










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAfrican Entomology
On-line version ISSN 2224-8854
Print version ISSN 1021-3589
AE vol.31 Pretoria 2023
http://dx.doi.org/10.17159/2254-8854/2023/a16178
RESEARCH ARTICLE
Arthropod associates of Kenyan buffelgrass (Cenchrus ciliaris): a field survey for biological control candidates of a globally important invasive grass
Colin R. MorrisonI, II; Robert M. PlowesI, II; Ivy Ng'iruIII, IV, V; Aaron C. RhodesI, II; Dino J. MartinsIII, VI; Lawrence E. GilbertI, II
IDepartment of Integrative Biology, The University of Texas at Austin, Austin, United States of America
IIBrackenridge Field Laboratory, The University of Texas at Austin, Austin, United States of America
IIIMpala Research Centre, Nanyuki, Kenya
IVSchool of Biosciences, Cardiff University, Cardiff, United Kingdom
VUK Centre for Ecology and Hydrology, Wallingford, United Kingdom
VITurkana Basin Institute, Stony Brook University, New York, United States of America
ABSTRACT
Invasive species are the second largest contributor to biodiversity loss and drivers of ecosystem change. Buffelgrass is a C4, perennial grass native to Africa and Asia that was widely introduced across tropical and subtropical rangelands as livestock forage. Buffelgrass reduces native biodiversity and ecosystem stability in its introduced range when it escapes livestock pastures. Understanding the ecology of insects associated with buffelgrass in its native range may provide an understanding of invasion processes and biological control opportunities where buffelgrass has become an invasive challenge. Here, we present the results of a six-year survey of herbivorous arthropods of a native buffelgrass population from Kenya. Buffelgrass was examined for externally and internally feeding insects of vegetative and reproductive tissues. We also categorised buffelgrass detritivores and parasitoids that may use buffelgrass herbivores as hosts. The samples were photographed and Sanger sequenced to identify them to the lowest possible taxonomic ranking. We collected information on sample abundances, phenologies, tissues consumed, and putative diet breadths. We identified 25 morphospecies representing seven orders and 16 families. The putative host plant specialisation was as high as 67% for Diptera. Phenological variation in herbivore presence correlated with seasonal rainfall and provided a guide for when to conduct follow-up biological control agent searches. The most abundant herbivore was a gall midge (Diptera: Cecidomyiidae) that inhabits buffelgrass culms. Additional research should focus on bringing this species into containment where host choice trials can be conducted to determine if it is truly monophagous and assess its impact on buffelgrass growth.
Keywords: biological control, invasive alien grass, Cecidomyiidae, gall midge, herbivore, introduced, specialist
INTRODUCTION
Buffelgrass, also known as African foxtail grass (Cenchrus ciliaris L., Poaceae), is a C4 perennial bunchgrass native to tropical and subtropical Africa and southwestern Asia (USDA 2010). It is fire-adapted, drought-tolerant, and able to tolerate heavy ungulate grazing through its rapid growth rate and substantial root storage capacity (Burquez-Montijo et al. 2002; Halvorson 2003; Marshall et al. 2012; Martins 2022; Rhodes et al. 2023). These factors combine to make it valuable for cattle pasture introductions yet a formidable challenge when it escapes and becomes invasive (Marshall et al. 2012; Rhodes et al. 2021, 2023). Buffelgrass has been widely introduced globally across tropical and subtropical rangelands because of its economic value as a pasture grass (Walker and Weston 1990) and for restoring land degraded by heavy industry (Harwood et al. 1999; Carrol et al. 2000).
Despite the economic advantages that buffelgrass brings to some land managers, its continued use is controversial because it can lower the ecological stability of communities by reducing native biodiversity and ecosystem services (Marshall et al. 2012). Invasive plants like buffelgrass drastically alter the composition and function of native arthropod communities, likely leading to further impacts on invaded ecosystems (Litt et al. 2014). Buffelgrass directly outcompetes many native plants for water and nutrients (Castellanos et al. 2016; Farrell and Gornish 2019). Buffelgrass also indirectly alters plant community composition by increasing fire frequency and intensity, replacing fire-sensitive natives (Burquez-Montijo et al. 2002). Moreover, buffelgrass root and leaf chemical exudates may deter the germination and growth of natives via allelopathy (Fulbright and Fulbright 1990), similar to other sympatric invasive grasses (Morrison et al. 2023). These factors all contribute to establishing a strong priority effect that makes buffelgrass-dominated habitats difficult to restore (Stevens and Fehmi 2011).
In its native range, buffelgrass primarily grows in open savannas of Africa and arid regions such as the Turkana Basin of eastern Africa (Marshall et al. 2012). In the United States of America (U.S.A.), buffelgrass was first introduced to the south-western states in 1917 (Hanselka 1988). In 1946, the United States Department of Agriculture, Soil Conservation Service imported many cultivars, including a drought-adapted variety from the Turkana Basin (t-4464) to Texas and Arizona USA (Cox et al. 1988), where it survived well in arid conditions. Seed producers sold 7 million kg of t-4464 buffelgrass seed between 1949 and 1985, facilitating its establishment on over 4 million ha in Texas and over 6 million ha in Mexico (Cox et al. 1988). During this period, systematic brush clearing of hundreds of thousands of hectares in South Texas and Mexico created a disturbance that facilitated widespread buffelgrass establishment (Hamilton et al. 2004; Franklin et al. 2006). In the Sonoran Desert of North America, introduced buffelgrass has invaded hundreds of thousands ha, including ecologically sensitive habitats such as Organ Pipe National Monument and Saguaro National Park in Arizona (Marshall et al. 2012). Buffelgrass is also considered invasive in the Hawaiian Islands, South America, and Australia (Marshall et al. 2012). In north-west Australia, buffelgrass was accidentally introduced by cameleers in the 1870s where it gradually naturalised. Beginning in the 1920s, buffelgrass importation into Australia for livestock pasture began (Smyth et al. 2009) and it subsequently became one of the most abundant pastoral grasses on the continent. Unfortunately, it also escaped from livestock pastures and now occupies extensive natural areas in Eucalyptus forests, other woodlands, and grasslands throughout the continent. The consequence is that buffelgrass increased the frequency and intensity of fires in Australia which contributed to the loss of many river red gum trees (Eucalyptus camaldulensis Dehnh.; Myrtaceae), a keystone species of arid Australian environments, and created a feedback loop that favours buffelgrass regeneration over native vegetation (Marshall et al. 2012).
Comparing factors that differ between an invasive's native and introduced ranges can identify mismatches that can be exploited to manage its establishment or spread in novel environments (Sutton et al. 2019). For example, while buffelgrass evolved under heavy ungulate and arthropod herbivory pressure in its native range (Cerling et al. 2015; Sinclair 1975; Olff et al. 2002), insect herbivory is minimal in the areas where it was introduced (Alcala 2005; Palmer and Mound 2020; Sommers et al. 2022). This discrepancy suggests that invasive buffelgrass populations are experiencing an escape from the natural enemies they evolved with (Keane and Crawley 2002), contributing to the competitive factors that make it a successful invader. This coevolutionary mismatch could be leveraged in an arthropod biological control programme if highly specialised buffelgrass arthropods exist in the native range that will not shift to native hosts in its novel environments.
