










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkAfrican Entomology
versión On-line ISSN 2224-8854
versión impresa ISSN 1021-3589
AE vol.31 Pretoria 2023
http://dx.doi.org/10.17159/2254-8854/2023/a15613
RESEARCH ARTICLE
Evaluating the establishment of a new water hyacinth biological control agent in South Africa
Benjamin E MillerI; Julie A CoetzeeII; Martin P HillI
ICentre for Biological Control, Department of Zoology and Entomology, Rhodes University, Makhanda, South Africa
IICentre for Biological Control, Department of Botany, Rhodes University, Makhanda, South Africa
ABSTRACT
Megamelus scutellaris Berg (Hemiptera: Delphacidae) is the most recent of nine biological control agents developed to manage invasive water hyacinth, Pontederia (=Eichhornia) crassipes Mart. (Pontederiaceae), in South Africa. More than a million M. scutellaris have been mass-reared and released since the first introduction of the agent into South Africa in 2013, successfully establishing overwintering populations at 32 sites in seven of the nine provinces. Establishment has also been recorded at seven of these sites through natural dispersal from sites where they had established. Inundative releases, where large numbers of M. scutellaris are released regularly, have resulted in excellent establishment, and caused a significant reduction in water hyacinth cover in areas where, historically, biological control seemed unlikely due to excessive eutrophication. Although M. scutellaris has established well throughout South Africa through classical biological control methods, this study also showed that inundative releases of biological control agents over multiple seasons results in the most effective control of the weed, especially at cool temperate and eutrophic sites.
Keywords: Biological control, Delphacidae, mass rearing, Megamelus scutellaris, Pontederia crassipes, post-release evaluation
INTRODUCTION
Many of South Africa's waterways face invasion from floating aquatic weeds, of which water hyacinth, Pontederia crassipes Mart. (Pontederiaceae), remains the worst despite decades of interventions to mitigate its spread (Coetzee et al. 2021). The invasiveness of water hyacinth, which originated from South America, has been exacerbated by highly eutrophic waterways that stimulate the plant's rapid growth and reproduction (Coetzee and Hill 2012). While biological control has reduced the invasiveness of water hyacinth in South Africa, at many sites, the populations of the weed remains above a tolerable level (Tipping et al. 2014a; Moran et al. 2021).
To date, nine species of biological control agents (eight arthropods and one fungal pathogen) have been released on water hyacinth in South Africa (Hill and Coetzee 2017). The complement of nine different control agents totals more than has been released anywhere that the plant is invasive (Hill and Coetzee 2017). Despite the large suite of introduced natural enemies, classical biological control of water hyacinth is constrained by cooler temperatures experienced in the more temperate areas of South Africa, as low temperatures reduce the fecundity of beneficial insects released for biological control while increasing their development times (Byrne et al. 2010; May and Coetzee 2013). The local climate can significantly influence the potential establishment of a biological control agent population, particularly in areas that experience winter frosting (Byrne et al. 2010). Additionally, water hyacinth will rapidly recover from winter frost damage during the spring. However, the insect populations, which were depleted by the cold weather, will only reach their maximum density by the late summer. Therefore, there is a delay in the effectiveness of biological control (Byrne 2010; Reddy et al. 2019; Miller et al. 2021).
Since the first releases of biocontrol agents in the late 1980s and 1990s, several species have established on water hyacinth in South Africa, particularly the weevils Neochetina eichhorniae Warner and N. bruchi Hustache (Coleoptera: Brachyceridae) (Hill and Cilliers 1999), which feed on the laminae and mine the petioles of the plants (Bashir et al. 1984). However, given variable levels of success of biological control of P. crassipes, Hill and Olckers (2001) suggested that there was a need for additional control agents that were able to reproduce quickly, could respond to environmental perturbations more effectively, and would complement the current suite of biological control agent species. To this end, the phloem-feeding planthopper, Megamelus scutellaris Berg (Hemiptera: Delphacidae) was released in South Africa in 2013 (Coetzee 2013). High levels of feeding activity by M. scutellaris causes significant damage at the cellular level, thus reducing the photosynthetic efficiency of water hyacinth (Tipping et al. 2011; Miller et al. 2019). Additionally, because M. scutellaris feeds by inserting its stylets directly into the plant's vascular tissues (Sosa et al. 2007), the feeding wounds provide an entry point for infection by phytopathogens (Sutton et al. 2016).
Megamelus scutellaris is multivoltine with overlapping generations, and its life cycle is relatively short compared to some of the other biological control agents (Sosa et al. 2005). Eggs, usually laid between the leaf and the petiole, hatch approximately seven days after oviposition in outdoor environments. The nymphs develop from first to fifth instar (imago) approximately 25 days after hatching, depending on temperature (Sosa et al. 2005). While other biological control agents of water hyacinth live a portion of their lifecycles within the petioles or laminae as larvae, M. scutellaris nymphs are vagile, which is advantageous on systems that employ integrated management practices involving manual removal or herbicide applications to remove water hyacinth (Goode et al. 2021). If water hyacinth is destroyed by physical removal or herbicide sprays, the larvae of holometabolous species like the Neochetina spp. are destroyed with the plants. However, the more mobile M. scutellaris can move to the remaining plants in the system, increasing the likelihood that biological control will persist despite the other integrated control practices (Goode et al. 2021).
