










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
versão On-line ISSN 2224-7904
versão impressa ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.43 no.2 Stellenbosch 2022
http://dx.doi.org/10.21548/43-2-4897
ARTICLES
Potential of Entomopathogenic Nematodes for the Control of Plangia gramínea (Orthoptera: Tettigoniidae) under Laboratory Conditions
F. du Preez*; A. P. Malan; P. Addison
Department of Conservation Ecology and Entomology, Faculty of AgriSciences, Stellenbosch University, Private Bag X1, Matieland 7602, Stellenbosch, South Africa
ABSTRACT
Plangia gramínea, locally known as a katydid or "krompokkel", is a minor pest of vineyards in the Western Cape province of South Africa. It feeds on leaves, and sporadically on the skin of grapevine berries. Under natural conditions, katydids are not of much agricultural importance, but pest outbreaks during favourable conditions can result in significant foliar damage. Observations indicate an increase in katydid abundance and damage intensity in recent years. Currently, no agrochemicals are registered for the control of this species, and its present natural enemies are unlikely to provide sufficient control without augmentation. In this study, 12 entomopathogenic nematode (EPN) species were evaluated against the nymphs of Plangia graminea in laboratory bioassays, and mortality by infection was investigated. Seven locally occurring nematode species achieved significant mortality, with H. zealandica, H. indica, S. jeffreyense and S. yirgalemense being found to perform the best (> 90% mortality).
Key words: Biological control, EPN, Heterorhabditis, integrated pest management, katydid, Steinernema
INTRODUCTION
Plangia graminea (Serville) (Orthoptera: Tettigoniidae) belongs to a group of insects generally known as katydids or bush crickets, ("krompokkels" in Afrikaans) and is a minor and sporadic pest of vineyards in South Africa. It primarily feeds on the foliage of grapevines (Fig. 1). Young nymphs (4 mm - 7 mm) seem to mimic black beetles (apart from for their long antennae) to evade predation within the foliage, while the adults camouflage well within the leafy canopy, which makes their monitoring, especially in the case of the adults, difficult (Du Preez, 2019). Similar to most katydids of the Phaneropterinae subfamily, P. graminea has only one generation per year (Bailey & Rentz, 1990), with soil not being utilised for any of its life stages (Du Preez, 2019).
Observations by Allsopp (2012) have indicated an increase in the extent of katydid abundance and damage intensity in the Cape Winelands district, possibly due to changes in agrochemical use or weather conditions. Katydids seem to occur in most of the Western Cape province, especially in the Cape Winelands region (Du Preez, 2019), but, due to their inconspicuous nature, their distribution is likely to have been underestimated.
In grapevine, the eggs of P. graminea were observed to start hatching in early spring (September), during the onset of bud break. Nymphs were seen to undergo several moults over the course of three months, with the population peaking between late October and November. Adults started to emerge from late November, with at least half of the katydid population maturing into adults by December (Doubell, 2017; Du Preez, 2019). The adults were observed to be highly mobile, with the ability to disperse to, and to lay eggs in, adjacent vineyards or vegetation. Females of P. graminea tend to lay their eggs under the bark of grapevine, in contrast to other Phaneropterinae, which usually lay their eggs in the epidermal layer of leaves (Picker et al., 2004) where they overwinter until spring of the following season (Doubell, 2017; Du Preez, 2019). Pest outbreaks in vineyards can cause significant foliar destruction, which not only degrades the vigour and growth of vines, but also affects grape berry health and quality. The outbreaks seem concentrated around certain hotspots within the Cape Winelands region, with the causative factors still unknown (Du Preez, 2019).
Biological control agents include generalist predators such as birds, lizards, spiders (growers, pers. comm.) and parasitoid wasps (Doubell, 2017). The entomopathogenic fungus (EPF) Metarhizium anisopliae (Metchnikoff) Sorokin (Hypocreales: Clavicipitaceae) was identified from a katydid cadaver (PPRI 12353) by Doubell (2017), and Beauveria bassiana (Bals.-Criv.) Vuill. (Hypocreales: Cordycipitaceae) is available commercially as Bio-Insek® for the control of "krompokkel", mealybug and snout beetle (Agro-Organics, 2010). In South Africa, the commercial product Green Muscle® (L6198), with the active ingredient Metarhizium anisopliae var. acridum, is registered for use against locusts and grasshoppers (Hatting et al., 2018).
