










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAfrican Entomology
versão On-line ISSN 2224-8854
versão impressa ISSN 1021-3589
AE vol.32 Pretoria 2024
http://dx.doi.org/10.17159/2254-8854/2024/a17422
SHORT COMMUNICATION
New data on an old pest complex: The status of Phlyctinus callosus Schönherr and Phlyctinus xerophilus Haran (Coleoptera: Curculionidae) in South Africa
Steffan P. HansenI; Julien M. HaranII; Shelley A. JohnsonI; Noémie M.-C. HévinIII; Pia AddisonI
IDepartment of Conservation Ecology and Entomology, Faculty of AgriSciences, Stellenbosch University, Stellenbosch, South Africa
IICBGP, CIRAD, INRAE, IRD, InstitutAgro, Univ. Montpellier, Montpellier, France
IIICNRS, Institut des Sciences de l'Évolution de Montpellier (Université de Montpellier, CNRS, IRD, EPHE), Montpellier, France
ABSTRACT
Phlyctinus is an endemic weevil genus of the Cape Floristic Region that comprises eight to ten species, previously classified under the monotypic P. callosus sensu lato concept. Two species of this genus, Phlyctinus callosus and P. xerophilus, are of economic concern to the deciduous fruit and grapevine industry in the Western Cape province of South Africa, causing primary damages and being of phytosanitary concern at the adult stage. The taxonomic revision conducted in 2020 raised doubts whether one of the two species was a newly emerging pest and/or what species was considered in studies before the clarification of species boundaries. Also unclear is whether these species can be controlled similarly using current control measures. The aim of this study therefore is to review the pest status of the two species over time based on museum records, field collections and historical publications, and to clarify how control measure(s) are impacted. We infer that neither species appear to be emerging pests in the region, both being of agricultural significance for at least 80 years. Phlyctinus callosus is mostly distributed along the southern coast, while P. xerophilus is distributed in inland valleys. The two species show similar biology, physiology and ecology in agro-ecosystems, and can be excluded from the host plant canopy using stem-barriers, adults being flightless. As such, it is shown that the naming of P. xerophilus as a pest is mainly a taxonomical clarification within a pre-existing species complex.
Keywords: cryptic species complex, deciduous fruit, grapevine, phytosanitary, taxonomical revision, Western Cape province
INTRODUCTION
Phlyctinus Schönherr (Coleoptera: Curculionidae: Entiminae) - also named the banded fruit weevil, v-band weevil or vine weevil - is a polyphagous broad-nosed weevil genus endemic to the Cape Floristic Region (CFR) in the Western Cape province of South Africa (Pringle et al. 2015; Haran et al. 2020; Hévin et al. 2022). This genus has been considered a pest of grapevine and deciduous fruit trees across the majority of south-western Cape growing regions since the late 1800's (Lounsbury 1896; Prinsloo and Uys 2015).
Recent taxonomic revisions including morphological, molecular, geographical and ecological data conducted on Phlyctinus species revealed the genus to be a complex of species containing approximately eight to ten species (and some additional genetic lineages pending a formal analysis), six of which have been named (Haran et al. 2020; Hévin et al. 202; Laborieux 2023). Of this species complex, Phlyctinus callosus sensu stricto Schönherr (s. str., hereafter P. callosus) and Phlyctinus xerophilus sensu lato Haran (s. lat., hereafter P. xerophilus) are the only two species found associated with commercial agriculture (Haran et al. 2020; Hévin et al. 2022). To current knowledge, P. callosus naturally occurs along the south coast of the CFR, and has subsequently expanded its range inland and internationally to St. Helena Island, Réunion Island, Norfolk Island, Tasmania, New Zealand and Western Australia, likely through human-mediated transport. By contrast, P. xerophilus appears naturally restricted to the inland valleys and lower mountain slopes in the CFR. Phlyctinus species are naturally restricted to areas receiving more than 300 mm of rainfall annually (Hévin et al. 2022).