Similar to other delphacids, M. scutellaris adults show wing dimorphism, where the adults are either brachypterous (having underdeveloped wings) or macropterous (having developed wings for dispersal). This morphological variation is only presented in adults and remains permanent (Zera and Denno 1997). Under optimal conditions, most M. scutellaris will be brachypterous, while macroptery is most common when intraspecific competition is high or when host-plant quality is poor (Denno and Roderick 1992; Fitzgerald and Tipping 2013). Brachyptery is favoured in healthy systems as the metabolic cost of flight muscles reduces fecundity, therefore, flightless adults have larger ovaries and produce more offspring (Zera and Denno 1997). Macroptery has been observed at numerous sites in South Africa (Supplementary Figure 1), and this is considered to be important for dispersal to new sites.
Megamelus scutellaris was imported to South Africa from the U.S.A. and tested for host-specificity, whereafter it was released onto water hyacinth infested systems. The first release was made on the Kubusi River in the Eastern Cape province (-32.565; 27.489) in 2013. Since then, the agent has been released onto several systems within South Africa, covering variable climatic regions such as the Mediterranean climate zones of the Western Cape which have hot dry summers and winter rainfall, the humid subtropical zones typical of the KwaZulu-Natal east coast which have very hot summers and mild winters, and the drier subtropical highlands with cold and dry winters (NorthWest and Gauteng provinces) (Peel et al. 2007). The only other country currently where M. scutellaris is a classical biological control agent outside of South America is the U.S.A., where the agent was first released in 2010 (Tipping et al. 2014b).
Given the variety of South African climatic zones, climatic mismatches are a concern. Such mismatches between the climates of the native and host ranges are responsible for the failures of a significant proportion of classical biological control programmes, as the natural enemies usually inhabit a narrower fundamental niche than their host plants (Harms et al. 2021). May and Coetzee (2013) reported that M. scutellaris would be less likely to establish and persist in the cooler interior of South Africa, particularly when temperatures were below its developmental threshold of 11.5 °C, while others determined that the survival and fecundity of M. scutellaris in rearing cultures is significantly reduced below 18 °C and above 33 °C (Grodowitz et al. 2017). Thus, the upper and lower temperature thresholds of M. scutellaris might be exceeded in parts of South Africa where these temperatures are commonplace.
However, recent research has suggested that the thermal tolerances of some insects are more plastic than was previously thought. Ten years after release in South Africa, the established range of the water hyacinth mirid, Eccritotarsus catarinensis Carvalho (Hemiptera: Miridae) in South Africa, was found to be broader than initially anticipated from thermal physiology models (Coetzee et al. 2007; Coetzee et al. 2008). Griffith et al. (2019) determined that E. catarinensis had established at both the warmest and coldest release sites in South Africa, while Porter et al. (2019) demonstrated the plastic nature of the thermal limits of E. catarinensis. These studies illustrate that control agent populations can locally adapt in sites that are climatically variable after establishment, making these populations more thermally suitable over time. Therefore, M. scutellaris may be better suited to cooler sites than May and Coetzee (2013) initially anticipated, as the thermal tolerances of field and rearing populations of the insect have changed over time since the first quarantine colony was tested in 2009 (C. Trivella, unpub.).
While predictive thermal physiology models are powerful tools, they cannot account for behavioural thermoregulation, thermal plasticity, or variable microclimates within the structure of water hyacinth mats which may contribute to the increased thermal ranges experienced by the biological control agents. The difficulty in predicting the distributions of a biological control agent once it has been released from rearing facilities speaks to the importance of post-release evaluations in modern biological control implementation practices. Therefore, we aimed to determine where M. scutellaris has established in South Africa. Through a synthesis of information relating to current release strategies, we aim to formulate a new best-practice approach for future releases of the agent such that establishment and effective biological control is implemented. The information gathered from these analyses can then be used to implement strategic adaptive management policies toward managing water hyacinth in South Africa and, in the future, in other African countries.
MATERIALS AND METHODS
Field trips were conducted during the summer months (October-April) between 2017 and 2021 to monitor all sites where M. scutellaris had been released. Most of the sites were sampled during March and April when the insect population is highest (Miller et al. 2021), but incidental sites were recorded as they were found. Site selection was guided by the release records from 2013 to 2020, and 34 sites in total were investigated based on these release records. Some other sites where M. scutellaris was suspected of having dispersed were also surveyed on an ad hoc basis. These sites were prioritised based on their proximity to waterbodies where M. scutellaris had swarmed recently, while other populations were found as part of annual surveys to monitor invasive weeds around South Africa (Coetzee et al. 2021).
All M. scutellaris releases are coordinated by the Centre for Biological Control, with the overwhelming majority of insects being released from the Waainek Mass Rearing Facility. Some insect releases have been made from other cultures maintained by the South African Sugar Research Institute (SASRI) in the KwaZulu-Natal province, as well as some from the City of Cape Town (COCT) facility in the Western Cape and some stations set up in strategic areas with community partners (Hill et al. 2021). These records are included.
