










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAfrican Entomology
versão On-line ISSN 2224-8854
versão impressa ISSN 1021-3589
AE vol.30 Pretoria 2022
http://dx.doi.org/10.17159/2254-8854/2022/a11778
RESEARCH ARTICLE
Description of the male of Rediviva steineri Kuhlmann, 2012 (Hymenoptera: Melittidae), an endemic oil-collecting bee species from South Africa
Annalie MelinI, II; Jonathan F ColvilleIII
ICompton Herbarium, South African National Biodiversity Institute, Claremont, South Africa
IIAfrican Centre for Coastal Palaeoscience, Nelson Mandela University, Port Elizabeth, South Africa
IIIStatistics in Ecology, Environment and Conservation, Department of Statistical Sciences, University of Cape Town, Cape Town, South Africa
ABSTRACT
We describe the male of Rediviva steineri Kuhlmann, 2012; distinguished by the shape and the margin of the apical plate of the eighth sterna and the vestiture of metasomal terga two and three. Five males were collected at the type locality of the female (Farm Doornbosch, Western Cape, South Africa). We examined the types of three other closely related species (R. pallidula, R. brunnea and R. whiteheadi) and provide images of the eighth sterna for comparison with R. steineri and update Whitehead et al.'s (2008) identification key for the males of these species.
Keywords: Apoidea; Great Escarpment; Melittinae taxonomy
INTRODUCTION
Thirty-three species, in five subgenera of the genus Rediviva Friese (Melittidae: Melittinae) have been described so far (Michez et al. 2009; Kahnt et al. 2017; Kuhlmann et al. 2020). All species are endemic to South Africa and Lesotho and form part of a specialised pollination system with oil-secreting host plants (Vogel 1984; Vogel & Michener 1985). Rediviva have been the subject of detailed taxonomic work (Whitehead & Steiner 1992, 2001; Whitehead et al. 2008; Kuhlmann 2012), pollination coevolution studies (e.g. Steiner & Whitehead 1990, 1991; Pauw 2006; Pauw et al. 2017) and research on trait evolution (e.g. Melin et al. 2020).
One of the most recent species to be described was Rediviva steineri (Kuhlmann 2012), based on a pinned female specimen that was collected several decades ago in 1988 by pollination biologist Kim Steiner and deposited in Iziko Museums of South Africa (Cape Town). In addition to the female holotype, two other female specimens were identified, with the males remaining unknown (Kuhlmann 2012).
As part of a larger study on the summer-rainfall occurring Rediviva, we collected both males (n = 5) and females (n = 6) over a two-day period (23 & 25 March 2021) at the type locality of R. steineri near the town of Murraysburg in the Western Cape. Here we describe the male for R. steineri: the shape of the distal expanded part of the eighth sterna (apical plate) was most important (Whitehead & Steiner 2001) but the form of the sixth and seventh sterna and genitalia contributed to defining the male. We provide photographs of these key features and the habitus of the male. We also compare these to the males of three other closely related summer-rainfall Rediviva species: R. pallidula Whitehead & Steiner, R. brunnea Whitehead & Steiner and R. whiteheadi Kuhlmann.
MATERIAL and METHODS
All specimens are deposited in the Entomology Collection of Iziko Museums of South Africa, Cape Town, South Africa (SAMC). Our specimens were compared to the specimens of closely related species housed in the SAMC collections: the male holotype of R. brunnea (SAM-HYM-B003004), the male paratype of R. whiteheadi (SAM-HYM-B009872) and the male paratype of R. pallidula (SAM-HYM-B007349).
Dissection: Male genitalia and associated sterna were removed from fresh specimens, using a Zeiss Stemi 305 stereo microscope. The dissected parts were extracted with the bent tip of a fine needle and later stored in microvials associated with the specimens concerned. Images were created based on stacked images taken using Leica Application Suite software (Ver. 4.7.1) and a Leica Z16 APO stereoscope. Photographs were enhanced in colour, with the contrast and plates prepared using GIMP (GIMP Development Team, 2.10.20).
