










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAfrican Entomology
On-line version ISSN 2224-8854
Print version ISSN 1021-3589
AE vol.31 Pretoria 2023
http://dx.doi.org/10.17159/2254-8854/2023/a14047
RESEARCH ARTICLE
Commercial banana and macadamia plantations in a savanna matrix support high levels of arthropod diversity
Vhuawelo SimbaI; Inam YekwayoII; Tarombera MwabvuIII, IV
ISchool of Agricultural Sciences, Faculty of Agriculture and Natural Sciences, University of Mpumalanga, Mbombela, South Africa
IIDepartment of Biological and Environmental Sciences, Walter Sisulu University, Mthatha, South Africa
IIISchool of Biology and Environmental Sciences, Faculty of Agriculture and Natural Sciences, University of Mpumalanga, Mbombela, South Africa
IVSchool of Life Sciences, University of KwaZulu-Natal, Durban, South Africa
ABSTRACT
Expansion and intensification of agroecosystems is one of the major causes of habitat loss in the savanna biome in South Africa. As such, this study sought to determine the influence of commercial subtropical fruit plantations (banana and macadamia) on species richness, abundance, and composition of surface-active arthropods compared to the savanna biome. Given that pesticides and herbicides are applied from spring to early autumn in banana and macadamia plantations, we sampled in winter to reduce the potential impact of pesticides and herbicides. Surface-active arthropods were sampled using pitfall traps. Habitat type did not affect species richness and abundance of ants and spiders, as well as species richness of beetles. However, significantly greater abundance of beetles was recorded in the macadamia plantation compared to the banana and savanna. This could have been due to greater abundance of herbivorous beetles and other insects, which would have increased the diversity of predatory beetles. Furthermore, unlike the banana plantation, the macadamia plantation was characterised by a deep leaf litter layer and the presence of weeds and grasses, which probably increased abundance of beetles. Species composition indicated that the studied arthropod taxa associate with specific habitats, as demonstrated by the three habitats supporting different species composition. Despite savanna habitat not supporting high species richness or abundance of surface-active arthropods, we recorded the highest number of unique species of ants and spiders in the savanna rather than in the plantations. These results highlight the importance of natural landscapes in conservation of surface-active arthropods.
Keywords: agroecosystem, ants, beetles, diversity, natural landscapes, spiders
INTRODUCTION
The vegetation in the savanna biome is characterised by high diversity of plant species composed of grasses, shrubs, and scattered trees (Rutherford et al. 2006). The mixture of herbaceous and woody plants is supported by the summer rainfall and gives the savanna biome a particularly complex structure (Rutherford et al. 2006), providing diverse microhabitats for many arthropod species (Botha et al. 2016). Different arthropod taxa prefer different environmental and habitat conditions (Yekwayo et al. 2017); therefore, high diversity of arthropods is likely to be associated with greater habitat heterogeneity. The availability of microhabitats, such as debris and logs, resulting from high diversity of plants in the savanna vegetation, increases food resources and nesting sites for surface-active arthropods (Loyola et al. 2006). Several studies have documented different surface-active arthropods, including, millipedes, scorpions, centipedes (Druce et al. 2007), termites (Doube 2018), ants, spiders, and millipedes (Mwabvu & Yekwayo 2020), and dung beetles (La Scaleia et al. 2018) in the savanna. Surface-active arthropods provide different ecosystem services that are crucial for the functioning of the savanna ecosystems (Marchant 2010) while also benefitting humans (Samways et al. 2012).
Despite the vital role of the savanna in biodiversity conservation, about 50% of savanna landscapes are now converted into agroecosystems to meet the demand of the growing human population (Lichtenberg et al. 2017; Riggio et al. 2013). Although the rate of conversion of the savanna biome is high, the low levels of endemicity and the high number of protected areas within this biome makes the savanna biome a low priority for biodiversity conservation (Driver et al. 2005). However, the endemicity level and conservation status of the savanna biome is based on assessments of plants, birds, and mammals (Driver et al. 2005), with relatively fewer studies focusing on arthropod groups, such as spiders (Cardoso et al. 2019; Foord et al. 2020).
Surface-active arthropods are sensitive to changes that occur in their habitats (Lichtenberg et al. 2017). Changes in habitats that are associated with the reduction in plant diversity decrease food resources and nesting sites for arthropods (Rhoades et al. 2018). Marasas et al. (2010) recorded low species richness and abundance of carabid beetles in wheat plots compared to semi-natural plots, which are likely to have more resources for surface-active arthropods. However, contrasting findings were reported by Magagula & Samways (2001), who found greater density and species richness of ladybirds in citrus orchards than in natural habitats, even though there were species unique to the natural habitats. Ladybirds dominated citrus orchards because of high abundance of prey and the high dispersal ability of some species, which allowed movement across habitats (Magagula & Samways 2001). However, many species of surface-active arthropods are characterised by low dispersal ability, as such the species may not be able to move to alternative habitats (Yekwayo et al. 2017). Thus, surface-active arthropods are affected by anthropogenic activities that result in habitat fragmentation (Moir et al. 2005).
In addition to habitat loss, intensive management practices, which are intended to increase yields in agroecosystems, can affect the diversity of surface-active arthropods negatively (Isbell et al. 2017). The application of pesticides may kill non-targeted beneficial surface-active arthropods (Olfert et al. 2002). For example, Hafsi et al. (2016) observed greater abundance of the Coccinellidae in peach plots that were not treated with pesticides than those that were treated. In addition, the use of herbicides along the plantation edges and between rows of crop plants reduces microhabitats for some surface-active arthropods (Sarabi 2019). Furthermore, high abundance of predatory arthropods was recorded in experimental plots that had weeds between apple trees compared to control plots that had no weeds (Wyss 1992). Variation in leaf litter depth also influences temperature, moisture, and prey abundance (Halaj et al. 2008). Therefore, clearing of leaf litter in agroecosystems can also affect surface-active arthropods negatively. For instance, in a wet tropical forest there was greater diversity of surface-active arthropods in sites that had a thick layer of leaf litter compared with sites that were bare (Sayer et al. 2006). However, in KwaZulu-Natal, Eckert et al. (2019) found that arthropod assemblages in grasslands and pine plantations were not influenced by the percentage of leaf litter cover. Thus, assessing communities of surface-active arthropods in agroecosystems is important for agricultural sustainability and mitigation of the negative effects of agricultural practices on ecosystem services.
Ecosystem services provided by surface-active arthropods can improve the production of crops in agroecosystems. These ecosystem services include, pollination (Bommarco et al. 2012), cycling of nutrients (Doube 2018), decomposition of organic matter (Ahmad & Ahmad 2009), control of pests (Poeydebat et al. 2017) and predation (Marie et al. 2018). For example, ants and certain beetles were recorded as pollinators in mango orchards in South Africa (Simba et al. 2018). Furthermore, dung beetles associate with specific mammalian dung, and protected areas are known to support greater diversity of mammals than forest plantations (Pryke et al. 2022). As such, the occurrence of high species richness of dung beetles in habitats with greater plant diversity, such as protected areas and wider corridors (van Schalkwyk et al. 2017) is not surprising considering the contribution of dung beetles in plant growth (Sitters et al. 2014). When these dung beetles remove and bury dung, they increase carbon and nitrogen storage in soils (Sitters et al. 2014). In another study, Pryke et al. (2013) recorded greater diversity of dung beetles in grassland and pine plantations compared to the natural forest. The activity of dung beetles and termites in the soil influences plant growth indirectly by improving the texture of the soil, aeration in the soil and infiltration of water (Doube 2018). Furthermore, even though activities of termites lead to the formation of mounds, there are plant species that have been reported to grow on those mounds (Kelly & Samways 2011), which may increase vegetation heterogeneity. Other important surface-active arthropods include natural enemies of pests, such as weaver ants that control the infestation of bagworm in oil palm orchards (Pierre & Idris 2013) and ground beetles that control the olive fruit fly in olive orchards (Albertini et al. 2017). Unfortunately, agroecosystems that are managed intensively can reduce the diversity of arthropods unlike natural landscapes (Luke et al. 2014).