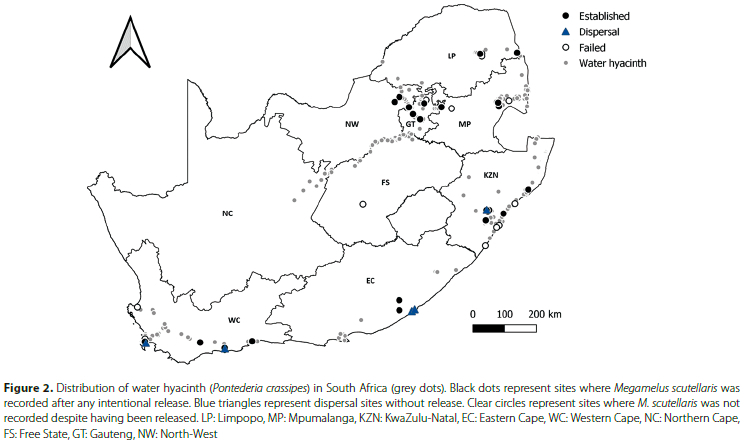
A visual investigation to look for signs of M. scutellaris was performed at each site, usually nearest to the point where the insects were released. If the insects were not found through a brief visual inspection, water hyacinth plants in 10 different areas of the water hyacinth mat were submerged using a plastic 70-litre bin with the bottom removed to make a tube. A cross made from fencing wire was affixed to the bottom of the sampler to push the plants down (see Miller et al. 2021 for the detailed sampling protocol). Once submerged, biological control agents on the plants float to the surface, and the M. scutellaris (whose light body colour contrasted with the black plastic) could be easily identified. The numbers of M. scutellaris per sample are not presented in this study, as the focus was on the presence or absence of the insects. Populations of M. scutellaris were only considered established if they were found after at least one entire winter since release, and these were assumed to be the progeny of the founding population. Megamelus scutellaris was assumed to have not established if it was not found at a site. The data were then grouped into sites where released populations of M. scutellaris had established or sites where the insect was present but releases had not been made (dispersal). These were plotted on a map using QGIS v. 3.24.3.
RESULTS
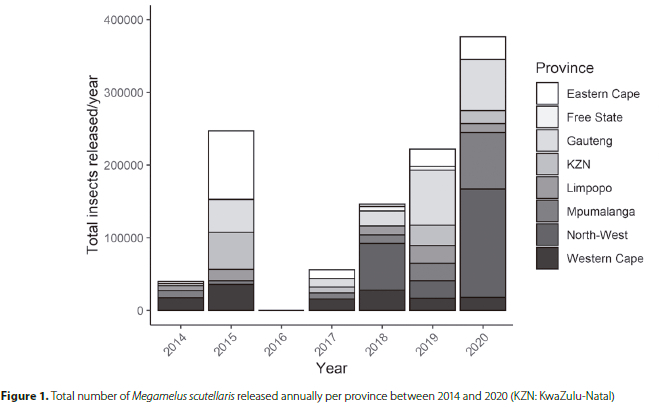
Cumulatively, 1 087 345 individual M. scutellaris were released from the Waainek Mass Rearing facility at 108 sites in South Africa between 2014 and 2020 (Figure 1). Between 2014 and 2015, the majority of releases were made in the Eastern Cape (96 500 at 8 sites) KwaZulu-Natal (53 500 at 8 sites), the Western Cape (53 345 at 11 sites), and Gauteng (45 000, 5 releases at one site). Very few releases were made in the Free State province, with the province only accounting for 12 000 (1.1%) of the total number of M. scutellaris released in South Africa, where three releases were made at one site, and establishment was not successful. The Free State province does not have many water hyacinth sites, except for the Vaal River, where the plants form patches that are typically managed using herbicide. More recently, increased release effort in the North-West province has accounted for 21.8% of the total number of M. scutellaris released in South Africa (39.6% of all M. scutellaris released in the year 2020), the majority of which were released on the Hartbeespoort Dam. The Gauteng province accounted for 20.6% of the total number of M. scutellaris released between 2013 and 2020. Due to problems with rearing, no M. scutellaris were released in 2016.
