Ammoneen Norddeutschlands. 36 pp. Bonn: A. Henry [ Links ]
SCHLÜTER, C. 1871-76. Cephalopoden der oberen deutschen Kreide. Palaeontographica 21: 1-24, pls 1-8 (1871); 21: 25-120, pls 9-35 (1872); 24: 1-144 (121-264) + 10, pls 36-55 (1876). [ Links ]
WRIGHT, C.W. 1996. Treatise on Invertebrate Paleontology. Part L, Mollusca 4: Cretaceous Ammonoidea. xx + 1-362 (with contributions by J.H. Calloman [sic] and M.K. Howarth). Lawrence, Kansas and Boulder, Colorado: Geological Society of America and University of Kansas. [ Links ]
ZITTEL, K. VON, 1884. Cephalopoda. p. 329-522. In: ZITTEL, K.A. VON Handbuch der Palaeontologie. Band 1, Abt. 2. Lief. 3. Oldenbourg: München & Leipzig. [ Links ]
Received 9 August 2011
Accepted 22 October 2011


{kind=link}
{kind=link}