










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAfrican Entomology
versão On-line ISSN 2224-8854
versão impressa ISSN 1021-3589
AE vol.31 Pretoria 2023
http://dx.doi.org/10.17159/2254-8854/2022/a13584
RESEARCH ARTICLE
Mosquito community composition in Central District, Botswana: insights from a malaria endemic to non-endemic gradient
M BuxtonI,II; C NyamukondiwaII,III; M KesamangII; RJ WassermanIII, II
IDepartment of Biological Sciences, Physical and Chemical Sciences, Botswana University of Agriculture and Natural Resources, Gaborone, Botswana
IIDepartment of Biological Sciences and Biotechnology, Botswana International University of Science and Technology, Palapye, Botswana
IIIDepartment of Zoology and Entomology, Rhodes University, Makhanda, South Africa
ABSTRACT
Spatial distribution of vector mosquitoes plays a critical role in the dynamics of associated diseases' spread across diverse landscapes. In Botswana, six Districts are implicated as malaria endemic zones, one of which is the Central District comprising both malaria endemic and non-endemic sub-districts. Despite being the biggest in the country, mosquito diversity in this District is under-explored, more so in the malaria non-endemic sub-districts. Here, we thus sampled mosquito adults and larvae from the malaria endemic sub-district of Bobirwa and non-endemic sub-districts of Palapye and Serowe, to determine spatial mosquito abundance and diversity in the District. Overall, all the sub-districts had a representation of key mosquito taxa of medical and economic importance (Aedes, Culex and Anopheles), irrespective of malarial endemicity status. Bobirwa had the highest number of mosquitoes sampled (429) although the greatest species richness (0.8511) was observed from Palapye. Moreover, Palapye also recorded a species from another genus, Culiseta longiareolata, a known natural vector of avian malaria parasites. Given global climate shift projections for the region, there is a need for continuous area-wide surveillance for vector mosquitoes and associated parasites in curbing the risk of emerging and re-emerging infections. While the Anopheles-centric approach to mosquito control is still necessary, a holistic approach, incorporating other vector incriminated mosquito species is warranted, particularly given shifting climates and the presence of invasive disease associated vector mosquito species.
Keywords: Bobirwa; endemicity status; mosquito-borne diseases; Palapye; Serowe; species composition; vectors
INTRODUCTION
Through the transmission of parasites such as protozoans, helminths and viruses, mosquitoes facilitate incapacitating diseases such as malaria, Zika, dengue, Wet Nile fever, chikungunya (Lee et al. 2018; Ralapanawa & Kularatne 2020). Globally, the introduction of invasive vector mosquitoes in new environments are on the rise (Akiner et al. 2016; Wilke et al. 2020) and so are numbers of other vector incriminated mosquito species and associated parasites (Braack et al. 2018). This has led to a concomitant surge in cases of vector-borne diseases (WHO 2014), with disease vector mosquitoes contributing increasingly to human morbidity and mortality worldwide (Ahmed et al. 2020; Boyer et al. 2020). Similarly, although less well documented, vector mosquitoes also contribute significantly to livestock and wildlife disease transmission (Mroz et al. 2017). The distribution of mosquitoes has been well studied in tropical and temperate regions worldwide (Atoni et al. 2019; Foster & Walker 2019; Martinet et al. 2019) with biotic and abiotic factors largely shaping mosquito community composition (Biteye et al. 2018; Akpan et al. 2019; Cane et al. 2020; Mouatassem et al. 2020). There is, however, continued scope for further exploration in this regard, given that changing environmental conditions and biological invasions are resulting in shifts in distribution of known vectors, and the presence of new vector mosquito species and associated pathogens (Weyl et al. 2019; Pereira dos Santos et al. 2018).
Climatic and human-mediated factors are regarded as crucial drivers of mosquito population dynamics (Monaghan et al. 2018; Lee et al. 2020; Wang et al. 2020). Given the recent increase in shifting climates and anthropogenic activities facilitating vector mosquito species (Schrama et al. 2020; Wang et al. 2020), it is highly likely that certain mosquitoes (Liu-Helmersson et al. 2019) and indeed their associated pathogens (Braack et al. 2018) may proliferate and translocate to novel environments. Many mosquitoes have also been reported as being invasive (Vaux et al. 2019; Fălcuţă et al. 2020) with potential of dispersing across different landscapes (Kampen et al. 2017; Strickman 2020). This necessitates the need for continual and revised spatio-temporal monitoring of known and potential vector species (Ludwig et al. 2019; El-Sayed & Kamel 2020). The geographic distribution and composition of mosquito communities is well documented in Africa (Jupp & Mcintosh 1990; Njabo et al. 2009; Alves et al. 2010; Johnson et al. 2020) albeit biased towards malaria-burdened areas (Coetzee et al. 2000; Wiebe et al. 2017). As such, anopheline mosquitoes are the focus of much of the biological, autecological and even community ecology studies in the region, particularly in southern Africa (Ferguson et al. 2010; Zengenene et al. 2020). As a result, little work has focused on culicine mosquito species in southern Africa (but see Pachka et al. 2016; Matowo et al. 2019; Kahamba et al. 2020).
Botswana mosquito research is largely reflective of that of the region, with mosquito work mainly explored within parts of the country that experience human malaria cases (Chirebvu & Chimbari 2015; Pachka et al. 2016; Tawe et al. 2017; Cornel et al. 2018; Kgoroebutswe et al. 2020). Malaria is currently the most apparent mosquito-borne infection in the country resulting in anopheline species research prioritisation, particularly within districts referred to as "malaria endemic", where historical malaria cases have been high. In contrast, the "non-endemic" malaria areas are largely neglected (Bango et al. 2020). Furthermore, the vector control monitoring programmes are also focused on anopheline species within the endemic localities. Areas that were previously known to be malaria free (e.g. Palapye, Serowe, Ghanzi, Kweneng) have, however, recently recorded both indigenous and imported malaria cases (Chihanga et al. 2016; Bango et al. 2020). For example, in recent years, malaria cases and deaths have been reported persistently across sub-districts in the Central District, with Bobirwa having the highest cases, Serowe exhibiting the least while Palapye region remained intermediate (Table S.1). But in addition to malaria, there are other mosquito-borne diseases in the region, vectored by non-anopheline mosquitoes, but are overlooked by the vector control monitoring programmes. For example, Culex spp. are known vectors of diseases for livestock and wild animals, although very little work has been conducted on this in the region. Similarly, globally invasive vector species have been detected in surrounding countries (Masaninga et al. 2014; Noden et al. 2014; Weyl et al. 2019), and very recently from a single sub-district in Central District of Botswana (Buxton et al. 2019), although no further investigation of their prevalence in Botswana has been explored. As such, nation-wide vector mosquito monitoring data remains scant. This has hindered understanding of mosquito species distribution and predictive modelling of vector-borne disease outbreaks. This also has critical implication on modelling vector borne disease risk in areas or for taxa regarded of 'less concern'. An understanding of available mosquito species would facilitate development of policy for a move from the Anopheles-centric approach to a more inclusive vector management strategy, including screening for associated human, domestic and wildlife associated pathogens and parasites.