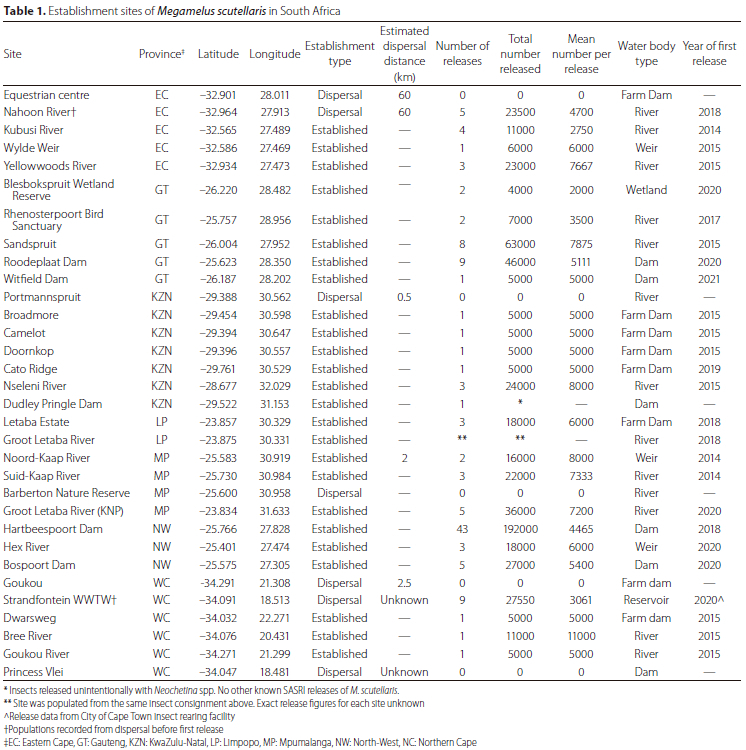
There are 32 sites where M. scutellaris is considered to have established in South Africa, while at nine sites establishment has failed (Figure 2). Of the 32 successful establishment sites, the insect has been recorded as established by natural dispersal to seven of these. Some of these dispersal sites were downstream of known release sites. Migration to downstream areas was anticipated as periodic flooding events moved the water hyacinth and M. scutellaris downstream. In contrast, establishment on Dudley Pringle Dam in KwaZulu-Natal was suspected to have originated from unintentional 'hitch-hikers' on water hyacinth leaves following releases of N. eichhorniae from a mass rearing site based at the South African Sugar Research Institute (SASRI). Until that point, no known releases of M. scutellaris were made from SASRI. Surprisingly, dispersal sites in East London (the Nahoon River and the Equestrian Centre Dam) were located approximately 60 km from the Kubusi River, the nearest site where M. scutellaris was released, suggesting long-distance dispersal by macropterous individuals. The Strandfontein Waste Water Treatment Works and Princess Vlei sites were also dispersal sites where the origin of the founding population remains unknown, but intentional releases soon followed at both sites. Overall, M. scutellaris has been recorded in seven of South Africa's nine provinces, favouring the warmer, subtropical areas or areas with a Mediterranean climate (Table 1). Water hyacinth is not a significant concern in the drier parts of South Africa's interior, except for sections of the Vaal River. However, M. scutellaris has not been released here as the river is usually managed with herbicide and manual removal (Department of Forestry, Fisheries and the Environment, pers. comm.).
The KwaZulu-Natal (KZN) province had the highest number of establishment sites (n = 7), where M. scutellaris was released just once, except the Nseleni River, where three releases were made (Table 1). The majority of M. scutellaris releases were made on lentic water bodies in the North-West (NW) and Gauteng (GT) provinces, where 237 000 and 224 000 M. scutellaris had been released in each province, respectively, despite releases only beginning in the North-West in 2018 (Figure 1). Most of the releases of M. scutellaris were focused on the Hartbeespoort and Roodeplaat dams. Despite being hypertrophic and high-elevation sites, M. scutellaris populations rapidly built up at both dams causing declines in the total cover of water hyacinth (Coetzee et al. 2022).
Despite predictions that M. scutellaris would not establish in some parts of South Africa due to climatic conditions thought to exceed its thermal capabilities, establishment over multiple years was recorded at both the coldest and hottest long-term monitoring sites in South Africa: the Kubusi River in the Eastern Cape province and at the Nseleni River in northern KwaZulu-Natal (Table 1). The Kubusi River is located in a cold, temperate region, with as many as 56 frost days recorded in a season and year-round rainfall (Byrne et al. 2010). In contrast, the Nseleni River falls within a sub-tropical climatic zone on the eastern coastline and is characterised by high temperatures and summer rains with an average maximum annual temperature of 26 °C (Byrne et al. 2010; Jones 2014).
DISCUSSION
Determining whether a biological control agent can establish in a novel environment is critical to the success of an invasive weed management project (McFayden 1998). Despite the climatic variability, M. scutellaris has been able to survive in a variety of regions within South Africa, successfully overwintering in both the coldest sites where biological control has been attempted, as well as the warmest, despite data showing that M. scutellaris would not persist where temperatures exceeded 30 °C (May & Coetzee 2013). Climate has also had a significant influence on the efficacy ofM. scutellaris as a classical biological control agent and on water hyacinth biological control as a whole in South Africa. Despite the short generation time of M. scutellaris, the time taken for population recovery in colder sites is still a concern, with the impact of biological control often lagging behind the exponential growth of the weed until late in the season when the populations fully recover (Miller et al. 2021). Therefore, implementing new inundative release strategies beginning early in spring has helped speed up the recovery of biological control agent by mass-releasing the insects shortly after winter to boost their populations in the field (Hill et al. 2021).
In the U.S.A., M. scutellaris has been released in several States, including Louisiana, California, and Florida (Grodowitz et al. 2014). Despite extensive releases, establishment has been reported as variable in the U.S.A., and this has been attributed to summer temperatures exceeding the upper thermal limit of 39.1 °C (Coetzee, unpub. data) for M. scutellaris (Grodowitz et al. 2014; Tipping et al. 2014b; Pratt et al. 2021). Another biotype of M. scutellaris collected from Paraguay has been released in the south-eastern U.S.A. (Foley et al. 2016). Because Paraguay falls within a warmer climatic zone than Argentina, the Paraguayan biotype of M. scutellaris may be more heat tolerant than the Argentine biotype initially released in the U.S.A. and South Africa. Hence, the Paraguayan biotypes might increase the establishment potential in areas that experience hot summers.
Prospecting for more climatically suitable biotypes can address climatic mismatches. However, there are challenges to collecting new biological control agent populations, particularly if cryptic species are collected (Paterson et al. 2019). Some species within a species complex may not be sufficiently host-specific and could threaten the safety of a biological control programme (Bickford et al. 2007).