Their larvae are typical of Entiminae: they are soil-living, feed on plant roots and crowns, and pupate in the soil (Marvaldi et al. 2014). The teneral adults then dig out of the soil using deciduous mandibular cusps (Marvaldi et al. 2014). Adult Phlyctinus are flightless and predominantly nocturnal, hiding in soil debris, under bark and in plant material during the day, and emerging to feed on aerial plant parts like leaves, stems and fruit at night (Annecke and Moran 1982; Prinsloo and Uys 2015).
As the flightless adults need to crawl up host plant stems/trunks (or use 'bridges' like adjoining plants or structures) to gain access to the host plant canopy, the use of physical exclusion barriers (sometimes treated with insecticide) placed on trunks or bridges can significantly reduce access of the adult weevils to the host plant canopy (Swartz 1988; Barnes et al. 1994, 1995, 1996; Pryke and Samways 2007). It is the direct feeding damage caused by the adults on fruit, stems, leaves and buds of apple, nectarine and grapevine, and to a smaller extent pear, plum, blueberry and raspberry, that are of economic concern in South Africa (Barnes and Giliomee 1992; Prinsloo and Uys 2015). Phlyctinus adults sheltering in fruit and grapes destined for export cause phytosanitary concern to certain markets like the European Union, the USA and Israel (Marais and Barnes 2004; PPIS 2006; USDA-APHIS 2007; Bragard et al. 2021).
Given the phytosanitary status of Phlyctinus, the newly described species complex and more particularly the description of P. xerophilus, may be cause for concern to export markets for South African deciduous fruit and table grapes (Bragard et al. 2021). Hence, the question: Is Phlyctinus xerophilus an emerging pest in need of revised management and export protocol, or does it merely represent greater taxonomical clarity on a historical pest complex? This study aims to clarify the true risks posed by P. callosus and P. xerophilus based on field observations, museum records and existing management and phytosanitary measures in South Africa.
MATERIALS AND METHODS
During the taxonomic revision of Phlyctinus by Haran et al. (2020), dry-mounted adult Phlyctinus specimens were borrowed from various South African museums (South African National Collection of Insects, Pretoria (SANC) and Stellenbosch University Insect Collection, Stellenbosch (SUIC), as well as several museums whose records were not used during the current study) and European museums (whose records were not used in the current study). Extensive field-collections were also conducted throughout the native range of Phlyctinus from 2017-2020 for the taxonomic revisions by Haran et al. (2020) and Hévin et al. (2022), with specimens being identified by morphological and molecular methods. The first and second authors also conducted extensive fieldwork, including collections and observations, in south-western Cape agro-ecosystems from 2017 to 2023. The current study made use of these records, and a more extensive series of specimens identified in South African museum collections, including the Agricultural Research Council, Infruitec-Nietvoorbij collection, that provided valuable information on historical pest status. During the course of the current study, dry-mounted female specimens with protruding apodemes (Figure 1a) could be identified to a greater degree of certainty than previous studies as P. xerophilus female apodemes have a characteristic sloping tip (Figure 1b); a feature not shared by any other species group in the genus (S Hansen and J Haran, pers. obs.; Figure 1c). All records of species groups other than P. callosus and P. xerophilus were excluded from the study, as none of these species have been recorded as damaging commercial agriculture in either the current or previous studies.

Distribution and agricultural host plant records from museum specimens, (specimen or series of specimens recorded from a certain growing area, sometimes linked with records of being found on or damaging a certain host), historical publications on Phlyctinus damage from specific agricultural regions, and field-observations of damage, were used to establish pest status. A total of 184 series of specimens (consisting of one or multiple specimens collected in the same locality, Suppl. Material 1) were used during the course of this study. All co-ordinates were converted to DMS. When co-ordinates were not presented on a specimen label, the co-ordinates of the location were estimated. All co-ordinates were checked for accuracy and where location/ co-ordinates presented on specimen label were aberrant, co-ordinates of location were corrected. Google Earth Pro was used to estimate, verify and correct co-ordinates. Maps were made using QGIS v. 3.10.