Given its primacy and centrality, and attracting national developmental attention, Central District is an area requiring research prioritisation in Botswana. Owing to the confluence of so many trade and touristic routes, the risk of introduction of new vectors and or associated pathogens/parasites in this district may be high. The consequential effects of these introductions to trade and tourism could also be highly significant. The aim of the study was to conduct an assessment of the mosquito species composition in human inhabited areas in the district. More specifically, species composition was contrasted across a malaria endemic to non-endemic sub-district gradient. We hypothesised that (i) abundance of Culex spp. would be more than other mosquito species due to the influence of favourable anthropogenic-degraded habitats in highly urbanised sub-districts (ii) anopheline species would dominate in the malaria endemic Bobirwa where malaria and associated vectors and pathogens are indigenous and historically prevalent, and that (iii) invasive Aedes spp. would be present in the Palapye and Serowe sub-districts given their increased urban size and proximity to South Africa where this species has been reported (Weyl et al. 2019), and their previous presence being recorded in Palapye (Buxton et al. 2019). The results of this study will serve as important findings for future work for these understudied regions (Tawe et al. 2017; Bango et al. 2020; Kgoroebutswe et al. 2020) and enlighten communities, researchers and practitioners of potential risk vectors, and associated disease transmission. The findings will also have implications in the monitoring of vector proliferation due to anthropogenic and climate driven factors prevailing in the region that favour mosquito success and associated infections (e.g. Endo & Eltahir 2020).
MATERIALS AND METHODS
Mosquito community collection
Mosquito samples were collected from understudied sub-districts in Central District, Botswana. This District straddles a malaria-endemicity gradient, consisting of three malaria-endemic and three non-endemic zones (Figure 1). Samples were collected from both the malaria endemic (Bobirwa) and the non-endemic sub-districts (Palapye and Serowe) between November and January austral summer of 2019/2020 (Figure 1; Table S.2). Sampling was conducted during this period as it falls within the rainy season for the area, the time of year when mosquito abundances are highest and where communities identified risk of bites to be the greatest (Buxton et al. 2020). Nine collection points were identified per sub-district based on proximity to human habitation (six for adults) and the presence of breeding sites (three for larvae). Adult and larval mosquito species were sampled from a total of nine collection points per sub-district, monitored repeatedly over 3 days (adults). The adults were trapped using a Biogents BG-GAT (Model: 10719, Biogents AG, Germany) baited with decaying hay. Samples were stored individually in 2 ml Eppendorf tubes with desiccating silica gel beads. Mosquito larvae were sampled with a 1000 μm mesh net (40 cm × 60 cm) from three separate aquatic habitats within each sub-district with standardised search times of 10 min per sampling site (Lee et al. 2020). Each larval specimen was preserved in 2 ml Eppendorf tubes using 80% alcohol.
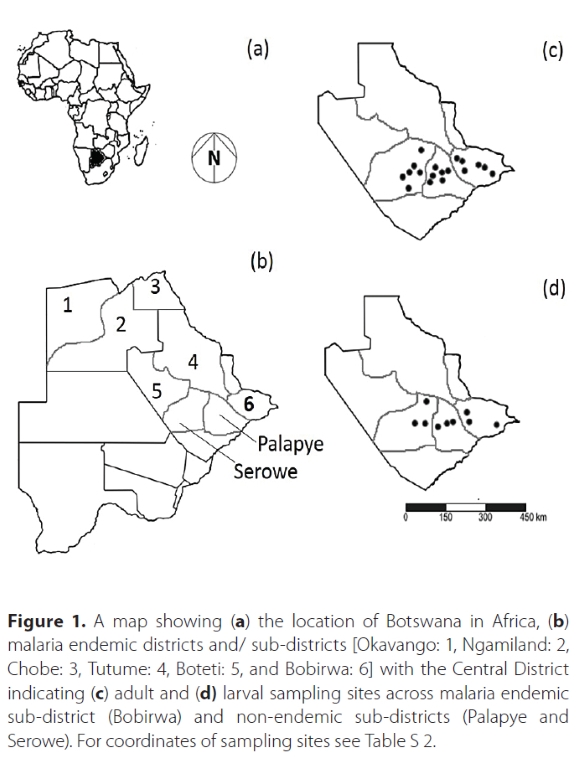
Identification
All adult and larval mosquito specimens were first identified using morphological features and adults were further sexed male and female based on the antenna (Folmer et al. 1994; Gillies & Coetzee 1987; Jupp 1996; Rueda et al. 2004). Modified protocols of Buxton et al. (2019) were subsequently used to confirm species identity with molecular techniques. Five individuals of both adults and larvae were randomly selected across species and subjected to polymerase chain reaction (PCR) for the amplification of the cytochrome oxidase subunit I (COI) using the universal primers LCO 1490 and HCO 2198. For adults, a leg was used in the amplification whilst for larval specimens, a portion of the abdomen was utilised (Bass et al. 2007). Dissected mosquito parts were added directly to the 50 μl PCR reaction mix consisting of 25 μl of 2× Phusion U Multiplex PCR Mix (Thermo Fisher Scientific, U.S.A.), 9 μl nuclease-free water (VWR International LLC, U.S.A.) and 0.8 μM of each primer. The PCR conditions were: initial denaturation at 98 °C for 30 s, 35 cycles of denaturation at 98 °C for 10 s, annealing at 48 °C for 30 s and extension at 72 °C for 30 s. Subsequently, a final extension at 72 °C for 7 min was performed. Visualisation of amplicons was done under 1% agarose gel stained with ethidium bromide. Purification of 40 μl PCR product was performed using a JET PCR purification kit (Thermo Fisher Scientific, U.S.A.) according to manufacturer's recommended protocols, followed by elution with 20 μl nuclease free water (VWR International LLC, U.S.A.). Amplicons were sequenced in Inqaba Biotechnical Industries (Pretoria, South Africa). The sequences were trimmed and contigs were assembled using a CodonCode Aligner 8.0.2 software package and subjected to nucleotide blast for species homology identification. Sequences were further deposited in Genbank to obtain species accession numbers.
Statistical analyses
Data on mosquito community composition were analysed using the Primer version 6 statistical software package, within which the total species number (S), total number of individuals (N), Margalef's species richness (d), species diversity (H) and Pielou's evenness (J') were determined per site. The same software package was then used to assess similarities in community composition. Although mixed methods were employed, given that the effort was standardised across all three sub-districts (6 and 3 sites each for adult and larval collection, respectively), a combined species list along with abundances per species was created per sub-district. With this matrix, a Bray-Curtis similarity matrix was used to construct a dendrogram to assess similarities across sub-districts. The Simper analysis in Primer version 6 was then used to assess the levels of dissimilarity between the sites (sub-districts) and the species contributing the greatest towards these differences.
