










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.44 n.1 Stellenbosch 2023
http://dx.doi.org/10.21548/44-1-5325
ARTICLES
Ch1 (Vitis vinifera L.) Rootstock Control of Scion Response to Water Stress in Some Commercial Grapevine Cultivars
H. RahmaniI; V. RasoliII, *; V. AbdossiI; M. Ghanbari JahromiI
IDepartment of Horticulture, Science and Agronomy, Science and Research Branch, Islamic Azad University, Tehran, Iran
IITemperate Fruit Research Center, Horticultural Sciences Research Institute, Agricultural Research, Education and Extension Organization (AREEO), Karaj, Iran
ABSTRACT
Drought stress is the most important environmental factor limiting the growth, development and yield of crop plants and horticulture. In this study, the induction of drought resistance in commercial cultivars (as scion) by the Chi rootstock was evaluated based on physiological and biochemical responses. Factorial experiment was performed in a completely randomised design (CRD) with three replications in the greenhouse. Plant materials consisted of five commercial grapevine cultivars (own-rooted plants of Black seedless, Flame seedless, Turkman seedless, Sultana and Shahani, and the scion of these cultivars grafted onto Ch1 rootstock). Drought stress treatment comprised stopping irrigation for 30 days from July to August, and not stopping irrigation was the control. Drought stress significantly reduced the membrane stability index (MSI) and the relative water content (RWC), and increased electrolyte leakage (EL), catalase, hydrogen peroxide, proline, ascorbic acid, guaiacol peroxidase, protein, sodium and potassium levels (P < 0.05). The EL and MSI in the Ch1 rootstock under drought stress decreased by 18.38% and 14.86% respectively. The Ch1 rootstock significantly increased the amount of proline, total protein and enzyme activity of guaiacol peroxidase and catalase in cultivars in a drought stress environment. The amount of hydrogen peroxide decreased in all cultivars grafted on Ch1 in both drought stress and non-drought stress environments, by 14.3% and 18.9% respectively. Sultana cultivar grafted on Ch1 rootstock showed the highest drought resistance. Therefore, it can be concluded that the Ch1 rootstock is recommended as a rootstock for inducing drought resistance in grapevine cultivar scions.
Keywords: own rooted grapevine, grafting, drought tolerance, catalase, proline, membrane stability index
INTRODUCTION
Iran is one of the major producers of grapes, with 3.02 million tons of production, and is ranked 10th in the world after China, Italy, the United States of America, Spain, France, Turkey, India, Argentina and Chile (FAO, 2020). In most parts of Iran, as in other parts of the world, drought is the most important environmental limiting factor of the growth, development and yield of crop plants and horticulture (Chaves et al., 2002; Rasoli et al., 2015).
Being one of the centres of origin of grapevine in the world, Iran has a high genetic diversity of grapes. Iranian grapevine cultivars and genotypes are very diverse in tolerating all kinds of biotic and abiotic stresses. Studies conducted to evaluate the drought resistance of commercial Iranian grapevine cultivars show that they have moderate resistance (Rasoli et al., 2015). Therefore, the use of drought-resistant rootstocks to reduce the effects of drought stress has greater importance in the country (Rasoli et al., 2015). Identifying or establishing biotic and abiotic stress-resistant rootstocks is one of the important strategies of fruit tree breeding programmes (Rasoli et al., 2015). Furthermore, the rootstocks directly affect the resistance, adaptability, tolerance and quality of grape production and indirectly affect control of the growth, ripening time and berry size (Shaffer et al., 2004).
Although grapevines are relatively drought-tolerant, a lack of rainfall in the spring and summer causes the growth cycle of grapevines to be negatively affected by high temperatures, increasing evapotranspiration, and finally causing drought stress. Since the response to drought stress varies according to the genetic background of the plant, using drought-resistant genotypes and rootstocks is the one of appropriate strategies to improve fruit trees growing in drought conditions (Cattivelli et al., 2008; Lovisolo et al., 2010; Liu et al., 2012).
In a study of the effect of drought-resistant rootstocks on the Cabernet Sauvignon cultivar, it was reported that 110R, 140Rug, 1103P and 44-53M rootstocks had the highest inducible effect on leaf water potential and the photosynthesis rate of scions in drought stress conditions (Nikolaou et al., 2003).
Grapevine rootstocks of 110R and SO4 have increased drought stress tolerance in Cardinal and Superior seedless scions; of which Cardinal with SO4 and Superior seedless with 110R were the best grafting combinations. In addition, drought tolerance was positively and significantly associated with the accumulation of soluble sugars and proline, as well as the balance of K+ and Na+ in the leaves (Toumi et al., 2007). In addition, Tramontini et al. (2013) reported that there was an effective rootstock-scion interaction to water absorption from the soil and the susceptibility of stomatal control in drought conditions.