Entomopathogenic nematodes (EPNs) are insect-parasitic roundworms that naturally occur in soil worldwide. Nematodes of the families Heterorhabditidae and Steinernematidae (Rhabditida) are associated with the symbiotic bacteria Photorhabdus and Xenorhabdus (Enterobacteriales: Enterobacteriaceae), respectively (Stock & Goodrich-Blair, 2012), which are pathogenic to insects (Kaya & Gaugler, 1993). Through inundative releases, the nematodes can be utilised as biological control agents against a wide range of insect species, causing significant mortality within 48 h by finding insects in cryptic habitats and having the ability to persist post application (Dillman & Sternberg, 2012).
Local research evaluated above-ground applications of certain local EPN isolates against the adults of the banded fruit weevil, Phlyctinus callosus (Schönherr) (Coleoptera: Curculionidae) (Ferreira & Malan, 2014; Dlamini et al., 2019), the vine mealybug, Planococcus ficus (Signoret) (Le Vieux & Malan, 2013, 2015; Platt et al., 2018, 2019a, b), the citrus mealybug, Planococcus citri (Risso) (Van Niekerk & Malan, 2012) and codling moth, Cydia pomonella L. (Lepidoptera: Tortricidae) (De Waal et al., 2011, 2013; Odendaal et al., 2016a, b). The diapausing larval population of codling moth overwinters in cryptic habitats, for example in old pruning wounds and cracks in the bark of apple trees, which offer an opportunity to use nematodes as a biological control agent prior to their emergence during the next growing season. The performance of EPNs has also been evaluated in laboratory and field bioassays against the false codling moth, Thaumatotibia leucotreta (Meyrick) (Malan et al., 2011; Malan & Moore, 2016; Steyn et al., 2019) and the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Malan & Manrakhan, 2009; James et al., 2018).
EPNs have not yet been evaluated as a biocontrol agent for katydids. A study by MacVean & Capinera (1992), which evaluated the performance of Steinernema carpocapsae (Weiser) Wouts, Mrácek, Gerdin & Bedding against the Mormon cricket Anabrus simplex Hald. (Orthoptera: Tettigoniidae), found that the nematode neither successfully infected nor reduced the survival rate of the cricket.
The aim of the current study was to evaluate the pathogenicity of in vivo-cultured South African species and one exotic species of EPN against the nymphs of P. graminea. Screening was conducted under optimum laboratory conditions to evaluate mortality resulting from EPN infection.
MATERIALS AND METHODS
Source of insects
Nymphs of P. graminea were obtained from multiple wine grape vineyards in the Western Cape. Three sites with a persistent katydid presence were prioritised for the collection of P. gramínea during the summer months of 2016 and 2017 (Fig. 2).
Nymphs were collected using rigid cylindrical plastic containers with perforated lids, after which they were taken to the laboratory of the Department of Conservation Ecology and Entomology at Stellenbosch University, with the aim of establishing a laboratory colony (Du Preez, 2019). The colony never successfully stabilised, and field-collected individuals were used for the purpose of performing laboratory bioassays. Field collected individuals were kept for a minimum of one day and for a maximum of three days, prior to the bioassays: firstly, to provide the opportunity to assess their health and to discard unsuitable individuals; and secondly, to limit the deterioration of their health over time, as was seen in laboratory colonies, for reasons currently unknown.
Source of nematodes
The EPNs used in the present study were obtained from the nematode collection stored at the Department of Conservation Ecology and Entomology, Stellenbosch University (Table 1). All species were locally isolated, except for Steinernema feltiae (Filipjev) Wouts, Mrácek, Gerdin & Bedding, which is an exotic species from Germany. Infective juveniles (IJs) were cultured in vivo, using the last-instar larvae of either the greater wax moth, Galleria mellonella (L.) (Lepidoptera: Pyralidae), or of the common mealworm Tenebrio molitor L. (Coleoptera: Tenebrionidae), at room temperature, using the methods described by Stock & Goodrich-Blair (2012) and by Van Zyl & Malan (2015).
The IJs were harvested over the course of two weeks and transferred to vented culture flasks (50 ml rec max, NUNC) that were stored horizontally at 14°C. The culture flasks were shaken biweekly to mitigate nematode clumping, and to aerate the mixture. The nematodes were used within three weeks of their culture and inspected for health (in terms of motility and mortality) prior to the commencement of bioassays.
Screening
Bioassays were prepared using six-well bioassay plates (BioLite 6-Well MultiDish, Thermo Scientific), which were lined with one circular filter paper disk (30 mm, Grade 1 Whatman, GE Healthcare Life Sciences) per well. The concentration of nematodes was determined using the technique employed by Glazer & Lewis (2000). Nematode concentrations of 200 IJs in 100 μ! of distilled water were inoculated onto each circular disk, whereas the control received 100 μ! of distilled water only.
