










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
versão On-line ISSN 2224-7904
versão impressa ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.43 no.1 Stellenbosch 2022
http://dx.doi.org/10.21548/43-1-4927
ARTICLES
Dry Matter Accumulation, Seasonal Uptake and Partitioning of Mineral Nutrients by Vitis vinifera L. cv. Sultanina Grapevines in the Lower Orange River Region of South Africa - A Preliminary Investigation
W.J. ConradieI; P.J. RaathI, II, *; A.R. MulidziIII; C.L. HowellIII
IFormerly ARC Infruitec-Nietvoorbij#, Private Bag X5026, 7599, Stellenbosch, South Africa
IIDepartment Horticultural Science, Stellenbosch University, Private Bag X1, 7602 Matieland, South Africa
IIIARC Infruitec-Nietvoorbij, Private Bag X5026, 7599 Stellenbosch, South Africa
ABSTRACT
Table grapes are harvested from November until early February in the Orange River Region of South Africa, where a functional leaf canopy is present for up to six months of post-harvest. Seasonal uptake and partitioning of mineral nutrients by 'Sultanina' grapevines in this long-season area were quantified in a preliminary investigation. Entire grapevines growing on two different soil types were sampled during the growing season. On sandy soil, further from the river, grapevine vigour was not excessive; however, on the fertile alluvial soil vigour was higher with active post-harvest re-growth. At budbreak, reserve N played an important role on the sandy soil, while it was less important on the alluvial one. During the post-harvest period, 34.3% of the annual N-requirement was absorbed by grapevines cultivated on sandy soil at post-harvest, while only 17.0% was absorbed on the alluvial soil. Approximately 4.0 kg N was utilized for the production of one ton of fresh grapes. A major fraction of the annual P-requirement (41.9%) was absorbed post-harvest by grapevines on the sandy soil. Grapevines on alluvial soil absorbed more P at pre-harvest; however, P was somehow lost post-harvest. Comparable amounts of K and Mg were absorbed by both selections of grapevines. Sandy soil grapevines absorbed K and Mg at post-harvest, while a net loss occurred for those of alluvial soil. Calcium utilized by sandy soil grapevines was less than half that utilized by those of alluvial soil. The seasonal absorption pattern of Ca was comparable for both soils. Results suggest that that seasonal uptake and partitioning of mineral nutrients are affected by soil type and grapevine vigour.
Key words: calcium, magnesium, nitrogen, phosphorus, potassium, table grapes
INTRODUCTION
To understand the mineral nutrient uptake of grapevines, knowledge about whole-vine nutrient contents (concentration x biomass) are exceptionally important (Schreiner 2004, and references therein; Pradubsuk, 2008; Pradubsuk & Davenport, 2010). Initial studies, aimed at quantifying seasonal uptake and partitioning of mineral nutrients, dealt with above-ground organs only, while the role of the permanent structure (roots & trunk) was not taken into account (Alexander, 1958; Lafon et al., 1965; Marocke et al., 1976; Rogiers et al., 2017; Ferrara. et al., 2018). In the early 1980's, a number of whole-vine studies were carried out in South Africa for Chenin blanc grapevines growing in sandy culture (Conradie, 1980; Conradie, 1981a; Conradie, 1981b; Conradie, 1986; Conradie, 1990; Conradie, 1991a; Conradie, 1991b; Conradie, 1992). Studies with entire, field-grown grapevines, although mainly aimed at studying nitrogen (N), have also been reported from other countries (Araujo & Williams, 1988; Löhnertz, 1988; Löhnertz, 1991; Williams, 1991; Williams & Biscay, 1991; Mullins et al., 1992; Hanson & Howell, 1995; Bates et al., 2002; Vos, 2003; Schreiner, 2004; Schreiner & Scagel., 2006 ; Holzapfel & Treeby, 2007; Tomasi et al., 2015; Ferrara et al., 2018). Seasonal uptake and partitioning of mineral nutrients were affected by factors such as cultivar type, climate, crop load, and irrigation, with comparable patterns obtained during the pre-harvest period (Vos, 2003; Conradie, 2004; Schreiner, 2004; Barnuud et al., 2014). During the post-harvest period, however, divergent patterns were obtained in the different studies.