Mosquito abundances (adults) data were analysed in STATISTICA, version 13.5.0.17 (Statsoft Inc., Tulsa, OK, U.S.A.). The data were checked for normality using Shapiro-Wilks test and fulfilled assumptions of analysis of variance (ANOVA). Factorial ANOVA was then used to analyse data, with abundances as dependent variables while the species, sex, sub-district were the independent factors.
RESULTS
A total of 1013 mosquito samples were collected across the three sub-districts sampled (754 adults and 259 larvae). Of these, 10 species from 4 genera (Aedes, Anopheles, Culex, Culiseta) were identified in either adult, larval stage, and/or both (Table 1). Aedine species identified similar to the partial COI gene isolate were Ae. vittatus and Ae. aegypti with the nucleotide sequence homology of 99.4 and 99.64% respectively. Aedes aegypti was identified and reported in all sub-districts however, Ae. vittatus was only found in Bobirwa. The anopheline species were identified as An. rhodesiensis (93%), An. quadriannulatus (98.51%), An. gambiae sensu stricto (s.s.) (98.95%), An. rufipes (95.53%) and an unidentified specimen with homology of 100% with an Anopheles sp. Bobirwa had one anopheline species (An. quadriannulatus) as compared to more than one species reported for the non-endemic sub-districts of Serowe (An. quadriannulatus and An. rufipes) and Palapye (An. rhodesiensis, An. gambiae s.s. and Anopheles sp.). Thus, the highest number of anopheline spp. Palapye were documented from Palapye from sampled sub-districts. Culex mosquito species identified comprised Cx. pipiens and Cx. sinaiticus with 99.85 and 99.23% homology respectively while Cs. longiareolata were identified at homology of 96.59% (Table 1). Mosquito species identified as adults were Ae. aegypti, Ae vittatus, Cx. sinaiticus and Cx pipiens. For the larval samples, Ae. aegypti, Ae. vittatus, An. rhodesiensis, An. quadriannulatus, An. gambiae s.s., An. rufipes, Anopheles sp., Cx. sinaiticus, Cx. pipiens and Cs. longiareolata were identified. Overall, larval sampling yielded more species and genera (Aedes, Anopheles, Culex and Culiseta) compared to adult (Aedes and Culex) collection. Subsequently, the sequences of all species identified here, were deposited in the Genbank and accession numbers confirmed for each species (Table 1).
Out of the 1013 total mosquito samples, Bobirwa had the highest number of mosquitoes (429) while Serowe produced the least (228), with 356 individuals collected from Palapye (Table 2). A total of four, five and six different species were reported in Serowe, Bobirwa and Palapye respectively (Table 2). The highest species richness was reported for Palapye (0.8511), the least Serowe (0.5526) while Bobirwa was intermediate (0.6599) (Table 2). Palapye exhibited the highest species diversity (1.304), while Serowe reported the least (0.7554) with Bobirwa intermediate (0.9433). The highest evenness was observed for Palapye while both Bobirwa and Serowe reported the least species evenness (Table 2).
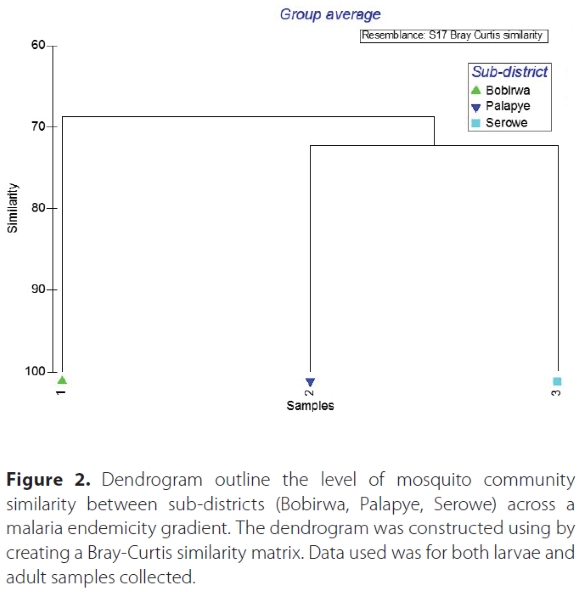
Average pairwise dissimilarity between Bobirwa and Serowe was 31.8%, Bobirwa and Palapye was 30.7% while Palapye and Serowe dissimilarity was 27.7% (Table 3). The dominant species contributing to dissimilarity between Bobirwa and Palapye was Cx. pipiens at 40.3%. Similarly, Cx. pipiens contributed to dissimilarity (59.8%) between Bobirwa and Serowe. For Palapye and Serowe sub-districts, Ae. aegypti contributed to the dissimilarity (21%) (Table 3). Overall, Palapye and Serowe (non-endemic) exhibited similar level of resemblance as opposed to Bobirwa (malaria endemic) (Figure 2).
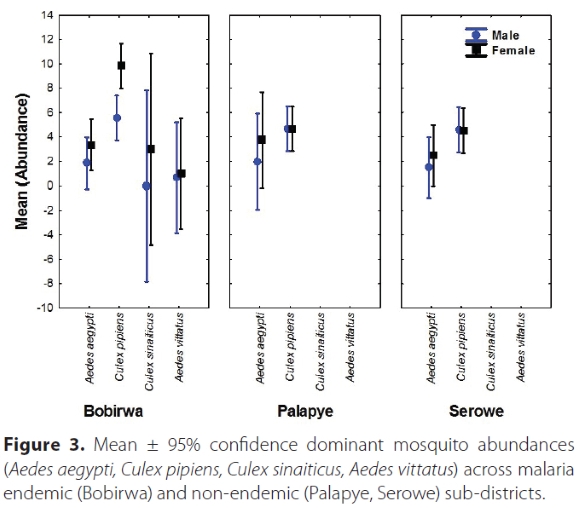
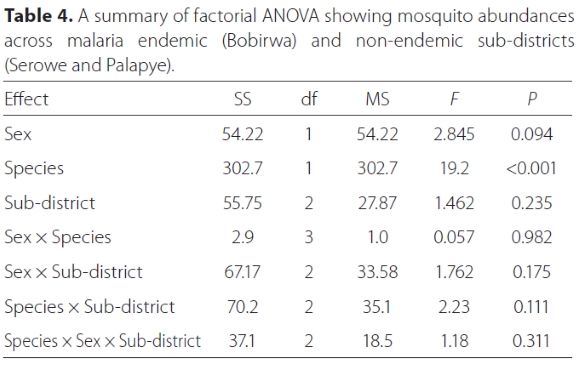
For the adults sampled, the abundances were significantly different for each species with Cx. pipiens exhibiting the highest abundances (Figure 3; Table 4). In addition, neither sex nor sub-districts were statistically significant factors (Table 4). Similarly, the interaction between Sex × Species, Sex × Sub-District, Species × Sub-district and Species × Sex × Sub-district were not significant (Table 4).