Introducing host-specific insect herbivore biological control for invasive grasses is showing increasing success and efficient workflows are being developed for scaling up their release into the environment (Sutton et al. 2019). A recent example of successfully integrated biological control of an invasive grass in North America is giant reed (Arundo donax L., Poaceae). Reduction in giant reed growth and reproductive output was achieved using a suite of host specific arthropods from its native Mediterranean habitat including monophagous leaf-mining midges (Goolsby et al. 2017), scale bugs (Goolsby et al. 2009), stem-galling flies (Escobar et al. 2020) and stem-galling wasps (Goolsby and Moran 2009). The success of this programme along riparian corridors throughout North America is largely attributable to the sustainability and amount of damage inflicted on giant reed culm biomass by the gall wasp population (Moran et al. 2017). The giant reed programme was exemplar of how identifying and vetting multiple potential agents that feed on different plant tissues increases the chance of a successful and sustainable invasive biological control effort (Schwarzländer et al. 2018). We predict that buffelgrass density and spread can be reduced if populations are pressured with consistent damage from a suite of herbivores. This is supported by the observation that the diversity and seasonal movement of herbivores is an important aspect of the ecology of the savannas where buffelgrass occurs in Kenya (Crego et al. 2020, 2021). In addition, buffelgrass's competitive ability is reduced when biomass reduction is achieved through targeted grazing (Clarke et al. 2005; Rhodes et al. 2021, 2023). Identifying potentially host-specific arthropods in buffelgrass' native range would greatly advance the understanding of buffelgrass invasion ecology and reveal potential biological control candidates.
Our overarching goal was to identify the diversity of herbivorous arthropods that could be considered biological control agents of introduced buffelgrass populations by conducting field surveys in its native range in Kenya. We collected buffelgrass leaf, stem, root, and reproductive tissues from several sites across the Mpala Research Centre in central Laikipia County for six years to summarise the list of herbivorous arthropod species that consume it. Our questions were: 1) Which species of arthropods consistently consumed buffelgrass and how abundant were they across sites? 2) What types of damage did the herbivores inflict on different tissues? and 3) Which months of the year were herbivores most active, and did they track seasonal variation in rainfall? For objectives one and two, we predicted that more abundant buffelgrass herbivores would be internally feeding insects that are specialised on particular tissues (e.g., only eating culms or only eating seeds), and would be found at multiple sites across the property. Conversely, we predicted that less abundant herbivores would be host plant generalists that feed on buffelgrass leaves, and would be found at fewer sites. For the third objective, we predicted that arthropod abundances would increase following average seasonal rainfall peaks because of rain's positive effect on plant growth consistent with a time-lagged growth-tracking herbivory scenario (Kasenene and Roininen 1999; Price and Hunter 2015). In addition, we predicted that the growth-tracking responses of herbivores from particular orders, and within order feeding guilds that utilise different host tissues, would be specific to the seasonal phenology of the buffelgrass tissues that those taxa consume.
MATERIALS AND METHODS
Study site
We conducted a field study at the Mpala Research Centre in Laikipia County of Central Kenya (0.292643 N, 36.898501 E).
Mpala is a 19 400 ha wildlife research area that ranges in elevation from 1650 to 1800 m amsl, with a low density cattle herd (< 3 000 head). Precipitation is variable in this region, historically ranging from 500 to 650 mm yr-1 with two rainfall peaks occurring from March to June and October to December, and a pronounced dry season from December to March (Caylor et al. 2022). Vegetation is characterised as bushland savanna composed of species-rich grasslands interspersed with perennial shrubs such as Croton dichogamus Pax (Euphorbiaceae), Grewia L. spp. (Malvaceae), and Rhus vulgaris Meikle (Anacardiaceae) with an overstorey dominated by Boscia angustifolia A.Rich (Capparaceae), Vachellia Wight & Arn. and Senegalia Raf. species (Fabaceae). The soils where buffelgrass is present are red sandy loams (Ahn and Greiger 1987). Buffelgrass has a patchy, low-density distribution across the property (authors, pers. obs.).
Collection methods
Targeted searches for immature (eggs, nymphs, larvae) and adult buffelgrass-associated arthropods were conducted at nine sites overall, spanning a south to north precipitation gradient (Titcomb et al. 2017), across the Mpala Research Centre property between 2017 and 2022. Three collection methods were used to sample our sites, at different times with variable sampling efforts, over the course of the study. Whole-plant arthropod visual searches included leaves, stems, roots, and reproductive tissues of buffelgrass plants within approximately 50 m2 areas; 37 surveys were conducted using this method at all nine sites between January and December from 2017 to 2022. During visual searches, sections of tissue that appeared to be infested with internal feeders, or had galls present, were harvested from the plants so that samples could be dissected under a stereo microscope or reared out in plastic bags. In a separate collecting approach, approximately 200 green culms per buffelgrass sample were also harvested from plants at the root base and placed in 61 χ 61 χ 91 cm white pop-up cages (BioQuip Products, U.S.A.) so that samples emerged into a confined space where they were collected with an aspirator; 15 surveys were conducted using this method at four sites from June to September 2022. Additionally, buffelgrass litter was collected and placed into Winkler traps (BioQuip Products, U.S.A.) and samples were included in the dataset if their morphospecies identification matched samples collected with targeted searches in which tissue consumption was confirmed; 15 surveys were conducted with this method at eight sites between January and December from 2018 to 2021. All specimens were preserved in vials with 99% ethanol and then assigned to morphospecies. The specimens were photographed with a Swift 5MPx camera (Hong Kong) attached to a Zeiss stereoscope (Germany). Non-herbivorous arthropods (e.g., parasitoids), and others deemed to be present on plants incidentally (e.g., detritivores), were also encountered in this survey and are presented here to provide context on native buffelgrass ecological interactions.
Arthropod associate identification and metadata
Following photography, all arthropod specimens were assigned to unique morphospecies and barcoded at the CO1 locus. A region of approximately 450-750 bp of the CO1 locus was amplified with degenerate primer sets LCO1490-F/HCO2198-R (Folmer et al. 1994), or ZBJArt-F1c/ZBJArt-R2c (Zeale et al. 2011) for specimens that failed to amplify with Folmer primers, followed by Sanger sequence analysis. Sequence similarity search queries were conducted with the NCBI GenBank and Barcode of Life (Ratnasingham and Hebert 2007) databases to assign the morphospecies to the lowest reliable taxonomic level (>80% sequence similarity = order, >90% = family, and >95% = genus). The sequences were deposited in GenBank and the accession identifiers have been provided (Table S2). Field identification notes and tissue collection data were used to assign functional feeding guilds to samples. The herbivore feeding guilds we included were piercing/sucking, gall-forming, culm-boring, leaf-mining, leaf-chewing, and seed predator.
