










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAfrican Entomology
On-line version ISSN 2224-8854
Print version ISSN 1021-3589
AE vol.30 Pretoria 2022
http://dx.doi.org/10.17159/2254-8854/2022/a13967
RESEARCH ARTICLE
Notes on the lifecycle and distribution of Sphodromantis gastrica (Stâl, 1858) (Mantodea: Mantidae) in South Africa
B Greyvenstein; H du Plessis; J Van den Berg
Unit for Environmental Sciences and Management, North-West University, Potchefstroom, South Africa
ABSTRACT
Limited information is available regarding the distribution of the African mantis, Sphodromantis gastrica (Stal) (Mantodea: Mantidae) and no information is available on its biology. The aim of this study was to determine the distribution of S.gastrica in South Africa based on historic insect collection records and citizen science platforms occurrence data, and to study its basic biology and developmental parameters under captive breeding conditions. A total of 153 South African museum records of Sphodromantis spp. were recorded during this study, while 39 Research Grade observations from iNaturalist were also included. These records indicated that S. gastrica occurred in all provinces of South Africa. The incubation time of the S. gastrica oothecae were approximately 10 weeks and each ootheca contained an average of 84 egg chambers. The mean fertility rate was 54.6% while the survival rate until adulthood was 41.8%. The mean lifespan of S. gastrica individuals were approximately 332 days and females lived longer than males. This study will provide a baseline as it is the first published data on the distribution and biology of this common and charismatic insect in South Africa.
Keywords: African mantis, biology, citizen science, museum records, ootheca
INTRODUCTION
Sphodromantis gastrica (Stâl) (Mantodea: Mantidae), also known as the African/Common green mantis, is one of 38 species in the genus (Roy 2010). Sphodromantis gastrica is suggested to occur throughout sub-Sahara Africa and is one of the largest and most common mantid species in the region (Picker et al. 2019; Erhmann 2002; Kaltenbach 1996). Scientific literature regarding the distribution and biology of this and other Sphodromantis species is scarce and the few studies that were done on species in this genus focussed on behavioural aspects and species interactions.
Ene (1964) described a study that investigated parthenogenesis in Mantodea. In the latter study, the eggs of a Sphodromantis sp. were also artificially inseminated with spermatozoa of Mantis religiosa L. (Mantodea: Mantidae), unfortunately the outcome was not reported or discussed further. Abasa and Mathenge (1971) investigated the use of S. gastrica as a potential biological control agent for the Giant Looper (Ascotis selenaria) (Denis & Schiffermüller) (Lepidoptera: Geometridae) in coffee plantations in Kenya. In the latter study, S. gastrica consumed 44% of the A. selenaria larvae provided to it under laboratory conditions, and was shown to potentially reduce pest infestations by 50% per tree. However, since S. gastrica and mantids in general are not specialist predators various other potentially beneficial insects could also be consumed (Abasa and Mathenge 1971).
A closely related subspecies, the giant African mantis, Sphodromantis viridis Forsskâl (subspecies undetermined but is thought to be S. viridis vischeri) which occurs in North, Central and East Africa has recently been listed as an alien species after it expanded its distribution to various European and Mediterranean countries (Battiston et al. 2020; Oliveira and Ferreira 2019; van der Heyden 2018; Battiston et al. 2017). The seasonality and oviposition site preference of S. viridis was investigated by Younes and Gabre (2003) who reported that it prefers to lay oothecae on stems of Acacia trees instead of on the branches and leaves.
While S. gastrica is listed in various Mantodea checklists which include some notes on their distribution (Patel and Singh 2016; Kaltenbach 1996) information gaps exist regarding the distribution and biology of all Sphodromantis spp. (Roy 2010). In this study we investigated the distribution of S. gastrica in South Africa, based on historic insect collection records in museums and occurrence data from citizen science platforms, as well as the biology and developmental parameters of S. gastrica under captive breeding.
MATERIALS AND METHODS
Species distribution data base
Distribution records of S. gastrica and Sphodromantis spp. were collected during visits to the following institutions that host curated insect collections in South Africa: Ditsong Museum of Natural History (Pretoria) (DNMNH), Agricultural Research Council (Biosystematics Division, Pretoria) (ARC), National Museum (Bloemfontein) (NMB), Albany Museum (Makhanda) (AMG), Rhodes University (Makhanda) (RU), Durban Natural Science Museum (DNSM), Iziko South African Museum (Cape Town) (Iziko) and KwaZulu-Natal Museum (Pietermaritzburg) (NMSA). Most specimens in these collections were previously identified by visiting taxonomists while many were sent for identification to the Vienna Museum in Germany, the University of Drexel in Philadelphia, USA, the Muséum National d'Histoire Naturelle (MNHN) in Paris, France and the research collection of Nicolas Moulin in Montérolier, France.