In warm countries, such as South Africa, there is a functional canopy for up to four months after harvest (Conradie, 1991a; Leão & Chaves, 2020). Roots grow vigorously during this specific period, resulting in active uptake of N, phosphorus (P), potassium (K), calcium (Ca) and magnesium (Mg) (Conradie, 1980; Conradie, 1981a; Conradie, 1981b; Pradubsuk & Davenport, 2010; Rogiers et al., 2017). The amount of N absorbed during the post-harvest period varied between 27% and 38% of the annual requirement, with the largest fraction (68%) of N absorbed during this period being retained in grapevine roots, trunks and cordons (Conradie, 1986; Pradubsuk & Davenport, 2010). Nitrogen absorbed during the post-harvest period account for up to 60% of the total amount of N-reserves present at the start of the next season (Conradie, 1992; Masclaux-Daubresse et al., 2010; Ferrara et al. 2018). In cool areas, where the leaf canopy is functional for only a short period after harvest (Hanson & Howell, 1995; Bates et al., 2002), nutrients are less effectively absorbed during this stage (Mârtensson, 2020). Consequently, even though roots may accumulate N during this short post-harvest period (Bates et al., 2002; Schreiner & Scagel, 2006), an apparent loss of N has been reported in short-season growing areas (Hanson & Howell, 1995).
In contrast to short-season growing areas, an exceptionally long post-harvest period may be experienced in extremely warm areas, such as the Orange River Region of South Africa. Although grapevines in other regions of the country are mainly harvested in February, table grapes in the Orange River Region are generally harvested from November until the beginning of February, resulting in a post-harvest period of up to six months. Active vegetative growth is a common phenomenon during this period, especially on the alluvial soils in the flood-plain of the Orange River, while it occurs to a lesser extent on the sandy soils further from the river. The dry matter partitioning, seasonal uptake, and partitioning of mineral nutrients have not yet been properly quantified for such a long-season growing area. Therefore, the objective of this preliminary investigation was to address above-mentioned aspects, using field-grown grapevines on two different soil types commonly found in the region.
MATERIALS AND METHODS
Sandy soil further away from the river
The study was carried out in a commercial Vitis vinifera L. (cv. Sultanina) vineyard, planted in 1989 on a sandy soil further from the river near Augrabies. This locality, situated at an altitude of 650 m and 28° 66' South latitude, is in a class V climatic region (Winkler, 1962). The well-aerated soil belonged to the Plooysburg form (Soil Classification Working Group, 1991), and consisted of 600 mm to 900 mm of gravelly, sandy loam on undulating, nodular limestone (Aridisol, USDA soil classification). Grapevines were grafted onto 110 Richter rootstock, planted 3.0 m x 1.8 m, trained on a gable trellis with four cordon arms (Zeeman, 1981) and cane pruned, allowing ten canes per grapevine and 12 to 18 nodes per cane. Normal management practices, as applicable to table grape vineyards, were followed. The vineyard was irrigated by means of 4.0 L/h drippers, spaced 0.6 m apart. Annual fertilisation, generally applied in three increments (after budbreak, fruit-set, post-harvest), amounted to 120 kg N/ha, 32 kg P/ha and 76 kg K/ha. Entire grapevines, of which there were two at each sampling date, were randomly harvested six times during the 1999/2000 season - i.e. before bud break (8-10 August), fruit set (22-24 September), harvest (10-12 December), end February, end April (start leaf senescence), and end June (end leaf fall). Due to the destructive nature of the sampling of the grapevines as well as the preliminary nature of the study, no more than two of them could be removed at each sampling time to minimize losses to the producers. The root system of each grapevine was manually excavated down to a depth of 0.75 m, from a 3.24 m2 area (1.8 m x 1.8 m). Roots were carefully removed from the soil by hand. Excavated grapevines were separated into fine roots (diameter < 2 mm), medium and thick roots (diameter > 2mm), trunk (including below-ground portion), cordon wood (older than one year), shoots (current season's growth), leaves, and clusters. Separated tissues were dried in a forced-air oven at 60°C until sample weight became constant. Dry tissues were weighed and ground to pass through a 40-mesh screen. Total N concentration of each sample was determined by combustion (Campbell, 1991), while P, K, Ca, and Mg were determined following dry ashing in a microwave furnace and uptake into acidified aqueous solution by use of a Varian 200 inductively coupled atomic emission spectrometer.