DISCUSSION
The study revealed that across the malaria endemic to non-endemic gradient, multiple species of potential vector significance were present with the highest species richness reported in a non-endemic sub-district (Palapye). This study also showed that Cx. pipiens was more abundant compared to other mosquito species sampled. Anopheline species instead, dominated in the non-malaria endemic sub-district (Palapye) while the malaria endemic Bobirwa reported only one anopheline species (An. quadriannulatus). The sub-districts bordering South Africa (Bobirwa and Palapye) as well as Serowe, which is further way, reported invasive Ae. aegypti. The native Ae. vittatus was, however, only reported from Bobirwa. Overall, the abundances of male and female mosquitoes were the same across sub-districts. Key mosquito taxa (Aedes, Anopheles and Culex) were recorded across all the sub-districts with Culiseta spp. reported only in Palapye. The species reported here, have vectorial capacity for infectious diseases such as dengue, yellow fever, Zika, chikungunya (Aedes), malaria (Anopheles) and Rift Valley, West Nile fevers, filariasis, various encephalitis (Culex) and avian malaria (Culiseta and Culex) (Braack et al. 2018; Lee et al. 2018; Ralapanawa & Kularatne 2020). The presence of these incriminating vectors represents a significant biosecurity threat to in the District. In particular, if associated pathogens and parasites are present in the region, this may threaten the health of the affected populations through associated vector-borne disease burdens (Ewen et al. 2012).
All three sampled sub-districts reported Ae. aegypti, a novel invasive species mainly across tropical and sub-tropical regions and originally from West Africa (Powell et al. 2018). The species colonises artificial water holding systems or microenvironments (e.g. used tyres and tins) at disposal of human habitation (Powell & Tabachnick 2013). Despite Ae. aegypti being sampled from homesteads of rural-urban lifestyle, the species has never been assessed for transmission ofpathogens to humans in the country. Thus, future studies should explore pathogen incrimination to assess the risk of infection. Currently, Ae. aegypti is distributed worldwide (e.g. Díez-Fernández et al. 2018) as an invasive species (Alarcón-Elbal et al. 2020) breeding in a variety of habitats (e.g. rock pools, artificial containers, tree holes) (Eshag et al. 2019). The species has the potential to spread of a wide variety of arboviruses to humans, livestock and wildlife (Díez-Fernández et al. 2018). With regard to its role as an emerging threat to public health (see Lwande et al. 2020), a fine-scale distribution of the species countrywide and regionally, is highly warranted. Botswana is a tourism hotspot contributing to the country's second largest economic sector (e.g. ~8.5% of GDP in 2014) after diamonds, receiving an increasing influx of tourists annually (Mbaiwa 2017). Should tourists arrive from regions where dengue and other Ae. aegypti associated diseases are prevalent, there is a chance of importing these pathogens and parasites within human hosts. The presence of healthy populations of Ae. aegypti should be a concern for authorities. Aedes vittatus was also reported in the endemic Bobirwa sub-district, although in fewer abundances compared to the invasive congeneric Ae. aegypti. The species is potentially a significant health threat mainly to humans. For example, Ae. vittatus is known to transmit important pathogens to humans viz, yellow fever, dengue, chikungunya and Zika viruses (Sudeep & Shil 2017).
The study further reported Cx. pipiens as the most abundant and widely distributed species from sampled areas, congruent with Karki et al. (2016). This may be mainly due to the ability of the species to colonise various habitats of diverse water quality (Geoffrey et al. 2017; Amusan & Ogbogu 2020), making them near omnipresent. Although environmental parameters play a crucial role in Cx. pipiens development and growth (Pachka et al. 2016), life stage physiological traits (e.g. aestivation and diapause) and seasonal variability (Ewing et al. 2019) promoting population exacerbation is less explored in the country. Despite the vectorial role of Cx. pipiens in humans (Mavridis et al. 2018) and livestock (Pachka et al. 2016) transmission cycles, it is a key carrier of avian malaria parasites (e.g. Plasmodium) (Schoener et al. 2019). To this end, no study on the local context has explored host-parasite interaction to ascertain the role of Cx. pipiens in avian host species. This is crucial since the country benefits from wildlife through tourism schemes. In addition, Cs. longiareolata reported here is associated with the spread of avian malaria parasites (Brahim et al. 2019). Botswana is a country well-known for its wetlands of importance to migratory bird species (Ramberg et al. 2006) likely to translocate parasites and pathogens in time and space. Therefore, there it is essential to disentangle and understand the functioning of the tripartite ecological interaction in curbing emerging and re-emerging disease associated with wildlife.
Anopheles spp. were reported from all sub-districts regardless of the malaria endemicity status, more so the geographical area is largely similar in terms of land cover and climatic conditions. This has potential for the burden of malaria (Janko et al. 2018), to further extend to the non-endemic areas given the possibility of parasite introduction. More so, Anopheles spp. were reported in Palapye (An. gambiae s.s., An. rhodesiensis, Anopheles sp.) and Serowe (An. rufipes, An. quadriannulatus), both formally designated as non-malarious zones. The prevalence of An. rufipes and An. quadriannulatus reported here, confirm previous studies documenting the same species in the Central District (Tawe et al. 2017; Bango et al. 2020). Anopheles rhodesiensis yielded relatively lower similarity of BLAST (93%) compared to other species identified. This could be due to poor sequence quality that may have resulted in several unassigned nucleotides, which may potentially affect the BLAST algorithm and overall reduced percentage identification output (see discussions in e.g. Afizah et al. 2019). Although An. quadriannulatus is regarded as a non-malaria vector (Lobo et al. 2008), all other Anopheles species sampled in this study are human malaria vectors (e.g. Abdulla-Khan 1998; Sinka et al. 2010; Tawe et al. 2017). Anopheles quadriannulatus was sampled along the river near homesteads, likely to exploit human habitation for dietary and resting needs, in corroboration with reports by Pates et al. (2006). This has implications for its current susceptibility status to Plasmodium parasites (Takken et al. 1999). The study further reports unidentified Anopheles sp. in non-endemic area (Palapye). This warrants taxonomic classification and investigation regarding vector incrimination. In the country, the malaria pathogen (Plasmodium) is mainly vectored by An. arabiensis (Tawe et al. 2017) which was not reported, although other studies documented the species within the Central District and elsewhere in the country (Abdulla-Khan 1998; Bango et al. 2020; Kgoroebutswe et al. 2020).
