










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
versão On-line ISSN 2224-7904
versão impressa ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.43 no.2 Stellenbosch 2022
http://dx.doi.org/10.21548/43-2-5329
ARTICLES
The Promise of Using Protoplasts in Grapevine Research
S.S Derman; M.A Vivier*
South African Grape and Wine Research Institute, Department of Viticulture and Oenology, Stellenbosch University, Stellenbosch, South Africa
ABSTRACT
Protoplasting refers to the isolation of protoplasts from plant material. These protoplasts are cells bound only by their plasma membranes following the removal of their cell walls. In grapevine research, protoplasting efforts and successes have been reported from 1985 onwards, but recent research has shown a renewed focus on grapevine-derived protoplasts, particularly due to the possibility of using protoplasts in applications such as gene-editing approaches. Grapevine species are generally considered recalcitrant to manipulation in culture and have also proven recalcitrant with regards to protoplasting and particularly in the regeneration of plants from protoplasts. This review will highlight the important milestones in grapevine protoplasting while outlining the current efforts to establish grapevine protoplasting as a routine technique.
Keywords: Grapevine, Vitis vinifera, protoplast, CRISPR-Cas9
INTRODUCTION
Plant cells have unique characteristics compared to other eukaryotic cells, one of the most evident being their rigid cell walls. When the plant cell wall is absent, the cell is known as a protoplast. Protoplasts are fragile and quite difficult to isolate and handle, but they provide the unique option to reduce a complex and highly organised multicellular organism like a plant, to single cells. With the cell walls removed, the "cells" are released from their multicellular structures, tissues and organs. Protoplasts are useful for a range of scientific applications where a per cell approach is beneficial, and complements studies that are conducted on a tissue, organ, or whole plant level where the aim is to incorporate the intercellular communications and control systems present in a plant body and/or understand the performance of the plant as a whole (Efroni & Birnbaum, 2016; Faraco et al., 2011).
The extreme fragility of protoplasts is limiting as their isolation and handling requires highly skilled operators (Davey et al., 2005). Once isolated, protoplasts can stay alive for hours to days, depending on the culturing conditions. For most applications, protoplasts are used in transient assays, where the half-life and viability of the isolated cells needs to be sufficient for the specific assay (Marx, 2016; Davey et al., 2005). Importantly, plant cells are considered totipotent, meaning that each plant cell (or derived protoplast) theoretically has the capacity to (re)produce a complete plant again (Eeckhaut et al., 2013).
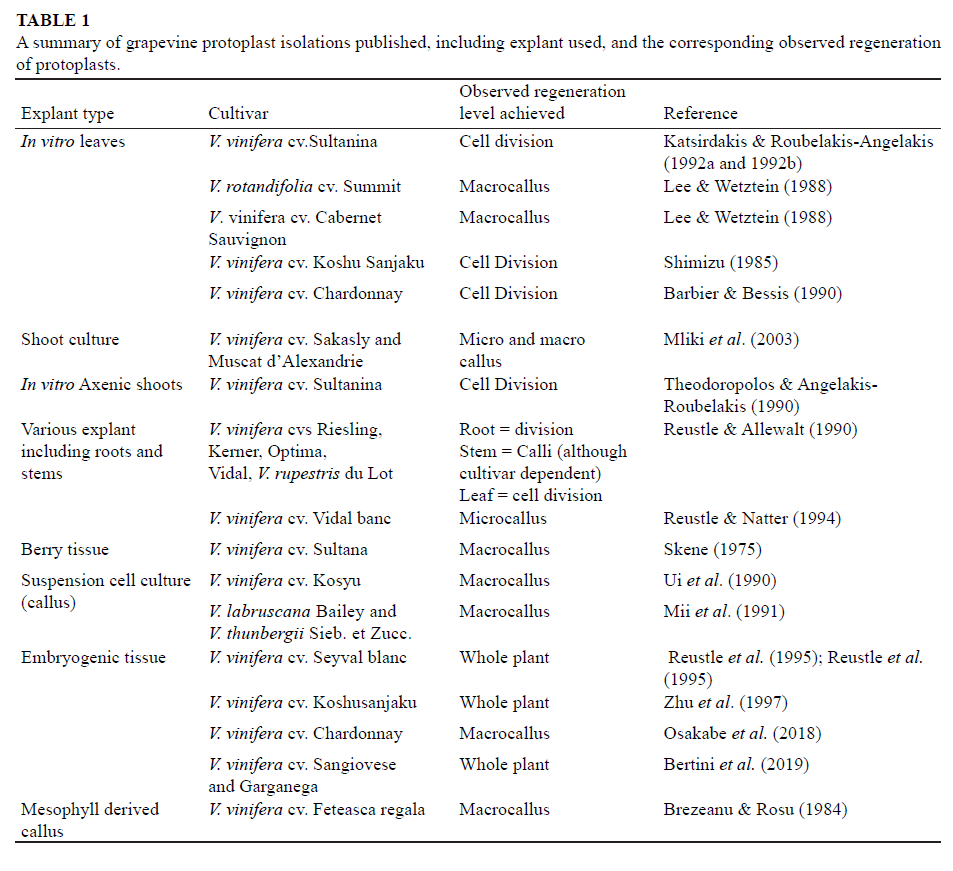
Totipotency is demonstrated in plant tissue culture techniques where plant organs, tissues and cells are manipulated in culture, using specialised culturing conditions and plant hormone combinations. Plant species are generally grouped according to the ease of culturing them in vitro, specifically their responsiveness to clonal propagation, dedifferentiation, embryogenesis and organ/ plantlet regeneration techniques. Those that are "difficult to work with", displaying low efficiencies in the normal in vitro culturing techniques are considered recalcitrant (Benson, 2000). Although protoplast regeneration to form plants have been successfully implemented in a number of species (Eeckhaut et al., 2013), the process itself is universally considered inefficient, irrespective if a species is considered easily cultured or recalcitrant. However, some plant species, such as grapevine, have become notorious for their inability to regenerate from protoplasts (Table 1).
There are many factors that are known to be problematic when isolating, culturing, and using protoplasts for experimental purposes, which are all important to consider when establishing a protoplast-based system, especially for recalcitrant plant species such as grapevine (Papadakis et al., 2001b; Papadakis et al., 2009; Malnoy et al., 2016; Osakabe et al., 2018; Bertini et al., 2019; Saumonneau et al., 2012; Marchive et al., 2013). The scope of this review is to briefly introduce plant protoplasts in general, before summarising the history and current state of the art in terms of grapevine protoplast research. Furthermore, perspectives regarding the usefulness of protoplasts in grapevine research during the adoption of novel gene-editing technologies will be discussed.