One katydid nymph was added to each of the six wells using soft forceps, with a glass rectangle being placed over the tray as each well was filled. Using sleight of hand, the original tray cover was slid into place over the glass cover once all the wells had been filled. With rubber bands securing the tray lids, the trays were transferred into 2-L plastic icecream containers, each of which was lined with paper towels moistened with distilled water, which were then each closed with a lid, to maintain high humidity levels. The containers were incubated in a growth chamber at 25°C, in the dark, for 48 h. The mortality of the katydids was determined by gently poking the insects with forceps. The dissection kit equipment, glassware and other potential sources of contaminants were submerged in boiling water and dried, prior to the handling of each treatment and batch.
The number of katydids within each treatment was scaled to use the maximum number of insects available at the time, which resulted in unequal treatment sizes (Table 2). In addition, the potential for high natural mortality prompted the authors to use a higher number of insects in the control group, relative to treatments, in an attempt to collect reliable natural mortality data and to reduce statistical error in this group. Finally, not all EPN species were available for use at the same time (due to the irregularity of when insects were available), thus not all EPN species were assessed through the same number of repetitions. Of note, Heterorhabditis baujardi, H. zealandica, Steinernema innovationi and Steinernema khoisanae only had one repetition. Repetitions were executed on different days, except for H. baujardi and H. zealandica, due to a shortage of katydid hosts, and for S. innovationi and S. khoisanae, for which IJs were not available at the time.
Penetration
Following the conclusion of screening bioassays, cadavers resulting from the different treatments were placed on a sieve, gently rinsed with a handheld water jet and patted dry on paper towelling, to remove surface nematodes. The cadavers were then placed in 90-mm-diameter Petri dishes, each lined with one circular filter paper disk (85 mm, Grade 1 Whatman, GE Healthcare Life Sciences), inoculated with 800 μl of distilled water and incubated at 25°C and > 95% RH in the dark for 24-36 h, to allow for IJ growth and development. The infectivity of the nematodes was determined by dissecting the cadavers and assessing the presence of nematodes. Large sample sizes necessitated the storage of the cadavers at 14°C, to slow down the growth and development of nematodes, which were later evaluated to assess the extent of IJ penetration.
Data analysis
The data obtained were analysed using Microsoft Excel 2010 for the purpose of descriptive statistics, and processed by Statistica 13.3 (Tibco Software Inc., 2017) for comparative analysis.
RESULTS
Screening
Residuals of the mortality response were considered to be normally distributed (Shapiro-Wilk's W = 0.984, P = 0.267), thus permitting the use of a one-way ANOVA. However, Levene's test for the homogeneity of variances failed, necessitating the use of a Games-Howell post hoc analysis to evaluate the responses of the different nematode species.
The highest percentage mortality was obtained by H. zealandica Poinar (n = 12; 100%), H. indica Poinar, Karunakar & David (n = 42; 95.24% ± 3.07%), S. Jeffreyense Malan, Knoetze & Tiedt (n = 60; 93.33% ± 3.69%) and S. yirgalemense Nguyen, Tesfamariam, Gozel, Gaugler & Adams (n = 84; 91.67% ± 3.81%). No significant differences in mortality were found between these EPN species, but significant differences were found compared to the control (P < 0.01). Heterorhabditis noenieputensis (n = 72; 70.83% ± 9.65%, P = 0.034) and H. bacteriophora Poinar (n = 60; 65% ± 8.03%, P = 0.041) also achieved significant mortality relative to the control treatment (n = 128; 25% ± 3.7%). The performance of H. zealandica was found to differ significantly from that of H. bacteriophora (P = 0.047), but no other treatments were found to perform significantly differently from one another (Fig. 3).
Steinernema feltiae (n = 30; 66.67% ± 9.13%), S. litchii Steyn, Knoetze, Tiedt & Malan (n = 30; 66.67% ± 11.79%), S. sacchari Nthenga, Knoetze, Berry, Tiedt & Malan (n = 30; 63.33% ± 19.29%), S. khoisanae Nguyen, Malan & Gozel (n = 18; 44.44% ± 29.4%) and S. innovationi Çimen, Lee, Hatting, Hazir & Stock (n = 12; 8.34% ± 8.34%) were found not to cause mortality significantly different from the control. Although H. baujardi Phan, Subbotin, Nguyen & Moens (n = 12; 83.34% ± 16.67%) achieved high average mortality, the related results did not compute in the Games-Howell post hoc analysis, due to the high variance within the treatment, thus eliminating the possibility of performing a comparative analysis for this species.