RESULTS AND DISCUSSION
Historical association with agricultural host plants
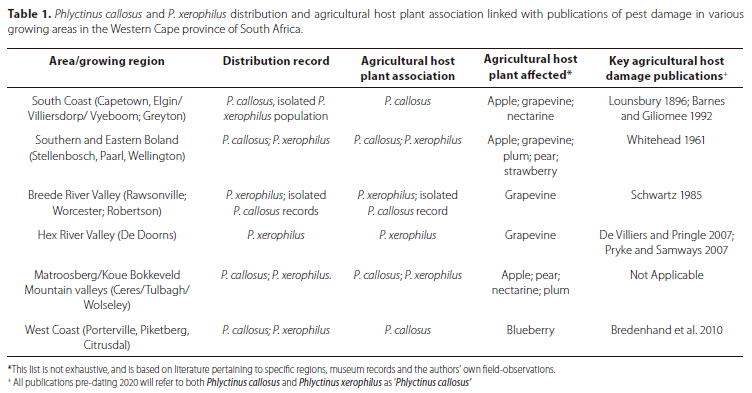
Museum records infer historical association of both P. callosus (1936 on grapevine in Stellenbosch area) and P. xerophilus (1938 on grapevine in Worcester) with agricultural host plants for at least 80 years (Suppl. Material 1, Figure 2). Both species are recorded to have caused historical damage to various deciduous fruit types, grapevines and berries, although only some lineages within both P. callosus and P. xerophilus appear to be highly polyphagous and associated with agricultural crops (Hévin et al. 2022; Laborieux 2023). All populations recorded on ornamental plants or exotic weeds like Plantago lanceolata Linnaeus (Lamiales: Plantaginaceae) (Suppl. Material 1) probably can feed on and damage agricultural crops as well. No other Phlyctinus species was found associated with agricultural hosts, either in the present museum records or recent studies (Haran et al. 2020; Hévin et al. 2022). For now, it is unclear why only P. callosus and P. xerophilus successfully shifted onto agricultural hosts to the extent of being economically significant pests; although this is an area of active research (Laborieux 2023).
Historical and recent (post-2000) distribution records of P. callosus and P. xerophilus are generally similar (Figure 2; Suppl. Material 1), with more extensive records being obtained post-2000 thanks to recent intensive field work. There is evidence of human mediated transport of both species outside their natural distribution ranges in southern Africa (Suppl. Material 1). Phlyctinus callosus seems to be the only Phlyctinus species to have been established outside of southern Africa, specifically in different areas of Oceania, St. Helena Island and Réunion Island (Haran et al. 2020). Phlyctinus callosus is dominant in its natural distribution range on the south coast and seems to be expanding its range in the Western Cape and even the Eastern Cape, appearing far outside its natural distribution range (including within the natural distribution range of P. xerophilus) due to human mediated transport and the creation of suitable habitat (Figure 2; Suppl. Material 1). Phlyctinus callosus appears to be displacing P. xerophilus in disturbed habitats in the natural range of the latter in the Eastern Boland, especially around the Stellenbosch suburban and agricultural area (Figure 2; Suppl. Material 1). According to recent molecular work, the species record of P. xerophilus on the south coast (Table 1, Figure 2) consists of an apparently isolated population that may comprise a separate species within the current P. xerophilus species concept (Laborieux 2023). This potential new species was not recorded as damaging crops in museum records or field observations. Furthermore, mixed populations of P. callosus and P. xerophilus are occasionally found living together on the same site (Suppl. Material 1).
Historical distribution and host plant records, linkage to publications reporting pest-status in specific regions and field-observations from 2017 to 2023, all indicate that both P. callosus and P. xerophilus are not newly emerging pests. All pre-2020 records of "Phlyctinus callosus" pest damage in South Africa can therefore be considered to include both P. callosus and P. xerophilus.
