










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkJournal of the Southern African Institute of Mining and Metallurgy
versión On-line ISSN 2411-9717
versión impresa ISSN 2225-6253
J. S. Afr. Inst. Min. Metall. vol.113 no.10 Johannesburg ene. 2013
CONFERENCE PAPERS
The use of change management and behaviour-based training in an improvement project aimed at creating organizational sustainability in a multicultural mining environment in Zambia
H.B. PrinslooI; J.O. ClaassenII
IUniversity of the Free State, HB Prinsloo Consulting Services
IIHB Prinsloo Consulting Services
ABSTRACT
Change management and training initiatives in remote rural settings in Africa are generally difficult to implement successfully due to factors such as varying levels of education, multiple nationalities and languages as well as cultures in one workplace.
The challenge has always been the ability to implement change management and training initiatives that will have an impact despite these differences. Essentially this requires that the change and training interventions employed are applicable and should work well with a wide range of individuals sitting in one room - in practice this can vary from semi-literate workers to engineers with PhDs.
In this paper we will share the theory behind behaviour-based change management and training and present a case study that reviews an actual supply chain improvement and sustainability implementation at a Zambian copper mine. The case study explains the results achieved as well as the interventions employed to achieve these results in a multidimensional organizational environment.
This paper is relevant to delegates who are faced with implementing changes or training in a similar mining environment.
Keywords: change management, behaviour-based training, learning style, P2P process.
Introduction - the problem
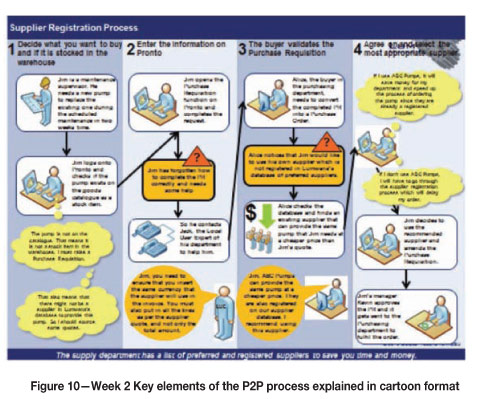
Shortly after transitioning from a construction phase to a production phase the client, Africa's largest copper mine (also producing uranium and cobalt), an opencast copper mine project in the North West Province of Zambia, 220 km west of the Zambian Copperbelt and 65 km west of the town of Solwezi, found that their Procure to Pay (P2P) process was not necessarily coping with or ideal for a remote rural mining operation in production phase. Role players in the (P2P) process were from different nationalities and cultural backgrounds - in order of prevalence: Zambian, Australian, South African, Canadian, UK/British, Ukrainian, Kenyan, and Ghanaian.
The intervention
An international consulting firm was contracted to solve the problem by improving and redesigning the P2P process (Figure 1).
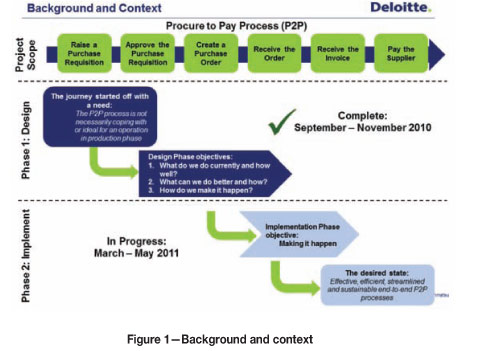
Once the process redesign phase was completed, the consulting firm started implementing the improved P2P process, and soon found that traditional classroom-based process presentation training was not effective in causing the role players in the P2P process to change their behaviour. It was clear that a different approach was needed. The consulting firm contracted a change management consultant - a behaviour-based training specialist - to design and implement a programme that would 'make it happen'. The consultant opted for two one-week on-site interventions.
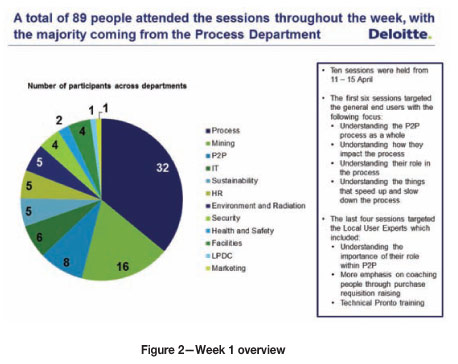
During the first week, 10 sessions were held to help the participants experience what the problems with the P2P process were and how they were affecting the different role players in the process (Figure 2). Various behaviour-based games (Figure 3) were played with the participants, each game illustrating some critical behavioural component that is problematic for the P2P process.

The most notable behaviour-based game that was administered was a 'Snakes and Ladders' game (Figure 4) designed around highlighting the positive and negative behaviours that form part of the P2P process. By simply playing the game, delegates learnt the P2P process without ever looking at a process flow (Figure 5).

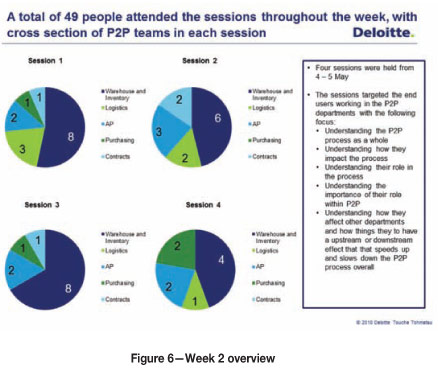
Approximately four weeks after the first week, interventions were conducted in a second and final week (Figure 6).
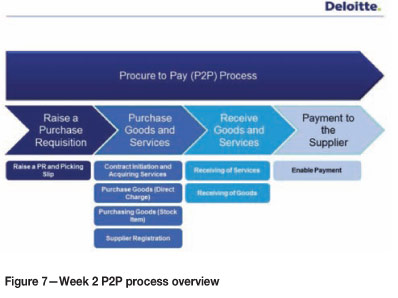
The interventions during the second week were aimed at a detailed understanding of the P2P process, which was broken down into four main sub-processes (Figure 7).
The training manual provided:
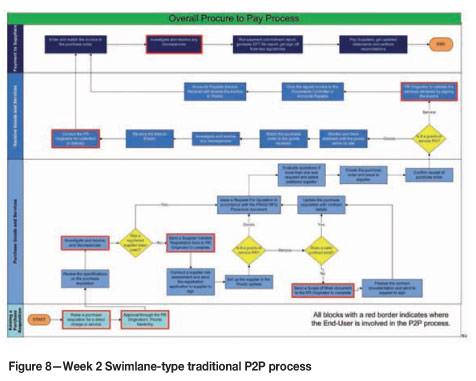
> Gave both a traditional process flow and cartoons of certain critical process elements (Figure 8)
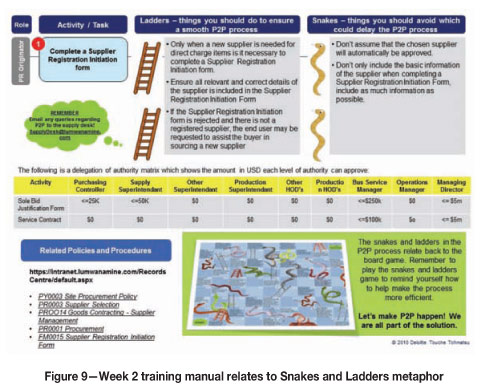
> Related to the metaphor of the Snakes and Ladders game that participants received during the first week (Figure 9).
At the end of the process training workshop, participants were divided into four teams, and allowed to draw a process and a theme card (Figure 11). In order to embed their process learning and make it practical, each team then had to act out the process on their process card, based on the story on their theme card. Participants voted for the winning performance, for which the winning team received a small prize.