The danger of collecting cryptic species and the costs involved in importing new populations of biological control agents can be prohibitive. Therefore, research into cold-hardening insects prior to release is becoming more frequent (Zhou et al. 2011; Mukherjee et al. 2014). Griffith et al. (2019) determined that populations of E. catarinensis in the field could undergo microevolutionary processes driven by climate. These adaptations resulted in slight expansions of the insects' thermal tolerances in the field. They showed that laboratory populations of an insect could be cold-hardened before release in the field. Cold-hardening can increase a biological control agent's ability to survive in colder areas, suggesting that, over time, insects might be able to acclimate more effectively to climates that do not match their host ranges, and this technique should be investigated for potential applications in the field (Griffith et al. 2019).
Additionally, temperatures within the canopy of water hyacinth are more stable than atmospheric conditions and, thus, provide a more suitable microclimate for the insects (Grodowitz et al. 2017). For example, the three-dimensional structure of water hyacinth mats provides a temperature buffer to M. scutellaris living on the plants. Within a closed canopy of water hyacinth, the temperatures are usually lower than the surrounding air due to cooling effects from water moving through the xylem and shading from the leaves (Grodowitz et al. 2014). Therefore, M. scutellaris might behaviourally thermoregulate by moving up and down petioles to maintain a stable temperature, usually remaining near the water's surface when temperatures are warm and ascending petioles in cooler weather (Grodowitz et al. 2014). However, research suggests that acclimating insects to local temperatures can be achieved prerelease to increase the propensity for population establishment at release sites that might have climates different from the rearing temperatures (Griffith et al. 2019).
One of the major limitations when implementing a biological control programme is determining how many control agents to release and how frequently to perform releases. A single, large release provides a robust founding population but this remains susceptible to disturbances and stochastic events, such as floods and herbicide applications. In contrast, multiple releases of smaller populations allow the implementer to mitigate single stochastic events, but at the expense of a large founding population. Smaller releases could also increase susceptibility to potential Allée effects (Grevstad 1999). The current practice for utilising M. scutellaris has been to regularly release as many insects as possible from mass-rearing facilities while maintaining sustainable populations and avoiding excessive bottlenecking (Coetzee et al. 2022). The number of agents released was typically between 1 000 and 15 000 M. scutellaris per week during the summer months (Centre for Biological Control, Release Records). While single releases have led to successful establishment at 10 of the sites, the highest populations of M. scutellaris were always observed at sites where multiple releases had been made. Not all sites had M. scutellaris population densities that would be considered damaging, but their survival year-on-year indicates that this could be achieved in future. Damage can be difficult to assess at an early stage (Miller et al. 2019).
Inundative techniques have yielded unprecedented success at two large, hypertrophic, and economically important reservoirs in the Highveld region of South Africa. The Hartbeespoort and Roodeplaat dams had regular releases of M. scutellaris between 2019 and 2020 totalling around 238 000 insects released between the dams from 52 individual release events. Due to the increased release effort, the cover of water hyacinth significantly reduced at both sites, with surface cover decreasing at the Hartbeespoort Dam from over 37% to less than 6% during the summer time (pers obs; Coetzee et al. 2022). Elsewhere in Africa, success has been achieved on Lake Victoria through the inundative release of Neochetina spp. (Wilson et al. 2007). Therefore, inundation has a record of success in the literature, and should be regarded as the best practice for future release strategies. Selecting a candidate site to implement a mass rearing strategy must be carefully thought out, and not all sites will be suitable, as inundation requires a significant allocation of insect and human resources to these sites. Therefore, before applying inundative techniques, one must determine if the site is likely to be disturbed by other management methods, such as herbicide applications or major manual/mechanical removal initiatives that could destroy the populations of biological control agents. Additionally, it is best to ensure that the water body is large enough so that the hydrology and wind effect will allow the mat to break up as the agents cause damage, thus enabling the dying plants to sink (Hill and Olckers 2001).
Measuring the success of a biological control programme can be difficult, especially with floating aquatic plants that move with currents and wind. Due to the size of these projects and the high degree of stochasticity, metrics of success are often imprecise and, therefore, difficult to describe quantitatively (Hoffmann et al. 2019). Furthermore, post-release studies are often neglected due to most of the available funding being directed towards the pre-release assessment of a prospective agent (Schaffner et al. 2020). The types of data collected and the terminology used to classify success are usually case-specific and inconsistent, making accurate comparisons difficult (Moran et al. 2021). In systems invaded by water hyacinth, there is a great need to slow the rate of invasion to reduce damage and limit seed production. This often makes collecting exact empirical data difficult, as several stakeholders are often involved in the release processes. In contrast, the release sites are often varied, subject to disturbances, and spatially separate. New avenues are being investigated using combinations of citizen science data to help monitor plant and insect populations. In addition, remote sensing (using Sentinel-2 satellite imagery) enables users to visualise the site-wide changes in plant coverage to more accurately measure changes over time in relation to biological control agent populations (Singh et al. 2020; Hill et al. 2021; Coetzee et al. 2022).