Descriptive statistics
For ecological metadata associated with the samples, we reported the abundances of each morphospecies, the number of sites where they were present, their seasonal phenologies, the tissues where they were collected, the type of damage they inflicted to plants, and their putative diet breadth based on literature searches and natural history knowledge of the taxa. The percentage of herbivores consuming each tissue, and the types of damage inflicted, were calculated by dividing the abundance collected from a particular organ (e.g., culms), or that inflicted a particular type of damage (e.g., gall-formation) by the total number of observations. To describe the buffelgrass arthropod herbivore assemblage, we calculated the species richness, evenness, and Shannon's diversity using ecological diversity index functions in the 'vegan' package (Oksanen et al. 2020) in the R statistical analysis environment, version 4.1.1 (R Core Team 2021). We also calculated the specialist ratio of each herbivore order which was defined as the number of putative monophagous specialist morphospecies in an arthropod order divided by the total number of morphospecies in that order. A species accumulation curve was estimated using the poolaccum function in the 'vegan' package following a published workflow (Sutton 2020) to visualise observed buffelgrass herbivore species richness. Surveys, or sampling effort, was defined as all collections made in a given site and year for the species accumulation curve. Kernel density estimates, or a smoothed histogram, of arthropod order abundances, and abundances of herbivores consuming particular tissues, over time was calculated and visualised using a Gaussian smoothing function and a default bandwidth estimate of one with the geom_density argument from the 'ggplot' R package (Wickham 2016).
RESULTS
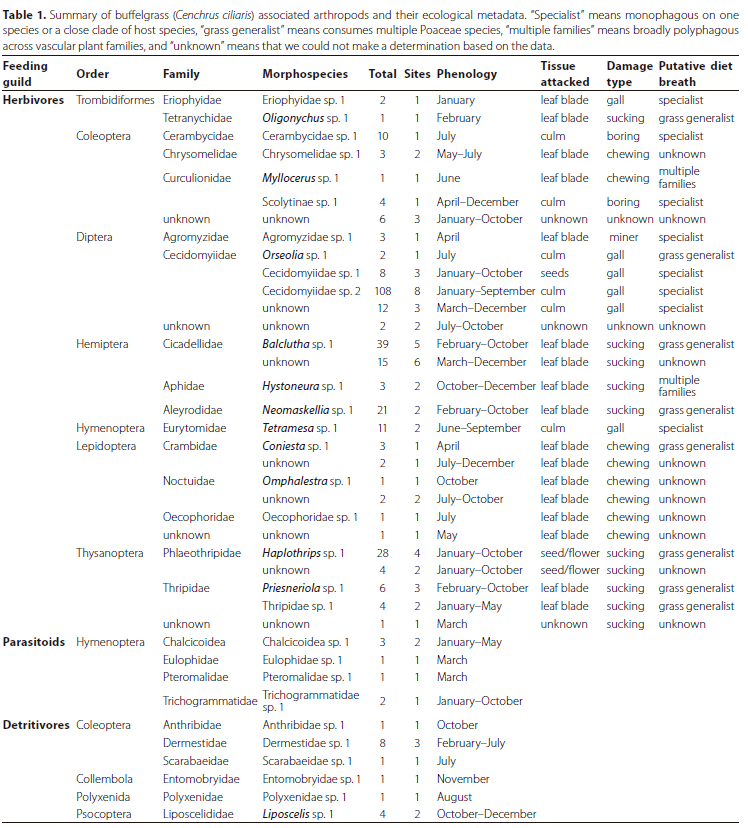
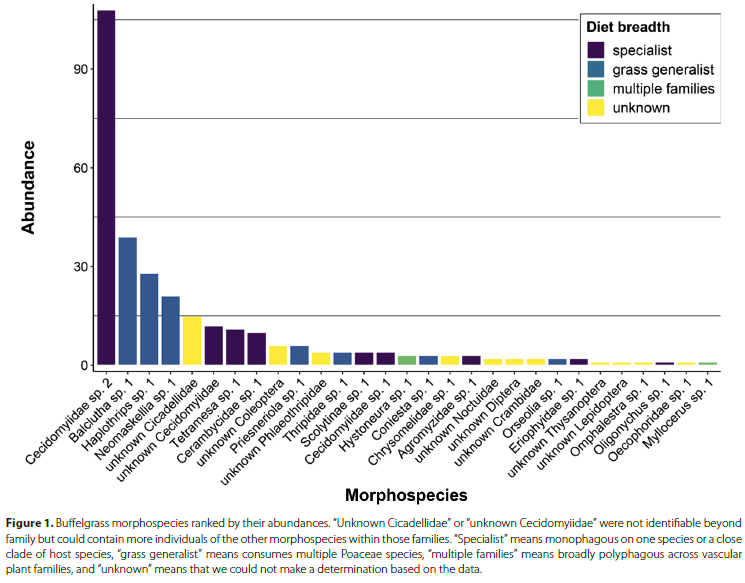
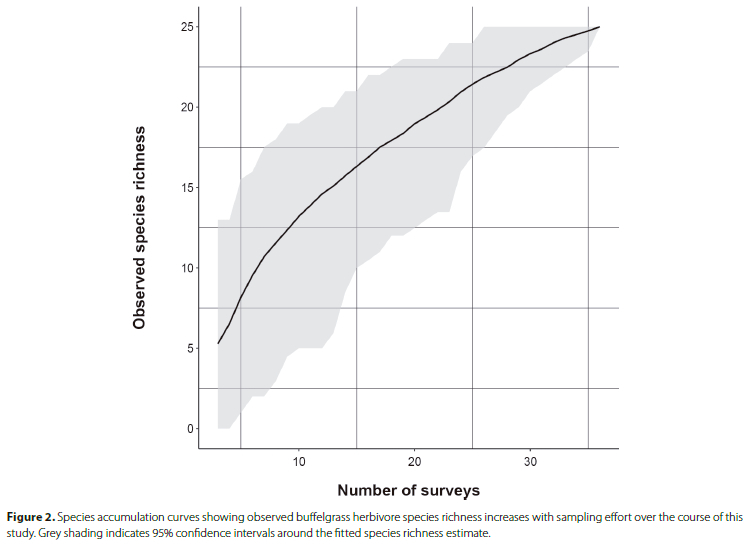
Our first objectives were to identify the arthropod species that consume buffelgrass across Mpala and quantify their abundances. We found herbivores associated with buffelgrass at 9 sites across Mpala. In total, we collected 337 specimens (300 herbivores) and identified them to 25 morphospecies (Table 1; Figure 1). The slope of the species accumulation curve indicated that we observed many of the herbivores that use buffelgrass as a hostplant in this community, but more surveys would probably yield additional species (Figure 2). Buffelgrass was consumed by a phylogenetically diverse assemblage ofherbivores that included seven orders represented by 16 families. The most abundant orders were Diptera (n = 131), Hemiptera (n = 78), Thysanoptera (n = 43), and Coleoptera (n = 24). The most abundant families from these orders were gall midges (Cecidomyiidae, Diptera; n = 126), followed by leaf hoppers (Cicadellidae, Hemiptera; n = 54), thrips (Phlaeothripidae, Thysanoptera; n = 32), and white flies (Aleyorodidae, Hemiptera, n = 21). The species richness, Shannon diversity index, and species evenness of these orders varied considerably among orders (Table S1). Trombidiformes (Acari) and Diptera had low Shannon diversity indices, Lepidoptera and Coleoptera had the highest Shannon diversity indices. Diptera had the lowest species evenness because that assemblage was dominated by one abundant morphospecies, "Cecidomyiidae sp. 2" (Figure 3); the other orders had higher evenness because they were represented by similar numbers of individuals among morphospecies (Table S1). Overall, the entire buffelgrass herbivore assemblage had a specialist to generalist ratio of 28%. The Hymenoptera (specialist ratio = 100%), Diptera (specialist ratio = 67%), Trombidiformes (specialist ratio = 50%), and Coleoptera (specialist ratio = 40%) had host specialist morphospecies while the Hemiptera, Thysanoptera and Lepidoptera had no specialists, or specialist ratios of 0 (Table S1). We also observed four hymenopteran parasitoid morphospecies that were likely using buffelgrass herbivore larvae as hosts, as well as a variety of detritivorous arthropods from four orders that were associated with buffelgrass (Table 1).