Fertile alluvial soil in flood-plain of Orange River
The study was carried out in an own-rooted Sultanina vineyard, planted in 1998 in the Blouputs area. This locality, situated at an altitude of 455 m and 28° 50' South latitude, is also a class V climatic region (Winkler 1962). During the summer months (i.e. from December to February), maximum temperatures at Blouputs follow similar trends to those observed at Augrabies but are generally about 2°C higher. Furthermore, maximum and minimum temperatures for the two years of the field study were similar (data not shown). The soil, a loamy-sand texture, belonged to the Dundee form (Soil Classification Working Group, 1991) and consisted of alluvial deposits (Entisol, USDA soil classification). Grapevines were planted 3.3 m x 1.8 m and the vineyard was flood-irrigated. Training system, pruning, and management practices were identical to those at the sandy soil further from the river. Entire grapevines, of which there were two at each sampling date, were harvested six times during the 2001/2002 season - i.e. before bud break (6-8 August), fruit-set (19-21 September), harvest (12-14 December), end February, end April (start leaf senescence), and end June (end leaf fall). Analytical procedures were similar to those described above.
The amounts of the different macro-elements contained in each of the organs at each of the six sampling dates were calculated by multiplying the dry weight of each organ with the concentration of the different elements (B) in the organ harvested for analyses.
The amount of N, P, K, Mg, and Ca in the plant tissues were calculated as follows:

where A is the amount contained (kg/ha), DMP is the dry weight in t/ha, B is the plant element concentration (%) and 10 is the conversion factor to obtain kg/ha.
Due to logistical and budgetary constraints, it should be noted that the use of isotopes was beyond the scope of the study. As the objective of the investigation was to quantify dry matter partitioning, seasonal uptake, and partitioning of mineral nutrients in a long-season growing area in a preliminary field study, the nature of the study was such that no treatments were applied that could be replicated.
RESULTS AND DISCUSSION
Accumulation of dry matter
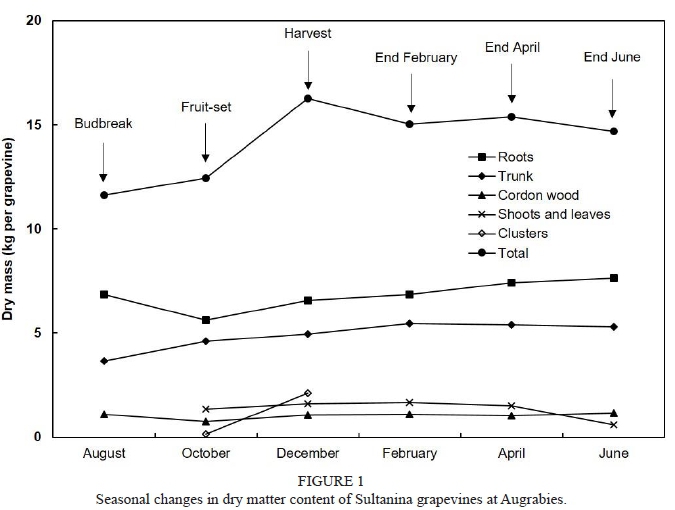
For the grapevines on the sandy soil further from the river near Augrabies, dry mass of roots (6.85 kg) accounted for 59% of total dry mass (11.61 kg) at budbreak (Fig. 1). In studies with field-grown grapevines (Williams, 1991; Williams & Biscay, 1991; Mullins et al., 1992; Hanson & Howell, 1995; Bates et al., 2002; Schreiner & Scagel, 2006; Grechi et al., 2007; Kodur et al., 2010), dry mass of roots rarely exceeded 3 kg per grapevine. Even though variability of vine size and soil properties can make it difficult to obtain accurate biomass data for roots (Schreiner, 2004; Kodur et al., 2010), above-mentioned indicated that the root system was well developed at Augrabies. In accordance with previous results (Conradie, 1980; Schreiner & Scagel, 2006), dry mass of roots declined between budbreak and fruit-set, whereafter there was a gradual increase. On account of grapevines not being sampled at veraison, it was not possible to ascertain whether dry mass of roots declined between veraison and harvest (Conradie, 1980; Williams, 1991). Dry mass of the trunk showed a gradual increase throughout the growing season.