CONCLUSION
Hill and Olckers (2001) recommended considering the importation and release of additional biological control agents to improve the prospects for water hyacinth management. Megamelus scutellaris has proved to be a valuable addition to the suite of biological control agents already released in South Africa, as it is very damaging when at high densities and easy to rear. The data presented in this paper show that M. scutellaris can establish in a wide range of climates and be effective at high altitude and highly eutrophic sites, where it has contributed to significantly reducing the coverage of water hyacinth on several large dams (Coetzee et al. 2022). The current recommended best practice for releasing M. scutellaris is regular releases as part of a strategic and adaptive management plan that includes all stakeholders (Hill et al. 2021), and these data show that various release protocols can lead to establishment. However, major success requires a more targeted approach (Coetzee et al. 2022). Future research should aim to develop more efficient rearing and release protocols to produce the greatest numbers of healthy M. scutellaris for the lowest cost to promote more regular releases in South Africa, while cost minimising will also enable M. scutellaris to be prioritised for release in other African countries where water hyacinth also has devastating consequences.
ACKNOWLEDGEMENTS
The National Research Foundation SARCHi and the Natural Resource Management Programme of the South African Department of Forestry, Fisheries, and the Environment are acknowledged for funding this research. Any opinion, conclusion or recommendation in this material is that of the authors and the NRF does not accept any liability in this regard.
ORCID IDs
Benjamin Miller - https://orcid.org/0000-0002-1409-690X
Julie Coetzee - https://orcid.org/0000-0002-0364-3349
Martin Hill - https://orcid.org/0000-0003-0579-5298
REFERENCES
Bashir MO, El Abjar ZE, Irving NS. 1984. Observations on the effect of the weevils Neochetina eichhorniae Warner and Neochetina bruchi Hustache on the growth of water hyacinth. Hydrobiologia. 110(1):95-98. https://doi.org/10.1007/BF00025780. [ Links ]
Bickford D, Lohman DJ, Sodhi NS, Ng PKL, Meier R, Winker K, Ingram KK, Das I. 2007. Cryptic species as a window on diversity and conservation. Trends in ecology & evolution (Amsterdam) 22(3):148-155. https://doi.org/10.1016/j.tree.2006.11.004. [ Links ]
Byrne M, Hill MP, Robertson M, King A, Jadhav A, Katembo N, Wilson J, Brudvig R, Fisher J. 2010. Integrated management of water hyacinth in South Africa: development of an integrated management plan for water hyacinth control, combining biological control, herbicidal control and nutrient control, tailored to the climatic regions of South Africa. Water Research Commission TT 454/10: 1-302. [ Links ]
Coetzee JA. 2013. Permission to release the planthopper, Megamelus scutellaris Berg. (Hemiptera: Delphacidae), a potential biological control agent of water hyacinth, Eichhornia crassipes. Makhanda: Department of Zoology & Entomology, Rhodes University. pp 1-34. [ Links ]
Coetzee JA, Hill MP. 2012. The role of eutrophication in the biological control of water hyacinth, Eichhornia crassipes, in South Africa. BioControl. 57(2):247-261. https://doi.org/10.1007/s10526-011-9426-y. [ Links ]
Coetzee JA, Bownes A, Martin GD, Miller BE, Smith R, Weyl PSR, Hill MP. 2021. A review of the biocontrol programmes against aquatic weeds in South Africa. African Entomology. 29(3):935-964. https://doi.org/10.4001/003.029.0935. [ Links ]
Coetzee JA, Byrne MJ, Hill MP. 2007. Predicting the distribution of Eccritotarsus catarinensis, a natural enemy released on water hyacinth in South Africa. Entomologia Experimentalis et Applicata. 125(3):237-247. https://doi.org/10.1111/j.1570-7458.2007.00622.x. [ Links ]
Coetzee JA, Hill MP, Byrne MJ. 2008. Ten years after the release of the water hyacinth mirid Eccritotarsus catarinensis in South Africa: what have we learnt? In: Anonymous Proceedings of the XII International Symposium on Biological Control ofWeeds, La Grande Motte, France, 22-27 April 2007. Wallingford: CAB International. Pp 512-515. [ Links ]
Coetzee JA, Miller BE, Kinsler D, Sebola K, Hill MP. 2022. It's a numbers game: inundative biological control of water hyacinth (Pontederia crassipes), using Megamelus scutellaris (Hemiptera: Delphacidae) yields success at a high elevation, hypertrophic reservoir in South Africa. Biocontrol Science and Technology. 32(11):1302-1311. https://doi.org/10.1080/09583157.2022.2109594. [ Links ]
Denno RF, Roderick GK. 1992. Density-related dispersal in planthoppers: effects of interspecific crowding. Ecology. 73(4):1323-1334. https://doi.org/10.2307/1940679. [ Links ]
Fitzgerald D, Tipping PW. 2013. Effect of insect density and host plant quality on wing-form in Megamelus scutellaris (Hemiptera: delphacidae). Florida Entomologist. 96(1):124-130. https://doi.org/10.1653/024.096.0116. [ Links ]
Foley JR, Minteer C, Tipping PW. 2016. Differences in seasonal variation between two biotypes of Megamelus scutellaris (Hemiptera: Delphacidae), a biological control agent for Eichhornia crassipes (Pontederiaceae) in Florida. Florida Entomologist. 99(3):569-571. https://doi.org/10.1653/024.099.0340. [ Links ]
Goode ABC, Tipping PW, Minteer CR, Pokorny EN, Knowles BK, Foley JR, Valmonte RJ. 2021. Megamelus scutellaris (Berg) (Hemiptera: Delphacidae) biology and population dynamics in the highly variable landscape of southern Florida. Biological Control. 160:104679. https://doi.org/10.1016/j.biocontrol.2021.104679. [ Links ]
Grevstad FS. 1999. Factors Influencing the chance of population establishment: implications for release strategies in biocontrol. Ecological Applications. 9(4):1439-1447. https://doi.org/10.1890/1051-0761(1999)009[1439:FITCOP]2.0.CO;2. [ Links ]
Griffith TC, Paterson ID, Owen CA, Coetzee JA. 2019. Thermal plasticity and microevolution enhance establishment success and persistence of a water hyacinth biological control agent. Entomologia Experimentalis et Applicata. 167(7):616-625. https://doi.org/10.1111/eea.12814. [ Links ]
Grodowitz MJ, Harms NE, Freedman JE. 2017. The influence of fluctuating temperature on Megamelus scutellaris Berg (Hemiptera: Delphacidae). US Army Engineer Research and Development Center, Vicksburg, MS. 39180: 1-5.