The species abundances varied across the sub-districts in the current study with Cx. pipiens dominated the dissimilarity. In particular, Cx. pipiens abundances were high in the malaria endemic area. Although seasonal abundance and fine scale distribution of this species is not investigated in Bobirwa, it remains a threat not only as a biting nuisance but also as a potential threat for disease (e.g. avian malaria, West Nile and Rift valley fevers) in the region. The study also highlights that mosquito sex ratios are similar across the sub-districts. However, other studies have demonstrated a skewed collection of significantly high numbers of females compared to males mainly associated with trapping methodology (McPhatter & Gerry 2017).
Different mosquito trapping methodology can potentially yield contrasting species results, as well as numbers (e.g. Lee et al. 2020). However, the present study only used a single Biogents BG-GAT trap type that targets adult mosquitoes. Therefore, the inclusion of other adult sampling techniques should be considered in future studies assessing mosquito diversity in the region. While hay was effectively used to lure many species of adult mosquitoes, not all species may be equally attracted to the trap type used in the study. As such, this could lead to skewed species representation in the sampled sites. An additional limitation in the present study was the lack of extended temporal sampling events and the slight mismatch in temporal synchronicity between sub-districts. This study only sampled austral summer months (November to January) and as such may not have yielded all species that occur in the study region. Similarly, even the marginal temporal separation between sampling of sub-districts could have had implications for species and abundances encountered. To this end, the interpretation of these results should be based on the methodological context presented here.
CONCLUSION
The current mosquito diversity survey of human-inhabited areas outlined the presence of key vector mosquito taxa (Aedes, Anopheles, Culex) across all sampled districts regardless of malaria endemicity status, including the presence of a previously unrecorded invasive species, Ae. aegypti (but see Buxton et al. 2019). More mosquito species were sampled from aquatic habitats (during early life stages) than were adults in the terrestrial environment, highlighting the importance of the incorporation of mosquito breeding sites into monitoring and education programmes. A recent study from the region highlighted that many members of vulnerable human communities are unaware that mosquitoes breed in aquatic habitats (Buxton et al. 2020). Given the reverence of water in this arid region, education and research programmes exploring exclusion of mosquitoes from water bodies in proximity to households could be highly effective for localised mosquito control. However, surveillance of mosquito species also remains essential in the Central District, to strengthen vector management and reduce disease spread.
ACKNOWLEDGEMENTS
We (MB, CN, MK and RJW) acknowledge funding from the Botswana International University of Science and Technology (BIUST) [REF: DVC/2/1/13 XI and DVC/RDI/2/1/7 V (18)] and providing infrastructure for the study. Also, we thank the Ministry of Environment, Natural Resources Conservation and Tourism (Botswana) for issuing of a research study permit (ENT 8/36/4XXXXII(14)). In addition, we sincerely acknowledge the contributions made by A. Moikothai, O. Moikothai and M.P. Buxton in data collection and processing.
CONFLICT OF INTEREST
All authors declare no conflict of interests.
REFERENCES
Abdulla-Khan R. 1998. A survey of the Anopheline mosquito fauna of Botswana, with special reference to the malaria vectors [Ph.D. thesis]. University of the Witwatersrand, Johannesburg, South Africa. [ Links ]
Afizah AN, Torno MM, Jannah JN, Azahari AH, Asuad MK, Nazni WA, Lee HL. 2019. DNA barcoding complementing morphological taxonomic identification of mosquitoes in Peninsular Malaysia. Southeast Asian Journal of Tropical Public Health 50: 36-46. [ Links ]
Ahmed AM, Mohammed AT, Vu TT, Khattab M, Doheim MF, Ashraf Mohamed A, Abdelhamed MM, Shamandy BE, Dawod MT, Alesaei WA, et al.. 2020. Prevalence and burden of dengue infection in Europe: A systematic review and meta-analysis. Reviews of Medical Virology 30(2): e2093. https://doi.org/10.1002/rmv.2093 [ Links ]
Akiner MM, Demirci B, Babuadze G, Robert V, Schaffner F. 2016. Spread of the invasive mosquitoes Aedes aegypti and Aedes albopictus in the Black Sea region increases risk of chikungunya, dengue, and Zika outbreaks in Europe. PLoS Neglected Tropical Diseases 10(4): e0004664. https://doi.org/10.1371/journal.pntd.0004664 [ Links ]
Alarcón-Elbal PM, Rodríguez-Sosa MA, Newman BC, Sutton WB. 2020. The first record of Aedes vittatus (Diptera: Culicidae) in the Dominican Republic: public health implications of a potential invasive mosquito species in the Americas. Journal of Medical Entomology 57(6): 2016-2021. https://doi.org/10.1093/jme/tjaa128 [ Links ]
Alves J, Gomes B, Rodrigues R, Silva J, Arez AP, Pinto J, Sousa CA. 2010. Mosquito fauna on the Cape Verde Islands (West Africa): an update on species distribution and a new finding. Journal of Vector Ecology 35(2): 307-312. https://doi.org/10.1111/j.1948-7134.2010.00087.x [ Links ]
Amusan B, Ogbogu S. 2020. Surveillance of mosquito larvae in various microhabitats in a University Campus in southwestern Nigeria. UNED Research Journal 12(1): e2605. https://doi.org/10.22458/urj.v12i1.2605 [ Links ]
Atoni E, Zhao L, Karungu S, Obanda V, Agwanda B, Xia H, Yuan Z. 2019. The discovery and global distribution of novel mosquito-associated viruses in the last decade (2007-2017). Reviews in Medical Virology 29(6) :e2079. https://doi.org/10.1002/rmv.2079 [ Links ]