Biology and control strategies
Applied research into microbial biological control and critical thermal limits between P. callosus and P. xerophilus showed these species to be generally similar in biological and physiological parameters tested (Hansen 2023; Hansen et al. 2023; Hansen et al. 2024). Major differences in behaviour and ecology between the two species were not observed within orchards and vineyards during extensive fieldwork by the first and second author from 2017 to 2023 (S. Hansen and J. Haran, pers. obs.).
Physical exclusion and removal of weevils from produce, such as the use of exclusion barriers placed on crop plant trunks, physical removal of weevils from produce and pack houses, and good harvest and pack house management (Marais and Barnes 2004; PPIS 2006; USDA-APHIS 2007), should be equally effective on both P. callosus and P. xerophilus adults. The use of effective stem exclusion barriers, usually consisting of batting (sometimes drenched in insecticide) and/or a sticky/greasy surface (e.g. Plantex"), can greatly reduce the incidence and damage of Phlyctinus on crops (Swartz 1988; Barnes et al. 1994, 1995, 1996), and should show similar efficacy on both P. callosus and P. xerophilus given their similar ecology, size and flightless nature.
The efficacy of stem barriers has been tested and shown in different growing areas and crops, across the range of P. callosus and P. xerophilus in South Africa (Swartz 1988; Barnes et al. 1994, 1995, 1996). Stem barriers also show efficacy against other flightless entimine weevil pests (e.g. genus Eremnus Schönherr) present in Western Cape deciduous fruit orchards and vineyards (Swartz 1988, Prinsloo and Uys 2015).
Similarities in upper and lower critical thermal limits have been shown under two laboratory acclimation regimes for field-collected P. callosus and P. xerophilus adults (Hansen 2023). Although temperature tolerances shown under phytosanitary treatments may differ from the more ecologically relevant tests in Hansen (2023), this is a promising preliminary result in phytosanitary treatments involving cold or heat-treatments of export produce to destroy Phlyctinus adults (Myburg and Kriegler 1967; Johnson and Neven 2011; Smit et al. 2018). Susceptibility of P. callosus and P. xerophilus adults to post-harvest fumigation, such as ethyl formate fumigation (Smit et al. 2020), remains to be assessed.
CONCLUSION
Good taxonomy is paramount, and cryptic species should never be ignored in applied research (Bortolus 2008). Morphological and molecular studies continue to define species status and relationships in the taxonomically challenging genus Phlyctinus. The historical and present pest-status of P. callosus and P. xerophilus in the Western Cape of South Africa has been revised in the light of current control and phytosanitary measures. It was found that broad historical ranges and pest-status (on which a current effective phytosanitary protocol was developed) are similar to recent studies and observations (2000-2023), and that neither species constitutes an emerging pest in this region. The naming of P. xerophilus as a pest can therefore be regarded principally as a taxonomic clarification within a species complex.
ACKNOWLEDGEMENTS
The authors wish to thank Riaan Stals (SANC), Elleunorah Allsopp and Muriel Knipe (ARC Infruitec-Nietboorbij) for providing many of the specimens and museum records used in this study. The authors also wish to thank E. Allsopp and M. de Villiers for valuable insights while preparing the manuscript. Material was collected under Cape Nature Permits Numbers CN35-28-13395 and CN44-30-4229, and with permission from private landowners. The study was funded by Hortgro Science and the South African Table Grape Industry (SATI), South Africa. This work is based on the research supported in part by the National Research Foundation of South Africa (Grant Number 132353).
AUTHOR CONTRIBUTIONS
Steffan P. Hansen: Conceptualisation; Data Curation; Investigation; Methodology; Resources; Visualisation; Writing - Original Draft; Writing - Review and Editing.
Julien M. Haran: Methodology; Resources; Validation; Visualisation; Writing - Original Draft; Writing - Review and Editing.
Shelley Johnson: Conceptualisation; Validation; Visualisation; Writing - Original Draft; Writing - Review and Editing.
Noémie M.-C. Hévin: Software; Writing - Original Draft; Writing - Review and Editing.