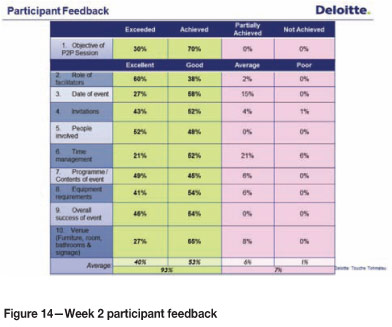
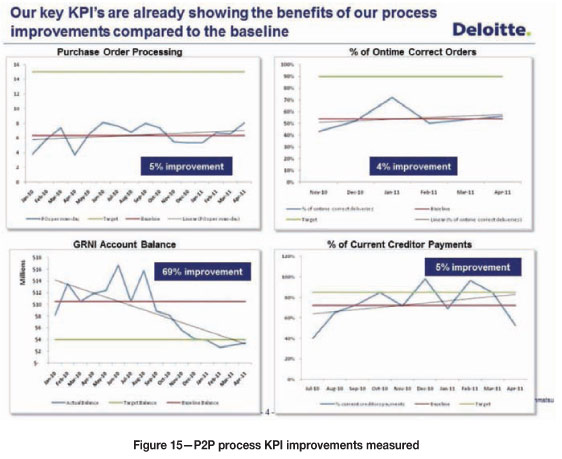
Behavioural measurements taken during the second week showed that attending one session only provided tangible results.
Why does it work?
People often comment beforehand that the interventions sound rather infantile, and say they doubt whether adults on their mine would be willing to participate. In reality, behaviour-based learning is fun, and totally immersive. People hardly realize they are learning and, as they experience the benefits of the training, readily participate in all the activities.
In order to understand why this type of behaviour-based training is such a successful method for training adults in a rural and multi cultural mining environment, we will explore recent research conducted in the fields of andragogy, the critical period, plasticity as well as learning styles.
Andragogy
Malcolm Knowles, often referred to as the 'father of adult education', is best known for further developing the concept of andragogy in the late 1960s. The theory of andragogy is defined as 'the art and science of helping adults learn' (Knowles, 1980, p. 43). His work helped professionalize the field of adult education and distinguished it from other areas of education (Sherow, 2006, p. 6). Based on Knowles' theory of andragogy, educating adults is perceived to be different from educating young adults (Knowles, 1980, p. 43). Abraham Maslow categorizes human needs into hierarchical levels: physiological needs; safety needs; needs for love, affection, and to belong; needs for esteem; and needs for self-actualization. These levels, to a large extent, influence the motivation of adults to learn (Boeree, 2006, p. 1).
The key andragogy assumptions about the adult learner are:
> Need to know purpose-Adults are problem-centred and interested in the immediate application of knowledge. Therefore adults expect learning to be useful and to address their felt needs. They need to know why they should learn something and how it will benefit them. The trainer should help adult students understand how what they will learn will be of use to them in the future
> Self-concept of responsibility-Unlike young learners, whose learning is future-directed and relies largely on others for direction during the learning process, adults are self-directed, with an independent self-concept and an ability to direct their own learning with the information at hand. Adults resent and resist situations in which they feel others are imposing learning content on them
> Amount and diversity of experience-Adults have life experiences to contribute to the learning process, and generally the topic of learning should be related to the learner's past experiences in order to have the greatest impact. Adults want to use what they know and want to be acknowledged for having that prior knowledge and relevant experience
> Real-life applicability-Adult learners must learn real-life facts or situations. If they cannot use what they are being taught then they are not benefiting from the information. They need to use information to learn a skill, to be more knowledgeable, or to teach others. With this they will be able to get a better job, get a promotion, or have the satisfaction of passing information on to others so that they, in turn, can use the information. If lots of information is taught, it needs to be layered - basic information first, followed by exercises and experiences that can be used and applied in real-life practical situations. The aim with layering is to set cognitive hooks in the mind so that you can identify issues and recognize critical information in practical, life-like situations
> Life- / task- /problem-centred-Adults want to see how what they are learning will apply to a task they need to perform or to a problem they need to solve. Just like real-life applicability, adults want information that will tie into their lives. Adults learn by doing, and retain knowledge and skills if they have immediate and repeated opportunities to practise what they have learnt. They also want tasks and problems to conquer. They will not get anything out of learning if they float along and are not challenged > Internal motivation is strongest--While adult learners may respond to external motivators like money, internal priorities are more important. Incentives such as increased job satisfaction, self-esteem, and quality of life are important in giving adults a reason to learn. Adults have goals and ambitions and use this motivation to push forward to accomplish this. Adults will not learn until they are ready and motivated to learn, and often require help to overcome inhibitions, behaviours, and beliefs about learning, their abilities, who they are, and how they will be able to apply the learning (Merriam, 2001, p. 3; Imel, 1989, p. 1; Conner, 2004, p. 12; Smith, 2002, p. 2).
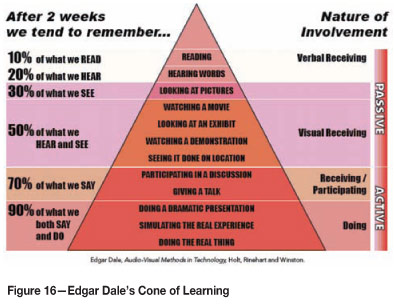
Although often criticized in the past as unscientific conjecture (Atherton, 2011, p. 1), Dale's Cone of Learning (1969) seems to summarize a lot of what adult learning is (Figure 16).


In essence, the active and practical nature of behaviour-based training is a significant determinant of its efficacy when training adults. However, the following recent research in the fields of cognitive psychology and neurology should also be noted and considered.
The Critical Period
The Critical Period hypothesis was originally proposed in the neurolinguistic literature by Penfield and Roberts (1959) and vigorously followed up by Lenneberg (1967) in his seminal book, Biological Foundations of Language. Lenneberg stated that language acquisition was an example of biologically constrained learning, and that it was normally acquired during a critical period, beginning early in life and ending at puberty. He also suggested a neural mechanism for this developmental change: he hypothesized that the critical period for language acquisition ended with the establishment of cortical lateralization of function, as the brain reached mature organization in late puberty.
As children start to mature, around the age of 20, cognitive plasticity decreases and the ability to learn some complex skills declines. Yun, Bazar, and Lee (2004) hypothesize that the advent of loss of plasticity in the brain relates to sexual maturation:
'Cognitive plasticity, a developmental trait that promotes acquisition of complex skills such as language or playing musical instruments, diminishes substantially during puberty. The loss of plasticity has been attributed to surge of sex steroids during adolescence, but the phenomenon remains poorly understood (Yun, Bazar, and Lee, 2004, p. 939).
Although loss of plasticity is still not fully understood, various studies have proven that a variety of neurobiological variables start declining relatively early in adulthood. Among the variables that have been found to exhibit nearly continuous age-related declines beginning when adults are in their twenties are measures of regional brain volume (Allen et al, 2005; Fotenos et al., 2005; Kruggel, 2006; Pieperhoff et al., 2008; Sowell et al., 2003), myelin integrity (Hsu, Leemans et al., 2008; Sullivan and Pfefferbaum, 2006), cortical thickness (Magnotta et al., 1999; Salat et al., 2004), serotonin receptor binding (Sheline et al., 2002), striatal dopamine binding (Erixon-Lindroth et al., 2005; Volkow et al., 2000), accumulation of neurofibrillary tangles (Del Tredici and Braak, 2008), and concentrations of various brain metabolites (Kadota et al., 2001). From an educational point of view, various sources also find diminished average achievement of older learners (Flege, Yeni-Komshian, and Liu, 1999; Stevens, 1999).
The second group of factors is the changes in cognition that occur with ageing. Although critical periods have not been posited in most cognitive domains, there are nonetheless age-related changes in cognitive processing. Some age-related changes in cognitive processes relevant to language learning are the decreased ability to learn paired associates (Salthouse, 1992), the increased difficulty with encoding new information (Craik and Jennings, 1992; Park et al., 2002; Rabinowitz, Craik, and Ackerman, 1982), and the reduced accuracy in recalling detail as opposed to gist (Hultsch and Dixon, 1990). Kemper (1992) pointed out that older adults' second-language proficiency, like their first-language proficiency, could also be affected by such factors as working memory capacity, cognitive processing speed, and attention. These factors all decline with age, and the decline is documented across the human lifespan. Such a reduction in cognitive resources would surely affect the ability to learn a new language. Older learners would find the task more difficult than younger ones, although no critical period would be involved (Hakuta, Bialystok, and Wiley, 2003: 6, p. 32).