Amongst other agroecosystems, subtropical fruit plantations have been expanding rapidly (Granatstein et al. 2016; Pio et al. 2018) because of their high contribution to the economies of countries (Campbell 2018). Banana and macadamia are among the most widely produced subtropical fruits that contribute significantly to the economy of South Africa (Diczbalis et al. 2014). Although arthropods have been studied in subtropical fruit plantations, the focus has been on arthropods as pollinators (Bommarco et al. 2012), pests and their natural enemies (Grout et al. 2001; Pierre & Idris 2013). In addition, checklists of spiders have been produced in South African agroecosystems (Dippenaar-Schoeman et al. 2001, 2005, 2013). However, ecological data on ants, beetles and spiders in banana and macadamia, particularly in South Africa, remain limited. Therefore, this study aimed to determine if species richness, abundance and composition of ants, beetles and spiders will differ between the two fruit plantations that are under specific agricultural practices. Furthermore, we compared species richness, abundance, and composition of the three taxa between the fruit plantations and the savanna vegetation. Several studies have reported increased diversity of surface-active arthropods in natural vegetation because of greater habitat heterogeneity, which is driven by the high diversity of plants (Mauda et al. 2018; Rhoades et al. 2018; Marasas et al. 2010). Therefore, we expected high species richness and abundance of ants, beetles and spiders in the savanna vegetation compared with the two fruit plantations. Furthermore, we expected the species composition of the three taxa to vary amongst the three habitat types.
MATERIAL AND METHODS
Study site
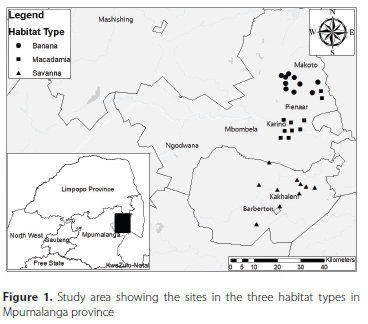
Surface-active arthropods were sampled from three habitat types, two in the Agricultural Research Council farms in Mbombela (macadamia) and Hazyview (banana and macadamia), and the savanna in Barberton Nature Reserve in the Mpumalanga province, South Africa (Figure 1). The two farms in Hazyview and in Mbombela are about 66 km apart. Barberton Nature Reserve is about 54 km from the farm in Mbombela and about 88 km from the farm in Hazyview. The macadamia and banana plots in Hazyview are between 15 and 20 years in age, while the macadamia trees in Mbombela are about 12 years old. Leaf litter depth was measured using a ruler. In the banana plantation leaf litter depth was 1.7-13.5 cm and in the macadamia plantation it was 4.5-14.7 cm, while in the savanna leaf litter depth was not measured due to its sparse distribution. However, leaf litter in macadamia sites in Hazyview was swept from under the tree canopies and piled up between the rows. Furthermore, the macadamia sites in Mbombela had grasses and up to six herbaceous plant species between rows of the trees, nlike banana and macadamia plantations in Hazyview which had no herbaceous plants nor grasses. The sites in Barberton Nature Reserve (BNR) were characterised by tall and short grass, scattered trees, and shrubs. In addition, BNR is rocky with some areas having steep slopes and dominated by shrubs of Vachellia spp.
The watering practices as well as control of pests and weeds in the two plantations differ between the two farms. Watering of the macadamia and banana plants in Hazyview occurs fortnightly in the dry winter compared with watering of the macadamia trees in Mbombela which occurs at higher frequency (weekly) between August and February. Both plantations in the two farms applied herbicides (glyphosate and paraquat) between September and March. However, in addition to the herbicides, two pyrethroid pesticides (beta-cyfluthrin and alpha cypermethrin) were applied in Mbombela.
Study design
Each habitat type had ten sites that were 10 m χ 10 m in size, making a total of 30 sites in the study (Figure 1). The sites in macadamia plantation and savanna vegetation were at least 300 m away from each other to prevent false replication (see Clark & Samways 1997). The banana plants are planted in smaller blocks that are separated by dirt roads that are about 8 m wide. Each block (minimum size about 8 000 m2) had a single site; thus, the roads were physical barriers between the sites; as Samways et al. (2010) indicated, sites or samples that are separated by a physical or geographical barrier can be considered independent of each other. However, we acknowledge the limitation (that we were unable to avoid) of having all banana sites in a single farm, as this may imply false replication (see Samways et al. 2010). To eliminate the edge effect, we adopted Gallé et al. (2020) methods and placed sites in macadamia plantation 10 m away from the edge, while sites in the banana were at the centre of the block. In all the vegetation types, each site was divided into four line transects, adjacent transects were about 2 m apart. In each line transect there were six pitfall traps, adjacent pitfall traps were about 2 m apart.
Sampling method
The spraying of pesticides and herbicides occurred between September and March in the two farms. Our target was to sample outside of the spraying season in order to ensure that the effects of pesticides on the activity of surface-active arthropods was reduced. As a result, sampling took place in July 2020. The pitfall trapping method was used because it is the most effective for sampling surface-active arthropods (Woodcock 2005; Samways et al. 2010). We sampled in three consecutive sampling events to ensure that the samples were representative in the study areas. Furthermore, sampling in three events increased the probability of capturing different species of ants, beetles and spiders, which are known to be active at different times depending on the behaviour of the species and environmental conditions (Samways et al. 2010). The pitfall traps were placed in the same location for all three sampling events. Each pitfall trap was half-filled with 50% ethylene glycol solution and inserted in the ground leaving the open end at the same level as the soil surface. Pitfall traps were collected after seven days because it is the appropriate minimum period for sampling arthropods (see Borgelt & New 2006). Traps were emptied into collection bottles, which had 100% ethyl alcohol. In the laboratory, specimens were extracted from the debris and stored in 70% ethyl alcohol. Species were identified to family and genus levels where possible, and assigned to morpho-species. Arthropods were identified using different guides (Dippenaar-Schoeman & Jocqué 1997; White 1998; Bouchard 2014; Dippenaar-Schoeman 2014; Fisher & Bolton 2016; Picker et al. 2019). Voucher specimens were housed temporarily in the Soil Invertebrates Laboratory at the University of Mpumalanga, and will be deposited for long-term storage at the KwaZulu-Natal Museum, Pietermaritzburg, South Africa. Leaf litter depth, which was measured randomly at four points in each site using a ruler was averaged.
Statistical analyses
The samples from each sampling event, at each site were pooled for analyses. Data were analysed separately for each of the three taxa (ants, beetles and spiders). Due to the high number of singletons and doubletons in our datasets, we used two non-parametric estimators (Chao2 and Jackknife2) to determine the effectiveness of our sampling (Samways et al. 2010). Species richness was estimated in PRIMER 7.
Species richness and abundance datasets of each taxon were tested for normality using the Shapiro-Wilk test in R (R Core Team 2020). Given the close proximity of our sites, especially in the banana plantation, we accounted for spatial autocorrelation by using the generalised least squares (GLS) and generalised linear mixed models (GLMM) with the penalised quasilikelihood estimation (Dorman et al. 2007). Species richness of ants and spiders, as well as abundance of spiders were analysed using GLS because these data were normally distributed. However, GLMMs were used for species richness of beetles and abundance of ants as well as beetles because these data were not normally distributed. The MASS package (Ripley et al. 2013) and Poisson distribution (Bolker et al. 2009) were used in GLMMs. In the models, GLS and GLMM, the longitude and latitude were included as a dummy variable with an exponential correlation (Dormann et al. 2007), which was the random factor. The type of habitats, banana, macadamia and savanna, was included in models as an explanatory variable. Boxplots were used to display significant differences in species richness and abundance among the three habitat types. Spearman's rank correlations in R (Crawley 2005) were used to determine significant correlation between response variables, species richness and abundance, of each taxon and leaf litter depth.