The general process of isolating plant protoplasts
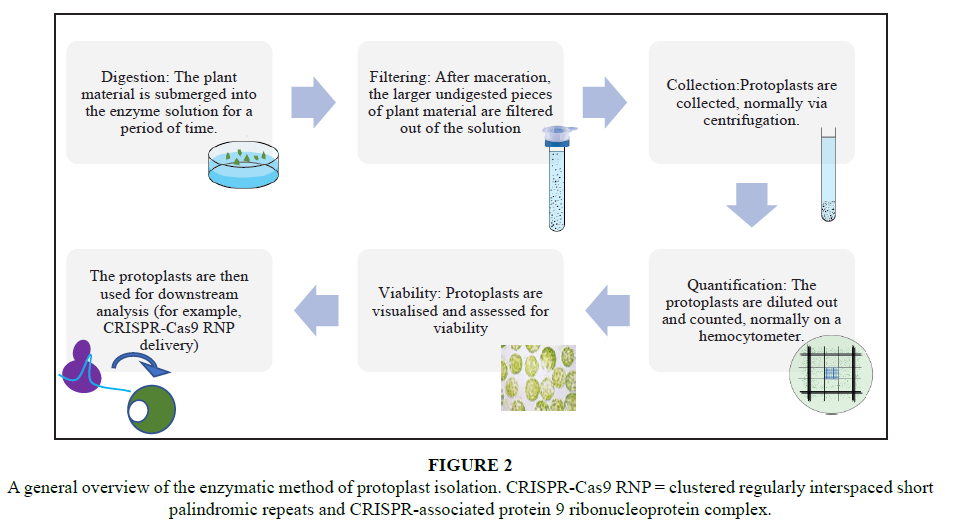
The first plant protoplast to ever be isolated derived from the plasmolysed cells of Stratiotes aloides in 1892 (Von Klercker, 1892). A process now known as the mechanical method of protoplast isolation involves the physical disruption of the cell walls through tissue damage. There are now (gentler) alternatives to remove the plant cell wall, including the commonly used enzymatic method, which relies on the use of enzymes to break down the typical components of the plant cell wall (Figure 1).
The enzymatic method of protoplast isolation involves exposing the desired plant material to an enzymatic solution (Figure 2). After an experimentally optimised digestion period (in which the cell wall is removed), the individual cells are separated from the rest of the digested tissue in order to exclude intact cells or cellular clumps based on size. This is normally carried out using a mesh filter. The cells are then washed multiple times in order to remove any cellular debris present in the isolate. The cells are enumerated under a microscope using a hemocytometer and finally diluted for plating, or analysed to establish the viability of cells and purity of the isolated protoplasts (Figure 2).
Parameters are adjusted based on the nature of the explant used (a pre-treatment of the tissue might be required prior to digestion), the composition of the cell wall (the type of enzymes used and the time allowed for digestion of the cell well), the size of the cells being isolated (size of pores within the mesh filter), the intended purity of the isolate (how many and what type of washing steps are included), as well as the resources available for the assessment of the isolate (can fluorescence microscopy be carried out or not).
The successful removal of the cellulose matrix surrounding the cell allows for these membrane-bound protoplasts to be used for many applications that are otherwise not possible when the cell wall acts as a physical barrier to the external environment. The use of protoplasts in such experiments have generated an encompassing experimental system for cellular biologists, providing an understanding of plant membrane biology, the structure and chemistry of the plasma membrane, the cytoplasmic organelles associated with the plasma membrane, the uptake of macromolecules and membrane transport into the protoplasts, primary and secondary metabolism through idioblasts formation, and organelle isolation such as the nuclei, chloroplast and vacuoles (Cove et al., 1979; Cocking et al., 1985; Eeckhaut et al., 2013). Although these protoplasting applications have been shown to be useful in a range of plant species, a limiting factor remains the phenomenon of recalcitrance in many other plant species, such as grapevine.
Understanding grapevine protoplast recalcitrance
Recalcitrance is often exhibited in in vitro grapevine experiments, specifically in the induction of embryogenesis and in protoplast regeneration. Garcia et al. (2019), Gambino et al. (2010), Guan et al. (2016) and Wójcikowska et al. (2020) describe the regulatory mechanisms involved in the process of embryogenesis and subsequently, the gene regulation or epigenetic modifications that could be responsible for this display of recalcitrance. For example, Gambino et al. (2010) described the importance of the WUSCHEL (WUS)-related homeobox (WOX) genes in coordinating the gene transcription involved in the early phases of embryogenesis in grapevine. Considering that a recalcitrant nature is displayed in grapevine somatic embryogenic calli, the protoplasts isolated therefrom would be expected to also display recalcitrance towards culturing.
Typically, there are three main points within the isolation and culturing procedures that protoplasts can exhibit a "recalcitrant" response, whereby the regeneration of these cells is arrested (Papadakis et al., 2001b). The first time-point in protoplast isolation at which the cell can exhibit a recalcitrant nature is during the isolation procedure itself or directly after isolation (Papadakis et al., 2009). The second recalcitrant stage is at the point at which a viable cell fails to divide. This specific response has received a lot of attention in terms of attempting to understand grapevine protoplast recalcitrance. The focus of these studies was on the possibility of oxidative stress during culturing, causing cell death, or the inability to divide (Roubelakis-Angelakis, 1993). As summarised in Papadakis et al. (2009), many studies up until 2009 suggested that specifically in grapevine tissue, a collapse in the defense mechanism against oxidative stress occurs, which in a normal cell, would be necessary in order to express totipotency.
The last point at which cells can exhibit recalcitrance is in their morphogenic response after cell wall reconstitution and cell division. Polyamine catabolism was looked at as a possible reason for this recalcitrance. Papadakis et al. (2009) observed a beneficial response on the protoplasts in culture when putrescine was added to the media. However, since recent studies have proven that with the correct handling and culturing conditions, it is possible to induce the correct morphogenic response of the cells, it appears that this point of recalcitrance can be overcome with culturing conditions specifically optimised for grapevine (Zhu et al., 1997; Bertini et al., 2019). However, this has only proven to be the case for very few cultivars.
As the application of protoplasts in plant biology again becomes popular, advancements are being made in terms of tracking and analysing their ability to divide and regenerate. Dawson et al. (2022) used a method previously only utilised on mammalian cells (high-throughput microscopic image analysis) to assess the early responses of tobacco leaf-derived protoplasts after isolation and culturing. Dawson et al. (2022) showed how high-throughput automated microscopy and the development of image processing pipelines can allow for quantification of various developmental properties of thousands of protoplasts during the initial days following cultivation by immobilisation in multi-well-plates. As these types of protoplast-monitoring techniques become more accessible, their use could aid in better understanding the recalcitrance displayed by (grapevine) protoplasts during regeneration. Despite the problems highlighted with recalcitrance, the body of work that reports success with grapevine protoplasting is encouraging.
Protoplast isolation from different grapevine source tissues is possible
The recalcitrance of grapevine protoplasts has resulted in the majority of grapevine protoplast studies since 1985 (Table 1 and the references therein) focusing on the optimisation of isolation and culturing procedures, in order to make sure the protoplasts remain viable and regenerative. Examples of the attempted optimisations include altering enzyme combinations, incubation periods and the type of explant used, or on parameters of the culturing steps, such as the method of culturing, the use of different hormone combinations in the media, as well as the effect of different preservatives on the culturing of protoplasts.