Measurements: Measurements were made using the calibrated Leica Z16 APO stereoscope with Leica Application Suite software (Ver. 4.7.1). Body length was measured from the anterior ocellus to the apex of the metasoma (Whitehead & Steiner 2001). We also provide a second standard measure of body size, namely inter-tegula distance (ITD) (Melin et al. 2019), measured between the tegulae at the wing bases. Puncture density was considered as a relationship between puncture diameter (d) and the distance between punctures (i) (Kuhlmann 2012). Punctures were considered sparse if separated by two or more puncture diameters (i > d), moderate if separated by one puncture diameter (i = d) and dense if separated by less than one puncture diameter (i < d).
Abbreviations: The following abbreviations (after Michener 2007) were used for morphological structures: S1, S2, etc. for the first, second, etc., metasomal sterna and T1, T2, etc. for the first, second, etc., metasomal terga.
TAXONOMY
Description of the male Rediviva steineri Kuhlmann, 2012. (Figures 1-4).

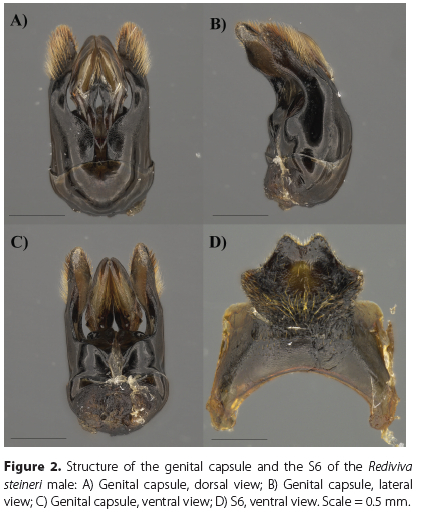
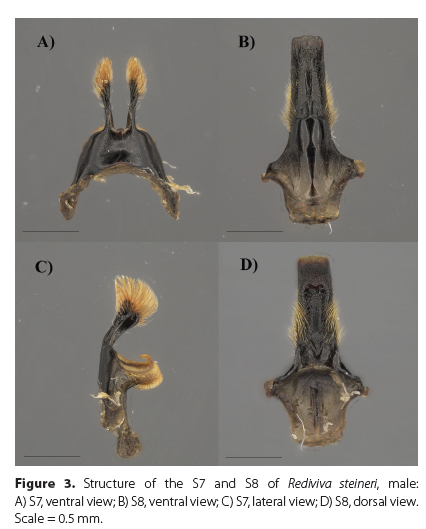

Diagnosis. The male of this species (Figure 1A) is characterized by a short malar space (Figure 1B), T1-T2 integument black (Figure 1C, E), clypeus moderately punctate ventrolaterally (Figure 1F), galea rounded apically, T2-T3 clothed with pale yellowish or orange vestiture (Figure 1C, E) and S8 as in Figures 3B, D, and 4B.
Using the key to Rediviva species of the summer-rainfall region of South Africa (Whitehead et al. 2008), the male of R. steineri runs to R. pallidula but it can be easily distinguished from this species first by the shape of the S8 (Figure 4A, B), followed by the form of the S6-S7 and genitalia (Figures 2-3; Figure 3 in Whitehead & Steiner 1992). The character of yellow or orange vestiture on T3 put forward by Whitehead et al. (2008) appears not to be a reliable separating character due to it being variable in some individuals (Figure 1C, E). In these instances, the male would run to R. brunnea but it can still be easily distinguished from both R. brunnea (Figure 4C) and R. whiteheadi (Figure 4D) by the shape of the S8.
Material examined: (5 specimens) 2 3, South Africa, Western Cape, 39 km east of Murraysburg, Farm Doornbosch, 32°01'25"S 24°06'29"E, 1565m, 23 March 2021, A. Melin, J.F. Colville, Host plant Cyphia triphylla, SAM-HYM-B023086 / SAM-HYM-B023087 (SAMC); 2 3 same data, but 25 March 2021, SAM-HYM-B023085 / B023089 (SAMC); 1 3 same data but host plant Diascia capsularis SAM-HYM-B023088.