The next objective was to catalogue the types of damage that buffelgrass herbivores inflict on different tissues. Most of the individual herbivores that we observed were consumers of grass culms (49% of the total observations), with gall-forming Cecidomyiidae being the most abundant culm-feeding family. Leaf blades were the second most common tissue consumed (36%), with the piercing/sucking Balclutha sp. 1 (Cicadeliidae) being the most common morphospecies feeding on leaves. Seeds and flowers were attacked by the smallest proportion of herbivores (12%), with the piercing/sucking Haplothrips sp. 1 (Phlaeothripidae) being the most abundant morphospecies observed consuming these tissues; the tissue consumed could not be determined for 3% of our observations. Overall, gallformation was the most common functional guild (46% of the total observations), followed by leaf piercing/sucking (41%); culm-boring (5%), leaf-chewing (5%), and leaf-mining (1%) comprised small fractions of the total functional feeding guilds; damage type could not be determined conclusively for 2% of the total observations. Cecidomyiidae gall midges were the most abundant gallers, Cicadellidae leaf hoppers and thrips were abundant piercing herbivores, Cerambycidae and Scolytinae (Curculionidae) were grouped as culm-boring Coleoptera and Lepidoptera comprised the external leaf chewers.
Our final objective was to catalogue in which months of the year buffelgrass herbivores were most active. Total herbivore abundances were high from January to April (39% of total observations) and again between July and October (51% oftotal). Few herbivores were observed from May to June or November to December (10% of total). These trends varied among the most abundant orders (Figure 4A). Diptera consumed buffelgrass year-round, with observations peaking in September. Hemiptera were also present year-round, with peaks of abundance in March and October. Thysanoptera were mainly observed from January to June, with a strong peak in April. Coleoptera were observed from March to August, with a strong peak in July. We observed phenological differences in the tissue types consumed as well (Figure 4B). Herbivore emergence from culms was highest between June and September, with another emergence period in January that was primarily driven by culm-feeding Cecidomyiidae larvae. Leaf blade feeding followed a bimodal pattern as hemipteran abundance peaked in February-March and October. Flower consumption peaked between April and May, and seed consumption had a strong peak between September and October, which tracked emergence patterns of Phlaeothripidae and Thripidae (Thysanoptera), respectively. Buffelgrass phenological periods (leaf flushing, setting seeds, etc.) is shown on Figure 4. Precipitation was variable among Mpala collection sites from 2017 to 2022 (Figure 4), with a wet season that lasted from February to April (20.1 mm ± SEM 3.1) followed by short bursts of precipitation in June (18.2 mm ± SEM 2.3) and December (25.2 mm ± SEM 3.7). July to November was the driest period at Mpala during this study (11.9 mm ± SEM 1.5).
DISCUSSION
This study contributed information on insects associated with grasses in East Africa where relatively little work has been done on phytophagous insects and Poaceae.
Several morphospecies that we observed may be good buffelgrass biological control candidates. The most abundant herbivores that we observed were three Cecidomyiidae gall midge morphospecies. These included Orseolia sp., which may be related to the specialist Orseolia species reported on Paspalum sp. in West Africa (Harris et al. 1999) or it may be Orseolia oryzae Wood-Mason (the "rice midge") which is a widely distributed generalist feeder of rice and other wild Poaceae species (Bonzi 1980). We also repeatedly collected two Cecidomyiidae morphospecies that we suspect are buffelgrass specialists based on the observation that species in this family tend to be highly host-specific (Dorchin et al. 2019; Gagné and Jaschhof 2021). "Cecidomyiidae sp. 1" was repeatedly detected from seeds and had barcodes that were distinct from other Cecidomyiidae on the GenBank and Barcode of Life databases (Table 1). Many gall midge species that specialise in consuming host seeds can lower population spread by damaging reproductive tissues, while avoiding culms and vegetative foliage (Barnes 1956; Neiman and Manglitz 1972; Kolesik et al. 2007; López et al. 2015). Buffelgrass spreads readily via high longevity seeds, with vegetative growth playing a lesser role in its dispersal (Marshall et al. 2012). Therefore, controlling the spread of seeds with a host-specific seed predator could be an attractive option for stakeholders interested in buffelgrass biological control. "Cecidomyiidae sp. 2" samples also had a unique barcode, as well as a distinct culm gall morphology. This morphospecies was the most abundant herbivore observed in this study, comprising over a third of our observations. It was detected at more Mpala sampling sites than the other herbivores (n = 8) and was present at varying frequencies from January through September. The suitability of culm-galling midges as biological control agents has been demonstrated in a variety of plant systems (Hinz and Müller-Schärer 2000; Impson et al. 2008; Paterson et al. 2014), including grasses that have become invasive in North America (Goolsby et al. 2017; Rector et al. 2021). However, culm-galling Cecidomyiidae remain poorly sampled in Africa, and their true host usage patterns are largely unknown (Harris and Dorchin 2012; Gagné and Jaschhof 2021). Follow-up host choice assays, surveys of arthropod herbivores on related grass species in buffelgrass' native range, morphological and molecular characterisation of these morphospecies are required to verify their host specificity and taxonomic affiliation.
In addition to gall-midges, we observed other host herbivores that target different tissues, and we suspect are possible host specialists based on the literature. These included leaf-mining flies (Agromyzidae, Diptera), culm-galling Tetramesa wasps (Eurytomidae, Hymenoptera), leaf-galling mites (Eriophyidae, Trombidiformes), culm-boring Coleoptera in the Scolytinae and Cerambycidae (Table 1). Our observed species richness accumulation curve indicated that additional sampling effort Mpala would uncover more arthropod species that consume buffelgrass, and some of these potential species could also be host-specific. The species accumulation curve also suggests that sampling buffelgrass arthropod herbivores at additional study sites across sub-Saharan Africa would reveal additional species worth consideration and compliment the results of the species reported here. These results strongly suggest that biological control could be a viable method for managing invasive buffelgrass populations and indicate that multiple functional guilds may be available to exert pressure on different plant tissues (Sutton et al. 2019). Further consideration of these insects as buffelgrass biocontrol candidates should also account for climatic suitability between native and potential introduction ranges.