The use of 5BB and 99R rootstocks increased drought stress tolerance in Italian grape cultivars compared to own-root seedlings (Sabir, 2016). The effect of the rootstock on yield, vegetative growth rate, yield, pruning weight and berry weight has been demonstrated in the Chardonnay and Cabernet Sauvignon cultivars (Migicovsky et al., 2021). Rootstocks also play an essential role in metabolites in the scion. It has been reported that grapevine scions grafted on 101.14 and M4 rootstocks had different responses in terms of oxidative stress, proteins, secondary metabolism, energy metabolism, hormones, modulation of C metabolism, osmotic responses, levels of ABA, stomatal control and mitochondrial function in the drought stress environment (Prinsi et al., 2018, 2021).
From a physiological point of view, stomatal closure in response to a decline in water status is one of the first responses to water stress to prevent hydraulic failure (Charrier et al., 2018; Cardone et al., 2019). Many scientists have described variation in stomatal control and have proposed a physiological classification of plants as isohydric or anisohydric. Isohydric species can maintain a constant midday leaf water potential (Ψ leaf) by closing their stomata, regardless of soil water availability; whereas anisohydric species maintain a higher stomatal aperture to optimise photosynthetic activity, but their Ψ leaf declines significantly as the soil water deficit increases (Charrier et al., 2018; Cardone et al., 2019). However, this framework is still debated, considering that grapevine cultivars can exhibit, for instance, both near iso- or anisohydric behaviours depending on the environmental conditions (Chaves et al., 2010; Dal Santo et al., 2016). In particular, a key role in the determination of the degree of iso/anisohydricity is played by the hydraulic properties of the soil (Domec & Johnson, 2012; Tramontini et al., 2014; Cardone et al., 2019) and by the rootstock (Merli et al., 2016; Cardone et al., 2019).
Ch1 is one of the Iranian rootstocks of grapevine that is resistant to drought stress, salinity and limey soil (Rasoli & Golmohammadi, 2010). In this study, the inducible effects of Ch1 were evaluated in relation to the physiological and biochemical behaviours of some grapevine cultivars under drought stress.
MATERIALS AND METHODS
Plant materials
Plant materials included Ch1 (as drought-tolerant rootstock) and five commercial grapevine cultivars (Black seedless, Flame seedless, Turkman seedless, Sultana and Shahani).
Ch1 was used as a rootstock and scions of the five cultivars were grafted on it. Own-rooted plants of commercial cultivars also were prepared.
Experimental conditions
Experimental treatments included three factors: a) five cultivars, b) own-rooted plant and grafted on Ch1 rootstock, and c) drought stress (with and without drought stress). All grafted and own-rooted cultivars were planted in 20 litre pots. Each pot was filled with cocopeat and perlite (3:1). A factorial experiment was conducted in a completely randomised design (CRD) with three replications (three pots and two plants in each pot for each treatment). Therefore, there were 60 pots, as follows:
5 (cultivars) × 2 (own-rooted and grafted plant) × 2 (with & without drought stress) × 3 (rep.) = 60
All pots were divided into two groups. The first group was exposed to drought and the second group was without drought stress. The drought stress treatment was performed by stopping irrigation for 30 days, from 15 July to 15 August 2021. The whole experiment lasted eights months - from February to the end of September. Each pot was irrigated with two litres of water every five days. All experiments were carried out in a greenhouse (with a temperature of 32 ± 2°C, humidity of 60%, and natural light).
Traits
Samples were taken from the leaves of each treatment separately on 2021-8-15 and physiological and biochemical studies were performed.
To measure the relative water content (RWC) of the leaves, 0.5 g of leaf disc (fresh weight - FW) was taken from each treatment and replication and kept immersed in distilled water for four hours and the weight was measured again (turgid weight - TW). The leaf specimens were then stored in a 70°C oven for 48 hours to dry completely. After this time, the weight of the samples was measured (dry weight -DW). The RWC was calculated using the following formula (Dhanda & Sethi, 1998).
Sairam's (1994) method was used to measure electrolyte leakage (EL) and the membrane stability index (MSI). According to this method, 0.1 g of leaves of each treatment were placed in 10 ml of twice-distilled water. The samples were immersed in 40°C water in a water bath (Vinteb Co., Ltd, Iran) for 30 minutes, after which their electrical conductivity (EC) was read using an EC meter (C1). The samples were then placed in a water bath at 100°C for 15 minutes and their electrical conductivity was recorded for the second time (C2). EL and MSI were calculated as follows:
MSI = (1 - C1/C2) × 100
EL = C1/C2
The method of Bradford (1976) was used to determine the protein concentration. Bates et al.'s (1973) method was used to measure the proline concentration using a hydrate solution of 3% sulfosalicylic acid. L-proline was used to draw the standard curve of proline at concentrations of 100, 50, 25, 12.5 and 0 μM, and toluene was used as a control (0 level). To measure the amount of hydrogen peroxide, the Alexieva et al. (2001) 0.1% trichloroacetic acid (TCA) method was used. To measure the quantitative concentration of the guaiacol peroxidase enzyme, the method proposed by Chance and Mahli (1995) was used. Nakano and Asada's (1981) method was used to measure the ascorbic peroxidase activity, while Dhindsa et al. (1981) was followed to measure the quantitative concentration of catalase activity. Leaf sodium and potassium were measured as suggested by Hamada and El-Enany (1994), based on leaf ash using a flame photometer (Jenwey Co. Ltd., England).