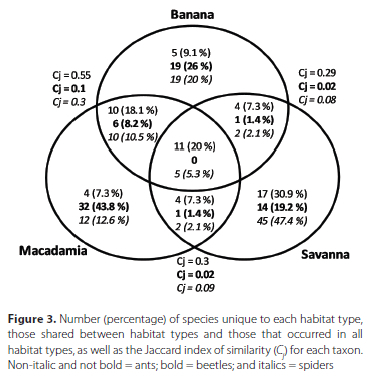
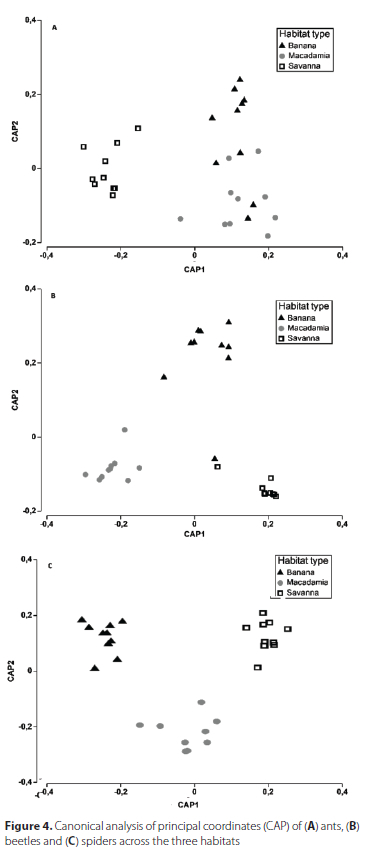
Variation in the composition of ants, beetles and spiders among the three habitat types were assessed with a multivariate generalised linear model using the "manyGLM" function in the mvabund package (Wang et al. 2012) in R. Arthropod abundance data were modelled as the response variable with a negative binomial distribution used as the best fit. Habitat type was modelled as an explanatory variable. The species composition of each taxon was visualised using the canonical analysis of principal coordinates in PRIMER 7 (Anderson & Willis 2003). In addition, similarities and dissimilarities in species unique to each habitat and those shared between and/or among habitats were visualised using Venn diagrams. Furthermore, the Jaccard index of similarity was calculated for each pair of habitat types. The formula used to calculate the Jaccard index of similarity is C. = j/ (a + b - j), where C is the Jaccard index of similarity, j is the number of species shared between two habitats, a is the number of species unique to the first habitat, and b refers to the number of species unique to the second habitat.
RESULTS
Ants had the highest number of individuals (4 368), followed by spiders (1 031) and beetles (733). The species accumulation curves did not reach an asymptote for ants (observed = 55, Choa2 = 63.45 ± 6.13, Jacknife2 = 70.69), beetles (observed = 73, Choa2 = 198 ± 54.19, Jacknife2 = 158.99) and spiders (observed = 95, Choa2 = 263.2 ± 70.35, Jacknife2 = 196.19). Furthermore, beetles had the highest number of singletons and doubletons contributing 73% of the total number of morpho-species, followed by spiders at 63%, while ants had the lowest percentage (27%).
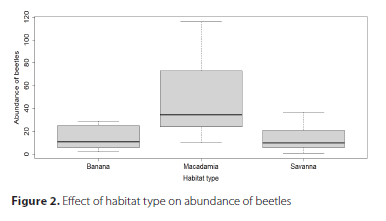
The three taxa showed varying responses to the type of habitat. Significantly greater abundance of beetles was recorded in the macadamia compared with the savanna and the banana (Table 1, Figure 2). Although species richness of beetles was nearly significant (p = 0.046), the macadamia had the highest percentage of unique morpho-species (43.8%) compared with the banana and the savanna (Figure 3). Even though no significant differences were recorded in species richness and abundance of ants and spiders among the three habitats (Table 1), the savanna had the highest percentage of unique species for both taxa (Figure 3).
Spearman's rank correlation showed that the effect of leaf litter depth on species richness and abundance is taxon-dependent. Species richness (r = 0.58, p = 0.0007) and abundance (r = 0.43, p = 0.02) of beetles positively correlated with leaf litter depth. However, a negative correlation between species richness of ants and leaf litter depth (r = -0.41, p = 0.02) was observed. On the other hand, leaf litter depth was not correlated with the abundance of ants (correlation = -0.03, p = 0.87) and spiders (correlation = -0.18, p = 0.33) nor species richness of spiders (correlation = -0.19, p = 0.32).
The multivariate generalised linear models showed that species composition of ants, beetles and spiders was significantly different among the three habitat types (Table 1; Figures 3, 4). However, the Jaccard index of similarity (C = 0.55) and canonical analysis of principal coordinates displayed an overlap in assemblages of ants between the banana and macadamia plantations (Figures 3, 4a).
DISCUSSION
Contrary to our expectations, the savanna vegetation did not support significantly greater species richness nor abundance of arthropods compared to the two plantations. Instead, species richness and abundance of ants and spiders, as well as species richness of beetles did not differ among the three habitat types. Although there are specialist species of ants and spiders, there are many generalist predators also (Abera-Kalibata et al. 2007; Dippenaar-Schoeman et al. 2001, 2005; Herwina et al. 2013; Pekár & Toft 2015). Species richness and abundance of arthropods may have increased in different habitat types because of the predatory nature of spiders and some species of ants and beetles, which are affected mostly by prey availability. Subtropical fruit plantations support high abundance of herbivorous insects (Herwina et al. 2013; Michalko et al. 2019), which may increase the abundance and species richness of predatory spiders (Herwina et al. 2013; Michalko et al. 2019; Tsai & Pekár 2019). Similarly, Bolu & Özgen (2018) reported a positive correlation between the abundance of aphids and Camponotus species that prey on aphids. Furthermore, ants are species rich and abundant in agricultural, natural, and urban landscapes, where they nest in microhabitats, such as rotten trunks, twigs, logs, leaf litter, plant roots and underneath rocks (Fisher & Bolton 2016; Herrera et al. 2020). Similarly, some salticids and thomisids have been recorded underneath loose bark and beneath leaves (Dippenaar-Schoeman et al. 2001). Furthermore, diverse species of spiders thrive in natural habitats (Foord et al. 2011), as well as, in subtropical fruit plantations (Dippenaar-Schoeman et al. 2001, 2005).
Although there were no significant differences in species richness of ants or spiders among the three habitats that we studied, the savanna had the highest percentage of unique species for both taxa, unlike beetles which were most rich in the macadamia plantations. This high number of unique species in the savanna compared with the other habitat types could have been due to greater vegetation diversity and density in the savanna compared to the two plantations. Downie et al. (1995) found a positive correlation between the distribution of species of spiders and the vegetation density. The high percentage of ant and spider species that were restricted to the savanna emphasises the importance of natural landscapes in the conservation of arthropods. Previous studies have also reported a decrease in species richness of ants in cultivated monocultures compared to heterogeneous natural vegetation (Perfecto 1990; Roth et al. 1994; Perfecto & Snelling 1995).