Sphodromantis specimens and distribution labels where photographed and digitised, after which the data was used to compile a distribution database. This database contains the following information for each specimen record: genus and species name (to the available level of identification), collector's details and collection date where available, and the geo-referenced locality. Scientific literature (Roy 2010; Ehrmann 2002) was used to determine the current nomenclature within the Sphodromantis genus, while the latest updates and changes applied to the classification system of the Order of Mantodea, as described by Schwarz and Roy (2019), were also incorporated in this paper. All locality data was georeferenced using the principles suggested by Wieczorek et al. (2004). Furthermore, the citizen science platform iNaturalist (iNaturalist 2022), in cooperation with the Global Biodiversity Information Facility (Gbif 2022) were used to increase the occurrence data of S. gastrica in South Africa. Only "Research grade" occurrences from the citizen science platforms were included since these records are the most reliable with regards to species identification. Oothecae records from the citizen science platforms were disregarded since it is unreliable in terms of species identification (Brannoch et al. 2017). Decimal degrees were used for developing the distribution maps for S. gastrica and Sphodromantis spp. in South Africa by means of GIS software (ESRI [ArcMaps], Version 10.6.1).
Rearing and biology of Sphodromantis gastrica
Specimens were collected in the Grassland biome in the North West province (Collection Permit number: HQ 08/05/17-152NW) of South Africa during the summer of2016/2017. Adults of these field-collected individuals were mated and nymphs that emerged from oothecae were used to rear a sufficient number of individuals to observe under captive breeding and rearing conditions. A sub-sample of the field-collected specimens was identified by Nicolas Moulin (honorary associate to MNHN), to confirm the species identification.
For breeding purposes, pairs of males and females were placed in glass containers. Large glass containers (30 cm x 20 cm x 20 cm) were used to ensure that ample space was available for the male to avoid sexual cannibalism before, during or after mating. To further limit the likelihood that females would cannibalise the males, ample food was provided before males were introduced into the breeding containers. After copulation concluded, males were removed from the breeding containers. The terrariums (15 cm x 10 cm x 20 cm) in which females were kept after mating was checked daily for the presence of oothecae. Oothecae were removed and put into small containers (5 cm diameter and 5 cm high) inside a desiccator. A humidity level of 68 ± 5% was maintained inside the closed desiccator, following the method described by Solomon (1951). Desiccators were kept in an insect rearing room at a temperature of 27 ± 1 °C with a 14L: 10D photoperiod until nymphs emerged from oothecae.
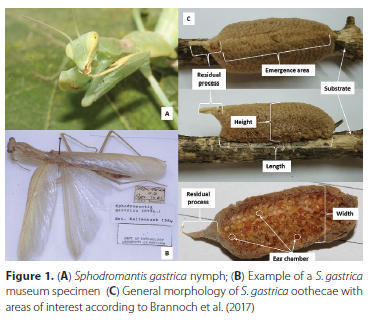
Rearing of nymphs was done under the above-mentioned conditions. Each nymph was placed into a terrarium (7 cm diameter and 15 cm high) with three holes (each 2 cm in diameter) covered with gauze to allow air flow. Thin twigs (5 mm x 10 cm) were placed inside each jar for climbing and hanging purposes, especially during moults. Food was provided every second day when fine water mist was also sprayed into each container. Live crickets (Acheta sp., Orthoptera: Gryllidae) of different sizes (nymphal instars, i.e. pinheads) were provided as food. After moulting to the second-instar, nymphs were removed from the communal terrariums and placed in separate terrariums to prevent cannibalism. Nymphs (Figure 1A) were reared until adulthood after which males and females were identified. This was done based on the number of abdominal segments and the appearance of the wings (McMonigle 2013; Fatimah et al. 2016; Brannoch et al. 2017).
The following life history parameters were recorded during this study: size of oothecae, number of egg chambers inside fertilised and unfertilised oothecae, numbers of days between moults and survival rate (based on nymphs reaching the adult phase). The mean number of days between moults and days to adulthood were calculated separately for males and females. The data were recorded for 63 individuals (22 males and 41 females) that completed their life cycles. Despite 157 nymphs hatching successfully, only 63 reached adulthood. The mean hatch and survival rates as well as duration of male and female life cycles was calculated.