Learners exposed to a language in adulthood show, on average, a lowered level of performance in many aspects of the language, though individual variation also increases with age (Johnson and Newport, 1989), to the extent that some individuals may approach the proficiency of early learners (Birdsong, 1992). These effects have been shown for both first and second languages, and for measures of proficiency including degree of accent, production, and comprehension of morphology and syntax, grammaticality judgements for morphology and syntax, and syntactic processing speed and accuracy (Newport, 2002, p. 738).
Age of exposure also affects the way language is represented in the brain, with similarities between behavioural and neural effects. PET (positron emission topography), FMRI (functional magnetic resonance imaging), and ERP (event-related potential) studies all show strong left hemisphere activation for processing the native language in bilinguals as well as monolinguals. However, when second languages are learned after the age of seven, the regions and patterns of activation are partially or completely nonoverlapping with those for the native language (Newport, 2002, p. 738).
Neural organization for late-learned languages is less lateralized and, like proficiency itself, displays a high degree of variability from individual to individual (Perani et al. 1996; Weber-Fox and Neville, 1996; Kim et al., 1997). This does not mean that adults cannot learn new skills, but the ability to learn declines and the brain becomes more selective about what it 'learns' and retains. The adult learner requires motivation that something is important to learn in order to be able to learn something new. As Michael Merzenich notes:
'Rather marvellously, the older brain only permits change when it judges that change to be important, rewarding, or goodfor it (Bruno, Merzenich, and Nudo, 2012, p. 4).
In their study of emigrant second-language learners, they found that age (of arrival) was the better measure of acquiring native language proficiency in English. However, the correlations between the attitudinal variables that they tested for (identification, self-consciousness, and motivation) also showed significant statistical correlations (Johnson and Newport, 1989, p. 84).
Generally it was believed that the loss of plasticity in the adult brain was responsible for the low level of recovery of function after damage to the nervous system. Experiments on monkeys and humans have generated a paradigm shift away from the (total) loss of plasticity theory, and conclusive proof has been found that not only is there a surprising amount of residual plasticity in the adult brain, but sensory inputs from one sense can quite literally substitute for another sense. These findings have invalidated long held theoretical assumptions about sensory function, such as 'Muller's law of specific nerve energies' (a given neuron in the brain can signal only a particular sensation) and 'place coding' (the experience evoked depends exclusively on the location of the excited neuron, not on the pattern of firing or the context in which it fires) (Ramcharan, 2005, p. 368).
Research into sensory substitution in the complete absence or the deterioration of a cognitive sense (like vision) shows that plasticity does exist after the critical period and that remapping and cross-modal recruitment of areas in the adult brain does take place after, for instance, the loss or severe deterioration of vision. Functional imaging of blind patients showed a cross-modal recruitment of the occipital cortex (generally used for processing of vision) during perceptual tasks such as braille reading, tactile perception, tactual object recognition, sound localization, and sound discrimination. This research finding shows that blind people can use their occipital lobe, generally used for vision, to perceive objects through the use of other sensory modalities, most often the tactile or auditory modality (Renier and de Volder, 2005).
Although motivation is a very significant determinant of the older brain successfully learning something new, Bieszczad and Weinberger (2010, pp. 229, 235), who researched the relationship between the strength of learning and the amount of cortical plasticity, found that the use of learning strategy or learning style actually trumps motivational level when learning-induced auditory cortical plasticity is measured. They trained two groups of rats (a highly motivated and a moderately motivated group) to solve an auditory-cued task to different degrees of correct performance and evaluated the amount of learning-related plasticity in the primary auditory cortex of the individual rats. This finding seems to indicate that learning strategy can be a critical factor in the development of learning-related cortical plasticity. Indeed, learning style and strategy can be more important than the level of learning.
The second great phase of plasticity
According to Merzenich and Sooy (2006, p. 2), in the second great phase of plasticity, plasticity is regulated as a function of behavioural context. The context-dependent (attention, brightness, reward or its expectation, positive surprise, punishment or threat) release of the neurotransmitters acetyl-choline, dopamine, noradrenaline, and serotonin (among others) contribute importantly to this regulation.
Controlled by brain processes that assure that it is primarily for the brain's benefit (i.e. correlated with goal achievement, expectation of reward, expectation of punishment, positive novelty), adult plasticity enables the specialization of the frontal lobe processing machinery necessary for establishing the thousands of skills and abilities that define each individual's operational capacities. While this phase of plasticity is in place to the end of life, its operational status degrades in older age. Given our understanding of brain plasticity processes, strategies designed to stabilize functionality or to reverse normal age-related losses of memory and other cognitive abilities, to address the progressive and ultimately catastrophic losses expressed in psychotic illnesses, and to address the development impairments expressed by the abnormally maturing brain of language, reading and cognitively impaired children have been developed and applied in human populations. This has led to two paradigm shifts. Firstly, that there is a tremendous latent plasticity even in the adult brain, and secondly, that the brain should be thought of, not as a hierarchy of organized autonomous modules, but rather a set of complex interacting networks that are in a state of dynamic equilibrium with the brain's environment (Ramcharan, 2005, p. 372).
Adult brain plasticity, although possible, remains more restricted in scope than during early and pre-pubescent development. There are certain conditions under which circuit rewiring may be facilitated in the mature brain. At a cellular and molecular level, adult plasticity is actively limited by the brain. Some of these 'brakes' are structural, such as perineuronal nets or myelin, which inhibit neurite outgrowth. Others are functional, acting directly upon excitatory/inhibitory balance within local circuits. Plasticity in adulthood can be induced either by lifting these brakes through invasive interventions or by exploiting endogenous permissive factors, such as neuromodulators. Using the amblyopic visual system as a model, Bavelier et al. (2010, p. 14694), enabled the genetic, pharmacological, and environmental removal of brakes in plasticity to enable the remarkable recovery of vision in adult rodents.
Once induced, synaptic rewiring appears to be executed by the action of extracellular proteases (Mataga et al., 2002; Oray et al., 2004), which induce dendritic spine motility and pruning prior to regrowth. Notably, these effects proceed in a laminar sequence (Oray et al., 2004) consistent with the progression of plasticity through the thalamocortical circuit (Trachtenberg et al., 2000). Likewise in the barrel cortex, a brief sensitive period for whisker receptive field tuning emerges concurrent with an increase in experience-dependent spine motility (Lendvai et al., 2000; Stern et al., 2001). Such abrupt and transient circuit reconfiguration is eventually recalibrated by homeostatic processes (Pozo and Goda, 2010), involving cell-intrinsic transcription factors (Greer and Greenberg, 2008; Chang et al., 2010) or surrounding glia-derived factors such as TNFa or the complement cascade (Kaneko et al., 2008; Stevens et al., 2007). It is notable in this context that one of the first successful approaches to re-introduce juvenile plasticity in the adult visual cortex was the direct transplantation of immature astrocytes (Müller and Best, 1989).
There is no doubt that humans demonstrate marked learning as a result of practice even in adulthood; yet adult learning appears qualitatively and quantitatively different when compared to what learning children may exhibit. It is effortful, often quite narrow in its scope, and most of the time incomplete (Newport et al., 2001).
One rather drastic example of adult plasticity in the case of amblyopia comes from 'experiments of nature' whereby amblyopic patients have lost vision in the 'good' eye. Under these conditions, visual acuity in the amblyopic eye sometimes spontaneously improves (Vereecken and Brabant, 1984; El Mallah et al., 2000; Rahi et al., 2002). These few reports are consistent with the notion that the connections from the amblyopic eye may be weakened, inhibited, or unattended rather than destroyed. Loss of the fellow eye would allow these existing connections to be reactivated. This could be the result of unmasking (Restani et al., 2009) or higher brain areas learning to attend to the previously inhibited signals from the amblyopic eye.