Statistical analysis
Analysis of variance (ANOVA) was performed using factorial completely randomised design (CRD). An error level of 5% was used for the accuracy of the test. The means were compared using Duncan's multiple range test (DMRT). Statistical analysis was performed by Gen-Stat ver. 12 statistical software. The graphs were drawn using Microsoft Excel software, ver. 2013.
RESULTS AND DISCUSSION
The independent effect means for comparing drought stress are shown in Table 1. Drought stress significantly reduced MSI and RWS, and increased EL, catalase, hydrogen peroxide, proline, ascorbic acid, guaiacol peroxidase, protein, leaf sodium and potassium (P < 0.05).
In the independent comparison of effect means of rootstock types on the studied traits, the mean of RWC, ascorbic acid and potassium in the grafted and own-root treatments was not significantly different (P > 0.05), while there were significant differences (P < 0.05) in the means of EL, MSI, catalase, hydrogen peroxide, guaiacol peroxidase, proline and sodium. The Ch1 rootstock significantly decreased EL, hydrogen peroxide and proline, and significantly increased catalase, MSI, protein and sodium compared to the own-root treatment (P < 0.05, Table 2).
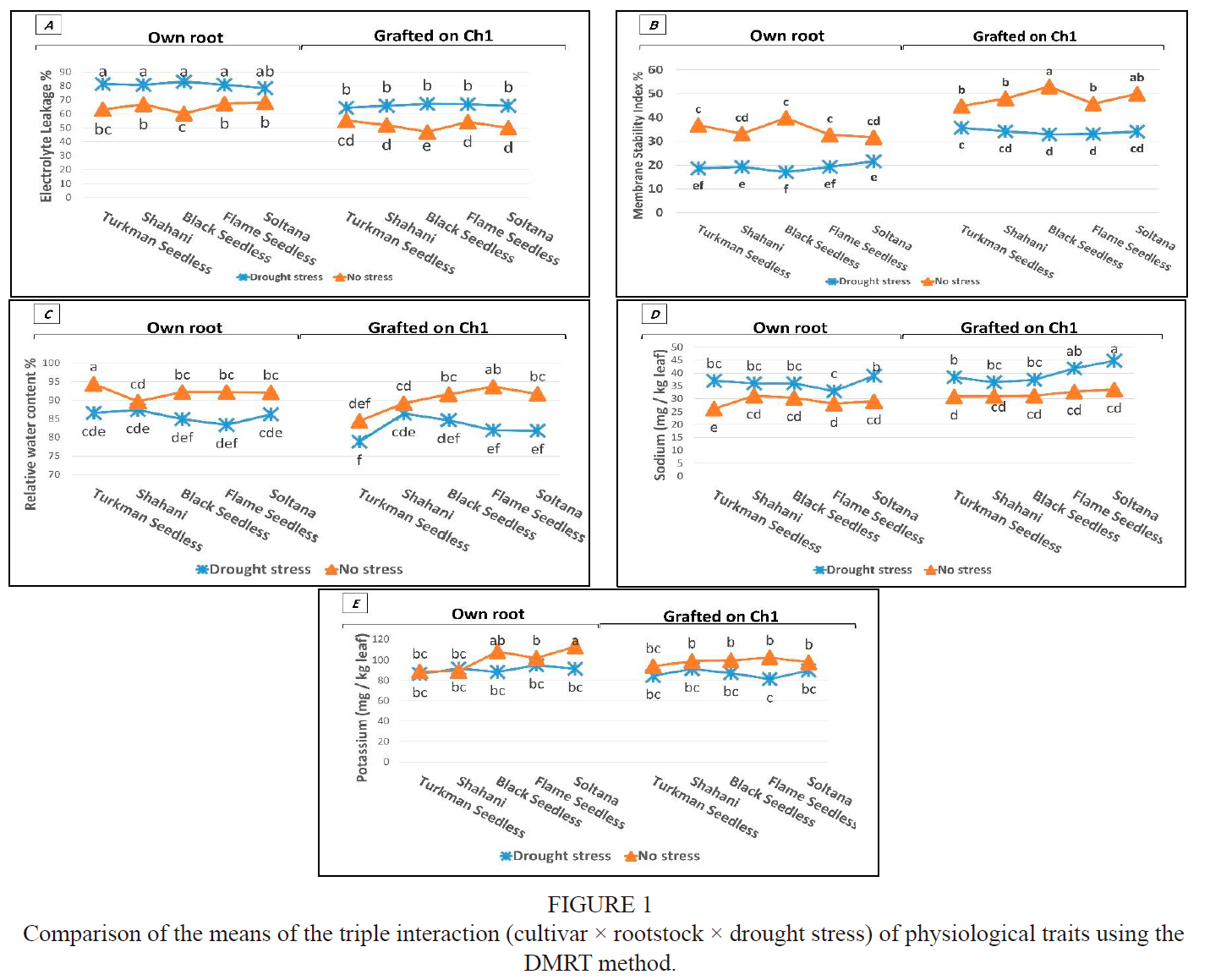
The comparison of the means of the triple interaction (cultivar χ rootstock χ drought stress) of physiological traits is shown in Fig. 1. The Ch1 rootstock reduced the value of EL to 18.38% in the drought stress treatment and to 20.69% in the treatment without drought stress. The lowest EL was in the Black seedless cultivar grafted on Ch1 in the without-stress environment, and the highest was in the own-rooted Black seedless, Shahani and Turkmen cultivars in the drought stress environment (Fig, 1a). The Ch1 rootstock increased the MSI of cultivars with and without drought stress by 14.86% and 13.48%, respectively. The lowest MSI was in the own-rooted Black seedless cultivar in the drought stress environment, and the highest was in grafted Black seedless and Sultana cultivars on Ch1 without drought stress (Fig. 1b). As drought stress increases, the amount of EL increases. Electrolyte leakage is associated with the preservation and integration of cell membranes under drought stress. A significant increase in electrolyte leakage in cultivars is most likely due to the high sensitivity of these cultivars to water deficiency and leaf water loss.