Other morphospecies that we observed could not be conclusively differentiated from congeners that are known to consume multiple Poaceae host plants. These putative "grass generalists" included Balclutha sp. (Cicadellidae, Hemiptera; Morgan et al. 2013), Neomaskellia sp. (Aleyrodidae, Hemiptera; Palmer 2009), Coniesta sp. (Crambidae, Lepidoptera; Kfir et al. 2002), Haplothrips and Priesneriola morphospecies (Thysanoptera; Mound and Ng 2009; Palmer and Mound 2020). However, these genera are poorly described and require further species identification and host breadth testing to confirm that these morphospecies indeed consume multiple Poaceae hosts.
Invasive grasses can produce a variety of non-target environmental impacts such as increasing fire frequency by accumulating dense biomass of flammable and senescent litter which exacerbate invasion (Burquez-Montijo et al. 2002; Brooks et al. 2004). Introduced buffelgrass populations produce copious leaf litter (McDonald and McPherson 2013) and few native detritivores have adapted to feed on and decompose litter from introduced grasses (Sands and Goolsby 2011). Therefore, arthropod detri-tivores from the native range have been proposed as potential biological control agents for senescing and dead invasive grass biomass if they meet host specificity requirements as agents (Sands and Goolsby 2011). The list of detritivores presented here could be evaluated for host specificity to explore the possibility of deploying them as biological controls (Table 1).
The hymenopteran parasitoids that we detected may be using larvae of the Cecidomyiidae or other specialist buffelgrass herbivores as hosts. Culm-galling Cecidomyiidae populations can suffer high mortality from parasitoids (Hawkins and Gagne 1989; Johnson et al. 2013). Thus, identifying parasitoids of these biological control candidates in their native range and potential release areas are research priorities because herbivore suppression by parasitoids is a major cause for biological control programme failure (Paynter et al. 2010).
The temporal precipitation data we presented is important for interpreting the phenology of herbivory in this system. We predicted that arthropod eclosion and abundance would track the seasonal growth pattern of buffelgrass tissues that they consume. This pattern has also been observed in other tropical savanna regions that vary in annual precipitation (Denlinger 1980; Pinheiro et al. 2002). Buffelgrass typically flushes out from root resources within a few days of 1-2 cm of cumulative rainfall after a dry period. Then the plants tend to remain in a growth phase in which they delay flowering until either there has been no rain for several weeks, or they have reached their full height at about four weeks. If there is only a single pulse of rain, they may flower within about two weeks. If there is sustained rainfall, buffelgrass will flower after about four weeks. The seed set begins 1 or 2 weeks after flowering (authors, pers. obs.). Our results support a time-lagged growth tracking herbivory scenario (Kasenene and Roininen 1999; Price and Hunter 2015). The abundance of dipteran and coleopteran herbivores peaked from July to September when gall midges and culm-boring beetles began emerging from grasses that had reached their seasonal growth peak with time to produce tall culms. Seed and flower predator thrip abundance peaked earlier in the rainy season when grasses began flowering and setting seeds. Piercing/sucking hemipterans showed a bimodal distribution in peak abundances that appeared to track leaf flushing early in the rainy season and then later in that season before vegetative tissues begin to senesce. Our tissue phenology consumption data showcased how buffelgrass herbivore assemblages responded to its growth and reproductive stages, and highlighted the periods in which specific feeding guilds are active. These data show how abiotic conditions affect plant growth and thus facilitate herbivore phenology over time. In addition, they provide a guide as to when follow-up buffelgrass biological control agent field surveys should be conducted. Further research on insect abundance, tissue preference, and phenology is needed to help understand these patterns, especially given the variable trends in climate and frequent droughts that were experienced in the region during our study (Caylor et al. 2022).
Plant invasions of rangelands are particularly serious because these ecosystems account for a significant portion of the Earth's terrestrial surface area (Lund 2007) and provide humans and wildlife with many ecosystem services (Havstad et al. 2007). In Australia and Arizona U.S.A., buffelgrass establishment is altering the fire regimes such that those regions are shifting from native desert communities to invasive species-dominated grasslands (D'antonio and Vitousek 1992; Burquez-Montijo et al. 2002), which diminishes the value of their cultural and ecological services. Buffelgrass is contributing to unprecedented wildfires in Hawaii which negatively affect the island's endemic flora (Marshall et al. 2012). In South Texas U.S.A., native vegetation is the foundation of natural wildlife diversity (Whittaker et al. 1979), including white-tailed deer and northern bobwhite quail, two economically important wildlife species that do not respond well to buffelgrass spread (Hernandez and Guthery 2012). These are just four regional examples of how these processes are happening and affecting native ecosystems across buffelgrass' entire introduced range (Marshall et al. 2012). The key to overcoming these challenges may be utilising herbivorous arthropod associates that are sufficiently host-specific and damaging to warrant their use as biological control agents (Sutton et al. 2019). The information summarised here can guide a focused search for biological control candidates for regions where stakeholders agree that non-native buffelgrass should be managed.
ACKNOWLEDGEMENTS
We thank the Insect-Plant Lab at Mpala Research Centre including Kennedy Saitot, George Koech, Godfrey Amooni, Katero Kamukunji, Godrey Gitimu, and Aimee Gaitho for assistance in the field and the Mpala Research Centre staff for logistical support of this research. Molecular work was done by Kaitlyn Williams, Shellsea Miller, and Caroline Chessher. Microscope photographs were taken by Qiyang Liu. Jonathan Reid Fischer assisted with the collation of the data. Clarke van Steenderen assisted with sequence processing. We thank NACOSTI for permission to conduct this study. The Lee and Ramona Bass Foundation provided funding for this research.
AUTHORS' CONTRIBUTIONS
CRM collated the data, summarised the results, and wrote the manuscript; IN collected the data and led the field crew; RMP and ACR managed data collection; RMP and LEG planned the project and secured funding; DJM oversaw the Kenyan field activities and logistics. All authors contributed to manuscript drafts and gave final approval for publication.