Learning can be divided into declarative memory, the conscious recall of people, places and events, and implicit memory, the unconscious acquisition of habits and skills with practice (Gilbert, Li, and Piech, 2012, p. 2743). Perceptual learning is a form of implicit memory, involving improvement in sensory discrimination or detection by repeated exposure to sensory stimuli. Whereas declarative memory is localized to the temporal lobe, perceptual learning involves functional changes that are widespread throughout the cortex and affects cortical function throughout life. The findings leading to this idea were unexpected. The classic Nobel Prize-winning studies of David Hubel and Torsten Weisel showed how the proper maturation of the developing visual cortex is critically dependent upon visual information received from the eyes. In what would today be considered highly unethical experiments, Hubel and Weisel (1959, 1960, 1961, 1962, and 2005) sewed shut one eye of newborn kittens and spider monkeys. They found that this monocular deprivation had dramatic effects on the visual part of the brain: the columns of cortical tissue that normally receive inputs from the closed eye failed to develop, while those that receive inputs from the other eye were significantly enlarged. The kittens also failed to develop visual cortical areas which normally receive inputs from both eyes and, as a result, did not have binocular vision.
But subsequently, evidence has accumulated that many other properties are subject to experience even in adulthood, and the mutability of these properties can account for the psychophysical changes occurring during perceptual learning, as well as functionally adaptive changes following central nervous system (CNS) lesions and damage (Gilbert, Li, and Piech, 2012, p. 2743).
In their research Gilbert, Li and Piech (2012, p. 2750) found that the visual cortex retains the capacity for experience-dependent plasticity into adulthood. This plasticity serves to encode the information acquired during perceptual learning, and can be used for the purpose of functional recovery following CNS lesions. Learning can involve global changes in the representation of shapes, shifting to different areas along the visual pathway. The areas involved can extend as far back as primary visual cortex. The way in which the functional properties are dynamically expressed in a task-dependent fashion implies that each cortical area acts as an adaptive processor, performing different calculations according to the immediate perceptual demands. The manifestation of the functional changes associated with perceptual learning suggests a circuit mechanism that involves an interaction between feedback connections and local cortical circuits.
The proposal that brain plasticity and learning are fostered by environmental factors is far from new (Greenough, et al., 1987). As early as the 1960s, Bennett et al. (1964) noted that adult rats housed in enriched cages had a far greater cortical weight than those housed in individual, standard laboratory cages. Two seemingly opposite manipulations in adult rats illustrate the power of environment. Oddly, amblyopic rats subjected to complete visual deprivation by dark exposure for ten days recover significant vision once allowed to see binocularly (He et al., 2007).
Another factor that has a positive effect on plasticity in the brain is aerobic exercise, with individuals who normally exercise outperforming those who do not on tasks as varied as dual-task performance, executive attention, or distractor rejection (for recent reviews, see Colcombe and Kramer, 2003; Kramer and Erickson, 2007; Hillman et al., 2008). In addition to its well documented impact on neurogenesis in animal models (Kempermann et al., 2000; Nithianantharajah and Hannan, 2006), aerobic fitness also leads to neuroanatomical and neurophysiological changes in older adults, including increased grey matter volume in the prefrontal and temporal areas (Colcombe and Kramer, 2003) and functional brain activity in a variety of areas such as superior parietal areas and the anterior cingulate cortex (Colcombe et al., 2004). Whether aerobic exercise can enhance brain plasticity in healthy, young adults unfortunately remains undocumented (Bavelier et al., 2010, p. 14694).
Another type of enriched environment extensively studied in humans, especially in the case of vision, is perceptual learning and, more recently, immersion in video games.
During perceptual learning using only their amblyopic eye, patients are required to practise a variety of visual tasks. A review of the extant studies (almost 200 amblyopic subjects distributed over 14 papers) reveals that such practice results in a long-lasting improvement in performance in amblyopic eyes (Levi and Li, 2009a). Similarly, recent studies indicate that enhancements after action video game play are also due to observers being better able to select and use the most reliable information for the task (Li et al., 2009a). Yet, unlike perceptual learning, whereby the observer typically learns the best template just for the trained task, this work suggests that action gamers learn to find the best template 'on the fly' as they are faced with new visual stimuli and new environments (Green et al., 2010b). Accordingly, fast-paced, action-packed games have already been documented to have a potent, positive impact on an array of skills, including perception, visuo-motor coordination, spatial cognition, attention, and decision-making to cite a few, illustrating the powerful effect of action game play in reshaping the adult brain (Gagnon, 1985; Dorval and Pe'pin, 1986; Greenfield et al., 1994; De Lisi and Wolford, 2002; Green and Bavelier, 2006; Quaiser-Pohl et al., 2006; Greenfield, 2009; Li et al., 2009b).
Action video game play may therefore improve the efficiency of probabilistic inference in neural circuits, which in turn would provide a mechanistic explanation for the broad transfer such training engenders (Green et al., 2010b). These plastic changes have been shown to be long-lasting, with beneficial effects noted 6 months to 2 years after the end of intervention (Feng et al., 2007; Li et al.,2009b).
As with enriched cages, the factors that conspire to induce brain plasticity within the action game experience remain to be systematically assessed. As a training paradigm, gaming differs from more standard methods on several dimensions.
Firstly, gaming tends to be more varied in the skills it requires than standard training, which typically focuses on just one aspect of performance, as exemplified in the field of perceptual learning. Such variation during training enhances transfer across tasks (Schmidt and Bjork, 1992; Kornell and Bjork, 2008).
Secondly, unlike standard training paradigms, gaming is an activity that is highly engrossing and also extremely rewarding. Reward, and the drive to perform as many correct responses per unit of time (Dye et al., 2009a), is likely to engage dopamine and possibly opiates and other neuromodu-lators. The relationship between game play and the reward system remains, however, an understudied domain. An early PET study indicated a large release of striatal dopamine during the play of a toy video game (Koepp et al., 1998), but significant experimental bias can affect the estimated size of the effect, calling for further replications (Egerton et al., 2009). This has significant implications in terms of simulators, simulations, and reward-based games.
Thirdly, action games constantly require divided attention and its efficient reallocation as task demands change, most likely engaging neuromodulatory systems such as acetyl-choline and dopamine (Rueda et al., 2005; Dye et al., 2009b), which are also known to enhance sensory processing and brain plasticity (Kilgard and Merzenich, 1998; Bao et al., 2001; Goard and Dan, 2009).
Finally, gaming is also associated with 'flow' or the sense that one is able to meet the challenges of one's environment with appropriate skills (Csikszentimihalyi, 1990). Flow is also characterized by a deep sense of enjoyment, which goes beyond satisfying a need and rather occurs when a person achieves something unexpected that has a sense of novelty. Playing entertainment video games is likely to increase flow. Accordingly, older adult stroke survivors report that participating in a virtual reality rehabilitation programme leads to increased involvement, enjoyment, and a sense of control over the environment (Farrow and Reid, 2004). The physiological bases of flow remain largely unknown, yet it would seem key for further studies to understand how it engages neuromodulatory systems as well as the regulation of the autonomous nervous system (Tang et al., 2007; Lutz et al.,2008).
The biology of the brain is heavily invested in the optimal timing and duration of plasticity, having evolved numerous molecular checks and balances to ensure an adaptable yet efficient organism. As we gain better knowledge of these mechanisms, we can assess the extent to which a sensitive period can be safely and noninvasively recapitulated by behavioural training such as perceptual learning (Levi and Li, 2009b), video game play (Green et al., 2010a), targeted pharmacological manipulation (Fluoxetine), or even brain stimulation to alter E/I balance directly (Fregni and Pascual-Leone, 2007; Thompson et al., 2008). A main challenge will of course be the ability to foster plasticity and relearning in one domain, while leaving unaltered cortical functions in other domains (Bavelier et al., 2010, p. 14698).
In terms of maturational decline, the tactile modality is the most abundant in terms of number and range of nerve endings and receptors, and generally does not decline as fast as, for instance, sight or hearing. Therefore a tactile-kinaes-thetic learning strategy would, on average, have the greatest propensity for a successful learning outcome in adults. In addition, tactile-kinaesthetic learning also fosters the creation of muscle memory (also referred to as motor learning), which is a form of procedural memory that involves consolidating a specific motor task into memory through repetition.