Similar studies have reported that it is more common to find preservation and integration of cell membranes in the leaves of drought-tolerant plants (Bajji & Kinet, 2002). Cell membranes are very sensitive to dehydration and are rapidly damaged in trees exposed to water stress. This damage varies in different cultivars (Karimi et al., 2015). Drought stress, like other abiotic stresses, leads to secondary oxidative stress by increasing the formation of reactive oxygen species (ROS). The accumulation of reactive oxygen species leads to peroxidation of the membrane lipids, destruction of the membranes and photosynthetic apparatus, and finally to the decomposition of chlorophyll (Noctor et al., 2014). A reduction of MSI in the leaves of Black seedless and Turkman seedless cultivars occurs simultaneously with a reduction in leaf RWC and cell dehydration under drought stress conditions. Previously, Sivritepe et al. (2008) and Karimi et al. (2015) reported the reduction of MSI due to drought stress in almonds and cherries.
The Ch1 rootstock only increased RWC by 1.5% in Flame seedless without drought stress. In other words, the Ch1 rootstock had no significant effect on the RWC of cultivars in different environments compared to own-rooted cultivars, but there was a significant difference between the cultivars. The highest and lowest RWC were observed in own-rooted Turkman seedless in a non-stress environment, and Turkman seedless grafted on Ch1 in a drought stress environment respectively (Fig. 1c). RWC is one of the most important, reliable and widely used indicators to identify drought-tolerant or sensitive cultivars. Grapevine plants with a strong stomatal control system are among the isohydric plants (Arve et al., 2011). This group of plants, in a deficiency of soil water, quickly close their stomata and prevent a further reduction in leaf water potential (Hugalde & Vila, 2014). Relatively high values of leaf RWC in Flame seedless and Tukman seedless cultivars can be attributed to the isohydric nature of the grapevine. The lack of effect of the Ch1 rootstock on RWC is probably due to the dependence of this index on the anatomical structure of the leaf. A high RWC means that the leaf has the ability to retain more water under stress (Chakraborty & Pradhan, 2011). Drought-tolerant cultivars maintain the water content of their cells at a higher level (Kafi et al, 2009).
Flexas and Medrano (2002) report that the RWC does not decrease significantly until the plant is exposed to severe drought stress, so cultivars that are able to maintain a relative amount of water at higher levels of drought stress can be introduced as tolerant cultivars. The amount of RWC in the leaves may be due to osmotic regulation or the roots' ability to absorb water (Arve et al., 2011). In a similar study, involving a 17-day stress period in grapevine cultivars, it was found that the RWC of one South Korean cultivar was less affected by drought stress than two commercial cultivars (Choi et al, 2013).