Leaf litter depth is among the variables that influence species richness and abundance of some arthropods. For instance, Moses et al. (2021) reported that species richness of ants in an undisturbed forest in Papua New Guinea increased with leaf litter depth, while abundance of ants decreased with leaf litter depth. However, another study that compared arthropod abundance and diversity in plots with different levels and quality of leaf litter found that arthropod diversity did not differ between heterogeneous and homogenous leaf litter (Donoso et al. 2010). In our study, leaf litter depth did not influence species richness of ants and spiders, probably because of the generalist nature of these taxa. However, we recorded that species richness and abundance of beetles correlated positively with leaf litter depth. Out of the three habitats, the macadamia plantation was characterised by a deeper layer of leaf litter, which would have provided a suitable habitat for beetle species that require leaf litter. Furthermore, a deep layer of leaf litter influences beetles, such as, ground beetles because of the reduced ground temperature and increased availability of prey within the leaf litter (Magura et al. 2005). Greater leaf litter depth also increases the abundance of dung beetles (da Silva & Hernández 2016).
As such, high abundance and the number of unique species of beetles that we recorded in the macadamia may have been due to greater leaf litter in most of our study sites.
The greater abundance and number ofspecies in the macadamia were attributed to herbivorous beetles in the Curculionidae and Chrysomelidae, as well as predatory beetles (e.g. Coccinellidae) (Table S1). In addition, within the Curculionidae there is a seed weevil (Kuschelorhynchus macadamiae) that has been reported as a threat to macadamia production (Khun et al. 2020). The presence of coccinellids is highly dependent on the presence of their prey (Magagula & Samways 2001) that would be expected to occur in both plantations. Furthermore, most species of beetles in fruit orchards are pests and their natural enemies (Sutton et al. 2014), as such, it is not surprising that we recorded significantly greater abundance and greater percentage of species that were unique to the macadamia.
Despite that habitat type did not affect abundance and species richness of ants and spiders, as well as species richness of beetles, species composition of all the three taxa were affected significantly. Variation in species composition amongst the three habitat types indicates some level of specialisation even among predatory arthropods. For example, carabids, which are natural enemies of pests in agricultural crops (Matta et al. 2017; Wyckhuys & O'Neil 2006), were represented by three morpho-species, one unique to the savanna, the other to the macadamia, while the third one occurred in both the banana and macadamia plantations (Table S1). Our results supported studies, which reported variation in species composition of ants in banana, macadamia, citrus and palmito (Matlock & de la Cruz 2003), as well as between natural vegetation and agricultural landscapes (de Castro Solar et al. 2016).
The three habitat types in our studyhad different characteristics. The savanna vegetation had tall grasses, herbaceous plants, scattered shrubs, and trees; unlike the macadamia plantations which, besides the about 5 m tall macadamia trees, had grasses and herbaceous plants and a layer ofleaflitter (in some sites). The banana plantations had about 4 m tall banana plants growing in wet soil due to constant irrigation, a layer of leaf litter and scattered patches of bare soil. The differences in microhabitats within the three habitat types could have caused each habitat type to support different species of arthropods. Although both the banana and macadamia had leaf litter, the type and density were different, and may have caused the significant differences in assemblages between these plantations. However, it is important to note that the Jaccard index of similarity and the canonical analysis of principal coordinates revealed overlap of ant assemblages between the two plantations, thus supporting Mauda et al. (2018) who reported that cultivated landscapes often share dominant species of ants. The overlap of ant assemblages between the macadamia and banana plantations could be due to generalist species (such as, those in Pheidole and Tetramorium) that are adapted to disturbed areas (García-Martínez et al. 2015). Generalist predators adapt well because when disturbance decreases the abundance of the preferred prey they can feed on any other prey that is available (Kwon et al. 2013; Mollot et al. 2012). Furthermore, a shift in prey preference is possible for predators, such as ants, because many species nest and feed on a variety of items. However, Eckert et al. (2022) indicated that the effect of environmental variables on ants vary depending on the vegetation type. For instance, in a natural forest, leaf litter cover and soil pH were the important variables affecting ant assemblages, while in a grassland, variables that mattered were soil compaction, pH, carbon, nitrogen and sulphur (Eckert et al. 2022).
The dominance of lycosids in the fruit plantations may be due to them being early colonisers of disturbed areas (Samu & Szinetár 2002). Unlike many surface-active arthropods, lycosids have greater dispersal ability, which facilitates invasion of, and thriving in disturbed habitats (Saqib et al. 2020). Species that were unique and abundant in the banana included a morpho-species in the Anthicidae, two in Copa and one in the Oonopidae. Most species of the Anthicidae are detritivores that are associated with agroecosystems that have high plant residues on the soil surface (Araújo et al. 2018). Furthermore, the Anthicidae is associated with high soil moisture (Telnov & Ghahari 2018), which was readily available in the banana compared with the other habitats. Similarly, species of Copa (Haddad 2013) and the Oonopidae (Ubick et al. 2005) are associated with leaf litter. Furthermore, oonopids feed on mites and springtails (Tiwari et al. 2021), which were common in the banana plantation. Therefore, irrigation and the layers of leaf litter in the banana plantation may have increased and retained soil and leaf litter moisture, thereby providing suitable habitats for mites and springtails and increased microbes on which they fed. The Nitidulidae are known to feed on the sap in fruits (Emekci & Moore 2015) and macadamia trees produce a lot of sap (Olesen et al. 2008), so the four morpho-species of nitidulids may have been feeding on the sap in the macadamia. Two nitidulids were in the banana and none were recorded in the savanna (Table S1).
Morpho-species of ants that were restricted to the savanna included species in genera that are associated with nesting in the ground and/or under rocks, e.g., Bothroponera sp. (Joma & Mackay 2017), Lepisiota sp. (Sharaf et al. 2020); Ocymyrmex sp. and Polyrhachis sp. (Andersen et al. 2000). These genera of ants build colonies in the ground and under rocks, this may explain their association with the savanna habitat where there is less disturbance. Furthermore, the restriction of Odontomachus sp. to the savanna habitat could be because the species feed on a variety of invertebrates (Raimundo et al. 2009), small vertebrates (Facure & Giaretta 2009), plant and insect exudates, and nutrient-rich fleshy fruits (Passos & Oliveira 2002, 2004). Thus, the savanna vegetation is more likely to have the preferred food resources for Odontomachus sp. compared with the two plantations. Furthermore, the abundance of scarabids in the savanna is probably a result of the continuous input of organic matter, such as leaf litter (although scattered) and the presence of mammalian dung on which scarabids depend (Pompeo et al. 2016). Most scarabids are sensitive to anthropogenic activities (Barretto et al. 2019). Therefore, their occurrence in the savanna habitat, which is less disturbed, was not surprising. Although morpho-species in the Gnaphosidae were present in all habitat types, there were morpho-species unique to each, with the savanna having the highest number. The occurrence of gnaphosids in all habitat types could be attributed to the fact that they are ant-eating spiders (Pekár 2004), and ants were abundant in the three habitats.
Our findings indicate that the conversion of savanna landscapes into subtropical fruit plantations does not reduce species richness and abundance of some predatory arthropods (ants and spiders). However, subtropical fruit plantations alter species composition of ants, beetles and spiders, as we found differences in assemblages among the three habitat types. Our results demonstrate that maintenance of diverse microhabitats in agroecosystems enhances arthropod conservation (see Geldenhuys et al. 2021). Considering the increasing habitat modification, and the importance and global decline in biological diversity, management practices in subtropical fruit plantations are critical in conserving surface-active arthropods.
ACKNOWLEDGEMENTS
We wish to thank the National Research Foundation (Competitive Support for Unrated Researchers - Grant number: 134117) for funding this project. Special thanks to the Mpumalanga Tourism and Parks Agency and the Agricultural Research Council (ARC), Mbombela and Hazyview, for granting us access to their properties. In addition, all field assistants, namely; Ntombikayise Ndwandwe, Sizwe Sambo, Simphiwe Sibeko, Sithabile Ndwandwe and Simphiwe Sibitane are thanked.