Penetration
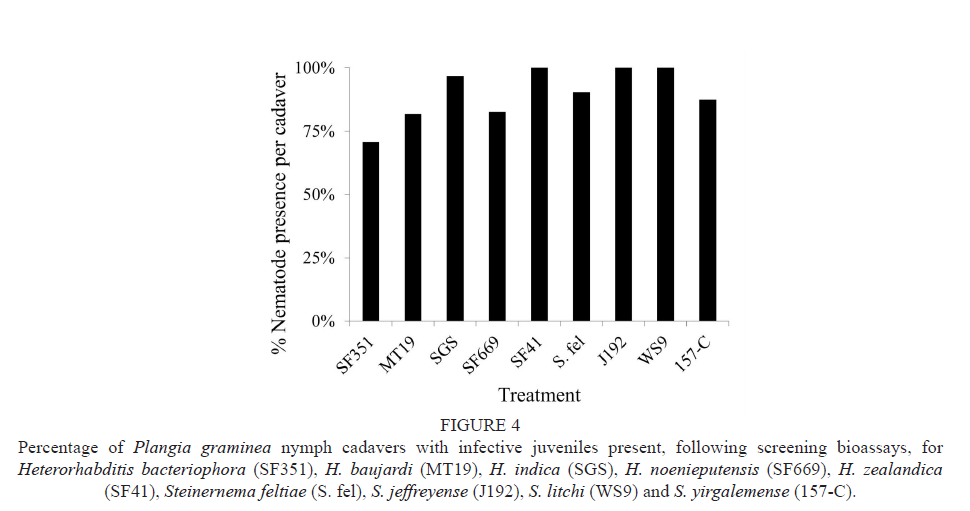
The cadavers of P. gramínea nymphs inoculated with S. innovationi, S. khoisanae and S. sacchari were unintentionally destroyed, following the screening bioassays, and penetration could thus not be confirmed for these species. In all other treatments, the presence of nematodes was confirmed for at least 70% of the cadavers (Fig. 4).
Heterorhabditis zealandica (n = 12), S. jeffreyense (n = 20) and S. litchi (n = 18) had nematodes present in 100% of the cadavers, followed by H. indica (n = 31; 97%), S. feltiae (n = 21; 90%), S. yirgalemense (n = 56; 88%), H. noenieputensis (n = 52; 83%), H. baujardi (n = 11; 82%) and H. bacteriophora (n = 48; 71%). Nematodes within the host cadaver were able to complete their lifecycle in vivo and to produce second-generation offspring with varying success (Fig. 5).
The presence of second-generation nematodes was confirmed in the P. gramínea cadavers inoculated with S. lítchíí (n = 18; 100%); S. jeffreyense (n = 20; 95%), H. zealandica (n = 12; 75%), H. noeníeputensís (n = 22; 73%), S. yírgalemense (n = 39; 67%), H. índica (n = 31; 52%), H. bacteriophora (n = 20; 50%), S. feltiae (n = 21; 48%) and H. baujardi (n = 11; 18%).
DISCUSSION
This is the first study to evaluate the biocontrol potential of EPNs for the control of Plangia graminea. Five EPN species, namely H. indica, H. zealandica, S. jeffreyense, S. yirgalemense and H. baujardi, achieved > 80% control in the laboratory environment, while H. noenieputensis (71%) and H. bacteriophora (65%) performed significantly better than the control (25%). Although H. zealandica performed significantly better than H. bacteriophora, no other statistically significant differences were found between the other treatments. The lack of significance is likely due to the relatively high control group mortality and variation that was found to occur within treatments, which could potentially be resolved in future bioassays by means of increasing the sample size and the number of repetitions, in addition to limiting the extent of control group mortality.
The five most effective EPN species, namely H. indica, H. zealandica, S. Jeffreyense, S. yirgalemense and H. baujardi, were found to have nematodes present in > 82% of the nymph cadavers, supporting the extent of mortality caused by EPN infection, and suggesting a high degree of pathogenicity of these species against the nymphs of P. gramínea. In addition, second- generation nematodes were discovered in the cadavers inoculated with H. indica (52%), H. zealandica (75%), S. jeffreyense (95%) and S. yirgalemense (67%), but with low presence in the case of H. baujardi (18%). The above indicates that nematodes can produce new generations in vivo, thus being theoretically able to produce a new cohort of IJs that can find and infect alternative hosts in soil environments, should infected katydid cadavers fall to the ground. The cadavers were not processed through a modified White trap (White, 1927) to evaluate the production rate of the IJs due to the destructive nature of dissection. The mortality of P. graminea ascribed to H. baujardi could not be evaluated statistically, due to the high variability within the treatment and the low sample size, whereas the treatments of S. feltiae, S. litchii, S. sacchari, S. khoisanae and S. innovationi did not result in significant mortality relative to the control.