ORCID IDs
Colin R. Morrison - https://orcid.org/0000-0003-2370-3617
Robert M. Plowes - https://orcid.org/0000-0001-5803-755X
Ivy Ng'iru - https://orcid.org/0000-0002-4766-6251
Aaron C. Rhodes - https://orcid.org/0000-0002-4501-0726
Dino J. Martins - https://orcid.org/0000-0003-1581-1642
Lawrence E. Gilbert - https://orcid.org/0000-0003-4418-2996
REFERENCES
Ahn PM, Geiger LC. 1987. Soils of Laikipia District. Nairobi: Ministry of Agriculture, National Agricultural Laboratories. [ Links ]
Alcala GC. 2005. Response of desert mule deer to habitat alterations in the lower Sonoran Desert. PhD Dissertation. University of Arizona, AZ, USA. [ Links ]
Barnes HF. 1956. Gall Midges of Economic Importance. Volume VII. Gall Midges of Cereal Crops. London: Crosby Lockwood & Son Ltd. [ Links ]
Bonzi SM. 1980. Wild host plants of rice gall midge Orseolia oryzae WM (Dipt. Cecidomyidae) in Upper Volta. WARDA Technical Newsletter. 2:5-6. [ Links ]
Brooks ML, D'antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D. 2004. Effects of invasive alien plants on fire regimes. BioScience. 54(7):677-688. https://doi.org/10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2 [ Links ]
Burquez-Montijo A, Miller ME, Martinez-Yrizar A. 2002. Mexican grasslands, thornscrub, and the transformation of the Sonoran Desert by invasive non-native buffelgrass (Pennisetum ciliare). In: Tellman B, editor. Invasive non-native species in the Sonoran region. Tucson: University of Arizona Press and Arizona-Sonora Desert Museum. p. 126-146. [ Links ]
Clarke PJ, Latz PK, Albrecht DE. 2005. Long-term changes in semi-arid vegetation: Invasion of an exotic perennial grass has larger effects than rainfall variability. Journal of Vegetation Science. 16:237-248. https://doi.org/10.1111/j.1654-1103.2005.tb02361.x [ Links ]
Carroll C, Merton L, Burger P. 2000. Impact of vegetative cover and slope on runoff, erosion, and water quality for field plots on a range of soil and spoil materials on central Queensland coal mines. Soil Research. 38:313-328. https://doi.org/10.1071/SR99052 [ Links ]
Castellanos AE, Celaya-Michel H, Rodriguez JC, Wilcox BP. 2016. Ecohydrological changes in semiarid ecosystems transformed from shrubland to buffelgrass savanna. Ecohydrology. 9:1663-1674. https://doi.org/10.1002/eco.1756 [ Links ]
Caylor KK, Gitonga JG, Martins DJ. 2022. Mpala Research Centre Meteorological and Hydrological Dataset [Data File]. Mpala Research Centre, Laikipia, Kenya.
Cerling TE, Andanje SA, Blumenthal SA, Brown FH, Chritz KL, Harris JM, et al. 2015. Dietary changes of large herbivores in the Turkana Basin, Kenya from 4 to 1 Ma. Proceedings of the National Academy of Sciences. 112:11467-11472. https://doi.org/10.1073/pnas.1513075112 [ Links ]
Cox JR, Martin-R MH, Ibarra F, Fourie JH, Rethman NFG, Wilcox DG. 1988. The influence of climate and soils on the distribution of four African grasses. Journal of Range Management. (41)2:127-139. https://doi.org/10.2307/3898948 [ Links ]
Crego RD, Ogutu JO, Wells HB, Ojwang GO, Martins DJ, Leimgruber P, Stabach JA. 2020. Spatiotemporal dynamics of wild herbivore species richness and occupancy across a savannah rangeland: Implications for conservation. Biological Conservation. 242:108436. https://doi.org/10.1016/j.biocon.2020.108436 [ Links ]
Crego RD, Wells HB, Ndung'u KS, Evans L, Njeri Nduguta R, Chege MA, et al. 2021. Moving through the mosaic: identifying critical linkage zones for large herbivores across a multiple-use African landscape. Landscape Ecology. 36:1325-1340. https://doi.org/10.1007/s10980-021-01232-8 [ Links ]
D'Antonio CMD, Vitousek PM. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change Annual Review of Ecology and Systematics. 23:63-87. https://doi.org/10.1146/annurev.es.23.110192.000431 [ Links ]
Denlinger DL. 1980. Seasonal and annual variation of insect abundance in the Nairobi National Park, Kenya. Biotropica. 12(2):100-106. https://doi.org/10.2307/2387725 [ Links ]
Dorchin N, Harris KM, Stireman III JO. 2019. Phylogeny of the gall midges (Diptera, Cecidomyiidae, Cecidomyiinae): Systematics, evolution of feeding modes and diversification rates. Molecular Phylogenetics and Evolution. 140:106602. https://doi.org/10.1016/j.ympev.2019.106602 [ Links ]
Escobar Y, Guermache F, Bon MC, Kerdellant E, Petoux L, Desurmont GA. 2020. Biology, ecology, and impact of Cryptonevra nigritarsis Duda, a potential biological control agent against the giant reed Arundo donax. Biological Control. 147:104287. https://doi.org/10.1016/j.biocontrol.2020.104287 [ Links ]
Farrell HL, Gornish ES. 2019. Pennisetum ciliare: a review of treatment efficacy, competitive traits, and restoration opportunities. Invasive Plant Science and Management. 12(4):203-213. https://doi.org/10.1017/inp.2019.28 [ Links ]
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology. 3:294-299. [ Links ]
Franklin KA, Lyons K, Nagler PL, Lampkin D, Glenn EP, Molina-Freaner F, et al. 2006. Buffelgrass (Pennisetum ciliare) land conversion and productivity in the plains of Sonora, Mexico. Biological Conservation. 127:62-71. https://doi.org/10.1016/j.biocon.2005.07.018 [ Links ]
Fulbright N, Fulbright TE. 1990. Germination of 2 legumes in leachate from introduced grasses. Rangeland Ecology and Management/ Journal of Range Management Archives. 43:466-467. https://doi.org.10.2307/3899014 [ Links ]
Gagné RJ, Jaschhof M. 2021. A catalog of the Cecidomyiidae (Diptera) of the world. 5th Edition. Washington, DC: United States Department of Agriculture & Entomological Society of Washington. [ Links ]
Goolsby JA, Moran P. 2009. Host range of Tetramesa romana Walker (Hymenoptera: Eurytomidae), a potential biological control of giant reed, Arundo donax L. in North America. Biological Control. 49(2):160-168. https://doi.org/10.1016/j.biocontrol.2009.01.019 [ Links ]
Goolsby JA, Moran PJ, Adamczyk JJ, Kirk AA, Jones WA, Marcos MA, Cortés E. 2009. Host range of the European, rhizome-stem feeding scale Rhizaspidiotus donacis (Hemiptera: Diaspididae), a candidate biological control agent for giant reed, Arundo donax (Poales: Poaceae) in North America. Biocontrol Science and Technology. 19:899-918. https://doi.org/10.1080/09583150903189099 [ Links ]
Goolsby JA, Vacek AT, Salinas C, Racelis A, Moran PJ, Kirk AA. 2017. Host range of the European leaf sheath mining midge, Lasioptera donacis Coutin (Diptera: Cecidomyiidae), a biological control of giant reed, Arundo donax L. Biocontrol Science and Technology. 27:781795. https://doi.org/10.1080/09583157.2017.1342222 [ Links ]
Halvorson WAGP. 2003. Factsheet for: Penisetum ciliare (L.). In: Halvorson WAGP, editor. USGS Weeds in the West project: Status of Introduced Plants in Southern Arizona Parks. Tucson: U.S. Geological Survey, National Parks Service. [ Links ]
Hamilton WT, McGinty A, Ueckert DN, Hanselka CW, Lee MR. 2004. Brush management: past, present, future, Volume 7. College Station: Texas A and M University Press.