When a movement is repeated over time, a long-term muscle memory is created for that task, eventually allowing it to be performed without conscious effort. This process decreases the need for attention and creates maximum efficiency within the motor and memory systems. Examples of muscle memory are found in many everyday activities that become automatic and improve with practice, such as riding a bicycle, typing on a keyboard, typing in a PIN, playing a melody or phrase on a musical instrument, and playing video games (Krakauer and Shadmehr, 2006, p. 58).
Learning styles
Brown (2000) defines learning styles as the manner in which individuals perceive and process information in learning situations. He argues that learning style preference is one aspect of learning style and refers to the choice of one learning situation or condition over another.
Mac Keracher (2004, p. 71) defines learning style as the characteristic cognitive, affective, social, and physiological behaviours that serve as relatively stable indicators of how learners perceive, interact with, and respond to the learning environment.
Generally, there are three main learning styles: visual, auditory, and kinaesthetic (also sometimes referred to as tactile-kinaesthetic).
> Visual learners think in pictures and learn best in visual images. They depend on the instructor's or facilitator's non-verbal cues such as body language to help with understanding. Sometimes, visual learners favour sitting in the front of the classroom
> Auditory learners discover information through listening and interpreting information by means of pitch, emphasis, and speed. These individuals gain knowledge from reading out loud in the classroom, and may not have a full understanding of information that is written
> Kinaesthetic learners learn best with an active 'hands-on' approach. These learners favour interaction with the physical world. Most of the time kinaesthetic learners have a difficult time staying on target and can become unfocused effortlessly (Gilakjani and Ahmadi, 2011, p. 470).
According to Dunn and Dunn (1978), only 20-30% of school-age children appear to be auditory learners, 40% are visual learners, and 30-40% are tactile/kinaesthetic or visual/tactile learners.
Learning style is important for many reasons, the three most pertinent being:
(1) People's individual learning styles vary because everyone is naturally different
(2) Learning styles offer the opportunity to teach using a wide range of methods in an effective way. Sticking to just one model unthinkingly will create a monotonous learning environment and not everyone will enjoy the lesson. In other words, learning and teaching will be just words and not rooted in reality
(3) If we really recognize the groups we are called to, we can manage many things in communication and education a lot better. Being aware of our audiences' learning styles, psychological qualities, and motivational differences will help us regulate our lessons appropriately and according to the conditions (McCarthy, 1982; Felder and Silverman, 1988; Coffield, 2004; Gilakjani and Ahmadi, 2011).
Learning style has an important role in the lives of individuals. If an individual knows his/her learning style he or she will be able to integrate it in the learning process and learn easier, faster, and more successfully. Another benefit of knowing one's learning style is that it will help the individual to become a more effective problem solver. The more successful the individual is at solving the problems that they face, the more control they will take over their own life(Biggs, 2001).
Many studies conducted on late pubescent and early adult students have recorded that the majority of students prefer the tactile-kinaesthetic learning style: These include Chinese university students studying in the USA (Reid, 1987), Chinese (PRC) university students (Melton, 1990), Chinese (Taiwan) university students (Jones, 1997), non-Chinese university students that emigrated into the USA (Rossi-Le, 1995), and Japanese university students (Hyland, 1993). In an analysis by Peacock (2001) the results of various studies internationally show that students prefer kinaesthetic learning styles above others, whereas the teaching methods are mostly designed to suit auditory students. Mounting neurological and neuropsychological studies concur with these learning style studies - consistently suggesting that a tactile-kinaesthetic learning style might be most beneficial to adult learners.
Acknowledgement
The authors gratefully acknowledge Ann Farndell, Anglo American Internal Communications Manager, South Africa, who asked the question that prompted the writing of this article.
References
Allen, J.S., Bruss, J., Brown, C.K., and Damasio, H. 2005. Normal neuroanatomical variation due to age: the major lobes and a parcellation of the temporal region. Neurobiology of Aging, vol. 26, no. 9. pp. 1245-60. [ Links ]
Atherton, J. S. 2011. Learning and Teaching; Enactive, iconic, and symbolic levels of representation. http://www.learningandteaching.info/learning/dale-bruner.htm [ Links ]
Bao, S., Chan, V.T. and Merzenich, M.M. 2001. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature, vol. 412. pp. 79-83. [ Links ]
Bavelier, D., Levi, D.M., Lí, R.W., Dan, Y., and Hensch, T.K. 2010. Removing brakes on adult brain plasticity: from molecular to behavioral interventions. Journal of Neuroscience, vol. 30, no. 45. pp. 14964-71. [ Links ]
Bennett, E.L., Diamond, M.C., Krech, D., and Rosenzweig, M.R. 1964. Chemical and anatomical plasticity of brain: changes in brain through experience, demanded by learning theories, are found in experiments with rats. Science, vol. 146. pp. 610-619. [ Links ]
Bieszczad, K.M. and Weinberger, N.M. 2010. Learning strategy trumps motivational level in determining learning-induced cortical plasticity. Neurobiology of Learning and Memory, vol. 93. pp. 229-239. [ Links ]
Biggs, J. 2001. Enhancing learning: a matter of style or approach? Perspectives on Thinking, Learning and Cognitive Styles. Sternberg, R.J. and Zhang, L.F. (eds). Lawrence Erlbaum and Associates, Mahwah, NJ. [ Links ]
Birdsong, D. 1992. Ultimate attainment in second language acquisition. Language, vol. 68. pp. 238-243. [ Links ]
Birren, J.E. and Schaie, K.W. (eds). 1990. Handbook of the Psychology of Aging. 3rd edn. Academic Press, San Diego, CA. [ Links ]
Boeree, C.G. 2006. Abraham Maslow. http://webspace.ship.edu/cgboer/maslow.html [ Links ]
Brown, H.D. 2000. Principles of Language Teaching and Learning. 4th edn. Longman, White Plains, NY. [ Links ]
Bruno, R., Merzenich, M., and Nudo, R. 2012, The fantastic plastic brain: recent research is proving the adult human brain is remarkably malleable and capable of new feats -- even in the last decades of life. It just needs the right prodding. http://www.kavlifoundation.org/science-spotlights/columbia-kibs-fantastic-plastic-brain#Aging [ Links ]
Chang, M.C., Park, J.M., Pelkey, K.A., Grabenstatter, H.L., XU, D., Linden, D.J., Sutula, T.P., Mcbain, C.J., and Worley, P.F. 2010. NARP regulates homeostatic scaling of excitatory synapses on parvalbumin-expressing interneurons. Nature Neuroscience, vol. 13, no. 9. pp. 1090-1097. [ Links ]