A distinction was also made between different types of oothecae, i.e., fertilised (produced by field collected and laboratory-mated females) and unfertilised (produced by laboratory-reared unmated females). The length, width and height of each ootheca were recorded, based on descriptions by Brannoch et al. (2017). The ootheca length was measured along the area of emergence, excluding the residual process (Greyvenstein et al. 2021; Brannoch et al. 2017). To determine the number of eggs per ootheca, oothecae were dorsally dissected along the length and inspected under a microscope according to the methods described by Greyvenstein et al. (2021; 2020; 2021). Measurements of ootheca parameters were done according to the method proposed by Brannoch et al. (2017) (Figure 1C).
Data analysis
The descriptive statistics (means and standard error) and the statistical analyses of the developmental parameters were done using Statistica Version 13.3 (TIBCO Software Inc. 2017) and SPSS Version 27 (IBM Corp. 2020). Shapiro-Wilk normality test was used to determine if the data was normally distributed. Subsequently, the parametric Student's t-test was used to determine if differences existed between the mean numbers of days between moults, adult longevity and mean number of days required by nymphs to reach adulthood between the males and females of S. gastrica. No statistical tests were done between the unfertilised and fertilised oothecae with regards to length, width, height and number of eggs as a low number (only four) of each oothecae type was collected/produced by female S. gastrica females during the study.
RESULTS
Distribution of Sphodromantis gastrica
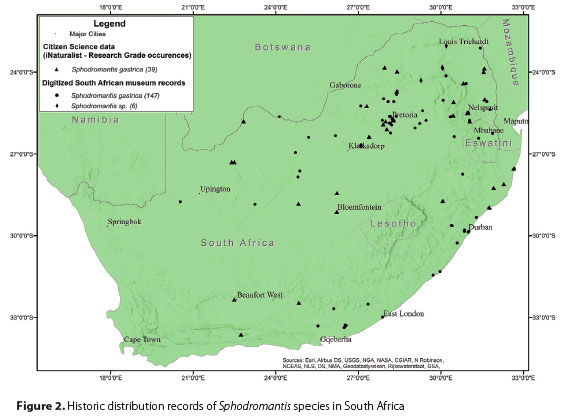
Historic distribution records were compiled from records that are available in the seven South African institutions mentioned above, and iNaturalist observations. The results presented in this paper should be viewed in this context since no museum records were included beyond those residing in South Africa.
A total of 192 records of Sphodromantis spp. specimens were recorded in museum collections and iNaturalist records. Six of the museum records were not identified to species level but were included as distribution records (Figure 2). The majority of the historic distribution records were from the northern parts of the country in the Mpumalanga and Limpopo provinces, with a few records from the Pretoria area in Gauteng. Only 16 records of specimens collected in KwaZulu-Natal province were found while in the Northern and Western Cape provinces only four and two distribution records of S. gástrica were recorded, respectively (Figure 2).
Biology of Sphodromantis gástrica
The oothecae of S. gástrica is somewhat rectangular in shape, with the base being longer than it is wide or high (Figure 1C). The dorsal side of the oothecae is attached to the substrate and in some instances somewhat encircled the substrate, for example in the case of thin branches. The oothecae usually have a light brown colour and a very short residual process and are thus not as elaborate as many other mantid species. The oothecae contained egg chambers arranged in rows following the shape of the oothecae with approximately six egg chambers per row.
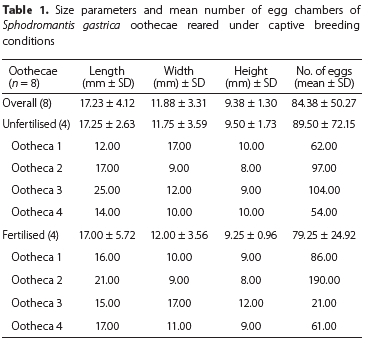
Eight oothecae were used during this study, four of which were produced by four laboratory-mated females and four by unmated laboratory-reared females. Although 41 of the latter females were reared, only four produced oothecae before they died.
The overall morphology, i.e., height, length, width and number of eggs per fertilised and unfertilised oothecae were similar (Table 1). Each ootheca contained approximately 84 eggs. Due to the low number of oothecae collected, morphological parameters could not be statistically compared. Fertilised oothecae were 17.0 mm (± 5.7) long and 12.0 mm (± 3.56) high.
Developmental parameters
A total of 157 nymphs hatched from the four fertilised oothecae. These nymphs were reared to adulthood and provided with food until natural death occurred. Only 63 (22 males and 41 females) completed their life cycle under laboratory conditions.