Grodowitz MJ, Johnson SJ, Harms NE. 2014. The Use of Megamelus scutellaris Berg in the southern United States as a biocontrol agent of water hyacinth (Eichhornia crassipes (Mart.)). U.S. Army Engineer Research and Development Center, Vicksburg, MS, 39180: 1-14.
Harms NE, Knight IA, Pratt PD, Reddy AM, Mukherjee A, Gong P, Coetzee J, Raghu S, Diaz R. 2021. Climate mismatch between introduced biological control agents and their invasive host plants: improving biological control of tropical weeds in temperate regions. Insects. 12(6):549. https://doi.org/10.3390/insects12060549. [ Links ]
Hill MP, Coetzee JA. 2017. The biological control of aquatic weeds in South Africa: current status and future challenges. Bothalia. 47(2):e1-e12. https://doi.org/10.4102/abc.v47i2.2152. [ Links ]
Hill MP, Conlong D, Zachariades C, Coetzee JA, Paterson ID, Miller BE, Foxcroft L, Van Der Westhuizen L. 2021. The role of mass-rearing in weed biological control projects in South Africa. African Entomology. 29(3):1030-1044. https://doi.org/10.4001/003.029.1030. [ Links ]
Hill MP, Cilliers CJ. 1999. A review of the arthropod natural enemies, and factors that influence their efficacy, in the biological control of water hyacinth, Eichhornia crassipes (Mart.) Solms-Laubach (Pontederiaceae), in South Africa. African Entomology. Memoir. (1):103-112.
Hill MP, Olckers T. 2001. Biological control initiatives against water hyacinth in South Africa: constraining factors, success and new courses of action. In: Second Meeting of the Global Working Group for the Biological and Integrated Control of Water Hyacinth, Beijing, China. ACIAR Proceedings: 33-38. [ Links ]
Hoffmann JH, Moran VC, Hill MP. 2019. Conceptualizing, categorizing and recording the outcomes of biological control of invasive plant species, at a population level. Biological Control. 133:134-137. https://doi.org/10.1016/j.biocontrol.2019.02.005. [ Links ]
Jones RW. 2014. Aquatic Invasions of the Nseleni River System: Causes, Consequences and Control. PhD Thesis, Rhodes University, South Africa. [ Links ]
May B, Coetzee JA. 2013. Comparisons of the thermal physiology of water hyacinth biological control agents: predicting establishment and distribution pre-and post-release. Entomologia Experimentalis et Applicata. 147(3):241-250. https://doi.org/10.1111/eea.12062. [ Links ]
McFadyen REC. 1998. Biological control of weeds. Annual Review of Entomology. 43(1):369-393. https://doi.org/10.1146/annurev.ento.43.1.369. [ Links ]
Miller BE, Coetzee J, Hill M. 2019. Chlorophyll fluorometry as a method of determining the effectiveness of a biological control agent in post-release evaluations. Biocontrol Science and Technology. 29(11):1118-1122. https://doi.org/10.1080/09583157.2019.1656165. [ Links ]
Miller BE, Coetzee JA, Hill MP. 2021. Mind the gap: the delayed recovery of a population of the biological control agent Megamelus scutellaris Berg. (Hemiptera: Delphacidae) on water hyacinth after winter. Bulletin of Entomological Research. 111(1):120-128. https://doi.org/10.1017/S0007485320000516. [ Links ]
Moran VC, Zachariades C, Hoffmann JH. 2021. Implementing a system in South Africa for categorizing the outcomes of weed biological control. Biological Control. 153:104431. https://doi.org/10.1016/j.biocontrol.2020.104431. [ Links ]
Mukherjee A, Knutson A, Hahn DA, Heinz KM. 2014. Biological control of giant salvinia (Salvinia molesta) in a temperate region: cold tolerance and low temperature oviposition of Cyrtobagous salviniae. BioControl (Dordrecht, Netherlands). 59(6):781-790. https://doi.org/10.1007/s10526-014-9617-4. [ Links ]
Paterson ID, Coetzee JA, Weyl P, Griffith TC, Voogt N, Hill MP. 2019. Cryptic species of a water hyacinth biological control agent revealed in South Africa: host specificity, impact, and thermal tolerance. Entomologia Experimentalis et Applicata. 167(7):682-691. https://doi.org/10.1111/eea.12812. [ Links ]
Peel MC, Finlayson BL, McMahon TA. 2007. Updated world map of the Köppen-Geiger climate classification. Hydrology and Earth System Sciences. 11(5):1633-1644. https://doi.org/10.5194/hess-11-1633-2007. [ Links ]