Pia Addison: Funding Acquisition; Project Administration; Supervision; Validation; Visualization; Writing - Original Draft; Writing - Review and Editing.
ORCIDS
Steffan P. Hansen: https://orcid.org/0000-0002-6319-4768
Julien M. Haran: https://orcid.org/0000-0001-9458-3785
Shelley Johnson: https://orcid.org/0009-0003-1291-4264
Noémie M.-C. Hévin: https://orcid.org/0000-0003-1730-0010
Pia Addison: https://orcid.org/0000-0002-8227-339X
REFERENCES
Annecke DP, Moran VC. 1982. Insects and mites of cultivated plants in South Africa. Durban: Butterworths. [ Links ]
Barnes BN, Giliomee JH. 1992. Fruit-feeding behaviour of banded fruit weevil, Phlyctinus callosus (Schönherr) (Col., Curculionidae), in apple orchards. Journal of Applied Entomology. 113(1-5):407-415. https://doi.org/10.1111/j.1439-0418.1992.tb00681.x [ Links ]
Barnes BN, Knipe MC, Calitz FJ. 1994. Trunk barriers provide effective control of banded fruit weevil on apples and nectarines. Deciduous Fruit Grower. 44:327-340. [ Links ]
Barnes BN, Knipe MC, Calitz FJ. 1995. Effective weevil control on apple trees with batting trunk barriers. Deciduous Fruit Grower. 45:376-378 [ Links ]
Barnes BN, Knipe MC, Calitz FJ. 1996. Latest results with trunk exclusion barriers for weevil control on apples (Jongste resultate met stamsperbande vir kalander beheer op appels). Deciduous Fruit Grower. 46:284-287. [ Links ]
Bortolus A. 2008. Error cascades in the biological sciences: the unwanted consequences of using bad taxonomy in ecology. Ambio. 37(2):114-118. https://doi.org/10.1579/0044-7447(2008)37[114:ECITBS]2.0.CO;2. [ Links ]
Bragard C, di Serio F, Gonthier P, Miret JAJM, Justesen AF, Magnusson S, Milonas P, Navas-Cortes JA, Parnell S, Potting R, et al. 2021. Pest categorization of Phlyctinus callosus. EFSA Journal. 19(8):6800. https://doi.org/10.2903/j.efsa.2021.6800 [ Links ]
Bredenhand E, Van Hoorn A, May F, Ferreira T, Johnson S. 2010. Evaluation of techniques for monitoring banded fruit weevil, Phlyctinus callosus (Schöenherr) (Coleoptera: Curculionidae), infestation in blueberry. African Entomology. 18(1):205-209. https://doi.org/10.4001/003.018.0118 [ Links ]
Hansen SP. 2023. Phlyctinus callosus (Coleoptera: Curculionidae) species complex linked to biological control and ecolog y. Doctoral thesis. Stellenbosch University, South Africa. [ Links ]
Hansen SP, Malan AP, Haran JM, Addison P. 2023. Susceptibility of adult Phlyctinus (Coleoptera: Curculionidae) to entomopathogens: A first look at potential differences in a newly revised species complex. Journal of Applied Entomology. jen.13214. https://doi.org/10.1111/jen.13214
Hansen SP, Malan AP, Haran JM, Dunn MD, Addison P. 2024. Steinernema yirgalemense (Steinernematidae) to control Phlyctinus callosus and Phlyctinus xerophilus (Curculionidae). Nematology. 9:1-16. https://doi.org/10.1163/15685411-bja10305 [ Links ]
Haran J, Hansen S, Benoit L, Addison P. 2020. Description of five new species in the genus Phlyctinus Schoenherr (Coleoptera, Curculionidae): a first step in deciphering the P. callosus complex. European Journal of Taxonomy. 669(669):1-29. https://doi.org/10.5852/ejt.2020.669 [ Links ]
Hévin NM-C, Hansen S, Addison P, Benoit L, Kergoat GJ, Haran J. 2022. Late Cenozoic environmental changes drove the diversification of a weevil genus endemic to the Cape Floristic Region. Zoologica Scripta. 51(6):724-741. https://doi.org/10.1111/zsc.12563 [ Links ]