Bango ZA, Tawe L, Muthoga CW, Paganotti GM. 2020. Past and current biological factors affecting malaria in the low transmission setting of Botswana: A review. Infection, Genetics & Evolution 85:104458. https://doi.org/10.1016/j.meegid.2020.104458 [ Links ]
Biteye B, Fall AG, Ciss M, Seck MT, Apolloni A, Fall M, Tran A, Gimonneau G. 2018. Ecological distribution and population dynamics of Rift Valley fever virus mosquito vectors (Diptera, Culicidae) in Senegal. Parasites & Vectors 11(1): 27. https://doi.org/10.1186/s13071-017-2591-9 [ Links ]
Boyer S, Marcombe S, Yean S, Fontenille D. 2020. High diversity of mosquito vectors in Cambodian primary schools and consequences for arbovirus transmission. PLoS One 15(6): e0233669. https://doi.org/10.1371/journal.pone.0233669 [ Links ]
Braack L, Gouveia de Almeida AP, Cornel AJ, Swanepoel R, de Jager C 2018. Mosquito-borne arboviruses of African origin: review of key viruses and vectors. Parasites & Vectors 11(1):29. https://doi.org/10.1186/s13071-017-2559-9 [ Links ]
Brahim M, Manel M, Imen M, Laid OM. 2019. Responses of the four larval stages (l1 to l4) of the avian malaria vector Culiseta longiareolata exposed to spinosad (naturally derived insecticide). Advances in Animal & Veterinary Science 7(7): 599-603. https://doi.org/10.17582/journal.aavs/2019/7.7.599.603 [ Links ]
Buxton M, Machekano H, Gotcha N, Nyamukondiwa C, Wasserman RJ. 2020. Are vulnerable communities thoroughly informed on mosquito bio-ecology and burden? Intenational Journal of Environmental Research and Public Health 17(21): 8196. https://doi.org/10.3390/ijerph17218196 [ Links ]
Buxton M, Lebani K, Nyamukondiwa C, Wasserman RJ. 2019. First record of Aedes (Stegomyia) aegypti (Linnaeus, 1762) (Diptera: Culicidae) in Botswana. BioInvasions Records 8(3): 551-557 https://doi.org/10.3391/bir.2019.8.3.10 [ Links ]
Cane RP, Hartley S, Gradwell B, Singe M. 2018. Spatial and temporal distribution, environmental drivers and community structure of mosquitoes in the Kaipara Harbour, New Zealand. Bulletin of Entomology Research 108(3): 305-313. https://doi.org/10.1017/S0007485317000736 [ Links ]
Chirebvu E, Chimbari MJ. Characteristics of Anopheles arabiensis larval habitats in Tubu village, Botswana. Journal of Vector Ecology 2015;40(1): 129-138. https://doi.org/10.1111/jvec.12141
Coetzee M, Craig M, Le Sueur D. 2000. Distribution of African malaria mosquitoes belonging to the Anopheles gambiae complex. Parasitol Today 16(2): 74-77. https://doi.org/10.1016/S0169-4758(99)01563-X [ Links ]
Cornel AJ, Lee Y, Almeida APG, Johnson T, Mouatcho J, Venter M, de Jager C, Braack L. 2018. Mosquito community composition in South Africa and some neighboring countries. Parasites & Vectors 11(1): 331. https://doi.org/10.1186/s13071-018-2824-6 [ Links ]
Díez-Fernández A, Martínez-De La Puente J, Ruiz S, Gutiérrez-López R, Soriguer R, Figuerola J. 2018 Aedes vittatus in Spain: current distribution, barcoding characterization and potential role as a vector of human diseases. Parasites & Vectors 11(1): 297. https://doi.org/10.1186/s13071-018-2879-4 [ Links ]
El-Sayed A, Kamel M. 2020. Climatic changes and their role in emergence and re-emergence of diseases. Environmental Science & Pollution Research Intrenational 27(18): 22336-22352. https://doi.org/10.1007/s11356-020-08896-w [ Links ]
Endo N, Eltahir EA. 2020.Increased risk of malaria transmission with warming temperature in the Ethiopian Highlands. Environmental Research Letters 15(5): 054006. https://doi.org/10.1088/1748-9326/ab7520 [ Links ]
Eshag OS, Bashir NH, Dukeen MY. 2019. Prevalence, habitat and productivity profiles of Aedes mosquitoes (Diptera: Culicidae) in Sennar state, Sudan. International Journal of Mosquito Research 6: 102-108. [ Links ]
Ewen JG, Bensch S, Blackburn TM, Bonneaud C, Brown R, Cassey P, Clarke RH, Pérez-Tris J. 2012. Establishment of exotic parasites: the origins and characteristics of an avian malaria community in an isolated island avifauna. Ecology Letters 15(10): 112-1119. https://doi.org/10.1111/j.1461-0248.2012.01833.x [ Links ]
Ewing DA, Purse BV, Cobbold CA, Schäfer SM, White SM. 2019. Uncovering mechanisms behind mosquito seasonality by integrating mathematical models and daily empirical population data: Culex pipiens in the UK. Parasites & Vectors 12(1): 74. https://doi.org/10.1186/s13071-019-3321-2 [ Links ]
Fălcuţă E, Prioteasa LF, Horváth C, Păstrav IR, Schaffner F, Mihalca AD. 2020. The invasive Asian tiger mosquito Aedes albopictus in Romania: towards a country-wide colonisation? Parasitology Research 119(3): 841-845. https://doi.org/10.1007/s00436-020-06620-8 [ Links ]
Ferguson HM, Dornhaus A, Beeche A, Borgemeister C, Gottlieb M, Mulla MS, Gimnig JE, Fish D, Killeen GF. 2010. Ecology: a prerequisite for malaria elimination and eradication. PLoS Medicine 7(8): e1000303. https://doi.org/10.1371/journal.pmed.1000303 [ Links ]
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. 1994. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294-299. [ Links ]
Foster WA, Walker ED. 2019. Mosquitoes (Culicidae). In: Mullen GR, Durden LA, editors. Medical & Veterinary Entomology 261. Vol. 325. London: Academic Press. https://doi.org/10.1016/B978-0-12-814043-7.00015-7
Geoffrey KK, Ngure NV, Kamau L, Bet D, Lugali R, Wangila A, Kiarie WM. 2017. Survey of urban mosquitoes species (Diptera: Culicidae) with focus on waste water channels as larval habitats in Nairobi industrial area, Kenya. African Journal of Health Sciences 30:120-138. [ Links ]
Gillett JD. 1972. Common African mosquitoes and their medical importance. London: William Heinemann; [ Links ].