AUTHORS' CONTRIBUTIONS
Vhuawelo Simba: investigation, methodology, formal analyses, data curation, writing (original draft).
Inam Yekwayo: conceptualisation, methodology, formal analyses, data curation, funding acquisition, resources, investigation, project administration, supervision, validation, writing (review and editing).
Tarombera Mwabvu: conceptualisation, methodology, data curation, resources, investigation, supervision, validation, writing (review and editing).
ORCID IDs
Vhuawelo Simba - https://orcid.org/0000-0003-4713-7584
Inam Yekwayo - https://orcid.org/0000-0002-7671-3643
Tarombera Mwabvu - https://orcid.org/0000-0002-8947-7811
REFERENCES
Abera-Kalibata AM, Gold CS, Van Driesche RG, Ragama P E. 2007. Composition, distribution, and relative abundance of ants in banana farming systems in Uganda. Biological Control 40(2):168-178. https://doi.org/10.1016/j.biocontrol.2006.11.006. [ Links ]
Ahmad A, Ahmad AH. 2009. A preliminary study on the decomposition and dipteran associated with exposed carcasses in an oil palm plantation in Bandar Baharu, Kedah, Malaysia. Tropical Biomedicine. 26(1):1-10. [ Links ]
Albertini A, Pizzolotto R, Petacchi R. 2017. Carabid patterns in olive orchards and woody semi-natural habitats: first implications for conservation biological control against Bactrocera oleae. Biological Control 62:71-83. https://doi.org/10.1007/s10526-016-9780-x. [ Links ]
Andersen AN, Azcárate FM, Cowie ID. 2000. Seed selection by an exceptionally rich community of harvester ants in the Australian seasonal tropics. Journal of Animal Ecology 69(6):975-984. https://doi.org/10.1111/j.1365-2656.2000.00452.x. [ Links ]
Anderson MJ, Willis TJ. 2003. Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84(2):511-525. https://doi.org/10.1890/0012-9658(2003)084[0511:CAOPCA]2.0.CO;2. [ Links ]
Araújo JDL, Pastori PL, Gomes VFF, Mendes PF, Nunes LAPL. 2018. Changes in the abundance and diversity of soil arthropods in the cultivation of fruit crops. Revista Ciência Agronömica 49(4):537-546. https://doi.org/10.5935/1806-6690.20180061. [ Links ]
Barretto JW, Cultid-Medina CA, Escobar F. 2019. Annual abundance and population structure of two dung beetle species in a human-modified landscape. Insects 10(1). https://doi.10.3390/insects10010002. [ Links ]
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MHH, White JSS. 2009. Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology and Evolution 24(3):127-135. https://doi.org/10.1016/j.tree.2008.10.008. [ Links ]
Bolu H, Özgen 1. 2018. Some observations about Formicidae (Hymenoptera) in almond agroecosystems in East-Southeast Anatolian Region of Turkey. Munis Entomology and Zoology. 13(2):495-498. [ Links ]
Bommarco R, Marini L, Vaissière BE. 2012. Insect pollination enhances seed yield, quality, and market value in oilseed rape. Oecologia 169:1025-1032. https://doi.org/10.1007/s00442-012-2271-6. [ Links ]
Borgelt A, New TR. 2006. Pitfall trapping for ants (Hymenoptera, Formicidae) in mesic Australia: what is the best trapping period? Journal of Insect Conservation 10:75-77. https://doi.org/10.1007/s10841-005-7549-0. [ Links ]
Botha M, Siebert SJ, Van den Berg J. 2016. Do arthropod assemblages fit the grassland and savanna biomes of South Africa? South African Journal of Science 112(9/10):1-10. https://doi.org/10.17159/sajs.2016/20150424. [ Links ]
Bouchard P. 2014. The Book of Beetles: A Life-Size Guide to Six Hundred of Nature's Gems. Chicago: University of Chicago Press. [ Links ]
Campbell CW. 2018. Tropical Fruits and Nuts. In CRC Handbook of tropical food crops. Boca Raton: CRC Press. https://doi.org/10.1201/9781351072809. [ Links ]
Cardoso P, Shirey V, Seppälä S, Henriques S, Draney ML, Foord S, Gibbons AT, Gomez LA, Kariko S, Malumbres-Olarte J, Milne M. 2019. Globally distributed occurrences utilised in 200 spider species conservation profiles (Arachnida, Araneae). Biodiversity Data Journal 7:e33264. https://doi.org/10.3897/BDJ.7.e33264. [ Links ]
Clark TE, Samways MJ. 1997. Sampling arthropod diversity for urban ecological landscaping in a species-rich southern hemisphere botanic garden. Journal of Insect Conservation 1:221-234. https://doi.org/10.1023/A:1018472118513. [ Links ]
Crawley MJ. 2005. Statistics: An Introduction Using R. USA: John Wiley,
da Silva PG, Hernandez MIM. 2016. Spatial variation of dung beetle assemblages associated with forest structure in remnants of southern Brazilian Atlantic Forest. Revista Brasileira de Entomologia 60(1):73-81. https://doi.org/10.1016/j.rbe.2015.11.001. [ Links ]
de Castro Solar RR, Barlow J, Andersen AN, Schoereder JH, Berenguer E, Ferreira JN, Gardner TA. 2016. Biodiversity consequences of land-use change and forest disturbance in the Amazon: A multi-scale assessment using ant communities. Biological Conservation 197:98-107. https://doi.org/10.1016/j.biocon.2016.03.005. [ Links ]
Diczbalis Y, Daniells J, Lambert S, Searle C. 2014. Plantation crops. In Horticulture: Plants for People and Places. South Africa: Springer. Dippenaar-Schoeman AS. 2014. Field Guide to the Spiders of South Africa. Cape Town: Lapa.
Dippenaar-Schoeman AS, Jocqué R. 1997. African Spiders: An Identification Manual. Pretoria: ARC-Plant Protection Research Institute. [ Links ]
Dippenaar-Schoeman AS, Van den Berg MA, Van den Berg AM, Van den Berg A. 2001. Spiders in macadamia orchards in the Mpumalanga Lowveld of South Africa: species diversity and abundance (Arachnida: Araneae). African Plant Protection 7(1):39-46. https://hdl.handle.net/10520/EJC87829. [ Links ]
Dippenaar-Schoeman AS, Van den Berg AM, Van den Berg MA, Foord SH. 2005. Spiders in avocado orchards in the Mpumalanga Lowveld of South Africa: species diversity and abundance (Arachnida: Araneae). African Plant Protection 11(1):8-16. https://hdl.handle.net/10520/EJC87789. [ Links ]
Dippenaar-Schoeman AS, Van den Berg AM, Haddad CR, Lyle R. 2013. Current knowledge of spiders in South African agroecosystems (Arachnida: Araneae). Transactions of the Royal Society of South Africa 68(1):57-74. http://dx.doi.org/10.1080/0035919X.2012755136. [ Links ]
Donoso DA, Johnston MK, Kaspari M. 2010. Trees as templates for tropical litter arthropod diversity. Oecologia 164(1):201-211. https://doi.org/10.1007/s00442-010-1607-3. [ Links ]
Dormann CF, Mcpherson JM, Araújo MB, Bivand R, Bolliger J, Carl G, Davies RG, Hirzel A, Jetz W, Kissling WD, Kühn I, Ohlemüller R, Peres-Neto PD, Reineking B, Schröder B, Schurr FM, Wilson R. 2007. Methods to account for spatial autocorrelation in the analysis of species distributional data: a review. Ecography 30(5):609-628. https://doi.org/10.1111/j.2007.0906-7590.05171.x. [ Links ]
Doube BM. 2018. Ecosystem services provided by dung beetles in Australia. Basic Applied Ecology. 26:35-49. https://doi.org/10.1016/j.baae.2017.09.008. [ Links ]
Downie IS, Butterfield JEL, Coulson JC. 1995. Habitat preferences of sub-montane spiders in Northen England. Ecography 18(1):51-61. https://www.jstor.org/stable/3683219. [ Links ]
Driver A, Maze K, Rouget M, Lombard AT, Nel J, Turpie JK, Cowling RM, Desmet P, Goodman P, Harris J, Jonas Z, Reyers B, Sink K, Strauss T. 2005. National spatial biodiversity assessment 2004: priorities for biodiversity conservation in South Africa. Pretoria: South African National Biodiversity Institute. [ Links ]
Druce D, Hamer M, Slotow R. 2007. Patterns of millipede (Diplopoda), centipede (Chilopoda) and scorpion (Scorpionida) diversity in savanna habitats within the Greater Makalali Conservancy, South Africa. African Zoology 42(2):204-215. https://doi.org/10.1080/15627020.2007.11407398. [ Links ]
Eckert M, Gaigher R, Pryke JS, Samways MJ. 2022. Conservation of complementary habitat types and small-scale spatial heterogeneity enhance soil arthropod diversity. Journal of Environmental Management 317:115482. https://doi.org/10.1016/j.jenvman.2022.115482. [ Links ]
Eckert M, Gaigher R, Pryke JS, Samways MJ. 2019. Rapid recovery of soil arthropod assemblages after exotic plantation tree removal from hydromorphic soils in a grassland-timber production mosaic. Restoration Ecology 27(6):1357-1368. https://doi.org/10.1111/rec.12991. [ Links ]
Emekci M, Moore D. 2015. Sap Beetles in Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges. Cham: Springer.