The Ch1 rootstock significantly increased the leaf sodium content of cultivars in both environments, which an increase in the stress and non-stress environments of 9.9% and 10.1%, respectively. The highest and lowest sodium levels were observed in Sultana grafted on Ch1 and in own-rooted Turkmen seedless, respectively (Fig. 1d). However, the Ch1 rootstock did not have a significant effect on the leaf potassium of cultivars in both environments compared to own-rooted cultivars (Fig. 1e). However, all concentrations of sodium and potassium in the leaves were within the normal range (Gärtel, 1996). This confirms the potential of the Ch1 rootstock to obtain balanced sodium to survive water stress, as previously reported by Tramontini et al. (2013). The effects of drought stress on grapevine nutrient uptake have been investigated in several independent studies, while the interaction between genotype and water deficiency is less well known (Kuster et al., 2013). In this study, the relatively higher mineral uptake by Sultana grafted on Ch1 may indicate a competitive uptake of sodium and potassium minerals under water scarcity.
The comparison of the mean of the triple interaction (cultivar × rootstock × drought stress) in biochemical traits is shown in Fig. 2. The Ch1 rootstock significantly increased the leaf catalase of cultivars in both environments, which an increase of 30.4% and 22.3% in the stress and non-stress environments respectively. The highest and lowest leaf catalase levels were observed in all cultivars grafted on Ch1 and in own-rooted Shahani, respectively (Fig. 2a). The amount of ascorbic peroxidase was significantly different in the cultivars, but the rootstock type had no significant effect on the amount of ascorbic peroxidase in the grapevines with and without drought stress (Fig. 2d). The amount of guaiacol peroxidase in cultivars grafted on Ch1 was significantly higher than in own-rooted cultivars in the drought stress environment, with 44.8% and 33.3% of guaiacol peroxidase in cultivars grafted on Ch1 in the drought (stress) and non-drought environments, respectively (Fig. 2e). The increased activity of antioxidants during drought stress indicates plant tolerance to drought stress (Noctor et al., 2014). The induction of antioxidant enzyme activity is a general adaptation strategy that plants use to overcome damage caused by oxidative stress (Noctor et al., 2014). Catalase, ascorbic peroxidase and guaiacol peroxidase are antioxidant enzymes that increase their activity under stress (Sharma & Dubey, 2005; Wang et al., 2005; Chakraborty & Pradhan, 2011).
The coordinated activity of these enzymes may act as an effective antioxidant defence against the effects of reactive oxygen species under drought stress conditions. The enzymes of a number of plants regulate the intracellular level of hydrogen peroxide, in which catalase, guaiacol peroxidase and ascorbic peroxidase are the most important (Noctor & Foyer, 1998). In the present study, the higher activity of antioxidant enzymes in cultivars grafted on Ch1 could help protect cell membranes and photosynthetic apparatus against oxidative damage and give these cultivars greater tolerance of drought stress. Lower values of oxidative stress indexes such as EL and hydrogen peroxide in Flame seedless grafted on Ch1 can also be attributed to the higher activity and better efficiency of the antioxidant enzymes. Higher activity of antioxidant enzymes in drought-resistant cultivars has been reported in relation to other types of environmental stresses, such as cold and salinity (Wang et al., 2009b).
In the present study, the amount of hydrogen peroxide was reduced in all the grapevine cultivars grafted on Ch1 in both environments, with reductions in the stress and non-stress environments of 14.3% and 18.9%, respectively. The highest and lowest levels of hydrogen peroxide were obtained in own-rooted and Sultana grafted on Ch1, respectively (Fig. 2b). The normal activity of cellular metabolism undergrowth periods leads to the regular production of reactive oxygen radicals. Biotic and abiotic stresses cause an imbalance between the production and purification of oxygen free radicals and thus increase their concentration in the cell (Kar, 2011). An increased content of hydrogen peroxide under drought conditions, especially in more sensitive cultivars and genotypes, had been confirmed in many studies (Chakraborty & Pradhan, 2011; Wang et al., 2012). The low amount of hydrogen peroxide can be due to the better performance of the antioxidant defence system (CAT and POD enzymes) in resistant cultivars (Chakraborty & Pradhan, 2011; Wang et al., 2012).
Drought stress increased the proline content by 29.6% compared to levels in the non-stress environment. The amount of proline in all grapevine cultivars grafted on Ch1 increased compared to the amount in own-rooted cultivars, with an increase of 32.8% in the environment with drought stress and 27.1% in the environment without drought stress. The highest and lowest proline levels under drought stress conditions were observed in Black seedless cultivar grafted on Ch1 and own-rooted Black seedless respectively. In a stress-free environment, the highest and lowest proline levels were observed in Black seedless grafted on Ch1 and own-rooted Turkmen seedless cultivars, respectively (Fig. 2c). The amount of proline in the grapevine increases with increasing drought stress. Compatible solvents such as proline do not interfere with normal cell biochemical reactions and act as osmotic protectors during osmotic stress. The physiological action of proline, sugars and other osmotic compounds maintains the stability of cell membranes due to their inhibitory effect on bonding between adjacent membranes during periods of stress. They also store lipids and ensure the stability of proteins through the formation of hydrogen bonds with linear proteins, gene regulation, and osmotic regulation (Shariat & Assareh, 2006). In addition to the finding in the present study, that there is an increase in proline content if drought stress is intensified, this situation has also been observed in studies on other plants such as cherries (Sivritepe et al., 2008), corn (Helal & Abdolaziz, 2008) and rice (Wang et al., 2009a).