All publications that have demonstrated the successful isolation of grapevine protoplasts for the purpose of regenerating and culturing tissues have made use of the enzymatic method of isolation (summarising in Table 1). Although many different enzyme combinations have been used in isolating grapevine protoplasts, most studies have used Macerozyme R-10 (a combination of pectinase, cellulase and hemicellulose), various different cellulases (most commonly from Aspergillus niger or Penicillium funiculosum), cellulysin (Trichoderma viride), dricelase and pectolyase in various concentrations. Currently, the most efficient enzyme combination for isolating protoplasts from embryogenic calli of Vitis spp. (the most regenerative explant) has been the mixed enzymatic method of combining macerozyme, cellulase and pectolyase together in a single incubation (Reustle et al., 1994; Reustle et al., 1995; Zhu et al., 1997; Osakabe et al., 2018; Bertini et al., 2019).
A wide range of grapevine starting material (explants) have been used in the attempts to isolate viable, regenerative protoplasts, each with their own successes and limitations. Some of the most typically used explants for grapevine protoplast isolation are represented in Table 1; somatic embryogenic calli, the most widely used explants for grapevine protoplast isolation, will be discussed further.
As summarised in Table 1, leaves, shoots, roots, berry tissue, suspension cells and mesophyll-derived callus have been used in attempts to isolate regenerative protoplasts from grapevine. To date, the most regenerative form of grapevine tissue to use for protoplast isolation is somatic embryogenic calli (Bertini et al., 2019, Osakabe et al., 2018, Zhu et al., 1997, Reustle et al., 1994, Reustle et al., 1995). The process of somatic embryogenesis, specifically in grapevine is time consuming and requires constant attention from a trained and skilled tissue culturist. With a roughly six-month period from anther/ovary/whole flower into somatic embryogenic calli, this process is not ideal, specifically considering the complete reliance on the availability of immature inflorescence which occurs in a short window period in the vineyard (Cadavid-Labrada et al., 2008). Despite the limitations of the embryogenic process, somatic embryogenic calli is the only explant that has given rise to grapevine protoplasts capable of complete plantlet regeneration (Table 1).
Additional grapevine explants with regenerative capacity for protoplast isolation
Despite all parts of the in vitro vine having been used in attempts to isolate regenerative protoplasts, due to the difficulty of establishing embryogenic tissue, it is still necessary that alternative explants are considered for protoplast isolation. Although the isolation of protoplasts from a specific tissue type may render a good yield, it does not necessarily mean that those protoplasts will possess a good regenerative ability, as was seen when isolating protoplasts from grapevine leaf tissue (Reustle & Aleweldt, 1990). The regenerative capacity of the protoplasts should therefore be an important factor if plant regeneration is the ultimate goal.
In selecting potential explants as protoplast sources, current research hypothesises that epigenetics may play a role since there is a link between the methylation state of the source tissue, and the regenerative ability of the cell derived therefrom. A study conducted by Osorio-Montalvo et al. (2018) profiled different types of plant tissue in terms of their DNA methylation profiles, the corresponding differentiation of that tissue, and their subsequent embryogenic potential (Figure 3). As made clear in Figure 3 tissue at a low state of differentiation (such as meristems, zygotic embryos and anthers) should be looked at when considering alternative explants to isolate protoplasts from, as they possess the same trend in low DNA methylation levels, low status of differentiation, but high embryogenic potential.
The only explant with a favorable epigenetic profile that has been tested in the isolation of grapevine protoplasts is somatic embryogenic calli. Based on regenerative capacities alone, meristems, zygotic embryos and anthers all provide interesting avenues for potentially regenerative protoplasts.
Culturing and regeneration of grapevine protoplasts are far from routine
Although there are many types of culturing methods that can be used, past studies that have focused on optimising the culturing of grapevine protoplasts have shown the predominant use of either embedding the protoplasts in sodium alginate layers or using a disc-culture method. The sodium alginate layered method was commonly used during the initial phases of grapevine protoplast regeneration (Reustle et al., 1994; Reustle et al., 1995; Jardak et al., 1999; Mliki et al., 2003). The embedded disc-culture method with gellan gum, is currently the most successful culturing method used for regeneration of grapevine protoplasts (Bertini et al., 2019; Osakabe et al., 2018; Malnoy et al., 2018; Zhu et al., 1997; Ui et al., 1990).
Briefly, the disc-culture method entails resuspending isolated protoplasts in a low melting point gellan gum containing a carbon source, an osmoticum, all required micro-elements, macro-elements and vitamins. The resuspended protoplasts are pipetted into a small petri-dish. Five discs are pipetted into each petri-dish and allowed to solidify. Once solidified, a liquid media comprising the same components without the gellan-gum is poured over the solid discs. The liquid media is replaced every two weeks, without the osmoticum (Bertini et al., 2019). Recently, Bertini et al. (2019) showed that embryogenesis was readily observed when culturing protoplasts via this method.
A study performed by Zhu et al. (1997) drew a comparison between a simple embedding of protoplasts, the disc-culture method, and the disc-culture method supplemented with liquid media containing activated charcoal. The results were significant in proving that not only was the disc-culturing method 41.2% more successful than the embedding culture method in promoting regeneration, but with the addition of activated charcoal, the number of embryos produced increased by over 400%.
Before 2018, there had only been two studies that reported the regeneration of whole plants from isolated grapevine protoplasts. One example was from the French cultivar V. vinifera cv. Seyval blanc (Reustle et al., 1994; Reustle et al., 1995) and the other from a Japanese cultivar V. vinifera cv. Koshusanjaku (Zhu et al., 1997). Although Reustle et al. (1995) showed that the regeneration of whole grapevine from protoplasts was possible, the efficiency of this protocol was not optimal. The highest regeneration frequency, in terms of protoplasts yielding regenerated whole plants, was 0.0013%.
Interestingly, what was proven in 1995 by Reustle et al. to be the best basal media for culturing grapevine protoplasts, is still used today by those working on whole plant regeneration in grapevine (Bertini et al., 2019). The basal media of Nitsch and Nitsch (1969) resulted in a significantly higher regeneration rate than any other basal media.
A relatively long period elapsed between the Zhu et al. (1997) publication in 1997 and 2019, when it was most recently proved that using two Italian cultivars, Garganega and Sangiovese, regeneration of a whole plant can be obtained from isolated grapevine protoplasts. Bertini et al. (2019) documented the formation of embryos from protoplast-derived callus within three months, with the first signs of cell division occurring within 10 days. They documented that from a single isolation, 87 Sangiovese and 78 Garganega embryos were recovered, but with only 55 Sangiovese and 33 Garganega germinating normally. Bertini et al. (2019) documented that 0.0054% of viable protoplasts regenerated into plants, which is 24% higher than what was reported in 1997 (Zhu et al., 1997). This is a substantial improvement from previous methods.