Facure KG, Giaretta AA. 2009. Semi-terrestrial tadpoles as vertebrate prey of trap-jaw ants (Odontomachus, Formicidae). Herpetology Notes 2(1):63-66. [ Links ]
Fisher BL, Bolton B. 2016. Ants of Africa and Madagascar: A Guide to Genera. California: University of California Press. [ Links ]
Foord SF, Dippenaar-Schoeman AS, Haddad CR, Lotz LN, Lyle R. 2011. The faunistic diversity of spiders (Arachnida: Araneae) of the savanna biome in South Africa. Transactions of the Royal Society of South Africa 66(3):170-201. https://doi.org/10.1080/0035919X.2011.639406. [ Links ]
Foord SH, Dippenaar-Schoeman AS, Haddad CR, Lyle R, Lotz LN, Sethusa T, Raimondo D. 2020. The South African National Red List of spiders: patterns, threats, and conservation. The Journal of Arachnology 48(2):110-118. https://doi.org/10.1636/0161-8202-48.2.110. [ Links ]
Gallé R, Geppert C, Földesi R, Tscharntke T, Batáry P. 2020. Arthropod functional traits shaped by landscape-scale field size, local agri-environment schemes and edge effects. Basic and Applied Ecology 48:102-111. https://doi.org/10.1016Ai.baae.2020.09.006. [ Links ]
García-Martínez MÁ, Martínez-Tlapa DL, Pérez-Toledo GR, Quiroz-Robledo LN, Castafto-Meneses G, Laborde J, Valenzuela-González JE. 2015. Taxonomic, species and functional group diversity of ants in a tropical anthropogenic landscape. Tropical Conservation Science 8:1017-1032. https://doi.org/10.1177/194008291500800412. [ Links ]
Geldenhuys M, Gaigher R, Pryke JM, Samways MJ.2021. Diverse herbaceous cover crops promote vineyard arthropod diversity across different management regimes. Agriculture, Ecosystems and Environment 307:107222. https://doi.org/10.1016/j.agee.2020.107222. [ Links ]
Granatstein D, Kirby E, Ostenson H, Willer H. 2016. Global situation for organic tree fruits. Scientia Horticulturae 208:3-12. https://doi.org/10.1016/j.scienta.2015.12.008. [ Links ]
Grout TG, Begemann GJ, Stephen PR. 2001. Monitoring and control of coffee bean weevil, Araecerus coffeae (Coleoptera: Anthribidae), in southern African citrus orchards. African Plant Protection 7(2): 67-72. https://hdl.handle.net/10520/EJC87841. [ Links ]
Haddad CR. 2013. Taxonomic notes on the spider genus Messapus Simon, 1898 (Araneae, Corinnidae), with the description of the new genera Copuetta and Wasaka and the first cladistic analysis of Afrotropical Castianeirinae. Zootaxa 3688(1):1-79. https://doi.org/10.11646/zootaxa.3688.1.1. [ Links ]
Hafsi A, Abbes K, Harbi A, Duyck PF, Chermiti B. 2016. Attract-and-kill systems efficiency against Ceratitis capitata (Diptera: Tephritidae) and effects on non-target insects in peach orchards. Journal of Applied Entomology 140(1-2):28-36. https://doi.org/10.1111/jen.12259. [ Links ]
Halaj J, Halpern CB, Yi H. 2008. Responses of litter-dwelling spiders and carabid beetles to varying levels and patterns of green-tree retention. Forest Ecology and Management 255(3-4):887-900. https://doi.org/10.1016/j.foreco.2007.09.083. [ Links ]
Herrera HW, Baert L, Dekoninck W, Causton CE, Sevilla CR, Pozo P, Hendrickx F. 2020. Distribution and habitat preferences of Galápagos ants (Hymenoptera: Formicidae). Belgian Journal of Entomology 93:1-60. http://dspace.espoch.edu.ec/handle/123456789/14062. [ Links ]
Herwina H, Nasir N, Jumjunidang, Yaherwandi. 2013. The composition of ant species on banana plants with banana bunchy-top virus (BBTV) symptoms in West Sumatra, Indonesia. Asian Myrmecology 5:151-161. [ Links ]
Isbell F, Adler PR, Eisenhauer N, Fornara D, Kimmel K, Kreme C, Letourneau DK, Liebman M, Polley HW, Quijas S, Scherer-Lorenzen M. 2017. Benefits of increasing plant diversity in sustainable agroecosystems. Journal of Ecology 105(4):871-879. https://doi.org/10.1111/1365-2745.12789. [ Links ]
Joma AM, Mackay WP. 2017. Revision of the African ants of the Bothroponera sulcata species complex (Hymenoptera: Formicidae: Ponerinae). Transactions of the American Entomological Society 143(1): 7-71. https://doi.org/10.3157/061.143.0103. [ Links ]
Kelly JA, Samways MJ. 2011. Mound-building termite (Blattaria: Isoptera) mound density and ecological correlates in a southern African savanna reserve. African Entomology 19(1):156-164. https://doi.org/10.10520/EJC32878. [ Links ]
Khun KK, Ash GJ, Stevens MM, Huwer RK, Wilson BA. 2020. Response of the macadamia seed weevil Kuschelorhynchus macadamiae (Coleoptera: Curculionidae) to Metarhizium anisopliae and Beauveria bassiana in laboratory bioassays. Journal of Invertebrate Pathology 174:107437. https://doi.org/10.1016/j.jip.2020.107437. [ Links ]
Kwon TS, Park YK, Lim JH, Ryou SH, Lee CM. 2013. Change of arthropod abundance in burned forests: different patterns according to functional guilds. Journal of Asia Pacific Entomology 16(3):321-328. [ Links ]
LaScaleia MC, Reynolds C, Magagula CN, Roets F, McCleery RA. 2018. Dung beetle richness decreases with increasing landscape structural heterogeneity in an African savanna-agricultural mosaic. Insect Conservation and Diversity 11(4):396-406. https://doi.org/10.1111/icad.12290. [ Links ]
Lichtenberg EM, Kennedy CM, Kremen C, Batary P, Berendse F, Bommarco R, Bosque-Pérez NA, Carvalheiro LG, Snyder WE, Williams NM, Winfree R. 2017. A global synthesis of the effects of diversified farming systems on arthropod diversity within fields and across agricultural landscapes. Global Change Biology 23(11):1-12. https://doi.org/10.1111/gcb.13714. [ Links ]
Loyola RD, Brito SL, Ferreira RL. 2006. Ecosystem Disturbances and Diversity Increase: Implications for Invertebrate Conservation. In Arthropod Diversity and Conservation: Topics in Biodiversity and Conservation. Dordrecht: Springer.