Description
Measurements (n = 5). Body length 9.6 mm (range 8.4710.47 mm), inter-tegular distance 2.3 mm (range 2-2.3 mm), malar space (length:width) 0.29 (range 0.24-0.37).
Integumental colour. Head, antennae, mesosoma, metasoma and forelegs black. Tarsus and tibia of hindlegs orange except for a median black area on the tibia of variable size and the first tarsomere, tarsal claw, and femur black. Basitarsus of the midleg orange, tarsomeres variable orange to black, tarsal claw, tibia, and femur black (Figure 1A).
Structure. Metasoma: S6 (Figure 2D) with median and lateral lobes well developed, translucent area on apical third; S7 (Figure 3A, C) median lobes long, spatulate, terminating in long-branched, yellow-orange hairs, lateral lobes large, striate, translucent; S8 (Figure 3B, D) long apical process with sublateral yellow hairs dorsally and ventrally, terminating in a spade-shaped apical plate with a crenulate apical margin. Genital capsule (Figure 2A-C) with gonostylus a little beyond the penis valves, with long branched yellow-orange hairs terminating with stiff piceous unbranched hairs on the apical third; volcellae conspicuous.
Sculpture. Head: Clypeus convex in profile, medially densely punctate, apically impunctate, ventrolaterally moderate; surface between punctures shiny. Mesosoma: Disc of mesoscutum moderately punctate, surface between punctures shiny; scutellum sparsely punctate, anteriorly becoming dense apically, surface between punctures shiny.
Vestiture. Head: Labrum covered with long yellow hairs and fimbria on apical margin orangish; clypeus and frons covered with long branched pale yellow to orange-yellow hairs (Figure 1B), vertex with branched black hairs around the ocelli, with black and yellow hairs intermixed along inner eye margin. Mesosoma: Mesoscutum, scutellum, metanotum, mesepisternum, propodeum covered in orange branched hairs, with sparse erect branched black hairs on the mesoscutal disc (Figure 1D); legs covered with yellow hairs (Figure 1A).
Metasoma: Pale yellow to orange apical tergal hair bands on T1-T6, apical tergal hair band of T1 inconspicuous (Figure 1C, E). Disc on T1 with sparse long sub-erect pale yellow to orange hairs; discs on T2-T3 covered in long pale yellow to orange hairs with median subapical patch of erect black hairs, lateral hairs on T3 sometimes partly intermixed with black hairs; discs on T4-T6 black, fimbria on T7 black.
Host flower records. Four males were collected from the nectar host plant Cyphia triphylla E.Phillips (Lobeliaceae), and one male was collected patrolling the oil-secreting host plant Diascia capsularis Benth. (Scrophulariaceae) for females in the late afternoon.
Distribution. Rediviva steineri is known from three localities along the southern extent of the Great Escarpment of South Africa (Figure 5 in Kuhlmann 2012). At the type locality, three vegetation types (Eastern Upper Karoo, Upper Karoo Hardeveld, Karoo Escarpment Grassland) and two biomes (Nama-Karoo Biome and Grassland Biome) are closely juxtaposed (Mucina & Rutherford 2006). This habitat pattern is also seen for the most western locality for R. stenieri, whereas its most eastern locality is found within a high-altitude area dominated by Karoo Escarpment Grassland vegetation of the Grassland Biome.
Comparative remarks. There is some variation noted in the apical margin of the S8 of R. pallidula (Figure 2D-F in Whitehead & Steiner 1992), which may result in confusion with R. steineri. However, R. pallidula can be clearly differentiated by the hairs on the gonostylus of the genital capsule that project at right angles, the median lobes of the S7 are short, and the median and lateral lobes of the S6 are not well developed (Figure 3 in Whitehead and Steiner 1992). R. steineri was assigned to the subgenus Rediviva s.str. through the associated female (Kuhlmann et al. 2020), here we can confirm that the shape of the median lobes and the presence of the large striate lateral lobes of the S7 are consistent with the subgeneric classification for related males.