Total protein increased significantly in cultivars grafted on Ch1. The Ch1 rootstock increased total protein by 22.1% and 26.4% in environments with and without drought stress, respectively. The highest total protein content was observed in Sultana grafted on the Ch1 cultivar under drought stress and the lowest was found in own-rooted Flame seedless without drought stress (Fig. 2f). The soluble protein content is one of the traits in plants affected by stress, and it decreases with increasing drought stress (Gill & Tuteia, 2010). The reason for this decrease is the increase in the activity of protein-degrading enzymes such as protease and the accumulation of amino acids such as proline. However, the use of the Ch1 rootstock in the present study prevented the reduction in total protein and subsequently reduced the effect of drought stress on the grafted cultivars. The reduction protein was less in cultivars grafted on Ch1 than in own-rooted cultivars, which may be due to a reduction in the complex variability of protein materials under drought stress (Bertamini et al., 2006), which indicates that cultivars grafted on Ch1 are less sensitive than own-rooted cultivars in drought stress condition.
CONCLUSIONS
The results of this study show that the Ch1 rootstock decreased EL and increased MSI by increasing osmotic regulation and the proper accumulation of sodium during drought stress in the studied cultivars. This indicates a high correlation between the decrease in EL and the increase in MDI and sodium storage in the cell wall, which prevents cell wall degradation during drought stress. In addition, when the level of antioxidant activity and proline content in the Ch1 rootstock was increased, it increased the tolerance of grafted cultivars in a drought stress environment, which in turn reduced hydrogen peroxide content and thus reduced cell damage during drought stress. In other words, there is an negative relationship between the increase in antioxidant activity and proline and the decrease in hydrogen peroxide content. On the other hand, unlike in other studies, the use of the Ch1 rootstock was found to prevent the reduction in total protein and subsequently reduced the effect of drought stress on grafted cultivars. Therefore, the Ch1 rootstock can be recommended as a rootstock for inducing drought resistance in grapevine cultivars. Among the grapevine cultivars, Sultana grafted on Ch1 rootstock showed the highest resistance to drought stress.
LITERATURE CITED
Alexieva, V., Sergiev, I., Mapelli, S. & Karanov, E., 2001. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 24(12), 1337-1344. [ Links ]
Arve, L.E., Torre, S., Olsen, J.E. & Tanino, K.K., 2011. Stomata responses to drought stress and air humidity. In: Shanker, A. & Venkateswarlu, B., (Eds) Abiotic Stress in Plants- Mechanisms and Adaptations. InTech Open Access Publisher, Vienna, Austria. pp. 42-44. [ Links ]
Bajji, M. & Kinet, J.M.S., 2002. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance in durum wheat. Plant Growth Regul. 36, 61-70. [ Links ]
Bates, L.S., Waldern, R.P. & Teave, I.D., 1973. Rapid determination of free proline for water-stress studies. Plant Soil 39, 205-207. [ Links ]
Bertamini, M., Zulini, L., Muthuchelian, K. & Nedunchezhian, N., 2006. Effect of water deficit on photosynthetic and other physiological responses in grapevine (Vitis vinifera L. cv. Riesling) plants. Photosynthetica 44, 151-154. [ Links ]
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principles of protein-dye binding. Anal. Biochem. 72, 248-254. [ Links ]
Cardone, M.F., Perniola, R., Catacchio, C.R., Alagna, F., Rotunno, S., Crupi, P., Antonacci, D., Velasco, R., Ventura, M. & Bergamini, C., 2019. Grapevine adaptation to drought: New candidate genes for the genotype-dependent response. BIO Web Conf. 15, 01016. [ Links ]
Cattivelli, L., Rizza, F., Badeck, F.W., Mazzucotelli, E., Mastrangelo, A.M. & Francia, E., 2008. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 105, 1-14. [ Links ]
Chakraborty, U. & Pradhan, B., 2011. Drought stress-induced oxidative stress and antioxidative responses in four wheat (Triticum aestivum L.) varieties. Arch. Acker Pflanzenbau Bodenkd. 58(6), 617-630. [ Links ]
Chance, B. & Maehly, A.C., 1995. Assay of catalase and peroxidase. In: Culowic, S.P. & Kaplan, N.O. (eds). Methods in enzymology, vol. 2. Academic Press. Inc., New York. pp. 764-765. [ Links ]