Of the five EPN species with high mortality, three were of the genus Heterorhabditis and two were Steinernema species. Two of the species, H. baujardi and H. indica, have not yet been tested against the pests associated with grapevine. During South African surveys for the occurrence of EPNs, H. bacteriophora has been found to be the most common species (Malan et al., 2006; Hatting et al., 2009; Malan et al., 2011), with it also being the only EPN used in the current study that is available commercially (Hatting et al., 2018). The methods of mass culturing this EPN have been well documented (Shapiro-Ilan & Gaugler, 2002; Inman et al., 2012), with its performance having been evaluated against multiple species since its discovery in 1976 (Smart, 1995; Van Lenteren, 2012). However, in the current study, the mortality that was caused among the katydids using H. bacteriophora was relatively low (65%) compared to the mortality that resulted from the administration of the other local isolates.
Heterorhabditis baujardi was recently reported from two independent surveys conducted in South Africa (Steyn et al., 2017b; Abate et al., 2018), with both claiming it to be the first record of this species. The species was also reported from Cameroon in Africa (Kanga et al., 2012). However, the potential to mass culture H. baujardi as a biocontrol agent is, as yet, unknown.
During 2016, the presence of H. indica (KU945293) was reported from Bonnievale, which is situated in the Western Cape province. Ehlers et al. (2000) successfully mass cultured H. indica from isolates in India. The success of this nematode in infecting P. graminea in the laboratory encourages future research into the use of this heterorhabditid for local mass culture. Evaluating the performance of this EPN against other insect pests of grapevine to determine its biocontrol potential would be beneficial.
Heterorhabditis zealandica was found to be the best performing EPN in the present study, with success in the mass culture of this nematode having already been demonstrated by Ferreira et al. (2014). This species was previously evaluated against the banded fruit weevil (Ferreira & Malan, 2014; Dlamini et al., 2019) and false codling moth (Malan et al., 2011; Malan & Moore, 2016). Both these insects are key pests of grapevine. Research into the mass culture and application of H. zealandica in integrated pest management programmes is also likely to increase awareness of the benefit of its control potential against katydids.
Steinernema jeffreyense was previously evaluated against codling moth and false codling moth in both laboratory and field environments (De Waal et al., 2011, 2013; Odendaal et al., 2016a; Steyn et al., 2019), as well as against the vine mealybug (Platt et al., 2018, 2019a, b). Methods for mass culturing this nematode species have been demonstrated by Dunn & Malan (2019).
The performance of S. yirgalemense was previously evaluated against false codling moth (Malan et al., 2011; Steyn et al., 2019), codling moth (De Waal et al., 2011) and mealybugs (Van Niekerk & Malan, 2012; Le Vieux & Malan, 2013, 2015; Platt et al., 2018) in both laboratory and field environments. It has also been evaluated in terms of above-ground application for the control of codling moth (Odendaal et al., 2016b). The nematode is currently regarded as the best candidate for biological control in South Africa, due to its demonstrated pathogenicity against multiple insect pests. It has already been successfully mass cultured, and research into its production (Ferreira et al., 2016; Dunn et al., 2019) and formulation (Kagimu & Malan, 2019) was under way at the time of the present research.
For the control of P. graminea in the field, EPNs would need to be applied above-ground onto the leafy canopy of grapevine, particularly when nymphs are still small (September to October) to have the best opportunity for suppression.
A significant constraint in the present study was the number of P. graminea nymphs available for bioassays. High mortality in the laboratory colony necessitated the use of field-collected individuals, as the strategies employed to increase katydid fitness and survival did not produce satisfactory results. Treatments and repetitions were scaled to use the maximum number of katydids available, resulting in unequal sample sizes between the treatments and repetitions. The sensitivity of P. graminea to suboptimal environments likely resulted in the increased control group mortality and variance within treatments that were observed. As a result of the above, the efficacy of treatments might have been underestimated, thus resulting in suppressed statistical significance. The findings pose the question of whether, and to what extent, the susceptibility of katydid nymphs to EPN infection is influenced by the environmental conditions in the laboratory, and how this is likely to translate to field applications in areas that are closer to their natural habitat.
Future research is recommended in terms of evaluating the field efficacies of the best-performing EPN species identified in the present study, as well as evaluating in vitro-produced EPNs, when they are available. Such research would build up an EPN control profile for P. graminea and help to motivate further research, development and investment into the use of biological control agents for insect pest control and for their adoption into integrated pest management programmes.
In conclusion, the results of the present study showed the high susceptibility of P. graminea to H. indica, H. zealandica, S. jeffreyense and S. yirgalemense, which, in turn, serves to confirm their pathogenicity. The results also demonstrated that these species can penetrate and to complete their life cycle within the host, thus indicating their ability to produce a new cohort of IJs with the potential to infect alternative insect hosts roaming in soil environments.