The use of cell mitotic stimulants in protoplast regeneration
Realising the importance of the optimisation of culturing conditions in cell culture and considering the developing understanding of the epigenetic regulation involved in embryogenesis, the use of mitotic stimulants known to modulate epigenetic profiles in plants should also be considered.
Although there is limited research on chemical compounds that can act as a cell mitotic stimulant in plant cell culture, a specific group of chemicals that are currently receiving attention are DNA methyltransferase (DMT) inhibitors, which have already proven to be beneficial in the induction of embryogenesis in cotton (Li et al., 2019). Understandably, as we gain a better understanding of the epigenetic factors that are responsible for the ability of cells to de-differentiate and re-differentiate in vitro, chemical compounds that have the ability to affect the methylation profiles of these cells will begin to become more popular. One of the most frequently studied DMT inhibitor compounds is zebularine. This specific compound, once incorporated in the cell, can covalently trap DNA methyltransferases and mediate their degradation, leading to passive loss of DNA methylation in the treated cells (Yoo et al., 2005; Stresemann & Lyko, 2008). As shown in Figure 3, a lower DNA methylation profile is seen to correspond with the ability to successfully go through embryogenesis.
The few studies that have tested the application of zebularine in its ability to stimulate cell division in vitro have documented its ability to promote embryogenesis. The plant somatic embryogenic process provokes many epigenetics changes including DNA methylation and histone modification. Li et al. (2019) showed that preventing DNA methylation by applying a chemical treatment (zebularine) to non-embryogenic calli resulted in an increased number of embryo formation, reaching the conclusion that "induced hypomethylation may facilitate higher plant regeneration ability". Although there is limited research conducted in the application of zebularine in plant cell culture thus far, its application in grapevine cultivars that have proven recalcitrant towards somatic embryogenesis may be interesting to pursue, as would the application of zebularine in the regeneration of plant tissue from single cells.
Isolated grapevine protoplasts have been useful in several cell-based assays
The limitation of biotechnological progress in grapevine is made clear when comparing the progress to that achieved in plant biology in general (specifically in model species) (Figure 4).
For example, aseptic tissue culture of the vine was only established 42 years after that of the first plant was cultured in vitro, whereas the first transgenic vine trailed behind the first transgenic tobacco plant by seven years (Bevan et al., 1983; Mullins et al., 1990). Comparing the date of all the major accomplishments in model species against those in grapevine, it is clear that the extension of technological innovation onto grapevine requires years of optimisation before becoming routine. For example, protoplasts were isolated from grapevine tissue 14 years after the first use of the technique to isolate protoplasts from plant tissue (Figure 4). The same trend is seen looking at the recovery of regenerative tissue from protoplasts (Takebe, 1971; Reustle et al., 1994; Reustle et al., 1995).
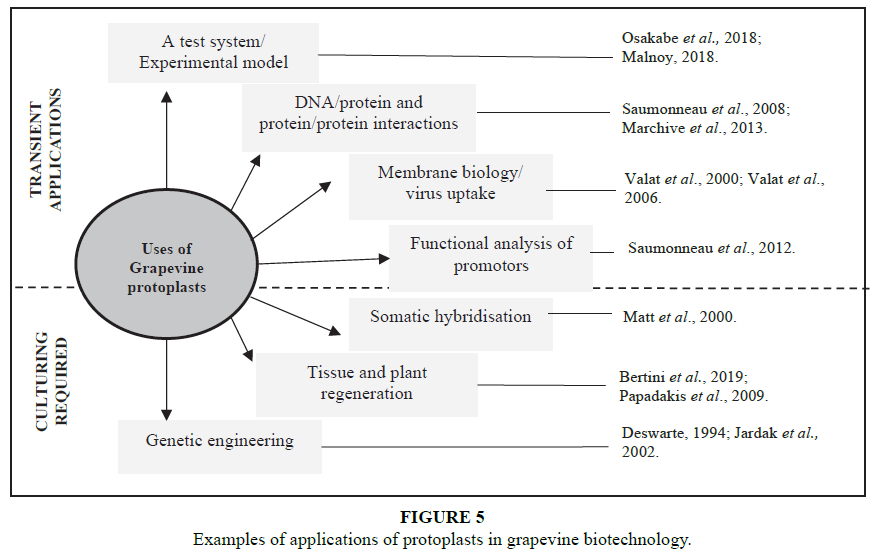
During the last 15 years, there has been focus on the optimisation of in vitro culturing and transformation of grapevine which has made substantial progress in attempt to ensure the study of the grapevine physiology and molecular biology remains on par with other major fruit crops worldwide (Papadakis et al., 2001a, Bouquet et al., 2006; Bouquet et al., 2008; Torregrosa et al., 2015). Among the techniques to benefit from adequate culturing of the vine is that of the protoplast-based biotechnologies. Unfortunately, protoplasts are under-exploited in many recalcitrant plant species (Benson, 2000). In the study of grapevine biology and biotechnology specifically, the use of protoplasts has been limited, with examples of the type of studies and their purpose displayed in Figure 5. Grapevine protoplasts have been used in multiple transient-based studies. In the understanding of grapevine fanleaf virus (GFLV) and grapevine resistance to GFLV, Valat et al. (2000) managed to develop a method in which protoplasts were isolated from the mesophyll of in vitro-grown plants and embryogenic cell suspensions. These protoplasts were then subjected to direct inoculation and electroporation of GFLV. Furthermore, Valat et al. (2006) utilised grapevine protoplasts as a means of assaying transgenic rootstock expressing the coat protein or movement protein gene of GFLV, in order to better understand the molecular mechanisms involved in grapevine resistance to GFLV.
In the study of the biological function of abscisic acid, stress and ripening induced (ASR) proteins in grapevine, Saumonneau et al. (2008) demonstrated multiple polymorphisms of grape ASRs at a cDNA and protein level and carried out their identification as chromosomal nonhistone proteins. This was achieved by using a yeast two-hybrid screening approach in conjunction with a bimolecular fluorescence complementation in grapevine protoplasts. This study was followed by the use of protoplasts in a dual luciferase system by Saumonneau et al. (2012), in which the transcriptional control of the expression of ASR genes in relation to glucose and ABA signaling was looked at. In 2000, Matt et al. (2000) developed an electrofusion protocol for the somatic hybridisation of grapevine protoplasts. Other studies that have also made use of the transient applications of grapevine protoplasts showcased the ability for us to better understand resistance to fungal pathogens such as downy mildew. An example of this was demonstrated by Marchive et al. (2013), who proved the ability of the transcription factor VvWRKY1 to trans-activate the promotors of genes involved in the jasmonic acid signaling pathway, and as a result, showed enhanced resistance of transgenic plants to downy mildew.