Charrier, G., Delzon, S., Domec, J.C., Zhang, L., Delmas C.E.L. & Merlin, I., 2018. Drought will not leave your glass empty: Low risk of hydraulic failure revealed by long-term drought observations in world's top wine regions. Sci. Adv. 4, 1-10. [ Links ]
Chaves, M.M., Pereira, J.O.S., Maroco, J.O., Rodrigues, M.L., Ricardo, C.N.P.P. & Osorio, M. L., 2002. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot. 89, 907-916. [ Links ]
Chaves, M.M., Zarrouk, O., Francisco, R., Costa, J.M., Santos, T. & Regalado, A.P., 2010. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 105(5), 661-76. [ Links ]
Choi, Y.J., Hur, Y.Y., Jung, S.M., Kim, S.H., Noh, J.H., Park, S.J. & Yun, H.K., 2013. Transcriptional analysis of Dehydrin1 genes responsive to dehydrating stress in grapevines. Hortic. Environ. Biotechnol. 54, 272-279. [ Links ]
Dal Santo, S., Palliotti, A., Zenoni, S., Tornielli, G.B., Fasoli, M. & Paci, P., 2016. Distinct transcriptome responses to water limitation in isohydric and anisohydric grapevine cultivars. BMC Genom. 17, 815-820. [ Links ]
Dhanda, S.S. & Sethi, G.S., 1998. Inheritance of excised-leaf water loss and relative water content in bread wheat (Triticum aestivum). Euphytica 104, 39-47. [ Links ]
Dhindsa, R.S., Plumb-Dhindsa P. & Thorpe, T.A., 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 32, 93-101. [ Links ]
Domec, J.C. & Johnson, D.M., 2012. Does homeostasis or disturbance of homeostasis in minimum leaf water potential explain the isohydric versus anisohydric behavior of Vitis vinifera L. cultivars? Tree Physiol. 32, 245-252. [ Links ]
FAO. 2020. Grape production statistics. Food and agriculture organization of the United Nations. Rome, Italy. [ Links ]
Flexas, J. & Medrano, H., 2002. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitation revisited. Ann. Bot. 89, 183-189. [ Links ]
Gärtel, W., 1996. Grapes. In: Bennett, W.F. (ed). Nutrient deficiencies and toxicities in crop plant. APS, St. Paul, MN. pp. 177-183. [ Links ]
Gill, S.S. & Tuteja, N., 2010. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909-930. [ Links ]
Hamada, A.M. & El-Enany, A.E., 1994. Effect of NaCl salinity on growth, pigment and mineral element contents, and gas exchange of broad bean and pea plants. Biol. Plant. 36, 75-81. [ Links ]
Helal, R.M. & Abdolaziz, S.M., 2008. Comparative response of drought tolerant and drought sensitive maize genotypes to water stress. Aust. J. Crop Sci. 1, 31-36. [ Links ]
Hugalde, I.P. & Vila, H.F., 2014. Isohydric or anisohydric behavior on vines an endless controversy? Rev. Investig. Agropec. 40(1), 75-82. [ Links ]
Kafi, M., Buarzouei, A., Salehi, M., Kamandi, A., Masoumi, A. & Nabati, J., 2009. Physiology of environmental stresses in plants. Mashhad University Jihad Press, Mashhad, Iran. pp. 451-455. [ Links ]
Kar, R.K., 2011. Plant responses to water stress: Role of reactive oxygen species. Plant Signal. Behav. 6, 1741-1745. [ Links ]
Karimi, S., Yadollahi, A., Arzani, K., Imani, A. & Aghaalikhani, M., 2015. Gas-exchange response of almond genotypes to water stress. Photosynthetica 53, 29-34. [ Links ]
Kuster, T.M., Arend, M., Günthardt-Goerg, M.S. & Schulin, R., 2013. Root growth of different oak provenances in two soils under drought stress and air warming conditions. Plant Soil 369, 61-71. [ Links ]
Liu, B.H., Cheng, L., Liang, D., Zou, Y.J. & Ma, F.W., 2012. Growth, gas exchange, water-use efficiency, and carbon isotope composition of 'Gale Gala' apple trees grafted onto 9 wild Chinese rootstocks in response to drought stress. Photosynthetica 50, 401-410. [ Links ]
Lovisolo, C., Perrone, I., Carra, A., Ferrandino, A., Flexas, J. & Medrano, H., 2010. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 37, 98-116. [ Links ]
Merli, M.C., Magnanini, E., Gatti, M., Pirez, F.J., Pueyo, I.B., Intrigliolo, D.S. & Poni, S., 2016. Water stress improves whole-canopy water use efficiency and berry composition of cv. Sangiovese (Vitis vinifera L.) grapevines grafted on the new drought-tolerant rootstock M4. Agric. Water Manag. 169, 106-111. [ Links ]
Migicovsky, Z., Cousins, P., Lindsay, M.J., Myles, S., Striegler, R.K., Verdegaal, P. & Chitwood, D.H., 2021. Grapevine rootstocks affect growth-related scion phenotypes. Plant Direct 5(5), e00324. [ Links ]