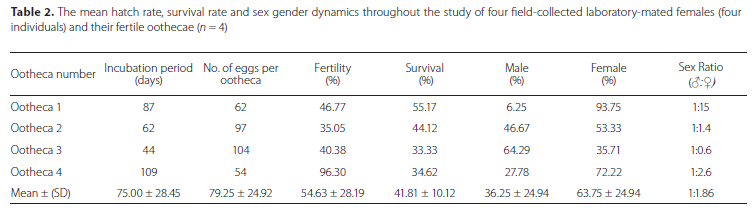
The mean incubation period of S. gástrica oothecae was 75 days (Table 2). However, one ootheca only required 44 days while another only hatched after 109 days, thus indicating great variation in the incubation period despite all oothecae being exposed to similar laboratory conditions. Although the size of the different oothecae were within narrow margins of each other (Table 1), the number of eggs per fertilised ootheca varied between 54 and 104 (Table 2).
The shortest incubation period (44 days) was recorded for the ootheca that contained the largest number of eggs (104 eggs). The mean hatch rate was 54.63% while the mean survival rate to the adult phase was 41.81% (Table 2). More females emerged per ootheca than males and only one ootheca was male dominated (64.2% males and 35.7% females). The overall sex ratio for S. gastrica was female skewed and estimated to be 1:1.86 (Table 2).
Most of the nymphs required seven instars to become adults (37 individuals), with only nine individuals that required more than seven instars (Table 3). Two nymphs required only five instars to become adult females, while one female required nine instars. Significant differences in the duration of only two of the developmental phases between male and female S. gastrica were recorded, i.e. duration of incubation period (p = 0.008) and duration of the 5th instar (p = 0.003) (Table 3). The incubation period of female eggs was significantly longer than that of males and male nymphs took significantly longer than females to complete the 5th instar. Overall, individual S. gástrica nymphs required approximately 251 days (± 9 months) to become adults and their entire lifecycle was completed in approximately 332 days (± 11 months) (Table 3). Although no statistically significant difference was recorded, female adult longevity (mean: 77 days) was longer than that of males (mean: 51 days) (Table 3).
DISCUSSION
Distribution of Sphodromantis gástrica
Distribution records in South African museums together with observation data collected from the citizen science platform (iNaturalist) and previous publications (Picker et al. 2019; Erhmann 2002; Kaltenbach 1996) indicate the distribution of S. gas trica to be throughout East and southern Africa. Kaltenb ach (1996) suggested that S. gástrica is a common and widespread species in southern Africa and also confirmed the presence of this species in Namibia, Zimbabwe and all the provinces of South Africa except Limpopo and the North and Eastern Cape provinces. However, this study confirmed the presence of S. gastrica in the latter provinces. Similar to Kaltenbach (1996), Ehrmann (2002) also listed Namibia, South Africa and Zimbabwe as the distribution area of S. gastrica. Other countries where the presence of S. gastrica was reported are Botswana, Eswatini, Ethiopia, Somalia, Tanzania (including Zanzibar), Uganda and Mozambique (Kaltenbach 1996; Roy 2010; Patel & Singh 2016). According to records of GBIF.org (2022a) the only other Sphodromantis species (excluding S. gastrica) ever recorded in South Africa was Sphodromantis rudolfae Rehn. The two specimens of S. rudolfae that were collected in the "Transvaal" (a province of South Africa prior to 1994) are currently in the Swedish Museum of Natural History (NRM) (Holston 2022G; BIF.org 2022a). This is however in contradiction to the distribution of both subspecies of S. rudolfae as recorded by Erhmann (2002), who reported this species to occur in northeast Africa (Ethiopia, Somalia). Other Sphodromantis species have been recorded in countries that neighbour South Africa are S. lineola (Burmeister) in Mozambique, Zimbabwe and Namibia (GBIF.org 2022b) and S. viridis in Namibia (GBIF.org 2022c). The possibility exists that the latter species occur in South Africa but that they have not been collected yet. According to Erhmann (2002), three other related species occur in South Africa i.e. Sphodromantis gracilis Lombardo, Sphodromantis lineola lineola (Burmeister) and Sphodromantis lineola pinguis (La Greca).