LITERATURE CITED
Abate, A.A., Slippers B., Wingfield, M.J., Malan, A.P. & Hurley, B.P., 2018. Diversity of entomopathogenic nematodes and their symbiotic bacteria in South African plantations and indigenous forests. Nematology 20, 355 -371. doi:10.1163/15685411-00003144 [ Links ]
Agro-Organics., 2010. Krompokkels, witluis / mealybug, snuitkewers / snoutbeetle. Newsletter 27. http://www.agro-organics.co.za/wp-content/uploads/2017/12/Bio-Insek-krompokkels-2010.pdf [ Links ]
Allsopp, E., 2012. Long-horned grasshoppers prevail again. https://www.wineland.co.za/long-horned-grasshoppers-prevail-again/ [ Links ]
Bailey, W.J. & Rentz, C.F., 1990. The Tettigoniidae: Biology, Systematics and Evolution, Crawford House Press, Bathurst. [ Links ]
De Waal, J.Y., Malan, A.P. & Addison, M.F., 2011. Efficacy of entomopathogenic nematodes (Rhabditida: Heterorhabditidae and Steinernematidae) against codling moth, Cydia pomonella (Lepidoptera: Tortricidae) in temperate regions. Biocontr. Sci. Technol. 21, 1161 - 1176. doi:10.1080/09583157.2011.607922 [ Links ]
De Waal, J.Y., Malan, A.P. & Addison, M.F., 2013. Effect of humidity and a superabsorbent polymer formulation on the efficacy of Heterorhabditis zealandica (Rhabditida: Heterorhabditidae) to control codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Biocontr. Sci. Technol. 23, 62 -78. doi:10.1080/09583157.2012.736472 [ Links ]
Dillman, A.R. & Sternberg, P.W., 2012. Entomopathogenic nematodes. Curr. Biol. 22, R430 - R431. doi:10.1016/j.cub.2012.03.047 [ Links ]
Dlamini, B., Malan, A.P. & Addison, P., 2019. Control of the banded fruit weevil, Phlyctinus callosus (Schönherr) (Coleoptera: Curculionidae) using entomopathogenic nematodes. Austral. Entomol. 58, 687 - 695. doi:10.1111/aen.12386 [ Links ]
Doubell, M., 2017. Katydid (Orthoptera: Tettigoniidae) bio-ecology in Western Cape vineyards. MSc thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. https://scholar.sun.ac.za/handle/10019.1/102630 [ Links ]
Dunn, M. & Malan, A.P., 2019. Production optimization of a novel entomopathogenic nematode, Steinernema jeffreyense, in shake flasks. BioControl 65, 223 - 233. doi:10.1007/s10526-019-09977-7 [ Links ]
Du Preez, F., 2019. Biological control of two sporadic grapevine pests, Plangia graminea and Lobesia vanillana using entomopathogenic nematodes. MScAgric thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. https://scholar.sun.ac.za/handle/10019.1/105714 [ Links ]
Ehlers, R.U., Niemann, I., Hollmer, S., Strauch, O., Jende, D., Shanmugasundaram, M., Mehta, U.K., Easwaramoorthy, S.K. & Burnell, A. 2000. Mass production potential of the bacto-helminthic biocontrol complex Heterorhabditis indica - Photorhabdus luminescens. Biocontrol Sci. Technol. 10, 607 - 616. doi:10.1080/095831500750016406 [ Links ]
Ferreira, T., Addison, M.F. & Malan, A.P. 2014. In vitro liquid culture of a South African isolate of Heterorhabditis zealandica for the control of insect pests. Afr. Entomol. 22, 80 - 92. doi:10.4001/003.022.0114 [ Links ]
Ferreira, T., Addison, M.F. & Malan, A.P. 2016. Development and population dynamics of Steinernema yirgalemense (Rhabditida: Steinernematidae) and growth characteristics of its associated Xenorhabdus indica symbiont in liquid culture. J. Helminthol. 90, 364 - 371. doi:10.1017/ S0022149X15000450 [ Links ]
Ferreira, T. & Malan, A.P. 2014. Potential of entomopathogenic nematodes for the control of the banded fruit weevil, Phlyctinus callosus (Schönherr) (Coleóptera: Curculionidae). J. Helminthol. 88, 293 - 301. doi:10.1017/ S0022149X13000175 [ Links ]
Glazer, I. & Lewis, E.E. 2000. Bioassays for entomopathogenic nematodes. In: Navon, A. & Ascher, K.R.S. (eds). Bioassays of Entomopathogenic Microbes and Nematodes, CABI, Wallingford. pp. 229 - 247. [ Links ]
Hatting, J., Stock, P.S. & Hazir, S. 2009. Diversity and distribution of entomopathogenic nematodes (Steinernematidae, Heterorhabditidae) in South Africa. J. Invertebr. Pathol. 102, 120 - 128. doi:10.1016/j. jip.2009.07.003 [ Links ]