Most recently, Hu et al. (2021) created a working model for the role of VpCDPK9 and VpCDPK13 in the regulation of powdery mildew resistance in grapevine. The ethodology in this study showcased the use of mesophyll-derived grapevine protoplasts in analysing protein phosphorylation in-vivo via Phos-tag SDS-PAGE and blotting, and in studying Co-immunoprecipitation (co-IP assay) via transient transformation of the protoplasts. None of these above-mentioned examples relied on successful culturing of grapevine protoplasts. The uses of grapevine protoplasts that require further culturing to determine the success of the experiment are discussed in the next sections.
Grapevine protoplasting combined with gene editing: the new frontier in grapevine improvement?
The previously mentioned applications of protoplasts are important in the study of all plant species, but understandably, crop plants are important in multiple aspects other than just that of research, with considerable commercial and financial stress being placed on crop-improvement-based research. Despite the limited application seen in Figure 5, grapevine protoplast research has recently seen a revival of interest, as new methods for genetic engineering, rely heavily on the use of protoplasts as sources materials.
As made clear by Vivier & Pretorius (2002) and Gray et al. (2015), there are many limitations arising from the lifecycle of the vine that do not permit the ease of application of conventional breeding techniques as with non-recalcitrant plant species. In 2020, technologies that bypass many of these limitations are available, most of which rely on a precision-breeding approach. This sees the genetic improvement of a plant without relying on conventional breeding, but rather a method of transferring only desirable genetic components among sexually compatible relatives without the genetic disruption imposed by meiosis (Gray et al., 2015). This general quest to alleviate the complete reliance on conventional breeding has led to a drastic chang in the way biotechnological tools are used to better understand the functioning of plants.
Currently, the most advanced biotechnological tool available for genetic engineering is the Clustered regularly interspaced short palindromic repeat regions - CRISPR-associated protein 9 (CRISPR-Cas9) technology (Samanta et al., 2016). As in any other crop plant, there are specific benefits of using this tool in grapevine, which have been addressed by many proponents for grapevine biotechnology (Nakajima et al., 2015; Ren et al., 2016; Wang et al., 2018). This method entails the direct delivery of the CRISPR-Cas9 components in a vector-independent manner into protoplasts, with subsequent regeneration from these protoplasts rendering gene-edited, non-chimeric plants. The tissue recovered from this means of genome editing will be free from vector backbone and any form of Agrobacterium, having the potential to bypass regulatory concerns imposed on traditional genetically modified plants (Sprink et al., 2016; Dalla Costa et al., 2018). However, a major prerequisite in applying this technology to any plant is to have a system to successfully perform protoplast isolation, transformation and whole plant regeneration from the genetically transformed protoplast.
A study in 2013 documented the first use of the CRISPR-Cas9 system in both a model plant (Arabidopsis thaliana) and an important crop plant (Oryza Sativa) (Feng et al., 2013). Within 6 years from this information being released, it was demonstrated that the implementation of the CRISPR-Cas9 tool in grapevine was possible, and in 2022 there are now numerous publications documenting the tools' use in grapevine (Nakajima et al., 2015, Wang et al., 2018, Ren et al., 2016, Malnoy et al., 2016, Osakabe et al., 2018,
Ren et al., 2019, Li et al., 2020, Ren et al., 2020, Sunita et al., 2020, Wan et al., 2020., Ren et al., 2022). In terms of Agrobacterium vector-based delivery, for example, Ren et al. (2016) showed that by transforming grapevine embryogenic calli, the technology is capable of inducing site-specific mutations no different to the application in model plants, with edited somatic embryos being recovered successfully. This method of vector-based delivery of the CRISPR-Cas9 system, and subsequent gene-editing is still relatively new, with the first eukaryotic cell to be successfully edited in 2013 (Cong et al., 2013). A review by Ren et al. (2022) precisely documents all applications of CRISPR-Cas9 mediated genome editing in grapevine, the future prospects of using this tool in grapevine research as well as the challenges facing the adoption of this tool in grapevine biology.
Both Osakabe et al. (2018) and Bertini et al. (2019) have subsequently proven that transfection of isolated protoplasts is possible, followed by the culturing of these protoplasts by the disc-culture method. Bertini et al. (2019) showed an optimised method for PEG-mediated transfection of protoplasts with a GFP reporter gene, whilst Osakabe et al. (2018) showed the delivery of the Cas9 protein and the sgRNA directly into the protoplasts. However, no regeneration was observed past microcalli (Osakabe et al., 2018). A Cas9 cleavage assay showed that the Cas9 protein was functional within the protoplast, cleaving the correct target gene, confirming that the Cas9 and the sgRNA was successfully delivered into the protoplasts.
In implementing these technologies for the purpose of genome editing, a number of aspects remain problematic, such as the off-target mutations induced by the CRISPR-Cas9 system, the limited transformation technologies vailable for the introduction of plant cells proven recalcitrant to transformation, the low efficiency of multiplexed editing and the ethical dilemma of the (current) classification of genetically edited plants as 'Genetically Modified Organisms' (Mao et al., 2019) in many countries.
Looking at what has already been achieved in terms of stable transformation of the CRISPR-Cas9 system in grapevine, if protoplast isolation and regeneration becomes routine, there is major potential for ribonucleoprotein (RNP)-based transient gene-editing (direct introduction of sgRNA and Cas9 in protein form) in the near future for grapevine.
CONCLUSIONS AND PERSPECTIVES
The advancements made in applying cutting-edge biotechnological tools in grapevine are currently limited by the recalcitrance of the Vitis genus. Looking at the unique limitations facing the field of grapevine crop improvement, the efforts put towards the development of new biotechnological tools and techniques are justified (Dalla Costa et al., 2018). Techniques such as protoplast isolation and culture in grapevine have been carried out for almost 50 years now, with continued optimisation still on-going today.
The CRISPR-Cas9 technology has proven to be an easily accessible, easy to use, highly efficient precision breeding tool, with applications now extending into DNA free gene editing. However, a major prerequisite in applying this technology in grapevine is to have a system for successful protoplast isolation, transformation and whole plant regeneration from genetically transformed protoplasts. It is therefore required that grapevine biotechnologists re-visit previously abandoned in vitro techniques deemed inefficient, such as that of isolating viable protoplasts from grapevine tissue that can be subjected to genetic transformation, as well as developing a means of successfully culturing these protoplasts back into a whole plant. Looking at what has already been achieved in terms of stable transformation of the CRISPR-Cas9 system in grapevine, if protoplast isolation and regeneration were to become a routine method of grapevine culture, there is promising potential for RNP-based gene-editing in the near future for grapevine.
As modern precision breeding techniques for crop improvement become more and more advanced, grapevine, along with many other recalcitrant plant species, are confined in their progress based on the in vitro techniques to which the plant is responsive. Considering the complexities of such a technique, continued optimisations are expected. It is impossible to fully explore the numerous variables that need to be considered during protoplast isolation, culture and further applications, and for this reason emphasis is placed on factors important to establishing a regeneration platform.