Coffield, F. 2004. Learning styles and pedagogy in post-16 learning: a systematic and critical review. Learning and Skills Research Centre, London. [ Links ]
Colcombe, S. and Kramer, A.F. 2003. Fitness effects on the cognitive function of older adults: a meta-analytic study. Psychological Science, vol. 14. pp. 125-130. [ Links ]
Colcombe, S.J., Kramer, A.F., Erickson, K.I., Scalf, P., Mcauley, E., Cohen, N.J., Webb, A., Jerome, G.J., Marquez, D.X., and Elavsky, S. 2004. Cardiovascular fitness, cortical plasticity, and aging. Proceedings of the US National Academy of Sciences, vol. 101. pp. 3316-3321. [ Links ]
Conner, M.L. 2004. Learn More Now: 10 Simple Steps to Learning Better, Smarter, and Faster. John Wiley & Sons, Hoboken, NJ. [ Links ]
Craik, F.I.M. and Jennings, J.M. 1992. Human Memory. The Handbook of Aging and Cognition. Craik, F.I.M. and Salthouse, T.A. (eds.). Erlbaum, Hillsdale, NJ. [ Links ]
Craik, F.I.M. and Salthouse, T.A. (eds). 1992. The Handbook of Aging and Cognition. Erlbaum, Hillsdale, NJ. [ Links ]
Csikszentimihalyi, M. 1990. Flow: the psychology of optimal experience. Harper Collins, New York, NY. [ Links ]
Dale, E. 1969. Audiovisual Methods in Teaching. 3rd edn. Dryden Press, New York. [ Links ]
Delisi, R. and Wolford, J.L. 2002. Improving children's mental rotation accuracy with computer game playing. Journal of Genetic Psychology, vol. 163. pp. 272-282. [ Links ]
Del tredici, K. and Braak, H. 2008. Neurofibrillary changes of the Alzheimer type in very elderly individuals: neither inevitable nor benign. Commentary on 'No disease in the brain of a 115 year-old woman'. Neurobiology of Aging, vol. 29. pp. 1133-1136. [ Links ]
Dorval, M. and Pe'pin, M. 1986. Effect of playing a video game on a measure of spatial visualization. Journal of Perceptual and Motor Skills, vol. 62. pp. 159-162. [ Links ]
Dunn, R. and Dunn, K. 1978. Teaching students through their individual Learning Styles: a Practical Approach. Prentice Hall, Reston, VA. [ Links ]
Dupoux, E. (ed.). 2001. Language, Brain and Cognitive Development: Essays in Honor of Jacques Mehler. MIT Press, Boston, MA. [ Links ]
Dye, M.W., Green, C.S., and Bavelier, D. 2009a. Increasing speed of processing with action video games. Current Directions in Psychological Science, vol. 18. pp. 321-326. [ Links ]
Dye, M.W., Green, C.S., and Bavelier, D. 2009b. The development of attention skills in action video game players. Neuropsychologia, vol. 47. pp. 1780-1789. [ Links ]
Egerton, A., Mehta, M.A., Montgomery, A.J., Lappin, J.M., Howes, O.D., Reeves, S.J., Cunningham, V.J., and Grasby, P.M. 2009. The dopaminergic basis of human behaviors: a review of molecular imaging studies. Neuroscience and Biobehavioral Reviews, vol. 33. pp. 1109-1132. [ Links ]
El mallah, M.K., Chakravarthy, U., and Hart, P.M. 2000. Amblyopia: is visual loss permanent? British Journal of Ophthalmology, vol. 84, no. 9. pp. 952-956. [ Links ]
Erixon-lindroth, N., Farde, L., Robins-Whalin, T.B., Sovago, J., Halldin, C., and Backman, L. 2005. The role of the striatal dopamine transporter in cognitive aging. Psychiatry Research: Neuroimaging, vol. 138. pp. 1-12. [ Links ]
Farrow, S. and Reid, D. 2004. Stroke survivors' perceptions of a leisure-based virtual reality program. Technology and Disability, vol. 16, no. 2. pp. 69-81. [ Links ]
Felder, R.M. and Silverman, L.K. 1988. Learning and teaching styles in engineering education. Engineering Education, vol. 78. pp. 674-681. [ Links ]
Feng, J., Spence, I., and Pratt, J. 2007. Playing an action videogame reduces gender differences in spatial cognition. Psychological Science, vol. 18. pp. 850-855. [ Links ]
Flege, J.E., Yeni-Komshian, G., and Liu, S. 1999. Age constraints on second language learning. Journal of Memory and Language, vol. 41. pp. 78-104. [ Links ]
Fotenos, A.F., Snyder, A.Z., Girton, L.E., Morris, J.C., and Buckner, R.L. 2005. Normative estimates of cross-sectional and longitudinal brain volume decline in aging and AD. Neurology, vol. 64, no. 6. pp. 1032-1039. [ Links ]
Fregni, F. and Pascual-leone, A. 2007. Technology insight: non-invasive brain stimulation in neurology- perspectives on the therapeutic potential of rTMS and tDCS. Nature Reviews Neurology, vol. 3. pp. 383-393. [ Links ]
Gagnon, D. 1985. Videogame and spatial skills: an exploratory study. Educational Communication & Technology Journal, vol. 33. pp. 263-275. [ Links ]
Gilakjani, A.P. and Ahmadi, S.M. 2011. The effect of visual, auditory, and kinaesthetic learning styles on language teaching. Proceedings of the 2011 international Conference on Social Science and Humanity. International Journal of Modeling and Optimization, vol. 5, no. 2. pp. 469-472. [ Links ]
Gilbert, C.D., Li, W., and Piech, V. 2009. Perceptual learning and adult cortical plasticity. Journal of Physiology, vol. 587. pp. 2743-2751. [ Links ]
Goard, M. and Dan, Y. 2009. Basal forebrain activation enhances cortical coding of natural scenes. Nature Neuroscience, vol. 12. pp. 1444-1449. [ Links ]
Green, C.S. and Bavelier, D. 2006. The cognitive neuroscience of video games. Digital Media: Transformations in Human Communication. Humphreys, L and Messaris, P. (eds.). Peter Lang Publishing, New York, NY. [ Links ]
Green, C.S., Lí, R., and Bavelier, D. 2010a. Perceptual learning during action video games. Topics in Cognitive Science, vol. 2. pp. 202-216. [ Links ]
Green, C.S., Pouget, A., and Bavelier, D. 2010b. Improved probabilistic inference as a general learning mechanism with action video games. Current Biology, vol. 20. pp. 1573-1579. [ Links ]
Greenfield, P.M. 2009. Technology and informal education: what is taught, what is learned? Science, vol. 323. pp. 69-71. [ Links ]
Greenfield, P.M., Dewinstanley, P., Kilpatrick, H., and Kaye, D. 1994. Action video games and informal education: effects on strategies for dividing visual attention. Journal of Applied Developmental Psychology, vol. 15. pp. 105-123. [ Links ]
Greenough, W.T., Black, J.E., and Wallace, C.S. 1987. Experience and brain development. Childhood Development, vol. 58. pp. 539-559. [ Links ]
Greer, P.L. and Greenberg, M.E. 2008. From synapse to nucleus: Calcium-dependent gene transcription in the control of synapse development and function. Neuron, vol. 59, no. 6. pp. 846-860. [ Links ]
Hakuta, K., Bialystok, E., and Wiley E.W. 2003. Critical evidence: a test of the critical-period hypothesis for second-language acquisition. Psychological Science, vol. 14, no. 1. pp. 31-38. [ Links ]
He, H.Y., Ray, B., Dennis, K., and Quinlan, E.M. 2007. Experience-dependent recovery of vision following chronic deprivation amblyopia. Nature Neuroscience, vol. 10. pp. 1134-1136. [ Links ]
Hillman, C.H., Erickson, K.I., and Kramer, A.F. 2008. Be smart, exercise your heart: exercise effects on brain and cognition. Nature Reviews Neuroscience, vol. 9. pp. 58-65. [ Links ]
Hsu, J-L., Leemans, A., Bai, C-H., Lee, C-H., Tsai, Y-F., Chiu, H-C., and Chen, WH. 2008. Gender differences and age-related white matter changes of the human brain: a diffusion tensor imaging study. NeuroImage, vol. 39. pp. 566-577. [ Links ]
Hubel, D.H. and Wiesel, T.N. 1959. Receptive fields of single neurones in the cat's striate cortex. Journal of Physiology, vol. 148. pp. 574-591. [ Links ]