Porter JD, Owen CA, Compton SG, Coetzee JA. 2019. Testing the thermal limits of Eccritotarsus catarinensis: a case of thermal plasticity. Biocontrol Science and Technology. 29(6):565-577. https://doi.org/10.1080/09583157.2019.1572712. [ Links ]
Pratt PD, Moran PM, Pitcairn MJ, Reddy A, O'Brien J. 2021. Biological control of invasive plants in California's Delta: Past, present, and future. Journal of Aquatic Plant Management. 59(S):55-66. [ Links ]
Reddy AM, Pratt PD, Hopper JV, Cibils-Stewart X, Walsh GC, Mc Kay F. 2019. Variation in cool temperature performance between populations of Neochetina eichhorniae (Coleoptera: Curculionidae) and implications for the biological control of water hyacinth, Eichhornia crassipes, in a temperate climate. Biological Control. 128:85-93. https://doi.org/10.1016/j.biocontrol.2018.09.016. [ Links ]
Schaffner U, Hill M, Dudley T, D'Antonio C. 2020.Post-release monitoring in classical biological control of weeds: assessing impact and testing pre-release hypotheses. Current Opinion in Insect Science. 38:99-106. https://doi.org/10.1016/j.cois.2020.02.008. [ Links ]
Singh G, Reynolds C, Byrne M, Rosman B. 2020. A Remote Sensing Method to Monitor Water, Aquatic Vegetation, and Invasive Water Hyacinth at National Extents. Remote Sensing (Basel). 12(24):4021. https://doi.org/10.3390/rs12244021. [ Links ]
Sosa AJ, Lenicov AMMDR, Mariani R, Cordo HA. 2005. Life history of Megamelus scutellaris with description ofimmature stages (Hemiptera: Delphacidae). Annals of the Entomological Society of America. 98(1):66-72. https://doi.org/10.1603/0013-8746(2005)098[0066:LHOMSW]2.0.CO;2. [ Links ]
Sosa AJ, Cordo HA, Sacco J. 2007. Preliminary evaluation of Megamelus scutellaris Berg (Hemiptera: Delphacidae), a candidate for biological control of waterhyacinth. Biological Control. 42(2):129-138. https://doi.org/10.1016/j.biocontrol.2007.04.012. [ Links ]
Sutton GF, Compton SG, Coetzee JA. 2016. Naturally occurring phytopathogens enhance biological control of water hyacinth (Eichhornia crassipes) by Megamelus scutellaris (Hemiptera: Delphacidae), even in eutrophic water. Biological Control. 103:261268. https://doi.org/10.1016/j.biocontrol.2016.10.003. [ Links ]
Tipping PW, Center TD, Sosa AJ, Dray FA. 2011. Host specificity assessment and potential impact of Megamelus scutellaris (Hemiptera: Delphacidae) on water hyacinth Eichhornia crassipes (Pontederiales: Pontederiaceae). Biocontrol Science and Technology. 21(1):75-87. https://doi.org/10.1080/09583157.2010.525739. [ Links ]
Tipping PW, Martin MR, Pokorny EN, Nimmo KR, Fitzgerald DL, Dray FA Jr, Center TD. 2014a. Current levels ofsuppression ofwater hyacinth in Florida USA by classical biological control agents. Biological Control. 71:65-69. https://doi.org/10.1016/j.biocontrol.2014.01.008. [ Links ]
Tipping PW, Sosa A, Pokorny EN, Foley J, Schmitz DC, Lane JS, Rodgers L, McCloud L, Livingston-Way P, Cole MS, et al. 2014b. Release and establishment of Megamelus scutellaris (Hemiptera: Delphacidae) on water hyacinth in Florida. Florida Entomologist. 97(2):804-806. https://doi.org/10.1653/024.097.0264. [ Links ]
Venter N, Hill MP, Hutchinson SL, Ripley BS. 2013. Weevil borne microbes contribute as much to the reduction of photosynthesis in water hyacinth as does herbivory. Biological Control. 64(2):138-142. https://doi.org/10.1016/j.biocontrol.2012.10.011. [ Links ]
Wilson JRU, Ajuonu O, Center TD, Hill MP, Julien MH, Katagira FF, Neuenschwander P, Njoka SW, Ogwang J, Reeder RH, et al. 2007. The decline of water hyacinth on Lake Victoria was due to biological control by Neochetina spp. Aquatic Botany. 87(1):90-93. https://doi.org/10.1016/j.aquabot.2006.06.006. [ Links ]
Zera AJ, Denno RF. 1997. Physiology and ecology of dispersal polymorphism in insects. Annual Review of Entomology. 42(1):207-230. https://doi.org/10.1146/annurev.ento.42.1.207. [ Links ]
Zhou Z, Guo J, Ai H, Li M, Wan F. 2011. Rapid cold-hardening response in Ophraella communa LeSage (Coleoptera: Chrysomelidae), a biological control agent of Ambrosia artemisiifolia L. Biocontrol Science and Technology. 21(2):215-224. https://doi.org/10.1080/09583157.2010.534549. [ Links ]
 Correspondence:
Correspondence:
Benjamin E Miller
Email: miller.benjamin93@gmail.com
Received: 12 February 2023
Accepted:16 August 2023


{kind=link}
{kind=link}
{kind=link}