Biology of Sphodromantis gastrica
Although some publications exist on the biology of other mantid species there is no specific information about the biology of S. gastrica. Observations on Sphodromantis spp. in South Africa date as far back as 1898 but are limited to brief reports on feeding and oviposition behaviour. Marshall (1902) reported that Sphodromantis sp. females could survive at least a month of starvation. Unfortunately, the only literature found on the biology of the Sphodromantis genus was from studies done on S. viridis viridis. Zohdy & Younes (2003) studied the biology of S. viridis and reported that females only took approximately 86 minutes to lay an ootheca as opposed to the four hours recorded by Williams & Buxton (1916). Although the study by Williams & Buxton (1916) contained a detailed schematic of the oothecae and protective layers that are present in S. viridis oothecae, no measurements were made and thus cannot be used to compare to results of this study. The oothecae of S. viridis which was studied by Zohdy & Younes (2003) are also visually similar to that of S. gastrica in this study. Unfortunately, no measurements or dissections of oothecae were made by the latter authors, making comparisons impossible. Studies on S. viridis indicated that their oothecae contain approximately 200 eggs (Biernacka 2020), which was also the case for S. lineola (Ene 1964). The latter two species therefore produce much higher number of eggs per oothecae than S. gastrica in this study (84.38 eggs). Zohdy & Younes (2003) recorded an average of ten oothecae per S. viridis female while S. gastrica females in this study only produced one ootheca each.
Developmental parameters
Due to the lack of information on the biology and developmental parameters of Sphodromantis, the biology and nymphal development of S. viridis (Zohdy and Younes) will be used for comparison purposes with the results obtained in this study of S. gastrica.
The mean incubation period for oothecae was 75.00 ± 28.45 days, which is shorter than that recorded for S. viridis (93 days) by Zohdy and Younes (2003) but longer than the average of 57 days recorded for Sphodromantis bioculata (Burmeister) (Adair 1914). Although egg incubation period could be a species-specific trait, it may also be influenced by environmental conditions. The study conducted by Zohdy and Younes (2003) was done at a lower temperature (25 °C ± 2 °C) than that used in this study and at a higher relative humidity (70 ± 5%). Sphodromantis gastrica in this study was kept at a relative humidity of 68 ± 5% for the duration of this study, since this more accurately represents the average conditions in the grassland biome of South Africa, where the specimens of this study were collected. The photoperiod also differed between these two studies with that used by Zohdy and Younes (2003) being a 13L:11D compared to the 14L:10D cycle in this study. These differences in environmental conditions could have influenced several of the biological activities and developmental parameters recorded in the current study and that of Zohdy and Younes (2003) on S. viridis.
The numbers of moults of Sphodromantis spp. seems highly variable. The maximum number ofmoults recorded for S. gastrica in this study was nine, but most individuals required only seven, which is lower than that of S. viridis which become adults after 12 instars (Zohdy and Younes 2003). Higher temperatures do not always result in fewer instars during the nymphal stage. Uvarov (1931) reported that temperature influenced the number of moults of S. viridis and that eight to nine moults were recorded at 25 °C, compared to nine to ten moults at 37 °C. In this study, a longer nymphal developmental period was recorded for S. gastrica males (163 days) and females (149 days) compared to S. viridis males (97 days) and females (103 days). Injury or loss of appendages have also been reported to influence the numbers of moults required by a S. bioculata nymphs to reach adulthood, probably because an extended development period allows time for the injured or lost appendage to be regenerated (Przibram and Megusar 1912).
Abahussain and Younes (2006) reported that high prey availability, especially during the first three instars of S. viridis, was positively related to development rate of nymphs but that development rate decreased when more than a certain number of prey individuals were consumed by mantid nymphs. Despite providing mantid nymphs with the same number of prey individuals, Abahussain and Younes (2006) reported that different developmental rates still occurred, which suggests a natural variation in consumption rates between individuals. Marshall (1902) reported differing behaviour among S. lineola individuals which were offered various butterfly species as prey. Some individuals would eat a certain butterfly with vigour while another would take a bite and drop the prey and start 'wiping its mouth' as if the butterfly was 'disgusting'. Such differences in consumption rates and preference could therefore also influence nymphal developmental rates.
Male S. gastrica took longer to develop into adults than females which is in contrast to the males of S. viridis which developed faster than the females (Zohdy and Younes 2003). Although S. viridis had overall shorter nymphal developmental periods than S. gastrica, the adult longevity of both males and females were almost double that of male and female S. gastrica individuals. The longer adult longevity of S. viridis could also explain the relatively high number of oothecae laid compared to S. gastrica.
Despite these differences between both studies and species (S. gastrica and S. viridis), longevity of female adults was longer than that of males. This is also the case for various other mantid species including Galepsus lenticularis (Saussure) (Greyvenstein et al. 2020), Tarachodes afzelii (Stal) (Ene 1964), Ephestiasula pictipes Giglio-Tos (Vanitha et al. 2016), Harpagomantis tricolor L. (Greyvenstein et al. 2021) and Popa spurca Stal (Greyvenstein et al. 2021). Various reasons for this phenomenon have been suggested, from changing sex ratios from male-dominant to becoming female-dominant and vice versa between seasons and years (Vanitha et al. 2016, Christensen and Brown 2018), to more time for females to mate and oviposit. Extended oviposition may also result in reduced sibling competition if resources are limited (Greyvenstein et al. 2020).