Hatting, J.L., Moore, S.D. & Malan, A.P. 2018. Microbial control of phytophagous invertebrate pests in South Africa: current status and future. J. Invertebr. Pathol. doi:10.1016/j.jip.2018.02.004 [ Links ]
Inman III, F.L., Singh, S. & Holmes, L.D. 2012. Mass production of the beneficial nematode Heterorhabditis bacteriophora and its bacterial symbiont Photorhabdus luminescens. Indian J. Microbiol. 52, 316 - 324. doi:10.1007/s12088-012-0270-2 [ Links ]
James, M., Malan, A.P. & Addison, P. 2018. Surveying and screening South African entomopathogenic nematodes for the control of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann). Crop Prot. 105, 41 -48. doi:10.1016/j.cropro.2017.11.008 [ Links ]
Kagimu, N. & Malan, A.P., 2019. Alginate and diatomaceous earth formulations of entomopathogenic nematodes. BioControl 64, 413 - 422. doi:10.1007/s10526-019-09945-1 [ Links ]
Kanga, F.N., Waeyenberge, L., Hauser, S. & Moens, M., 2012. Distribution of entomopathogenic nematodes in Southern Cameroon. J. Invertebr. Pathol. 109, 41 - 51. doi:10.1016/j.jip.2011.09.008 [ Links ]
Kaya, H.K. & Gaugler, R., 1993. Entomopathogenic nematodes. Annu. Rev. Entomol. 38, 181 - 206. doi:10.1146/annurev.en.38.010193.001145 [ Links ]
Le Vieux, P.D. & Malan, A.P., 2013. The potential use of entomopathogenic nematodes to control Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). S. Afr. J. Enol. Vitic. 34, 296 - 306. doi:10.21548/34-2-1109 [ Links ]
Le Vieux, P.D. & Malan, A.P., 2015. Prospects for using entomopathogenic nematodes to control the vine mealybug, Planococcus ficus, in South African vineyards. S. Afr. J. Enol. Vitic. 36, 59 - 70. doi:10.21548/36-1-937 [ Links ]
MacVean, C.M. & Capinera, J.L. 1992. Field evaluation of two microsporidian pathogens, an entomopathogenic nematode, and carbaryl for suppression of the Mormon cricket, Anabrus simplex Hald. (Orthoptera: Tettigoniidae). Biol. Control. 2, 59 - 65. doi:10.1016/1049-9644(92)90076-P [ Links ]
Malan, A.P., Nguyen, K.B. & Addison, M.F. 2006. Entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) from the southwestern parts of South Africa. Afr. Plant Prot. 12, 65 - 69. https://hdl.handle.net/10520/EJC87795 [ Links ]
Malan, A.P., Knoetze, R. & Moore, S.D. 2011. Isolation and identification of entomopathogenic nematodes from citrus orchards in South Africa and their biocontrol potential against false codling moth. J. Invertebr. Pathol. 108, 115 - 125. doi:10.1016/j.jip.2011.07.006 [ Links ]
Malan, A.P., Knoetze, R. & Tiedt, L. 2014. Heterorhabditis noenieputensis n. sp. (Rhabditida: Heterorhabditidae), a new entomopathogenic nematode from South Africa. J. Helminthol. 88, 139 - 151. doi:10.1017/ S0022149X12000806 [ Links ]
Malan, A.P. & Moore, S.D. 2016. Evaluation of local entomopathogenic nematodes for the control of false codling moth, Thaumatotibia leucotreta (Meyrick, 1913), in a citrus orchard in South Africa. Afr. Entomol. 24, 489 - 501. doi:10.4001/003.024.0489 [ Links ]
Malan, A.P. & Manrakhan, A. 2009. Susceptibility of the Mediterranean fruit fly (Ceratitis capitata) and the Natal fruit fly (Ceratitis rosa) to entomopathogenic nematodes. J. Invertebr. Pathol. 100, 47 - 49. doi:10.1016/j.jip.2008.09.007 [ Links ]
Nguyen, K., Malan, A. & Gozel, U., 2006. Steinernema khoisanae n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from South Africa. Nematology 8, 157 - 175. doi:10.1163/156854106777998728 [ Links ]
Nthenga, I., Knoetze, R., Berry, S., Tiedt, L.R. & Malan, A.P., 2014. Steinernema sacchari n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from South Africa. Nematology 16, 475 - 494. doi:10.1163/15685411-00002780 [ Links ]
Odendaal, D., Addison, M.F. & Malan, A.P. 2016a. Entomopathogenic nematodes for the control of the codling moth (Cydia pomonella L.) in field and laboratory trials. J. Helminthol. 90, 615 - 623. doi:10.1017/ S0022149X15000887 [ Links ]