As the field of protoplast-based biotechnology in grapevine starts to become increasingly more popular again, there are many avenues that require optimisation and clarification, ranging from standard in vitro culture optimisation, the possibility of using alternative explants for isolation, isolation parameters, the application of cell stimulants (methylation inhibitors), transfection conditions, and the recovery of transformed tissue without selection.
LITERATURE CITED
Barbier, M. & Bessis, R. 1990. Isolation and culture of grapevine cultivar Chardonnay leaf protoplasts. Euphytic, 47: 39-44. [ Links ]
Benson, E. 2000. In vitro Plant Recalcitrance: An Introduction. In Vitro Cellular & Developmental Biology. Plant, 36 (3): 141-148. [ Links ]
Bertini, E., Tornielli, G.B., Pezzotti, M. & Zenoni, S. 2019. Regeneration of plants from embryogenic callus-derived protoplasts of Garganega and Sangiovese grapevine (Vitis vinifera L.) cultivars. Plant Cell Tiss Organ Cult, 138: 239-246. [ Links ]
Bevan, M., Flavell, R. & Chilton, M.D. 1983. A chimaeric antibiotic resistance gene as a selectable marker for plant cell transformation. Nature, 304(5922): 184-187 [ Links ]
Bouquet, A., Torregrosa, L., Iocco, P. & Thomas, M.R. 2006. Grapevine (Vitis vinifera L.), in Wang, K. Agrobacterium Protocols Volume 2. Methods in Molecular Biology, NJ: Humana Press, Totowa. 273-285. [ Links ]
Bouquet, A., Torregrosa, L., Iocco, P. & Thomas, M.R. 2008. Grapes, in Kole, C & Hall, T. (eds.). Compendium of Transgenic Crop Plants: Transgenic Temperate Fruits and Nuts. Oxford: Blackwell Publishing. 189-232. [ Links ]
Brezeanu, A. & Rosu, A. 1984. Isolation and culture of cell protoplasts from mesophyll callus of Vitis vinifera L. Rev. Roum. Biol. Biol. Veg., 29: 33-37. [ Links ]
Cadavid-Labrada, A. 2008. Somatic embryogenesis and efficient regeneration of Vitis vinifera L.'Carménere'plants. Vitis 47 (1), 73-74. [ Links ]
Chaib J., Torregrosa L., Mackenzie D., Corena P., Bouquet A. and Thomas M.R.. 2010. The grapemicrovine - a model system for rapid forward and reverse genetics of grapevines. Plant J., 62:1083-1092. [ Links ]
Cocking, E.C. 1960. A method for the isolation of plant protoplasts and vacuoles. Nature (Land.), 187: 962-963. [ Links ]
Cocking, E.C. 1985. Somatic hybridization: Implications for agriculture, in Zaitlin, M., Day, P., Hollaender, A. (eds.) Biotechnology in Plant Science: Relevance to Agriculture in the Eighties. Orlando: Academic Press. 101113. [ Links ]
Cong, L., Ran, F.A., Cox, D., Lin, S., Barretto, R., Habib, N., Hsu, P.D., Wu, X., Jiang, W., Marraffini, L.A. & Zhang, F. 2013. Multiplex genome engineering using CRISPR/Cas systems. Science, 339:819-823. [ Links ]
Cove, D. 1979. The uses of isolated protoplasts in plant genetics. Heredity 43, 295-314. [ Links ]
Davey, M. R., Anthony, P., Power, J. B. & Lowe, K. C. 2005. Plant protoplasts: status and biotechnological perspectives. Biotechnol. Adv., 23(2), 131-171. [ Links ]
Dalla Costa, L., Malnoy, M. & Gribaudo, I. 2017. Breeding next generation tree fruits: technical and legal challenges. Hortic. Res., 4. [ Links ]
Dawson, J., Pandey, S., Yu, Q., Schaub, P., Wüst, F., Moradi, A.B., Dovzhenko, O., Palme, K. & Welsch, R. 2022. Determination of protoplast growth properties using quantitative single-cell tracking analysis. Plant methods, 78(1):1-15. [ Links ]
Deswarte, C., Rouquier, P., Roustan, J.P., Dargent, R. & Fallot, J. 1994. Ultrastructural changes produced in plantlet leaves and protoplasts of Vitis vinifera cv Cabernet Sauvignon by eutypine, a toxin from Eutypa lata. Vitis, 33: 185-188. [ Links ]
Eeckhaut, T., Lakshmanan, P.S., Deryckere, D. Von Bockstaele, E. & Van Huylenbroeck, J. 2013. Progress in plant protoplast research. Planta 238: 991-1003. [ Links ]
Efroni, I., Birnbaum, K.D. 2016. The potential of single-cell profiling in plants. Genome Biol., 17: 65. [ Links ]
Faraco, M., Di Sansebastiano, G.P., Spelt, K., Koes, R. & Quattrocchio, F. 2011. One Protoplast Is Not the Other! PlantPhysiology,156 (2): 474-478. [ Links ]
Feng, Z., Zhang, B., Ding, W., Liu, X., Yang, D. L., Wei, P., Cao, F., Zhu, S., Zhang, F., Mao, Y. & Zhu, J. K. 2013. Efficient genome editing in plants using a CRISPR/Cas system. Cell research, 23(10):1229-1232. [ Links ]
Gambino, G., Minuto, M., Boccacci, P., Perrone, I., Vallania, R. & Gribau-do, I. 2010. Characterization of expression dynamics of WOX homeodo-main transcription factors during somatic embryogenesis in Vitis vinifera. Journal of Experimental Botany, 62(3), 1089-1101. [ Links ]
Garcia, C., de Almeida, A. A. F., Costa, M., Britto, D., Valle, R., Royaert, S. & Marelli, J. P. 2019. Abnormalities in somatic embryogenesis caused by 2, 4-D: an overview. Plant Cell, Tissue and Organ Culture (PCTOC), 137(2): 193-212. [ Links ]
Gray, D.J., Li, Z.T., Grant, T.N.L., Dean, D.A., Trigiano, R.N. & Dhekney, S. A. 2015. The Application of Precision Breeding (PB) for Crop Improvement is Fully Consistent with the Plant Life Cycle: The Utility of PB for Grapevine. Precision Breeding ACTA Horticulturae, 1115: 49-56. [ Links ]
Guan, Y., Li, S.G, Fan, X. F. & Su, Z.H. 2016. Application of Somatic Embryogenesis in Woody Plants. Front. Plant Sci. 7(938). [ Links ]
Hu, Y., Cheng, Y., Yu, X., Liu, J., Yang, L., Gao, Y., Ke, G., Zhou, M., Mu, B., Xiao, S. & Wang, Y. 2021. Overexpression of two CDPKs from wild Chinese grapevine enhances powdery mildew resistance in Vitis vinifera and Arabidopsis. New Phytologist, 230(5): 2029-2046. [ Links ]
Jaillon, O, et al. 2007. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature vol. 449: 463-7. [ Links ]