Identification key to the males of Rediviva of the summer-rainfall region of South Africa
Revision of the key of Whitehead et al. (2008) for the males from couplet 9 to include the male of R. whiteheadi and R. steineri. For completeness, we include couplets 1 to 8 (Whitehead et al. 2008) for male diagnosis.
1. Malar space long, its length subequal to diameter of antennal scape..............................................................................R. colorata Michener
- Malar space short, shorter than diameter of antennal scape......2
2. Metasomal terga 1-2 partly to completely reddish-orange.............3 - Metasomal terga 1-2 black.....................................................................5
3. Integument of T1-T2 partly orange (orange area interrupted medially with black in R. rufocincta); integument of hind tibia and basitarsus blackish; vestiture on outer surfaces of hind tibia and basitarsus of male R. rufocincta orange and/or black (male of R. transkeiana unknown)...................................................................................4 - Integument of T1-T2 mostly completely orange; integument of hind tibia and basitarsus mostly orangish, basitarsus black distally; male with vestiture on outer surfaces of hind tibia and basitarsus mostly white.........R. rhodosoma Whitehead & Steiner
4. This couplet only relates to female diagnosis of R. rufocincta and R. transkeiana. For male R. rufocincta.............................................................6
5. Clypeus completely, densely punctate ...................................................................R. saetigera Whitehead & Steiner
- Clypeus moderately to sparsely punctate, ventral margin at least impunctate ........................................................................................................ 6
6. Male T2 with hairs on black part of disc black ....................................................................................... R. rufocincta Cockerell
- Male T2 with disc clothed with pallid vestiture................................7
7. Clypeus sparsely punctate ventrolaterally ..............................................................R. autumnalis Whitehead & Steiner - Clypeus moderately punctate ventrolaterally...................................8
8. Galea sharply pointed.................................................R. neliana Cockerell - Galea rounded apically..............................................................................9
9. Male T3 clothed in pale yellowish or orange vestiture....................10
- Male T3 with black or black hairs intermixed with yellow on disc and pale yellowish distal fringe................................................................11
10. Male distal expanded part of S8 (apical plate) with deep median emargination on distal margin (Figure 4A). S6-S7 and genitalia (Figure 3 in Whitehead & Steiner 1992) ....................................................................R. pallidula Whitehead & Steiner - Male distal margin of apical plate of S8 with three shallow emarginations (Figure 4B). S6-S7 and genitalia (Figures 2-3) ...........................................................................................R. steineri Kuhlmann
11. Male distal margin of apical plate of S8 with two shallow emarginations (Figure 4C)...............R. brunnea Whitehead & Steiner- Male distal margin of apical plate of S8 evenly rounded (Figure 4D)............................................................R. whiteheadi Kuhlmann
ACKNOWLEDGEMENTS
This work was supported by the National Research Foundation (PDP postdoctoral fellowship grant no. UID127738 to A.M) and the National Geographic Society (Early Career Grant no. EC-62339R-20 and COVID-19 Support Grant no. NGS-82264R-20 to A.M). Specimens were collected under the following permits: 0821 (Economic Development, Environmental Affairs and Tourism, Eastern Cape); 0056-AAA007-00225 (Cape Nature, Western Cape Province). Our thanks to Simon van Noort and Aisha Mayekiso from Iziko Museums of South Africa for loan of the type material of Rediviva pallidula, R. brunnea and R. whiteheadi. Thanks also goes to Megan Van der Bank (SANBI Marine Programme) for the use of the Leica stereoscope to photograph and measure specimens.