Seasonal changes in environmental conditions were not considered in this study since S. gastrica was reared in captivity and at constant temperature and photoperiod. Mesnier (1984) reported that photoperiod influenced ootheca production in S. lineola and that no egg laying occurred when rearing was done under conditions of continuous light. However, when a sub-sample ofthese females were exposed to a light:dark photoperiod of 12 hours, females immediately laid normal oothecae (Mesnier 1984). Although photoperiod could influence the incubation period of oothecae, other factors such as temperature may also influence the survival strategies of mantids during unfavourable periods. Overwintering strategies of the Mantodea differ between species but little information is available for this group of insects. Some species, for example Miomantis caffra Saussure (Mantidae), go into a facultative diapause phase during the ootheca stage (Ramsay 1984). Shcherbakov and Govorov (2021) reported that Severinia turcomaniae (Saussure) (Toxoderidae) overwintered as late-instar nymphs.
Further research should be conducted to determine the effect of different temperatures on S. gastrica development rates as well as the strategies of this species to survive the wide-ranging environmental conditions throughout its distribution area in South Africa. The occurrence of fire, which is common in the grassland where this species occurs, may also have a significant effect on the ecology of a predatory species such as S. gastrica that has a very long life cycle.
CONCLUSION
This study indicated that S. gastrica occurs throughout South Africa. Museum and iNaturalist records indicate that S. gastrica is the only species of this genus that occurs in South Africa. Although it is likely that other Sphodromantis species also occur in the country, research into this field is hampered by a lack of taxonomic expertise in the region and low numbers of Mantodea in South African museum collections compared to other insect groups. Due to the longevity of S. gastrica and long incubation period of its oothecae, S. gastrica could potentially expand its distribution range throughout Africa, similar to what was recorded for S. viridis in recent years. However, field studies under natural conditions should be conducted in future, as there is no artificial substitute for a species biology as it occurs in nature.
ACKNOWLEDGEMENTS
We would like to thank the following people at each of these institutions for allowing us to access the collections: Audrey Ndaba at DNMNH, Vivienne Uys at ARC (Biosystematics Division), Ashley- Kirk Spriggs and Burgert Muller at NMB, Helen James and Musa Mlambo at AMG, Martin Hill and Thabisa Mdlangu at RU, Kirstin Williams at DNSM, Tricia Pillay at NMSA, Aisha Mayekiso at Iziko. We also thank Simon van Noort at Iziko Museums of South Africa, Entomology Specify 6 database. The National Research Foundation of South Africa contributed funding to this project (Grant number: 101176). Lastly, we thank Nicolas Moulin for identifying the species used in this study.
ORCID IDs
B Greyvenstein - https://orcid.org/0000-0003-2033-7113
H du Plessis - https://orcid.org/0000-0003-1163-1468
J Van den Berg - https://orcid.org/0000-0002-6831-3180
REFERENCES
Abahussain M, Younes AA. 2006. Influence of prey availability on immature survival, development and predation of the mantid predator, Sphodromantis viridis Forskâl. Egyptian Journal of Biological Pest Control. 16: 63-66. [ Links ]
Abasa RO, Mathenge WM. 1971. The praying mantis, Sphodromantis sp. as a predator of the Giant Looper, Ascotis selenaria reciprocaria Walk in Kenya coffee: laboratory evaluation. East African Agricultural and Forestry Journal. 37(2): 177-180. https://doi.org/10.1080/00128325.1971.11662521 [ Links ]
Adair EW. 1914. Notes preliminaires pour servir à l'etude des Mantidae. Bulletin Société Entomologique d'Egypte. 6: 21-36. [ Links ]
Battiston R, Amerini R, Di Pietro W, Guariento LA, Bolognin L, Moretto E. 2020. A new alien mantis in Italy: is the Indochina mantis Hierodula patellifera chasing the train for Europe? Biodiversity Data Journal. 8:e50779. https://doi.org/10.3897/BDJ.8.e50779 [ Links ]
Battiston R, Andria S, Ruzzante G. 2017. The silent spreading of a giant mantis: A critical update on the distribution of Sphodromantis viridis (Forskâl, 1775) in the Mediterranean islands (Mantodea: Mantidae). Onychium. 13: 25-30. [ Links ]
Biernacka N. 2020. Gender relationship and their influence on cannibalism Sphodromantis gastrica and Sphodromantis viridis. Acta Juvenum. 5: 11-16. [ Links ]
Brannoch SK, Wieland F, Rivera J, Klass K, Béthoux O, Svenson GJ. 2017. Manual of praying mantis morphology, nomenclature, and practices (Insecta, Mantodea). ZooKeys. 696: 1-100. https://doi.org/10.3897/zookeys.696.12542 [ Links ]
Holston K. 2022. Entomological Collections (NHRS), Swedish Museum of Natural History (NRM). Version 26.783. Swedish Museum of Natural History. Occurrence dataset https://doi.org/10.15468/fpzyjx. [accessed 11 August 2022].