Gillies MT, Coetzee M. 1987. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). South African Institute for Medical Research: 55. [ Links ]
Habtewold T, Povelones M, Blagborough AM, Christophides GK. 2008. Transmission blocking immunity in the malaria non-vector mosquito Anopheles quadriannulatus, species A. PLoS Pathogens 4(5): e1000070. https://doi.org/10.1371/journal.ppat.1000070 [ Links ]
Janko MM, Irish SR, Reich BJ, Peterson M, Doctor SM, Mwandagalirwa MK, Likwela JL, Tshefu AK, Meshnick SR, Emch ME. 2018. The links between agriculture, Anopheles mosquitoes, and malaria risk in children younger than 5 years in the Democratic Republic of the Congo: a population-based, cross-sectional, spatial study. Lancet Planet Health 2(2):e74-e82. https://doi.org/10.1016/S2542-5196(18)30009-3 [ Links ]
Jayakrishnan L, Sudhikumar AV, Aneesh EM. 2018. Role of gut inhabitants on vectorial capacity of mosquitoes. Journal of Vector Borne Diseases 55(2): 69. https://doi.org/10.4103/0972-9062.242567 [ Links ]
Johnson T, Braack L, Guarido M, Venter M, Gouveia Almeida AP. 2020. Mosquito community composition and abundance at contrasting sites in northern South Africa, 2014-2017. Journal of Vector Ecology 45(1): 104-117. https://doi.org/10.1111/jvec.12378 [ Links ]
Jupp PG, Mcintosh BM. 1990. Aedes furcifer and other mosquitoes as vectors of chikungunya virus at Mica, northeastern Transvaal, South Africa. Journal of the American Mosquito Control Association 6: 415-420. [ Links ]
Kahamba NF, Limwagu AJ, Mapua SA, Msugupakulya BJ, Msaky DS, Kaindoa EW, Ngowo HS, Okumu FO. 2020. Habitat characteristics and insecticide susceptibility of Aedes aegypti in the Ifakara area, south-eastern Tanzania. Parasites & Vectors 13(1): 53. https://doi.org/10.1186/s13071-020-3920-y [ Links ]
Kampen H, Schuhbauer A, Walther D. 2017. Emerging mosquito species in Germany - a synopsis after 6 years of mosquito monitoring (2011-2016). Parasitology Research 116(12): 3253-3263. https://doi.org/10.1007/s00436-017-5619-3 [ Links ]
Karki S, Hame, GL, Anderson TK, Goldberg TL, Kitron UD, Krebs BL, Ruiz MO. 2016. Effect of trapping methods, weather, and landscape on estimates of the Culex vector mosquito abundance. Environmental Health Insights 10: EHI-S33384. [ Links ]
Kgoroebutswe TK, Ramatlho P, Reeder S, Makate N, Paganotti GM. 2020. Distribution of Anopheles mosquito species, their vectorial role and profiling of knock-down resistance mutations in Botswana. Parasitology Research 119(4): 1201-1208 https://doi.org/10.1007/s00436-020-06614-6 [ Links ]
Lee H, Halverson S, Ezinwa N. 2018. Mosquito-borne diseases. Primary Care: Clinics in Office Practice 45(3): 393-407. https://doi.org/10.1016/j.pop.2018.05.001 [ Links ]
Lee JM, Wasserman RJ, Gan JY, Wilson RF, Rahman S, Yek SH. 2020. Human activities attract harmful mosquitoes in a tropical urban landscape. EcoHealth 17(1): 52-63. https://doi.org/10.1007/s10393-019-01457-9 [ Links ]
Lemine AMM, Lemrabott MAO, Ebou MH, Lekweiry KM, Salem MSOA, Brahim KO, Basco L. 2017. Mosquitoes (Diptera: Culicidae) in Mauritania: a review of their biodiversity, distribution and medical importance. Parasites & Vectors 10(1): 35. https://doi.org/10.1186/s13071-017-1978-y [ Links ]
Liu-Helmersson J, Rocklöv J, Sewe M, Brännström Å. 2019. Climate change may enable Aedes aegypti infestation in major European cities by 2100. Environmental Research 172: 693-699. https://doi.org/10.1016/j.envres.2019.02.026 [ Links ]
Ludwig A, Zheng H, Vrbova L, Drebot MA, Iranpour M, Lindsay LR. 2019. Increased risk of endemic mosquito-borne diseases with climate change. Canada Communicable Disease Report 45: 90-97. [ Links ]
Lwande OW, Obanda V, Lindström A, Ahlm C, Evander M, Näslund J, Bucht G. 2020. Globe-trotting Aedes aegypti and Aedes albopictus: risk factors for arbovirus pandemics. Vector Borne & Zoonotic Diseases 20(2): 71-81. https://doi.org/10.1089/vbz.2019.2486 [ Links ]
Martinet JP, Ferté H, Failloux AB, Schaffner F, Depaquit J. 2019. Mosquitoes of north-western Europe as potential vectors of arboviruses: a review. Viruses 11(11): 1059. https://doi.org/10.3390/v11111059 [ Links ]
Masaninga F, Muleba M, Masendu H, Songolo P, Mweene-Ndumba I, Mazaba-Liwewe ML, Kamuliwo M, Ameneshewa B, Siziya S, Babaniyi OA.2014. Distribution of yellow fever vectors in North-western and Western Provinces, Zambia. Asian Pacific Journal of Tropical Medicine 7: S88-S92. https://doi.org/10.1016/S1995-7645(14)60210-8 [ Links ]
Matowo NS, Abbasi S, Munhenga G, Tanner M, Mapua SA, Oullo D, Koekemoer LL, Kaindoa E, Ngowo HS, Coetzee M, et al. 2019. Fine-scale spatial and temporal variations in insecticide resistance in Culex pipiens complex mosquitoes in rural south-eastern Tanzania. Parasites & Vectors 12(1): 413. https://doi.org/10.1186/s13071-019-3676-4 [ Links ]
Mavridis K, Fotakis EA, Kioulos I, Mpellou S, Konstantas S, Varela E, Gewehr S, Diamantopoulos V, Vontas J. 2018. Detection of West Nile Virus-Lineage 2 in Culex pipiens mosquitoes, associated with disease outbreak in Greece, 2017. Acta Tropica 182 :64-68. https://doi.org/10.1016/j.actatropica.2018.02.024 [ Links ]
Mbaiwa JE. 2017. Poverty or riches: who benefits from the booming tourism industry in Botswana? Journal of Contemporary African Studies 35(1): 93-112. https://doi.org/10.1080/02589001.2016.1270424 [ Links ]
McPhatter L, Gerry AC. 2017. Effect of CO2 concentration on mosquito collection rate using odor-baited suction traps. Journal of Vector Ecology 42(1): 44-50. https://doi.org/10.1111/jvec.12238 [ Links ]
Monaghan AJ, Sampson KM, Steinhoff DF, Ernst KC, Ebi KL, Jones B, Hayden MH. 2018. The potential impacts of 21st century climatic and population changes on human exposure to the virus vector mosquito Aedes aegypti. Climatic Change 146(3-4): 487-500. https://doi.org/10.1007/s10584-016-1679-0 [ Links ]
Mouatassem TF, El Ouali Lalami A, Faraj C, Rais N, Guemmouh R. 2020. Study of abiotic and biotic parameters affecting the abundance of mosquito larvae (Diptera: Culicidae) in the region of Fez (Morocco). International Journal of Zoology 2020: 5429472. https://doi.org/10.1155/2020/5429472 [ Links ]