Jardak, R. 1999. Utilisation des protoplastes pour la transformation génétique de la vigne (Vitis vinifera L.). Diplome d'Etudes Approfondies. Université de Tunis: Departement des Sciences Biologique. [ Links ]
Katsirdakis, K.C. & Roubelakis-Angelakis, K.A. 1992a. A modified culture medium and culture conditions increase viability and cell wall synthesis in grapevine (Vitis vinifera L. cv Sultanina) leaf protoplasts. Plant Cell Tiss. Org. Cult. 28: 255-260. [ Links ]
Katsirdakis, K.C. & Roubelakis-Angelakis, K.A. 1992b. Ultrastructural and biochemical aspects of cell wall regeneration in recalcitrant and regenerating leaf protoplasts. In vitro Cell Dev. Race-specific elicitors of Cladosporium fulvum promote translocation of cytosolic components of NADPH oxidase to the plasma membrane of tomato cells. Plant Cell Biol., 28 (9): 90-96. [ Links ]
Kazusa DNA Research Institute., The Cold Spring Harbor and Washington University Sequencing Consortium., The European Union Arabidopsis Genome Sequencing Consortium. et al. 2000. Sequence and analysis of chromosome 5 of the plant Arabidopsis thaliana . Nature 408: 823-826. [ Links ]
Lee, N. & Wetzstein H. Y. 1988. Protoplastisolation And Callus Production From Leaves Of Tissue-Cultured Vitis Spp. Plant Cell Rep. 7: 531-534. [ Links ]
Li, J., Wang, M., Li, Y., Zhang, Q., Lindsey, K., Daniell, H., Jin, S. & Zhang, X. 2019. Multi-omics analyses reveal epigenomics basis for cotton somatic embryogenesis through successive regeneration acclimation process. Plant Biotechnology Journal, 17: 435-450 [ Links ]
Li, M.Y., Jiao, Y.T., Wang, Y.T., Zhang, N., Wang, B.B., Liu, R.Q., Yin, X., Xu, Y. and Liu, G.T. 2020. CRISPR/Cas9-mediated VvPR4b editing decreases downy mildew resistance in grapevine (Vitis vinifera L.). Horticulture research, 7. [ Links ]
Malnoy, M., Viola, R., Jung, M., Koo, O., Kim, S., Kim, J., Velasco, R., & Kanchiswamy, C.N. 2016. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci., 7:1904. [ Links ]
Mao, Y., Botella, J., Liu, Y & Zhu, J. 2019. Gene editing in plants: progress and challenges, National Science Review, 6(3): 421-437. [ Links ]
Marchive, C., Leon, C., Kappel, C., Coutos-Thevenot, P., Corio-Costet, M.F., Delrot, S. & Lauvergeat, V. 2013. Over-expression of VvWRKYl in grapevines induces expression of jasmonic acid pathway-related genes and confers higher tolerance to the downy mildew. PLoS ONE, 8. [ Links ]
Marx, V. 2016. Plants: a tool box of cell-based assays. Nature methods, 13(7), 551-554. [ Links ]
Matt, A., Reustle, G.M. & Blaich, R. 2000. Somatic hybridization of grapevine protoplasts. Acta Hortic., 538: 411-414. [ Links ]
Mii, M., Zou, Y.M., Sugiyama, T., Yanagihara, S. & Lizuka, M. 1991. High-frequency callus formation from protoplasts of Vitis labruscana Bailey and Vitis thunbergii Sieb. et Zucc. by embedding in gellan gum. Scientia Hort., 46: 253-260. [ Links ]
Mliki, A., Jardak., R., Reustle, G. & Ghorbel, A. 2003. Isolation and culture of leaf protoplasts from Tunisian grapes. Journal International des Sciences de la Vigne et du Vin. 37(3): 145-153. [ Links ]
Morel G. 1944. Sur le développement des tissus devigne cultives in vitro. C.R. Soc. Biol. 138, 62. [ Links ]
Mullins M.G. and Srinivasan C. 1976. Somaticembryos and plantlets from an ancient clone of the grapevine (cv Cabernet-Sauvignon) by apomixis invitro. J. Exp. Bot. 27, 1022-1030. [ Links ]
Mullins, M.G., Tang, F.C.A. & Facciotti, D. 1990. Agrobacterium -mediated genetic transformation of grapevines: transgenic plants of Vitis rupestris Scheele and buds of Vitis vinifera L. Bio/Technology, 8: 1041-1045. [ Links ]
Nakajima, I., Ban, Y., Azuma, A., Onoue, N., Moriguchi, T., Yamamoto, T., Toki, S. & Endo, M. 2017. CRISPR/Cas9-mediated targeted mutagenesis in grape. PLoS One, 12 (95). [ Links ]
Nitsch, J.P. & Nitsch, C. 1969. Haploid plants from pollen grains. Science, 163: 85-87. [ Links ]
Osakabe, Y., Liang, Z., Ren, C., Nishitani, C., Osakabe, K., Wada, M., Komori, S., Malnoy, M., Velasco, R., Poli, M., Jung, M., Koo, O., Viola, R. & Nagamangala Kanchiswamy, C. 2018. CRISPR-Cas9-mediated genome editing in apple and grapevine. Nat Protoc., 13(12):2844-2863. [ Links ]
Osorio-Montalvo, P., Sáenz-Carbonell, L. & De-la Peña, C. 2018. 5-Azacytidine: A Promoter of Epigenetic Changes in the Quest to Improve Plant Somatic Embryogenesis. International Journal of Molecular Sciences, 19(10): 3182. [ Links ]
Papadakis, A., Fontes, N., Gerós, H. & Roubelakis-Angelakis, K. 2009. Progress in Grapevine protoplast Technology, in Roubelakis-Angelakis K.A. (eds.). Grapevine Molecular Physiology & Biotechnology. Dordrecht: Springer. [ Links ]
Papadakis, A., Reustle, G. & Roubelakis-Angelakis, K. 2001a. Protoplast Technology in Grapevine, in Roubelakis-Angelakis, K. (eds.). Molecular Biology & Biotechnology of the Grapevine. Dordrecht: Springer, 53-392. [ Links ]
Papadakis, A.K., Siminis, C.I., & Roubelakis - Angelakis, K.A. 2001b. Reduced activity of antioxidant machinery is correlated with suppression of totipotency in plant protoplasts. Plant Physiol., 126 (1) 434-444. [ Links ]
Ren, C., Liu, X., Zhang, Z., Wang, Y., Duan, W., Li, S. & Liang, Z. 2016. CRISPR/Cas9- mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep., 6, 32289. [ Links ]
Ren, F, Ren, C, Zhang, Z, Duan, W, Lecourieux D, Li, S., Liang, Z. 2019. Efficiency optimization of CRISPR/Cas9-mediated targeted mutagenesis in grape. Frontiers in Plant Science, 10:612. [ Links ]
Ren, C., Guo, Y., Kong, J., Lecourieux, F., Dai, Z., Li, S. and Liang, Z. 2020. Knockout of VvCCD8 gene in grapevine affects shoot branching. BMC plant biology, 20(1):1-8. [ Links ]