Hubel, D.H. and Wiesel, T.N. 1960. Receptive fields of optic nerve fibres in the spider monkey. Journal of Physiology, vol. 154. pp. 572-580. [ Links ]
Hubel, D.H. and Wiesel, T.N. 1961. Integrative action in the cat's lateral geniculate body. Journal of Physiology, vol. 155. pp. 385-398. [ Links ]
Hubel, D.H. and Wiesel, T.N. 1962. Receptive fields, binocular interaction and functional architecture in the cat's visual cortex. Journal of Physiology, vol. 160. pp. 106-154. [ Links ]
Hubel, D.H. and Wiesel, T.N. 2005. Brain and Visual Perception: The Story of a 25-year Collaboration. Oxford University Press, Oxford. [ Links ]
Hultsch, D.F. and Dixon, R.A. 1990. Learning and memory in aging. Handbook of the Psychology of Aging. 3rd edn. Birren, J.E. and Schaie, K.W. (eds.). Academic Press, San Diego, CA. [ Links ]
Humphreys, L. and Messaris, P. (eds.). 2006. Digital Media: Transformations in Human Communication. Peter Lang Publishing, New York, NY. [ Links ]
Hyland, K. 1993. Culture and learning: a study of the learning style preferences of Japanese students. RELC Journal, vol. 24, no. 2. pp. 69-91. [ Links ]
Imel, S. 998. Teaching adults: Is it different? ERIC Digest, No. 82. http://wiww.ericdigests.org/pre-9211/teaching.htm [ Links ]
Johnson, J.S. and Newport, E.L. 1989. Critical period effects in second language learning: the influence of maturational state on the acquisition of English as a second language. Cognitive Psychology, vol. 21. pp. 60-99. [ Links ]
Jones, N.B. 1997. Applying learning styles research to improve writing instruction. Paper presented at the RELC Seminar on Learners and Language Learning, Singapore, April 1997. [ Links ]
Kadota, T., Horinouchi, T., and Kuroda, C. 2001. Development and aging of the cerebrum: assessment with proton MR spectroscopy. American Journal of Neuroradiology, vol. 22. pp. 128-135. [ Links ]
Kaneko, M., Stellwagen, D., Malenka, R.C., and Stryker, M.P. 2008. Tumor necrosis factor- mediates one component of competitive, experience-dependent plasticity in developing visual cortex. Neuron, vol. 58, no. 5. pp. 673-680. [ Links ]
Kemper, S. 1992. Language and aging. The Handbook of Aging and Cognition. Craik, F.I.M. and Salthouse, T.A. (eds.). Erlbaum, Hillsdale, NJ. [ Links ]
Kempermann, G., Van Praag, H., and Gage, F.H. 2000. Activity-dependent regulation of neuronal plasticity and self-repair. Progress in Brain Research, vol. 127. pp. 35-48. [ Links ]
Kilgard, M.P. and Merzenich, M.M. 1998. Cortical map reorganization enabled by nucleus basalis activity. Science, vol. 279. pp. 1714-1718. [ Links ]
Kim, K.H.S., Relkin, N.R., Lee, K-M., and Hirsch, J. 1997. Distinct cortical areas associated with native and second languages. Nature, vol. 388. pp. 171-174. [ Links ]
Knowles, M.S. 1980. The Modern Practice of Adult Education: From Pedagogy to Andragogy. 2nd edn. Cambridge Books, New York, NY. [ Links ]
Koepp, M.J., Gunn, R.N., Lawrence, A.D., Cunningham, V.J., Dagher, A., Jones, T., Brooks, D.J., Bench, C.J., and Grasby, P.M. 1998. Evidence for striatal dopamine release during a video game. Nature, vol. 393. pp. 266 -268. [ Links ]
Kornell, N. and Bjork, R.A. 2008. Learning concepts and categories: is spacing the 'enemy of induction'? Psychological Science, vol. 19. pp. 585-592. [ Links ]
Krakauer, J.W. and Shadmehr, R. 2006. Consolidation of motor memory. Trends in Neurosciences, vol. 29. pp. 58-64. [ Links ]
Kramer, A.F. and Erickson, K.I. 2007. Capitalizing on cortical plasticity: influence of physical activity on cognition and brain function. Trends in Cognitive Sciences, vol. 11. pp. 342-348. [ Links ]
Kruggel, F. 2006. MRI-based volumetry of head compartments: normative values of healthy adults. NeuroImage, vol. 1. pp. 1-11. [ Links ]
Lendvai, B., Stern, E.A., Chen, B., and Svoboda, K. 2000. Experience-dependent plasticity of dendritic spines in the developing rat barrel cortex in vivo. Nature, vol. 404. pp. 876-881. [ Links ]
Lenneberg, E.H. 1967. Biological Foundations of Language. John Wiley and Sons, New York, NY. [ Links ]
Levi, D.M. and LI, R.W. 2009a. Improving the performance of the amblyopic visual system. Philosophical Transactions of the Royal Society London B (Biological Sciences), vol. 364. pp. 399-407. [ Links ]
Levi, D.M. and Lí, R.W. 2009b. Perceptual learning as a potential treatment for amblyopia: a mini-review. Vision Research, vol. 49. pp. 2535-2549. [ Links ]
Lí, R., Bejjanki, V.R., Lu, Z.L., Pouget, A., and Bavelier, D. 2009a. Playing action video games leads to better perceptual templates. Annual Meeting of the Vision Sciences Society, Naples, FL, May 2009. [ Links ]
Lí, R., Polat, U., Makous, W., and Bavelier, D. 2009b. Enhancing the contrast sensitivity function through action video game training. Nature Neuroscience, vol. 12. pp. 549 -551. [ Links ]
Lutz, A., Slagter, H.A., Dunne, J.D., and Davidson, R.J. 2008. Attention regulation and monitoring in meditation. Trends in Cognitive Sciences, vol. 12. pp. 163-169. [ Links ]
Mac Keracher, D. 2004. Making Sense of Adult Learning. 2nd edn. University of Toronto Press, Toronto. [ Links ]
Magnotta, V.A., Andreasen, N.C., Schultz, S.K., Harris, G., Cizadlo, T., Heckel, D., Nopoulos, P., and Flaum, M. 1999. Quantitative in vivo measurement of gyrification in the human brain: Changes associated with aging. Cerebral Cortex, vol. 9. pp. 151-160. [ Links ]
Mataga, N., Nagai, N., and Hensch, T.K. 2002. Permissive proteolytic activity for visual cortical plasticity. Proceedings of the National Academy of Sciences USA, vol. 99, no. 11. pp.7717-7721. [ Links ]
McCarthy, B. 1982. Improving staff development through CBAM and 4MAT. Educational Leadership, vol. 40. pp. 20-25 [ Links ]
Melton, C.D. 1990. Bridging the cultural gap: a study of Chinese students' learning style preferences. RELC Journal, vol. 21, no. 1. pp.29-54. [ Links ]
Merriam, S. B. 2001. Andragogy and self-directed learning: Pillars of adult learning theory. New Directionsfor Adult and Continuing Education, vol. 89. pp. 3-11. [ Links ]
Merzenich, M.M. and Sooy, F. 2006. Neuroplasticity contributions in children and adults to impairments in learning, memory, and movement. Sensory Perception: Basic Mechanisms and Human Diseases Conference (Perception Sensorielle: Mécanismes Fondamentaux et Déficits chez l'Homme), Paris, March 9-10, 2006. : http://www.pasteur.fr/applications/euroconf/sensoryperception/21Merzenich abstract.pdf. [ Links ]
Muller, C.M. and Best, J. 1989. Ocular dominance plasticity in adult cat visual cortex after transplantation of cultured astrocytes. Nature, vol. 342. pp. 427-230. [ Links ]
Nadel, L. (ed.). 2002. Encyclopaedia of Cognitive Science. Macmillan Reference, London. [ Links ]
Newport, E.L. 2002. Critical periods in language development. Encyclopaedia of Cognitive Science. Nadel, L. (ed.). 2002. Macmillan Reference, London. [ Links ]
Newport, E.L., Bavelier, D., and Neville, H.J.N. 2001. Critical thinking about critical period: Perspectives on a critical period for language acquisition. Language, Brain and Cognitive Development: Essays in Honor of Jacques Mehler. Dupoux, E. (ed.). MIT Press, Boston, MA. [ Links ]