Luke SH, Fayle TM, Eggleton P, Turner EC, Davies RG. 2014. Functional structure of ant and termite assemblages in old growth forest, logged forest and oil palm plantation in Malaysian Borneo. Biodiversity and Conservation 23(11):2817-2832. https://doi.prg/10.1007/s10531-014-0750-2. [ Links ]
Magagula CN, Samways MJ. 2001. Maintenance of lady beetle diversity across a heterogeneous African agricultural/savanna land mosaic. Biodiversity and Conservation 10:209-222. https://doi.org/10.1023/A:1008900302267. [ Links ]
Magura T, Tóthmérész B, Elek Z. 2005. Impacts of leaf-litter addition on carabids in a conifer plantation. Biodiversity and Conservation 14:475-491. https://doi.org/10.1007/s10531-004-7307-8. [ Links ]
Marasas ME, Sarandón SJ, Cicchino A. 2010. Semi-natural habitats and field margins in a typical agroecosystem of the Argentinean Pampas as a reservoir of carabid beetles. Journal of Sustainable Agriculture 34(2):153-168. https://doi.org/10.1080/10440040903482563. [ Links ]
Marchant R. 2010. Understanding complexity in savannas: climate, biodiversity and people. Current Opinion in Environmental Sustainability 2(1-2):101-108. https://doi.org/10.1016/j.cosust.2010.03.001. [ Links ]
Marie A, Mangenot A, Puech C, Aviron S, Plantegenest M, Pétillon J. 2018. Farming system and landscape characteristics differentially affect two dominant taxa of predatory arthropods. Agriculture, Ecosystems and Environment 259:98-110. https://doi.org/10.1016/j.agee.2018.02.031. [ Links ]
Matlock Jr RB, de la Cruz R. 2003. Ants as indicators of pesticide impacts in banana. Environmental Entomology 32(4):816-829. https://doi.org/10.1603/0046-225X-32.4.816. [ Links ]
Matta DHD, Cividanes FJ, Silva RJ, Batista MN, Otuka AK, Correia ET, Matos STS. 2017. Feeding habits of Carabidae (Coleoptera) associated with herbaceous plants and the phenology of coloured cotton. Acta Scientiarum Agronomy 39(2):135-142. https://doi.org/10.4025/actasciagron.v39i2.32593. [ Links ]
Mauda EV, Joseph GS, Seymour CL, Munyai TC, Foord SH. 2018. Changes in land use alter ant diversity, assemblage composition and dominant functional groups in African savannas. Biodiversity and Conservation 27:947-965.https://doi.org/10.1007/s10531-017-1474-x. [ Links ]
Michalko R, Pekár S, Dul'a M, Entling MH. 2019. Global patterns in the biocontrol efficacy of spiders: A meta-analysis. Global Ecology and Biogeography 28(9):1366-1378. https://doi.org/10.1111/heb.12927. [ Links ]
Moir ML, Brennan KEC, Koch JM, Majer JD, Fletcher MJ. 2005. Restoration of a forest ecosystem: the effects of vegetation and dispersal capabilities on the reassembly of plant-dwelling arthropods. Forest Ecology and Management 217(2-3):294-306. https://doi.org/10.1016/j.foreco.2005.06.012. [ Links ]
Mollot G, Tixier P, Lescourret F, Quilici S, Duyck PF. 2012. New primary resource increases predation on a pest in a banana agroecosystem. Agricultural and Forest Entomology 14(2):317-323. https://doi.org/10.1111/j.1461-9563.2012.00571.x/abstract. [ Links ]
Moses J, Fayle TM, Novotny V, Klimes P. 2021. Elevation and leaf litter interact in determining the structure of ant communities on a tropical mountain. Biotropica 53(3):906-919. https://doi.org/10.1111/btp.12914. [ Links ]
Mwabvu T, Yekwayo I. 2020. A checklist of ants (Formicidae), spiders (Araneae) and millipedes (Spirostreptida) of the savannah in Mpumalanga Province, South Africa. African Journal of Ecology 58(1):138-144. https://doi.org/10.1111/aje.12679. [ Links ]
Olesen T, Robertson D, Muldoon S, Meyer R. 2008. The role of carbohydrate reserves in evergreen tree development, with particular reference to macadamia. Scientia Horticulturae 117:7377. https://doi.org/10.1016/J.SCIENTA.2008.03.026. [ Links ]
Olfert O, Johnson GD, Brandt SA, Thomas AG. 2002. Use of arthropod diversity and abundance to evaluate cropping systems. Agronomy Journal 94(2):210-216. https://doi.org/10.2134/agronj2002.2100. [ Links ]
Passos L, Oliveira PS. 2002. Ants affect the distribution and performance of seedlings of Clusia criuva, a primarily bird-dispersed rain forest tree. Journal of Ecology 90(3):517-528. https://doi.org/10.1046/j.1365-2745.2002.00687.x. [ Links ]
Passos L, Oliveira PS. 2004. Interaction between ants and fruits of Guapira opposita (Nyctaginaceae) in a Brazilian sandy plain rainforest: ant effects on seeds and seedlings. Oecologia 139:376382. https://doi.org/10.1007/s00442-004-1531-5. [ Links ]
Pekár S. 2004. Predatory behaviour of two European ant-eating spiders (Araneae, Zodariidae). Journal of Arachnology 32(1):31-41. https://doi.org/10.1636/S02-15. [ Links ]
Pekár S, Toft S. 2015. Trophic specialisation in a predatory group: the case of prey-specialised spiders (Araneae). Biological reviews of the Cambridge Philosophical Society. 90(3):744-761. https://doi.org/10.1111/brv.12133. [ Links ]
Perfecto I. 1990. Indirect and direct effects in a tropical agroecosystem: the maize-pest-ant system in Nicaragua. Ecology 71(6):2125-2134. https://doi.org/10.2307/1938626 [ Links ]
Perfecto I, Snelling R. 1995. Biodiversity and the transformation of a tropical agroecosystem: ants in coffee plantations. Ecological Applications 5(4):1084-1097. https://doi.org/10.2307/2269356. [ Links ]
Picker M, Griffiths C, Weaving A. 2019. Field Guide to Insects of South Africa. 2nd Edition. Cape Town: Penguin Random House South Africa. [ Links ]
Pierre EM, Idris A. 2013. Studies on the predatory activities of Oecophylla smaragdina (Hymenoptera: Formicidae) on Pteroma pendula (Lepidoptera: Psychidae) in oil palm plantations in Teluk Intan, Perak (Malaysia). Asian Myrmecology 5(1):163-176. [ Links ]
Pio R, de Souza FBM, Kalcsits L, Bisi RB, Farias DH. 2018. Advances in the production of temperate fruits in the tropics. Acta Scientiarum. Agronomy 41:1807-8621. https://doi.org/10.4025/actasciagron.v41i1.39549. [ Links ]
Poeydebat C, Tixier P, De Bellaire LDL, Carval D. 2017. Plant richness enhances banana weevil regulation in a tropical agroecosystem by affecting a multitrophic food web. Biological Control 114:125-132. https://doi.org/10.1016/j.biocontrol.2017.08.009. [ Links ]
Pompeo PN, Filho LCIO, Filho OK, Mafra ÁL, Baretta CRDM, Baretta D. 2016. Coleoptera diversity (Arthropoda: Insecta) and soil properties under soil management systems in the highlands of Santa Catarina state, Brazil. Scientia Agraria 17(1):16-28. [ Links ]