Christensen T, Brown WD. 2018. Population structure, movement patterns, and frequency of multiple matings in Tenodera sinensis (Mantodea: mantidae). Environmental Entomology. 47(3): 676-683. https://doi.org/10.1093/ee/nvy048 [ Links ]
Ehrmann R. 2002. Mantodea: Gottesanbeterinnen der Welt. Munster, Germany: Natur und Tier Verlag.
Ene JC. 1964. The distribution and post-embryonic development of Tarachodes afzelii (Stâl), (Mantodea: eremiaphilidae). The Annals and Magazine of Natural History. 7(80): 493-511. https://doi.org/10.1080/00222936408651488 [ Links ]
ESRI. 2011. ArcGIS and ArcMAPS Desktop: Release 10. Redlands, California, USA: Environmental Systems Research Institute. [ Links ]
Fatimah S, Sultana R, Wagan MS. 2016. Study on the gender identification of praying mantids (Dictyoptera: mantodea). Journal of Entomology and Zoology Studies. 4: 529-531. [ Links ]
Global Biodiversity Information Facility. (GBIF.org). 2022a. Available at GBIF.org. GBIF Occurrence Download https://doi.org/10.15468/dl.su57zk [accessed 5 August 2022].
Global Biodiversity Information Facility. (GBIF.org). 2022b. Available at GBIF.org. GBIF Occurrence Download https://doi.org/10.15468/dl.kt3h7q [accessed 11 August 2022].
Global Biodiversity Information Facility. (GBIF.org). 2022c. Available at GBIF.org. GBIF Occurrence Download https://doi.org/10.15468/dl.3v2md5. [accessed 11 August 2022].
Greyvenstein B, du Plessis H, Van den Berg J. 2021. Life history of the false flower mantid (Harpagomantis tricolor Linnaeus, 1758) (Mantodea: Galinthiadidae) and its distribution in southern Africa. Journal of Orthoptera Research. 30(1): 17-26. https://doi.org/10.3897/jor.30.52816 [ Links ]
Greyvenstein B, du Plessis H, Moulin N, Van den Berg J. 2020. Distribution of Galepsus spp. in southern Africa and life history of Galepsus lenticularis (Mantodea: Tarachodidae). Insects. 11(2): 119. https://doi.org/10.3390/insects11020119 [ Links ]
Greyvenstein B, du Plessis H, Van den Berg J. 2021. Historic distribution and life history of the African twig mantid, Popa spurca (Mantodea: Deroplatyidae) in southern Africa. Oriental Insects. 55(3): 355-373. https://doi.org/10.1080/00305316.2020.1820916 [ Links ]
IBM Corporation. Released 2020. IBM SPSS Statistics for Windows, Version 27.0. Armonk, New York, USA: IBM Corporation.
INaturalist. 2022. https://www.inaturalist.org. [accessed 29 July 2022].