Nakano, Y. & Asada, K., 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867-880. [ Links ]
Nikolaou, N., Angelopoulos, K. & Karagiannidis, N., 2003. Effects of drought stress on mycorrhizal and non-mycorrhizal Cabernet Sauvignon grapevine, grafted onto various rootstocks. Exp. Agric. 39(3), 241-252. https://doi.org/10.1017/S001447970300125X [ Links ]
Noctor, G. & Foyer, C.H., 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 49, 249-279. [ Links ]
Noctor, G., Mhamdi, A. & Foyer, C.H., 2014. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 164, 1636-1648. [ Links ]
Prinsi, B., Simone, N.A., Failla, O., Scienza, A. & Espen, L., 2018. Root proteomic and metabolic analyses reveal specific responses to drought stress in differently tolerant grapevine rootstocks. BMC Plant Biol. 18, 126-134. [ Links ]
Prinsi, B., Simeoni, F., Galbiati, M., Meggio, F., Tonelli, C., Scienza, A. & Espen, L., 2021. Grapevine rootstocks differently affect physiological and molecular responses of the scion under water deficit condition. Agronomy 11, 289-298. [ Links ]
Rasoli, V. & Golmohammadi, M., 2010. Evaluation of drought tolerance of grape cultivars in Qazvin province. Seed Plant Improv. 1, 359-349. [ Links ]
Rasoli, V., Farshadfar, E. & Ahmadi, J., 2015. Evaluation of genotype χ environment interaction of grapevine genotypes (Vitis vinifera L.) by non parametric method. J. Agr. Sci. Tech. 17, 1279-1289. [ Links ]
Sabir, A., 2016. Vegetative and reproductive growth responses of grapevine cv. 'Italia' (Vitis vinifera L.) grafted on different rootstocks to contrasting soil water status. J. Agr. Sci. Tech. 18, 1681-1692. [ Links ]
Sairam, R.K., 1994. Effect of moisture stress on physiological activities of two contrasting wheat genotypes. Indian. J. Exp. Biol. 32, 584-593. [ Links ]
Shaffer, R., Sampaio, T.L., Pinkertorn, J. & Vasconcelos, M.C., 2004. Grapevine root stocks for Oregon vineyards. Oregon State University Extension Service, EM 8882. [ Links ]
Shariat, A. & Assareh, M.H., 2006. Effect of drought stress on pigments, prolin, soluble sugar and growth parameters of four eucalyptus species. J. Constr. Res. 78, 139-148. [ Links ]
Sharma, P. & Dubey, R.S., 2005. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 46, 209-221. [ Links ]
Sivritepe, N., Sivritepe, N., Erturk, U., Yerlikaya, C., Turkan, I., Bor, M. & Ozdemir, F., 2008. Response of the cherry rootstock to water stress induced in vitro. Biol. Plant. 52, 573-576. [ Links ]
Toumi, I., M'sehli, W., Bourgou, S., Jallouli, N., Bensalem-Fnayou, A., Ghorbel, A. & Mliki, A., 2007. Response of undrafted and grafted grapevine cultivars and rootstocks (Vitis sp.) to water stress. J. Int. Sci. Vigne Vin. 41, 85-94. [ Links ]
Tramontini, S., Döring, J., Vitali, M., Ferrandino, A., Stoll, M. & Lovisolo, C., 2014. Soil water-holding capacity mediates hydraulic and hormonal signals of near-isohydric and near-anisohydric Vitis cultivars in potted grapevines. Funct. Plant Biol. 41, 1119-1128. [ Links ]
Tramontini, S., Vitali, M., Centioni, L., Schubert, A. & Lovisolo, C., 2013. Rootstock control of scion response to water stress in grapevine. Environ. Exp. Bot. 93, 20-26. [ Links ]
Wang, F.Z., Wang, Q.B., Kwon, S.Y., Kwak, S.S. & Su, W.A., 2005. Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. J. Plant Physiol. 162, 465-472. [ Links ]
Wang, F., Zeng, B., Sun, Z. & Zhu, C., 2009a. Relationship between proline and Hg2+-induced oxidative stress in tolerant rice mutant. Arch. Environ. Contam. Toxicol. 56(4), 723-731. [ Links ]
Wang, S., Liang, D., Li, C., Hao, Y., Ma, F. & Shu, H., 2012. Influence of drought stress on the cellular ultrastructure and antioxidant system in leaves of drought-tolerant and drought-sensitive apple rootstocks. Plant Physiol. Biochem. 51, 81-89. [ Links ]
Wang, W.B., Kim, Y.H., Lee, H.S., Kim, K.Y., Deng, X.P. & Kwak, S.S., 2009b. Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 47, 570-577. [ Links ]
Submitted for publication: July 2022
Accepted for publication: December 2022
Acknowledgements: Head of the Agricultural and Natural Resources Research Center of Qazvin province and all the experts and technicians of the biological laboratory for their cooperation in conducting the experiments.
* Corresponding author: spiiqv@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}