Ren, C., Lin, Y. and Liang, Z. 2022. CRISPR/Cas genome editing in grapevine: recent advances, challenges and future prospects. Fruit Research, 2(1):1-9. [ Links ]
Reustle, G. & Alleweldt, G. 1990. Isolation and culture of grapevine protoplasts. Vitis, 29: 423-431. [ Links ]
Reustle, G. & Natter, I. 1994. Effect of polyvinylpyrrolidone and activated charcoal on formation of microcallus from grapevine protoplasts (Vitis spp). Vitis, 33: 117-122. [ Links ]
Reustle, G., Hurst, M. & Alleweldt, G. 1994. Regeneration of grapevine (Vitis sp.) protoplasts. Vitis, 33, 173-174. [ Links ]
Reustle, G., Hurst, M., & Alleweldt, G. 1995. Plant regeneration of grapevine (Vitis sp.) protoplasts isolated from embryogenic tissue. Plant Cell Rep., 15: 238-241. [ Links ]
Roubelakis-Angelakis, K.A. 1993. An assessment of possible factors contributing to recalcitrance of plant protoplasts, in Morphogenesis in Plants: Molecular Approaches. New York: Plenum. 201-220. [ Links ]
Samanta, M.K., Dey, A. & Gayen, S. 2016. CRISPR/Cas9: an advanced tool for editing plant genomes. Transgenic Res 25: 561-573. [ Links ]
Saumonneau, A., Agasse, A., Bidoyen, M.T., Lallemand, M., Cantereau, A., Medici, A., Laloi, M. & Atanassova, R. 2008. Interaction of grape ASR proteins with a DREB transcription factor in the nucleus. FEBS Lett., 582: 3281-3287. [ Links ]
Saumonneau, A., Laloi, M., Lallemand, M., Rabot, A., & Atanassova, R. 2012. Dissection of the transcriptional regulation of grape ASR and response to glucose and abscisic acid. Journal of experimental botany, 63(3): 14951510 [ Links ]
Shimizu, J.I. 1985. Cell regeneration and division of grape mesophyll protoplasts. J. Plant Physiol., 119: 405-418. [ Links ]
Skene, K.G.M. 1974. Culture of protoplasts from grapevine pericarp callus. Aust. J. Plant Physiol, 1: 371-376. [ Links ]
Skene, K.G.M. 1975. Production of callus from protoplasts of cultured grape pericarp. Vitis, 14: 177-180. [ Links ]
Sprink, T., Eriksson, D., Schiemann, J. & Hartung, F. 2016. Regulatory hurdles for genome editing: process- vs. product-based approaches in different regulatory contexts. Plant Cell Reports. 35 (7): 1493-1506. [ Links ]
Steward, F.C., Mapes, M.O., and Smlth, J. 1958. Growth and organized development of cultured cells. I. Growth and division of freely suspended cells. Am. J. Bot. 45: 693-703. [ Links ]
Stresemann, C. & Lyko, F. 2008. Modes of action of the DNA methyltransferase inhibitors azacytidine and decitabine. International journal of cancer, 123(1): 8-13. [ Links ]
Sunitha, S. & Rock, C.D., 2020. CRISPR/Cas9-mediated targeted mutagenesis of TAS4 and MYBA7 loci in grapevine rootstock 10114. Transgenic research, 29(3): 355-367. [ Links ]
Takebe, I., Labib, G. & Melchers, G. 1971. Regeneration of whole plants from isolated mesophyll protoplasts of tobacco. Naturwissenschaften, 58: 318-320. [ Links ]
Theodoropoulos, P.A. and K.A. Roubelakis-Angelakis. 1990. Progress in leaf protoplast isolation and culture from virus-free axenic shoot cultures of Fins vinifera L. Plant Cell Tiss. Org. Cult, 20: 15-23. [ Links ]
Torregrosa, L., Vialet, S., Adivèze, A., Iocco-Corena, P. & Thomas, M.R. 2015. Grapevine (Vitis vinifera L.). Agrobacterium Protocols Methods in Molecular Biology, 2: 1224. [ Links ]
Ui, S., Suzuki, M., Kubota, S., Masuda, H., Muraki, H., Yamakawa, Y. & Sato, T. 1990. Cooperative effect of activated charcoal and gellun gum on grape protoplast culture. Agric. Biol. Chem., 54: 207-209. [ Links ]
Valat, L., Fucks, M. & Burrus, M. 2006. Transgenic grapevine rootstock clones expressing the coat protein or movement protein genes of Grapevine fanleaf virus: Characterization and reaction to virus infection upon protoplast electroporation. Plant Sci, 170:739-747. [ Links ]
Valat, L., Toutain, S., Courtois, N., Gaire, F., Decout, E., Pinck, L., Mauro, M-C. & Burrus, M. 2000. GFLV replication in electroporated grapevine protoplasts. Plant Sci, 155:203-212 [ Links ]
Vivier, M. A. & Pretorius, L. S. 2000. Genetic Improvement of Grapevine: Tailoring Grape Varieties for The Third Millennium - A Review. S. Afr. J. Enol. Vitic., 21(1): 5-26. [ Links ]
Von Klercker, J. 1892. A method for isolating living protoplasts. Ofvers Vetensk. Akad. Forh. Stockholm. 49: 463. [ Links ]
Yoo, C., Cheng, J. & Jones, P.A. 2005. Zebularine: A new drug for epigenetic therapy. Biochemical Society transactions. 32: 910-2. [ Links ]
Wan, D.Y., Guo, Y., Cheng, Y., Hu, Y., Xiao, S., Wang, Y. & Wen, Y.Q. 2020. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Horticulture research, 7. [ Links ]
Wójcikowska, B., Wójcik, A. M. & Gaj, M. D. 2020. Epigenetic Regulation of Auxin-Induced Somatic Embryogenesis in Plants. International Journal of Molecular Sciences, 21(7): 2307. [ Links ]
Woo, J. W., Kim, J., Kwon, S. I., Corvalan, C., Cho, S. W., Kim, H., et al. 2015. DNA-free Genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotech. 33: 1162-1164 [ Links ]
Zhu, Y.M., Hoshino, Y., Nakano, M., Takahashi, E. & Mii, M. 1997. Highly efficient system of plant regeneration from protoplasts of grapevine (Vitis vinifera L.) through somatic embryogenesis by using embryogenic callus cultures and activated charcoal. Plant Science. 123(1): 151-157. [ Links ]
Submitted for publication: July 2022
Accepted for publication: October 2022
* Corresponding author: E-mail address: mav@sun.ac.za
Acknowledgements: The National Research Foundation, The Wine Industry Network for Expertise and Technology (Winetech) and Stellenbosch University for their financial support


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}