Nithianantharajah, J. and Hannan, A.J. 2006. Enriched environments, experience dependent plasticity and disorders of the nervous system. Nature Reviews Neuroscience, vol. 7. pp. 697-709. [ Links ]
Oray, S., Majewska, A., and Sur, M. 2004. Dendritic spine dynamics are regulated by monocular deprivation and extracellular matrix degradation. Neuron, vol. 44, no. 6. pp. 1021-1030. [ Links ]
Park, D.C., Lautenschlager, G., Hedden, T., Davidson, N.S., Smith, A.D., and Smith, P.K. 2002. Models of visuospatial and verbal memory across the adult life span. Psychology and Aging, vol. 17, no. 2. pp. 299-320. [ Links ]
Peacock, M. 2001. Match or Mismatch? Learning styles and teaching styles in EFL. International Journal of Applied Linguistics, vol. 11, no. 1. pp. 1-20. [ Links ]
PENFiELD, W. and RoBERTS, L. 1959. Speech and Brain Mechanisms. Princeton university Press, Princeton, NJ. [ Links ]
Perani, D., Dehaene, S., Grassi, F., Cohen, L., Cappa, S.F., Dupoux, E., Fazio, F., and Mehler, J. 1996. Brain processing of native and foreign languages. Neuroreport, vol. 7, no. 15-17. pp. 2439-2444. [ Links ]
Pieperhoff, P., Homke, L., Schneider, F., Habel, U., Shah, N.J., ZíLLES, K., and Amunts, K. 2008. Deformation field morphometry reveals age-related structural differences between the brains of adults up to 51 years. Journal of Neuroscience, vol. 28. pp. 828-842. [ Links ]
Pozo, K. and Goda, Y. 2010. Unraveling mechanisms of homeostatic synaptic plasticity. Neuron, vol. 66, no. 3. pp. 337-351. [ Links ]
Quaiser-Pohl, C., Geiser, C., and Lehmann, W. 2006. The relationship between computer-game preference, gender, and mental-rotation ability. Personality and Individual Differences, vol. 40. pp. 609-619. [ Links ]
Rabinowitz, J.C., Craik, F.I.M., and Ackerman, B.P. 1982. A processing resource account of age differences in recall. Canadian Journal of Psychology, vol. 36. pp. 325-344. [ Links ]
Rahi, J.S., Logan, S., Timms, C., Russel-Eggitt, I., and Taylor, D. 2002. Risk, causes, and outcomes of visual impairment after loss of vision in the non-amblyopic eye: a population-based study. Lancet, vol. 360, no. 9333. pp. 597-602. [ Links ]
Ramcharan, V.S. 2005. Plasticity and functional recovery in neurology. Clinical Medicine, vol. 5, no. 4. pp. 368-373. [ Links ]
Reid, J.M. 1987. The learning style preferences of ESL students. TESOL Quarterly, vol. 21, no. 1. pp. 87-111. [ Links ]
Reid, J.M. (ed.). 1995. Learning Styles in the ESL / EFL Classroom. Heinle & Heinle, Boston, MA. [ Links ]
Renier, L. and De Volder, A.G. 2005. Cognitive and brain mechanisms in sensory substitution of vision: a contribution to the study of human perception. Journal of Integrative Neuroscience, vol. 4, no. 4. pp. 489-503. [ Links ]
Restani, L., Cerri, C., Pietrasanta, M., Gianfranceschi, L., Maffie, L., and Caleo, M. 2009. Functional masking of deprived eye responses by callosal input during ocular dominance plasticity. Neuron, vol. 64, no. 5. pp. 707-718. [ Links ]
RoSSi-LE, L. 995. Learning styles and strategies in adult immigrant ESL students. Learning Styles in the ESL / EFL Classroom. Reid, J.M. (ed.). Heinle & Heinle, Boston, MA. [ Links ]
Rueda, M.R., Rothbart, M.K., Mccandliss, B.D., Saccomanno, L., and Posner, M.i. 2005. Training, maturation, and genetic influences on the development of executive attention. Proceedings of the US National Academy of Sciences, vol. 102. pp. 14931-14936. [ Links ]
Salat, D.H., Buckner, R.K., Snyder, A.Z., Greve, D.N., Desikan, R.S.R., Busa, E., Morris, J.C., Dale, A.M., and Fischl, A. 2004. Thinning of the cerebral cortex in aging. Cerebral Cortex, vol. 14. pp. 721-730. [ Links ]
Salthouse, T.A. 1992. Mechanisms of Age-cognition Relations in Adulthood. Erlbaum, Hillsdale, NJ. [ Links ]
Schmidt, R.A. and Bjork, R.A. 1992. New conceptualizations of practice: common principles in three paradigms suggest new concepts for training. Psychological Science, vol. 3. pp. 207-217. [ Links ]
Sheline, Y.I., Mintun, M.A., Moerlein, S.M., and Snyder, A.Z. 2002. Greater loss of 5-HT2A receptors in midlife than in late life. American Journal of Psychiatry, vol. 159. pp. 430-435. [ Links ]
Sherow, S. 2006. Theory to Practice: Adult Teaching and Learning Strategies For Tutors. Pennsylvania State University Institute for the Study of Adult Literacy, Pennsylvania, PA. [ Links ]
Smith, M. 2002. Malcolm Knowles, Informal Adult Education, Self-direction and Andragogy. http://www.infed.org/thinkers/et-knowl.htm [ Links ]
Sowell, E.R., Peterson, B.S., Thompson, P.M., Welcome, S.E., Henkenius, A.L., and Toga, A.W. 2003. Mapping cortical change across the human life span. Nature Neuroscience, vol. 6. pp. 309-315. [ Links ]
Stern, E.A., Maravall, M., and Svoboda, K. 2001. Rapid development and plasticity of layer 2/3 maps in rat barrel cortex in vivo. Neuron, vol. 31. pp. 305-315. [ Links ]
Sternberg, R.J. and Zhang, L.F. (eds.). 2001. Perspectives on Thinking, Learning and Cognitive Styles. Lawrence Erlbaum and Associates, Mahwah, NJ. [ Links ]
Stevens, G. 1999. Age at immigration and second language proficiency among foreign born adults. Language in Society, vol. 28. pp. 555-578. [ Links ]
Stevens, B., Allen, N.J., Vazquez, L.E., Howell, G.R., Christopherson, K.S., Nouri, N., Micheva, K.D., Mehalow, A.K., Huberman, A.D., Stafford, B.A.S., Litke, A.M., Lambris, J.D., Smith, S.J., John, S.W.M., and Barres, B.A. 2007. The classical complement cascade mediates CNS synapse elimination. Cell, vol. 131, no. 6. pp. 1164-1178. [ Links ]
Sullivan, E.V. and Pfefferbaum, A. 2006. Diffusion tensor imaging and aging. Neuroscience and Biobehavior Reviews, vol. 30. pp. 749-761. [ Links ]
Tang, Y.Y., Ma, Y., Wang, J., Fan, Y., Feng, S., Lu, Q., Yu, Q., Sui, D., Rothbart, M.K., Fan, M., and Posner, M.I. 2007. Short-term meditation training improves attention and self-regulation. Proceedings of the US National Academy of Sciences, vol. 104. pp. 17152-17156. [ Links ]
Thompson, B., Mansouri, B., Koski, L., and Hess, R.F. 2008. Brain plasticity in the adult: modulation of function in amblyopia with rTMS. Current Biology, vol. 18. pp. 1067-1071. [ Links ]
Trachtenberg, J.T., Trepel, C., and Stryker, M.P. 2000. Rapid extra granular plasticity in the absence of thalamocortical plasticity in the developing primary visual cortex. Science, vol. 287. pp. 2029-2032. [ Links ]
Vereecken, E.P. and Brabant, P. 1984. Prognosis for vision in amblyopia after loss of the good eye. Archives of Ophthalmology, vol. 102, no. 2. pp. 220-224. [ Links ]
VOLKOW, N.D., LOGAN, J., FOWLER, J.S., Wang, G.J., GUR, R.C., WONG, C., FELDER, C., Gatley, S.J., Ding, Y.S., Hitzemann, R., and Pappas, N. 2000. Association between age-related decline in brain dopamine activity and impairment in frontal and cingulate metabolism. American Journal of Psychiatry, vol. 157. pp. 75-80. [ Links ]
Weber-Fox, C. and Neville, H.J. 1996. Maturational constraints on functional specializations for language processing: ERP and behavioural evidence in bilingual speakers. Journal of Cognitive Neuroscience, vol. 8. pp. 231-256. [ Links ]
Yun, A.J., Bazar, K.A., and Lee, P.Y. 2004.Pineal attrition, loss of cognitive plasticity, and onset of puberty during the teen years: is it a modern maladaptation exposed by evolutionary displacement? Medical Hypotheses, vol. 63, no. 6. pp. 939-950. [ Links ]