Hanselka CW. 1988. Buffelgrass - South Texas wonder grass. Rangelands. 10:279-281. [ Links ]
Harris KM, Dorchin N. 2012. The taxonomic status of Jean-Jacques Kieffer's type specimens of Afrotropical Cecidomyiidae (Diptera). African Invertebrates. (53)1:169-174. [ Links ]
Harris KM, Williams CT, Okhidievbie O, Lasalle JE, Polaszek A. 1999. Description of a new species of Orseolia (Diptera: Cecidomyiidae) from Paspalum in West Africa, with notes on its parasitoids, ecology and relevance to natural biological control of the African rice gall midge, O. oryzivora. Bulletin of Entomological Research. 89:441-448. https://doi.org/10.1017/S0007485399000577 [ Links ]
Harwood MR, Hacker JB, Mott JJ. 1999. Field evaluation of seven grasses for use in the revegetation of lands disturbed by coal mining in Central Queensland. Australian Journal of Experimental Agriculture. 39:307-316. https://doi.org/10.1071/EA98119 [ Links ]
Havstad KM, Peters DP, Skaggs R, Brown J, Bestelmeyer B, Fredrickson E, Herrick J, Wright J. 2007. Ecological services to and from rangelands of the United States. Ecological Economics. 64:261-268. https://doi.org/10.1016/j.ecolecon.2007.08.005 [ Links ]
Hawkins BA, Gagné RJ. 1989. Determinants of assemblage size for the parasitoids of Cecidomyiidae (Diptera). Oecologia. 81(1):75-88. https://doi.org/10.1007/BF00377013 [ Links ]
Hernandez F, Guthery FS. 2012. Beef, brush, and bobwhites: quail management in cattle country. College Station: Texas A and M University Press.
Hinz HL, Müller-Schärer H. 2000. Influence of host condition on the performance of Rhopalomyia n. sp.(Diptera: Cecidomyiidae), a biological control agent for scentless chamomile, Tripleurospermum perforatum. Biological Control. 18:147-156. https://doi.org/10.1006/bcon.2000.0818 [ Links ]
Impson FAC, Kleinjan CA, Hoffmann JH, Post JA. 2008. Dasineura rubiformis (Diptera: Cecidomyiidae), a new biological control agent for Acacia mearnsii in South Africa. South African Journal of Science. 104:247-249. [ Links ]
Johnson PJ, Torrez VC, Buhl PN. 2013. A new species of Platygaster Latreille (Hymenoptera: Platygastridae) parasitizing Chilophaga virgati Gagné (Diptera: Cecidomyiidae). Zootaxa. 3630:184-190. https://doi.org/10.11646/zootaxa.3630.1.8 [ Links ]
Kasenene JM, Roininen H. 1999. Seasonality of insect herbivory on the leaves of Neoboutonia macrocalyx in the Kibale National Park, Uganda. African Journal of Ecology. 37:61-68. https://doi.org/10.1046/j.1365-2028.1999.00162.x [ Links ]
Keane RM, Crawley MJ. 2002. Exotic plant invasions and the enemy release hypothesis. Trends in Ecology and Evolution. 17:164-170. https://doi.org/10.1016/S0169-5347(02)02499-0 [ Links ]
Kfir R, Overholt WA, Khan ZR, Polaszek A. 2002. Biology and management of economically important lepidopteran cereal stem borers in Africa. Annual review of entomology. 47:701-31. https://doi.org/10.1146/annurev.ento.47.091201.145254 [ Links ]
Kolesik P, Sarfati MS, Brockerhoff EG, Kelly D. 2007. Description of Eucalyptodiplosis chionochloae sp. nov., a cecidomyiid feeding on inflorescences of Chionochloa (Poaceae) in New Zealand. New Zealand Journal of Zoology. 34:107-115. https://doi.org/10.1080/03014220709510069 [ Links ]
Litt AR, Cord EE, Fulbright TE, Schuster GL. 2014. Effects of invasive plants on arthropods. Conservation Biology. 28:1532-1549. https://doi.org/10.1111/cobi.12350 [ Links ]
López JMP, Johnson PJ, Gagné RJ, Boe A. 2015. A new species of Stenodiplosis (Diptera: Cecidomyiidae) on Spartina grasses (Poaceae) with notes on its biology and its parasitoid Tetrastichus bromi (Hymenoptera: Eulophidae). Zootaxa.4057:115-124. https://doi.org/10.11646/zootaxa.4057.1.7 [ Links ]
Lund HG. 2007. Accounting for the world's rangelands. Rangelands. 29:310. https://doi.org/10.2111/1551-501X(2007)29[3:AFTWR]2.0.CO;2 [ Links ]
Marshall VM, Lewis MM, Ostendorf B. 2012. Buffel grass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: a review. Journal of Arid Environments. 78:1-12. https://doi.org/10.1016/j.jaridenv.2011.11.005 [ Links ]
Martins DJ. 2022. Grasses of East Africa. Cape Town: Struik Nature, Penguin Random House South Africa (Pty) Ltd. [ Links ]
McDonald CJ, McPherson GR. 2013. Creating hotter fires in the Sonoran Desert: Buffelgrass produces copious fuels and high fire temperatures. Fire Ecology. 9:26-39. https://doi.org/10.4996/fireecology.0902026 [ Links ]
Moran PJ, Vacek AT, Racelis AE, Pratt PD, Goolsby JA. 2017. Impact of the arundo wasp, Tetramesa romana (Hymenoptera: Eurytomidae), on biomass of the invasive weed, Arundo donax (Poaceae: Arundinoideae), and on revegetation of riparian habitat along the Rio Grande in Texas. Biocontrol Science and Technology. 27(1):96-114. https://doi.org/10.1080/09583157.2016.1258453 [ Links ]
Morgan AR, Smith-Herron AJ, Cook JL. 2013. Rapid spread of Balclutha rubrostriata (Hemiptera: Cicadellidae) in Texas and southwestern Louisiana, USA with notes on its associated host plants. Florida Entomologist. 96:477-81. https://doi.org/10.1653/024.096.0213 [ Links ]
Morrison CR, Rhodes AC, Bowman EA, Plowes RM, Sedio BE, Gilbert LE. 2023. Adding insult to injury: Light competition and allelochemical weapons interact to facilitate grass invasion. Ecosphere 14:e4438. https://doi.org/10.1002/ecs2.4438 [ Links ]
Mound LA, Ng YF. 2009. An illustrated key to the genera of Thripinae (Thysanoptera) from Southeast Asia. Zootaxa. 2265:27-47. https://doi.org/10.11646/zootaxa.2265.1.2 [ Links ]
Neiman EL, Manglitz GR. 1972. The Biology and Ecology of the Bromegrass Seed Midge in Nebraska. Nebraska Agricultural Experiment Station Research Bulletin. 252:1-23. [ Links ]
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, et al. 2020. Community ecology package. R package version 2.5.7. https://CRAN.R-project.org/package=vegan