Mroz C, Gwida M, El-Ashker M, El-Diasty M, El-Beskawy M, Ziegler U, Eiden M, Groschup MH. 2017. Sero-prevalence of Rift Valley fever virus in livestock during inter-epidemic period in Egypt,. BMC Veterinary Research 13(1): 87. https://doi.org/10.1186/s12917-017-0993-8 [ Links ]
Njabo KY, Cornel AJ, Sehgal RNM, Loiseau C, Buermann W, Harrigan RJ, Pollinger J, Valkiünas G, Smith TB. 2009. Coquillettidia (Culicidae, Diptera) mosquitoes are natural vectors of avian malaria in Africa. Malaria Journal 8(1): 193. https://doi.org/10.1186/1475-2875-8-193 [ Links ]
Noden BH, Musuuo M, Aku-Akai L, Van Der Colf B, Chipare I, Wilkinson R. 2014. Risk assessment of flavivirus transmission in Namibia. Acta Tropica 137: 123-129. https://doi.org/10.1016/j.actatropica.2014.05.010 [ Links ]
Ogunmodede AF. 2020. Mosquitoes and their medical importance. Western Journal of Medical & Biomedical Sciences 1(1): 115-120. https://doi.org/10.46912/wjmbs.16 [ Links ]
Pachka H, Annelise T, Alan K, Power T, Patrick K, Véronique C, Janusz P, Ferran J. 2016. Rift Valley fever vector diversity and impact of meteorological and environmental factors on Culex pipiens dynamics in the Okavango Delta, Botswana. Parasites & Vectors 9: 434. https://doi.org/10.1186/s13071-016-1712-1 [ Links ]
Pates HV, Takken W, Curtis CF, Jamet H. 2006. Zoophilic Anopheles quadriannulatus species B found in a human habitation in Ethiopia. Annals of Tropical Medicine & Parasitology 100(2): 177-179. https://doi.org/10.1179/136485906X86374 [ Links ]
Pereira dos Santos T, Roiz D, Santos de Abreu FV, Luz SLB, Santalucia M, Jiolle D, Santos Neves MSA, Simard F, Lourenço-de-Oliveira R, Paupy C. 2018. Potential of Aedes albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerging Microbes & Infections 7(1):1-8. https://doi.org/10.1038/s41426-018-0194-y [ Links ]
Powell JR, Gloria-Soria A, Kotsakiozi P. 2018. Recent history of Aedes aegypti: vector genomics and epidemiology records. Bioscience 68(11): 854-860. https://doi.org/10.1093/biosci/biy119 [ Links ]
Powell JR, Tabachnick WJ. 2013. History of domestication and spread of Aedes aegypti - A review. Memórias do Instituto Oswaldo Cruz 108 suppl 1:11-17. https://doi.org/10.1590/0074-0276130395 [ Links ]
Ralapanawa U, Kularatne S. 2020. Aedes - one mosquito species, a few serious diseases. SF Internal Medicine 1: 1002. [ Links ]
Ramberg L, Hancock P, Lindholm M, Meyer T, Ringrose S, Sliva J, van As J, van der Post C. 2006. Species diversity of the Okavango Delta, Botswana. Aquatic Sciences. 68: 310-337. https://doi.org/10.1007/s00027-006-0857-y [ Links ]
Schoener ER, Harl J, Himmel T, Fragner K, Weissenböck H, Fuehrer HP. 2019. Protozoan parasites in Culex pipiens mosquitoes in Vienna. Parasitology Research 118(4): 1261-1269. https://doi.org/10.1007/s00436-019-06219-8 [ Links ]
Schrama M, Hunting ER, Beechler BR, Guarido MM, Govender D, Nijland W, van 't Zelfde M, Venter M, van Bodegom PM, Gorsich EE. 2020. Human practices promote presence and abundance of disease-transmitting mosquito species. Scientific Reports 10(1): 13543. https://doi.org/10.1038/s41598-020-69858-3 [ Links ]
Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, Patil AP, Temperley WH, Gething PW, Kabaria CW, et al. 2010. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasites & Vectors 3(1): 117. https://doi.org/10.1186/1756-3305-3-117 [ Links ]
Strickman D. 2020. Invasive mosquito species and potential introductions. In: Debboun M, Nava MR, Rueda LM, editors. Mosquitoes, communities, and public health in Texas. London: Academic Press. p. 307-315 https://doi.org/10.1016/B978-0-12-814545-6.00010-9 [ Links ]
Sudeep AB, Shil P. 2017. Aedes vittatus (Bigot) mosquito: an emerging threat to public health. Journal of Vector Borne Diseases 54(4): 295. https://doi.org/10.4103/0972-9062.225833 [ Links ]
Takken W, Eling W, Hooghof J, Dekker T, Hunt R, Coetzee M. 1999. Susceptibility of Anopheles quadriannulatus Theobald (Diptera: Culicidae) to Plasmodium falciparum. Transactions of the Royal Society of Tropical Medicine & Hygiene 93(6): 578-580. https://doi.org/10.1016/S0035-9203(99)90054-8 [ Links ]
Tawe L, Ramatlho P, Waniwa K, Muthoga CW, Makate N, Ntebela DS, Quaye IK, Pombi M, Paganotti GM. 2017. Preliminary survey on Anopheles species distribution in Botswana shows the presence of Anopheles gambiae & Anopheles funestus complexes. Malaria Journal 16(1): 106. https://doi.org/10.1186/s12936-017-1756-5 [ Links ]
Vaux AGC, Dallimore T, Cull B, Schaffner F, Strode C, Pflüger V, Murchie AK, Rea I, Newham Z, Mcginley L, et al. 2019. The challenge of invasive mosquito vectors in the UK during 2016-2018: a summary of the surveillance and control of Aedes albopictus. Medical and Veterinary Entomology 33(4): 443-452. https://doi.org/10.1111/mve.12396 [ Links ]
Wang Y, Yim SHL, Yang Y, Morin CW. 2020. The effect of urbanization and climate change on the mosquito population in the Pearl River Delta region of China. International Journal of Biometeorology 64(3): 501-512. https://doi.org/10.1007/s00484-019-01837-4 [ Links ]
Weyl OLF, Ellender BR, Wasserman RJ, Truter M, Dalu T, Zengeya T, Smit N. 2019. Alien freshwater fauna in South Africa. In: Measy J, van Wilgen B, editors. South African Biological Invasions and their Management. Cham, Switzerland: Springer. p. 153-183. [ Links ]
Wiebe A, Longbottom J, Gleave K, Shearer FM, Sinka ME, Massey NC, Cameron E, Bhatt S, Gething PW, Hemingway J, et al. 2017. Geographical distributions of African malaria vector sibling species and evidence for insecticide resistance. Malaria Journal 16(1): 85. https://doi.org/10.1186/s12936-017-1734-y [ Links ]
Wilke AB, Benelli G, Beier JC. 2020. Beyond frontiers: on invasive alien mosquito species in America and Europe. PLoS Neglected Tropical Diseases 14(1): e0007864. https://doi.org/10.1371/journal.pntd.0007864 [ Links ]
Zengenene MP, Soko W, Brooke BD, Koekemoer LL, Govere J, Mazarire TT, Mberikunashe J, Munhenga G. 2020. Anopheles species composition and breeding habitat characterisation in Chiredzi District, Zimbabwe. African Entomology 28(1): 84-94. https://doi.org/10.4001/003.028.0084 [ Links ]
 Correspondence:
Correspondence:
M. Buxton
Email:lodybuxton@hotmail.co.uk
Received: 20 November 2020
Accepted: 27 September 2021