Kaltenbach AP. 1996. Unterlagen für eine monographie der Mantodea des südlichen Afrika: 1. Artenbestand, geographische Verbreitung und Ausbreitungsgrenzen (Insecta: Mantodea). Annalen des Naturhistorischen Museums in Wien. 98: 193-346. [ Links ]
Marshall GAK. 1902.Five years' observations and experiments (18961901) on the bionomics of South African insects, chiefly directed to the investigation of mimicry and warning colours. Transactions of the Entomological Society of London. 1902 (3): 286-584. Available at https://www.biodiversitylibrary.org/page/13516829#page/357/mode/1up. [ Links ]
Mcmonigle O. 2013. Keeping the praying mantis. Greenville, USA: Coachwhip Publications. [ Links ]
Mesnier M. 1984. Patterns of laying behaviour and control of oviposition in insects: further experiments on Sphodromantis lineola (Dictyoptera). International Journal of Invertebrate Reproduction and Development. 7(1): 23-32. https://doi.org/10.1080/01688170.1984.10510068 [ Links ]
Oliveira D, Ferreira S. 2019. Extension of the known distribution of Sphodromantis viridis (Forskâl, 1775) in Portugal (Mantodea: mantidae). Boletín de la Sociedad Entomológica Aragonesa. 65: 251-252. [ Links ]
Patel S, Singh R. 2016. Updated checklist and distribution of Mantidae (Mantodea: Insecta) of the world. International Journal of Research Studies in Zoology. 2: 17-54. [ Links ]
Picker M, Griffiths C, Weaving A. 2019. Field guide to insects of South Africa. Cape Town, South Africa: Struik Nature [ Links ]
Przibram H, Megusar F. 1912. Wachstumsmessungen an Sphodromantis bioculata Burm. Archiv für Entwicklungsmechanik der Organismen 34(4): 680-741. https://doi.org/10.1007/BF02292211 [ Links ]
Ramsay GW. 1984. Miomantis caffra, a new mantid record (Mantodea: Mantidae) for New Zealand. New Zealand Entomologist. 8(1): 102104. https://doi.org/10.1080/00779962.1984.9722479 [ Links ]
Roy R. 2010. Mises au point sur le genre Sphodromantis Stâl, 1871 (Mantodea, Mantidae). Bulletin de la Société Entomologique de France. 115(3): 345-366. https://doi.org/10.3406/bsef.2010.2692 [ Links ]
Schwarz C, Roy R. 2019. The systematics of Mantodea revisited: an updated classification incorporating multiple data sources (Insecta: Dictyoptera). Annales de la Société Entomologique de France (N.S.). 55: 101196. [ Links ]
Shcherbakov E, Govorov V. 2021. Riders on the storm? A short note on the biology of Severinia turcomaniae (Saussure, 1872) (Mantodea: Toxoderidae). Annales de la Société Entomologique de France (N.S.). 57: 372-378. [ Links ]
Solomon ME. 1951. Control of humidity with potassium hydroxide, sulphuric acid or other solutions. Bulletin ofEntomological Research. 42(3):543-554. https://doi.org/10.1017/S0007485300028947 [ Links ]
TIBCO Software Inc. 2017. California, USA: TIBCO Software Inc.
Statistica (data analysis software system), version 13.3. Available at https://statistica.io. [Accessed 2 January 2022].
Uvarov BP. 1931. Insects and Climate. Transactions of the Entomological Society of London. 79(1): 1-232. https://doi.org/10.1111/j.1365-2311.1931.tb00696.x [ Links ]
Van Der Heyden, T. 2018. An interesting finding of a mantis on Crete/ Greece (Dictyoptera: Mantodea: Mantidae: Mantinae: Paramantini). BV news Publicaciones Científicas. 99: 142-145. [ Links ]
Vanitha K, Bhat PS, Raviprasad TN, Srikumar KK. 2016. Biology and behaviour ofEphestiasula pictipes (Wood-Mason) (Hymenopodidae: Mantodea) under captive breeding. International Journal of Pest Management. 62(4): 308-318. https://doi.org/10.1080/09670874.2016.1195026 [ Links ]
Wieczorek J, Guo Q, Hijmans R. 2004. The point-radius method for georeferencing locality descriptions and calculating associated uncertainty. Transactions of the Entomological Society of London ;18(8):745-767. https://doi.org/10.1080/13658810412331280211 [ Links ]
Williams CB, Buxton PA. 1916. On the biology of Sphodromantis guttata (Mantidae). Transactions of the Entomological Society of London. 64(1): 86-100. https://doi.org/10.1111/j.1365-2311.1916.tb03122.x [ Links ]
Wood-Mason J. 1878. Entomological notes 2. On the presence of a stridulating apparatus in certain Mantidae. Transactions of the Entomological Society of London. 1878: 259-270. [ Links ]
Younes AA, Gabre RM. 2003.Oviposition seasonality and site preference of the predaceous mantis, Sphodromantis viridis Forskâl (Mantodea: Mantidae). Egyptian Journal of Biological Pest Control. 13: 65-69. [ Links ]
Zohdy N, Younes AA. 2003. Biological study on the praying mantis (Sphodromantis viridis Forskâl) {Mantodea: mantidae}. Journal of the Egyptian-German Society of Zoology. 40: 63-76. [ Links ]
 Correspondence:
Correspondence:
B Greyvenstein
Email: biagrey90@gmail.com
Received: 18 May 2022
Accepted: 28 August 2022


{kind=link}
{kind=link}