Olff H, Ritchie ME, Prins HH. 2002. Global environmental controls of diversity in large herbivores. Nature. 415:901-904. https://doi.org/10.1038/415901a [ Links ]
Palmer CM. 2009. Buffel grass (Cenchrus ciliaris' L.) is a host for the 'sugarcane whitefly' Neomaskellia bergii (Signoret) (Hemiptera: Aleyrodidae) in central Australia. The Australian Entomologist. 36:89-95. [ Links ]
Palmer CM, Mound LA. 2020. The diversity of thrips (Insecta: Thysanoptera) on buffel grass (Cenchrus ciliaris) is markedly lower than on native grasses in an urban landscape. Journal of Urban Ecology. 6:juaa024. https://doi.org.10.1093/jue/juaa024 [ Links ]
Paynter Q, Fowler SV, Hugh Gourlay A, Groenteman R, Peterson PG, Smith L, Winks CJ. 2010. Predicting parasitoid accumulation on biological control agents of weeds. Journal of Applied Ecology. 47:575582. https://doi.org/10.1111/j.1365-2664.2010.01810.x [ Links ]
Paterson ID, Vitorino MD, De Cristo SC, Martin GD, Hill MP. 2014. Prioritisation of potential agents for the biological control of the invasive alien weed, Pereskia aculeata (Cactaceae), in South Africa. Biocontrol Science and Technology. 24:407-425. https://doi.org/10.1080/09583157.2013.864382 [ Links ]
Pinheiro F, Diniz IR, Coelho D, Bandeira MPS. 2002. Seasonal pattern of insect abundance in the Brazilian cerrado. Austral Ecology. 27:132136. https://doi.org/10.1046/j.1442-9993.2002.01165.x [ Links ]
Price PW, Hunter MD. 2015. Population dynamics of an insect herbivore over 32 years are driven by precipitation and host-plant effects: testing model predictions. Environmental Entomology. 44:463-473. https://doi.org/10.1093/ee/nvv039 [ Links ]
R Core Team. 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ratnasingham S, Hebert PD. 2007. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Molecular ecology notes. 7(3):355-364. https://doi.org/10.1111/j.1471-8286.2007.01678.x [ Links ]
Rector BG, Gagné RJ, Perilla López JM, Tonkel KC, Bon MC, Guermache F, Cristofaro M. 2021. Taxonomic description of Stenodiplosis tectori n. sp. (Diptera: Cecidomyiidae), a seed parasite of cheatgrass, Anisantha tectorum, based on morphological and mitochondrial DNA data. Insects. 12:755. https://doi.org/10.3390/insects12080755 [ Links ]
Rhodes AC, Rutledge J, DuPont B, Plowes RM, Gilbert LE. 2021. Targeted grazing of an invasive grass improves outcomes for native plant communities and wildlife habitat. Rangeland Ecology and Management. 75:41-50. https://doi.org/10.1016/j.rama.2020.11.007 [ Links ]
Rhodes AC, Plowes RM, Gilbert LE. 2023. Mitigating buffelgrass invasion through simulated targeted grazing: understanding restoration potential in a variable precipitation regime. Restoration Ecology. 31(6):e13923. https://doi.org/10.1111/rec.13923 [ Links ]
Sands D, Goolsby JA. 2011. The case for biological control of exotic African grasses in Australia and USA using introduced detritivores. In: Wu Y, Johnson T, Sing S, Raghu S, Wheeler G, Pratt P, et al. (eds). Proceedings of the XIII International Symposium on Biological Control of Weeds. Waikoloa, Hawaii, USA, 11-16 September 2011. p. 103-109.
Schwarzländer M, Hinz HL, Winston RL, Day MD. 2018. Biological control of weeds: an analysis of introductions, rates of establishment and estimates of success, worldwide. BioControl. 63:319-331. https://doi.org/10.1007/s10526-018-9890-8 [ Links ]
Sinclair AR. 1975. The resource limitation of trophic levels in tropical grassland ecosystems. The Journal of Animal Ecology. 497-520. https://doi.org/10.2307/3608
Smyth A, Friedel M, O'Malley C. 2009. The influence of buffel grass (Cenchrus ciliaris) on biodiversity in an arid Australian landscape. The Rangeland Journal. 31:307-320. https://doi.org/10.1071/RJ08026 [ Links ]
Sommers P, Davis A, Chesson P. 2022. Invasive buffel grass (Cenchrus ciliaris) increases water stress and reduces success of native perennial seedlings in southeastern Arizona. Biological Invasions. 24:18091826. https://doi.org/10.1007/s10530-022-02750-5 [ Links ]
Stevens JM, Fehmi JS. 2011. Early establishment of a native grass reduces the competitive effect of a non-native grass. Restoration Ecology. 19:399406. https://doi.org/10.1111/j.1526-100X.2009.00565.x [ Links ]
Sutton GF, Canavan K, Day MD, Den Breeyen A, Goolsby JA, Cristofaro M, McConnachie A, Paterson IF. 2019. Grasses as suitable targets for classical weed biological control. BioControl. 64:605-622. https://doi.org/10.1007/s10526-019-09968-8 [ Links ]
Sutton GF. 2020. Guy Sutton: Plotting species accumulation curves using ggplot. Retrieved from https://guysutton.netlify.app/posts/2020-10-29-automated-plotting-of-species-accumulation-curves-by-group/
Titcomb G, Allan BF, Ainsworth T, Henson L, Hedlund T, Pringle RM, et al. 2017. Interacting effects of wildlife loss and climate on ticks and tick-borne disease. Proceedings of the Royal Society B: Biological Sciences. 284:20170475. https://doi.org/10.1098/rspb.2017.0475 [ Links ]
United States Department of Agriculture. 2010. Pennisetum ciliare (L.) Link Buffelgrass. Plants Profile. United States Department of Agriculture, Natural Resources Conservation Service.
Walker B, Weston EJ. 1990. Pasture development in Queensland-a success story. Tropical Grasslands. 24:257-268. [ Links ]
Whittaker RH, Gilbert LE, Connell JH. 1979. Analysis of two-phase pattern in a mesquite grassland, Texas. The Journal of Ecology. 67:935-952. https://doi.org/10.2307/2259222 [ Links ]
Wickham H. 2016. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag. [ Links ]
Zeale MR, Butlin RK, Barker GL, Lees DC, Jones G. 2011. Taxon-specific PCR for DNA barcoding arthropod prey in bat faeces. Molecular Ecology Resources. 11:236-244. https://doi.org/10.1111/j.1755-0998.2010.02920.x [ Links ]
 Correspondence:
Correspondence:
Colin R Morrison
Email:crmorrison@utexas.edu
Received: 15 June 2023
Accepted: 18 August 2023


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}