REFERENCES
Kahnt B, Montgomery GA, Murray E, Kuhlmann M, Pauw A, Michez D, Paxton RJ, Danforth BN. 2017. Playing with extremes: Origins and evolution of exaggerated female forelegs in South African Rediviva bees. Molecular Phylogenetics and Evolution 115: 95-105. https://doi.org/10.1016/j.ympev.2017.07.025 [ Links ]
Kuhlmann M. 2012. Two new species of the South African endemic bee genus Rediviva Friese (Hymenoptera: Apoidea: Melittidae). Zootaxa 3517: 71-78. https://doi.org/10.11646/zootaxa.3517.L4 [ Links ]
Kuhlmann M, Jürgensen LS, Michez D. 2020. Subgeneric classification of the bee genus Rediviva Friese (Hymenoptera: Apiformes: Melittidae). Zootaxa 4790: 318-328. https://doi.org/10.11646/zootaxa.4790.2.7 [ Links ]
Melin A, Altwegg R, Manning JC, Colville JF. 2021. Allometric relationships shape foreleg evolution of long-legged oil bees (Melittidae: Rediviva). Evolution 75: 437-449. https://doi.org/10.1111/evo.14144 [ Links ]
Melin A, Krenn HW, Bowie RCK, Beale CM, Manning JC, Colville JF. 2019. The allometry of proboscis length in Melittidae (Hymenoptera: Apoidae) and an estimate of their foraging distance using museum collections. PLoS One 14: e0217839. https://doi.org/10.1371/journal.pone.0217839 [ Links ]
Michener CD. 2007. The bees of the world. Second edition. Baltimore: Johns Hopkins University Press. [ Links ]
Michez D, Patiny S, Danforth BN. 2009. Phylogeny of the bee family Melittidae (Hymenoptera: Anthophila) based on combined molecular and morphological data. Systematic Entomology 34:574597. https://doi.org/10.1111/j.1365-3113.2009.00479 [ Links ]
Mucina L, Rutherford MC (eds). 2006. The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia, Vol. 19, South African National Biodiversity Institute, Pretoria.
Pauw A. 2006. Floral syndromes accurately predict pollination by a specialized oil-collecting bee (Rediviva peringueyi, Melittidae) in a guild of South African orchids (Coryciinae). American Journal of Botany 93: 917-926. https://doi.org/10.3732/ajb.93.6.917 [ Links ]
Pauw A, Kahnt B, Kuhlmann M, Michez D, Montgomery GA, Murray E, Danforth BN. 2017. Long-legged bees make adaptive leaps: linking adaptation to coevolution in a plant - pollinator network. Proceedings of the Royal Society B: Biological Sciences 284: 20171707. https://doi.org/10.1098/rspb.2017.1707 [ Links ]
Steiner KE, Whitehead VB. 1990. Pollinator adaptation to oil-secreting flowers-Rediviva and Diascia. Evolution 44: 1701. https://doi.org/10.2307/2409348 [ Links ]
Steiner KE, Whitehead VB. 1991. Oil flowers and oil bees: Further evidence for pollinator adaptation. Evolution 45: 1493. https://doi.org/10.1111/j.1558-5646.1991.tb02651.x [ Links ]
Vogel S. 1984. The Diascia flower and its bee-an oil-based symbiosis in southern Africa. Acta Botanica Neerlandica 33: 509-518. [ Links ]
Vogel S, Michener CD. 1985. Long bee legs and oil-producing floral spurs, and a new Rediviva (Hymenoptera, Melittidae; Scrophulariaceae). Journal of Kansas Entomological Society 58: 359-364. [ Links ]
Whitehead VB, Steiner KE. 1992. Two new species of oil-collecting bees of the genus Rediviva from the summer rainfall region of South Africa, Hymenoptera, Apoidea, Melittidae. Annals of the South African Museum 102: 143-164. [ Links ]
Whitehead VB, Steiner KE. 2001. Oil-collecting bees of the winter rainfall area of South Africa (Melittidae, Rediviva). Annals of the South African Museum 108: 143-277. [ Links ]
Whitehead VB, Steiner KE, Eardley CD. 2008. Oil collecting bees mostly of the summer rainfall area of southern Africa (Hymenoptera: Melittidae: Rediviva). Journal of Kansas Entomological Society 81: 122-141. https://doi.org/10.2317/jkes-703.12.1 [ Links ]
 Correspondence:
Correspondence:
Annalie Melin
Email:annalie.melin@gmail.com
Received: 20 May 2021
Accepted: 19 August 2021