Johnson SA, Neven LG. 2011. Heat-controlled atmosphere postharvest treatments for Macchiademus diploterus (Hemiptera: Lygaeidae) and Phlyctinus callosus (Coleoptera:Curculionidae). Journal of Economic Entomology 104(2):398-404. https://doi.org/10.1603/EC10316 [ Links ]
Laborieux L. 2023. Host plant shifts and the emergence of new pest insects: the case of the weevil genus Phlyctinus. MSc thesis. University of Montpellier, France. [ Links ]
Lounsbury CP. 1896. The calandra (Phlyctinus callosus, Bohem). Agricultural Journal of the Cape of Good Hope. 9:63-64. [ Links ]
Marais E, Barnes BN. 2004. Weevils on apples and pears. Stellenbosch: ARC Infruitec-Nietvoorbij. p 7599. [ Links ]
Marvaldi AE, Lanteri AA, del Río MG, Oberprieler RG. 2014. 3.7.5. Entiminae Schoenherr, 1823. In: Leschen RA, Beutel RG (editors), Handbook of Zoology. Coleoptera, beetles-morphology and systematics. 3. Berlin: De Gruyter. p. 503-522. [ Links ]
PPIS. 2006. Bilateral quarantine agreement between the plant protection and inspection services of Israel (PPIS) and the National Department of Agriculture Directorate: Plant Health and Quality of the Republic of South Africa (DPHQ) regarding the conditions for the importation of fresh grapes (Vitis vinifera) from the Republic of South Africa into Israel. State of Israel, Ministry of Agriculture and Rural Development, Plant Protection and Inspection Services, Bet-Dagan, Israel.
Pringle KL, Barnes BN, Blomefield TL. 2015. Apple. In: Prinsloo GL, Uys VM (editors), Insects of Cultivated Plants and Natural Pastures in Southern Africa. Hatfield: The Entomological Society of Southern Africa. p. 350-365. [ Links ]
Prinsloo GL, Uys VM. 2015. Insects of cultivated plants and natural pastures in Southern Africa. Hatfield: The Entomological Society of Southern Africa. [ Links ]
Pryke JS, Samways MJ. 2007. Current control of phytosanitary insect pests in table grape vineyards of the Hex River Valley, South Africa. African Entomology. 15(1):25-36. https://doi.org/10.4001/1021-3589-15.1.25 [ Links ]
Schwartz A. 1988. Efficacy of trunk barriers for the control of key pests on trellised grapevines. South African Journal for Enology and Viticulture. 9(1):16-18. https://doi.org/10.21548/9-1-2304 [ Links ]
Smit R, Jooste MM, Johnson, SA. 2018. CATTS technology: Phytosanitary control and market expansion of chill sensitive Japanese plums for South Africa. Acta Horticulturae. (1194):201-208. https://doi.org/10.17660/ActaHortic.2018.1194.30
Smit R, Jooste MM, Addison MF, Johnson SA. (2020). Ethyl formate fumigation: Its effect on stone and pome fruit quality, and grain chinch bug (Macchiademus diploterus) mortality. Scientia Horticulturae. 261:108845. https://doi.org/10.1016Zj.scienta.2019.108845 [ Links ]
USDA-APHIS. 2007. Work plan for the USDA preclearance inspection and cold treatment of South African deciduous fruit designated for export to the United States of America. USA. South African Department of Agriculture, Land Reform and Rural Development.
Whitehead VB. 1961. Vine snoutbeetles: Some facts about them and their control. Deciduous Fruit Grower. 11(9):261-263. [ Links ]
 Correspondence:
Correspondence:
Steffan P. Hansen
Email: Shansen.rsa@gmail.com
Received: 8 December 2023
Accepted: 16 January 2024


{kind=link}
{kind=link}