Pryke JS, Roets F, Samways MJ. 2022. Large African herbivore diversity is essential in transformed landscapes for conserving dung beetle diversity. Journal of Applied Ecology 59(5):1372-1382. https://doi.org/10.1111/1365-2664.14152. [ Links ]
Pryke JS, Roets F, Samways MJ. 2013. Importance of habitat heterogeneity in remnant patches for conserving dung beetles. Biodiversity and Conservation 22:2857-2873. https://doi.org/10.1007/s10531-013-0559-4. [ Links ]
R Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Raimundo RL, Freitas AV, Oliveira PS. 2009. Seasonal patterns in activity rhythm and foraging ecology in the neotropical forest-dwelling ant, Odontomachus chelifer (Formicidae: Ponerinae). Annals of the Entomological Society of America 102(6):1151-1157. https://doi.org/10.1603/008.102.0625. [ Links ]
Rhoades PR, Davis TS, Tinkham WT, Hoffman CM. 2018. Effects of seasonality, forest structure, and understory plant richness on bee community assemblage in a southern Rocky Mountain mixed conifer forest. Annals of the Entomological Society of America 111(5):278-284. https://doi.org/10.1093/aesa/say021. [ Links ]
Riggio J, Jacobson A, Dollar L, Bauer H, Becker M, Dickman A, Funston P, Groom R, Henschel P, de Iongh H, Lichtenfeld L. 2013. The size of savannah Africa: a lion's (Panthera leo) view. Biodiversity and Conservation 22, 17-35. https://doi.org/10.1007/s10531-012-0381-4. [ Links ]
Ripley B, Venables B, Bates DM, Hornik K, Gebhardt A, Firth D, Ripley MB. 2013. Package 'MASS'. Cran R. 538:113-120. [ Links ]
Roth DS, Perfecto I, Rathcke B. 1994. The effects of management systems on ground-foraging ant diversity in Costa Rica. Ecological Applications. 4(3):423-436._https://doi.org/10.2307/1941947. [ Links ].
Rutherford MC, Mucina L, Powrie LW. 2006. Biomes and Bioregions of Southern Africa. In The vegetation of South Africa, Lesotho and Swaziland. South Africa: South African National Biodiversity Institute.
Samu F, Szinetár C. 2002. On the nature of agrobiont spiders. The Journal of Arachnology 30(2):389-402. https://doi.org/10.1636/0161-8202(2002)030[0389:OTNOAS]2.0.CO;2. [ Links ]
Samways MJ, Hamer M, Veldtman R. 2012. Development and Future of Insect Conservation in South Africa. In Insect Conservation: Past, Present a nd Prospects. Dordrecht: Springer.
Samways MJ, McGeoch MA, New TR. 2010. Insect Conservation: A Handbook of Approaches and Methods. New York: Oxford University Press. [ Links ]
Saqib HSA, Chen J, Chen W, Pozsgai G, Akutse KS, Ashraf MF, You M, Gurr GM. 2020. Local management and landscape structure determine the assemblage patterns of spiders in vegetable fields. Scientific Reports 10:1-11. https://doi.org/10.1038/s41598-020-71888-w. [ Links ]
Sarabi V. 2019. Factors that influence the level of weed seed predation: a review. Weed Biology and Management 19(3):61-74. https://doi.org/10.1111/wbm.12186. [ Links ]
Sayer EJ, Tanner EV, Lacey AL. 2006. Effects of litter manipulation on early-stage decomposition and meso-arthropod abundance in a tropical moist forest. Forest Ecology and Management 229(1-3): 285-293. https://doi.org/10.1016/j.foreco.2006.04.007. [ Links ]
Simba LD, Foord SH, Thébault E, van Veen FF, Joseph GS, Seymour CL. 2018. Indirect interactions between crops and natural vegetation through flower visitors: the importance of temporal as well as spatial spillover. Agriculture Ecosystem and Environment 253, 148-156. https://doi.org/10.1016/j.agee.2017.11.002. [ Links ]
Sitters J, Maechler MJ, Edwards PJ, Suter W, Olde Venterink H. 2014. Interactions between C: N: P stoichiometry and soil macrofauna control dung decomposition of savanna herbivores. Functional Ecology 28(3)776-786. https://doi.org/10.1111/1365-2435.12213. [ Links ]
Sutton TB, Aldwinckle HS, Agnello AM, Walgenbach JF. 2014. Compendium of Apple and Pear Diseases and Pests. USA: American Phytopathological Society.
Telnov D, Ghahari H. 2018. An annotated checklist of the Anthicidae and pediline Pyrochroidae (Insecta: Coleoptera) of Iran, with thirteen new country records. Zootaxa 4497(4):451-491. https://doi.org/10.11646/zootaxa.4497.4.1. [ Links ]
Tiwari AK, Singh G, Singh R. 2021. Faunal diversity of Oonopidae (Araneomorphae: Araneae: Arachnida) in India. Journal of Global Biosciences 10(1):8340-8351. https://ssrn.com/abstract=3916149. [ Links ]
Tsai YY, Pekár S. 2019. Prey acceptance and conditional foraging behavior in the cribellate-web spider Titanoeca quadriguttata (Araneae: Titanoecidae). The Journal of Arachnology 47(2):202-208. https://doi.org/10.1636/JoA-S-18-083. [ Links ]
Ubick D, Paquin P, Cushing PE, Roth VD. 2005. Spiders of North America: An identification manual. New Hampshire: American Arachnological Society. [ Links ]
van Schalkwyk J, Pryke JS, Samways MJ. 2017. Wide corridors with much environmental heterogeneity best conserve high dung beetle and ant diversity. Biodiversity and Conservation 26:1243-1256. https://doi.org/10.1007/s10531-017-1299-7. [ Links ]
Wang YI, Naumann U, Wright ST, Warton DI. 2012. mvabund-an R package for model-based analysis of multivariate abundance data. Methods in Ecology and Evolution 3(3):471-474. https://doi.org/10.1111/j.2041-210X.2012.00190.x. [ Links ]
White RE. 1998. The Beetles of North America. Boston: Houghton Mifflin Harcourt. [ Links ]
Woodcock BA. 2005. Pitfall Trapping in Ecological Studies. UK: Blackwell Publishing. [ Links ]
Wyckhuys KA, O'Neil RJ. 2006. Population dynamics of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) and associated arthropod natural enemies in Honduran subsistence maize. Crop Protection 25(11):1180-1190. https://doi.org/10.1016/jxropro.2006.03.003. [ Links ]
Wyss E. 1996. The effects of artificial weed strips on diversity and abundance of the arthropod fauna in a Swiss experimental apple orchard. Agriculture, Ecosystems and Environment 60(1):47-59. https://doi.org/10.1016/S0167-8809(96)01060-2. [ Links ]
Yekwayo I, Pryke JS, Roets F, Samways MJ. 2017. Responses of ground living arthropods to landscape contrast and context in a forest-grassland mosaic. Biodiversity and Conservation 26:631-651. https://doi.org/10.1007/s10531-016-1262-z. [ Links ]
 Correspondence:
Correspondence:
Inam Yekwayo
Email: iyekwayo@wsu.ac.za
Received: 06 June 2022
Accepted:20 October 2022


{kind=link}