Odendaal, D., Addison, M.F. & Malan, A.P. 2016b. Evaluation of above-ground application of entomopathogenic nematodes for the control of diapausing codling moth (Cydia pomonella L.) under natural conditions. Afr. Entomol. 24, 61 - 74. doi:10.4001/003.024.0061 [ Links ]
Picker, M., Griffiths, P. & Weaving, A., 2004. Field Guide to Insects of South Africa. Struik, Cape Town. [ Links ]
Platt, T., Stokwe, N.F. & Malan, A.P., 2018. Potential of local entomopathogenic nematodes for control of the vine mealybug, Planococcus ficus. S. Afr. J. Enol. Vitic. 39, 208 - 215. doi:10.21548/39-2-3158 [ Links ]
Platt, T., Stokwe, N.F. & Malan, A.P., 2019a. Foliar application of Steinernemayirgalemense to control Planococcus ficus: assessing adjuvants to improve efficacy. S. Afr. J. Enol. Vitic. 40(1), 13 - 19. doi:10.21548/40-1-2920 [ Links ]
Platt, T., Stokwe, N.F. & Malan, A.P., 2019b. Grapevine leaf application of Steinernema yirgalemense to control Planococcus ficus in vineyards conditions S. Afr. J. Enol. Vitic. 40(1), 75 - 83. doi:10.21548/40-1-3141 [ Links ]
Shapiro-Ilan, D. & Gaugler, R. 2002. Production technology for entomopathogenic nematodes and their bacterial symbionts. J. Ind. Microbiol. Biotechnol. 28, 137 - 146. doi:10.1038/sj/jim/7000230 [ Links ]
Smart, G.C., 1995. Entomopathogenic nematodes for the biological control of insects. J. Nematol. 27, 529 - 534. https://pubmed.ncbi.nlm.nih.gov/19277318/ [ Links ]
Steyn, W.P., Knoetze, R., Tiedt, L.R. & Malan, A.P., 2017a. Steinernema litchii n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from South Africa. Nematology 19, 1157 - 1177. doi:10.1163/15685411-00003116 [ Links ]
Steyn, W.P., Malan, A.P., Daneel, M.S. & Slabbert, R.M., 2017b. Entomopathogenic nematodes from north-eastern South Africa and their virulence against the false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae). Biocontr. Sci. Technol. 27, 1265 - 1278. doi:10. 1080/09583157.2017.1391174 [ Links ]
Steyn, V.M., Malan, A.P. & Addison, A., 2019. Control of false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae), using in vitro-cultured Steinernema jeffreyense and S. yirgalemense. Biol. Control 138(104052), 1 - 8. doi:10.1016/j.biocontrol.2019.104052 [ Links ]
Stock, S.P. & Goodrich-Blair, H., 2012. Nematode parasites, pathogens and associates of insects and invertebrates of economic importance. In: Lacey, L.A. (ed). Manual of Techniques in Invertebrate Pathology (2nd ed), Academic Press, Oxford. pp. 353 - 426. [ Links ]
Tibco Software Inc., 2017. Statistica (data analysis software system), version 13. [ Links ]
Van Lenteren, J.C., 2012. The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. BioControl 57, 1 - 20. doi:10.1007/s10526-011-9395-1 [ Links ]
Van Niekerk, S. & Malan, A.P., 2012. Potential of South African entomopathogenic nematodes (Heterorhabditidae and Steinernematidae) for control of the citrus mealybug, Planococcus citri (Pseudococcidae). J. Invertebr. Pathol. 111, 166 - 174. doi:10.1016/j.jip.2012.07.023 [ Links ]
Van Zyl, C. & Malan, A.P., 2015. Cost-effective culturing of Galleria mellonella and Tenebrio molitor and entomopathogenic nematode production in various hosts. Afr. Entomol. 23, 361 - 375. [ Links ]
White, G.F., 1927. A method for obtaining infective nematode larvae from cultures. Science 66, 302 - 303. doi:10.1126/science.66.1709.302-a [ Links ]
Submitted for publication: October 2021
Accepted for publication: August 2022
* Corresponding author: E-mail address: fdupreez@sun.ac.za
Acknowledgements: The authors wish to thank D.G. Nel (Centre for Statistical Consultation, Stellenbosch University) and K.L. Pringle (Department of Conservation Ecology and Entomology, Stellenbosch University) for their assistance with statistical analyses. Financial support was provided by the South African Table Grape Industry (SATI), Winetech and the Technology and Human Resources for Industry Programme (THRIP-TP14062571871)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}