Between fruit-set and harvest, the dry mass of vegetative growth (shoots & leaves) increased relatively slowly, from 1.35 kg per grapevine to 1.60 kg per grapevine. Although comparable rates were reported (Conradie, 1980; Hanson & Howell, 1995), other studies have found that shoots and leaves still accumulate dry material at a fast rate during this period (Williams, 1987a; Mullins et al., 1992; Schreiner & Scagel, 2006). Total dry mass amounted to 16.25 kg per grapevine at harvest, with the roots accounting for 40.3%, the trunk for 30.5%, the cordon wood for 6.5%, the shoots and leaves for 9.8%, and the clusters for 12.9%. At harvest, clusters (2.10 kg per grapevine) contained 57% of the aerial dry mass of new growth (clusters, shoots & leaves). In a study with field-grown Chenin blanc (Mullins et al., 1992), where the dry mass of clusters was much higher (5.20 kg per grapevine), clusters accounted for a comparable fraction (54.9%) of the aerial dry mass of new growth. This appears to support the view that clusters of a well-balanced vine should contain at least 50% newly grown above-ground biomass at harvest (Conradie, 1991; Bates et al., 2002). Where clusters contain a lower fraction of total dry mass, the dynamics of growth may be affected. Roots of the Augrabies grapevines accumulated dry matter during the post-harvest period, albeit at a relatively slow rate. Over the season (budbreak to end leaf-fall), dry mass increased by 3.07 kg per grapevine (i.e. from 11.61 kg to 14.68 kg per grapevine).
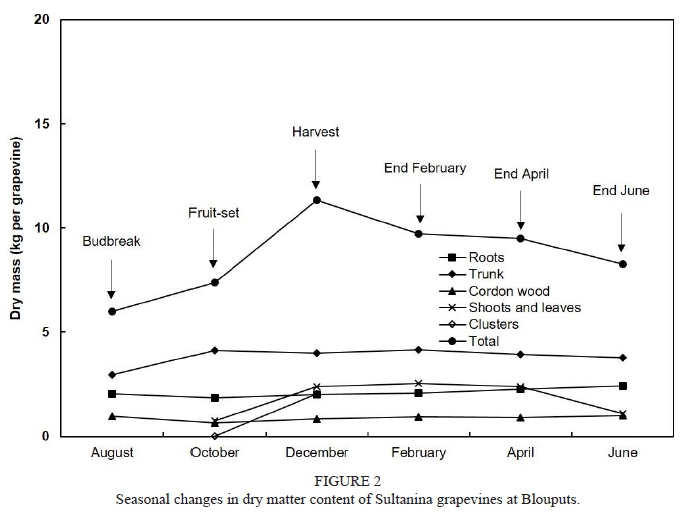
On the alluvial soil at Blouputs, dry mass of roots amounted to 2.05 kg per grapevine at budbreak (Fig. 2), which was substantially lower than the values obtained at Augrabies. This is possibly due to the younger own-rooted grapevines growing on a different soil form. However, total dry mass of roots was still higher than values obtained in other studies for mature, field-grown grapevines (Williams, 1991; Williams & Biscay, 1991; Hanson & Howell, 1995; Bates et al., 2002; Schreiner & Scagel, 2006; Grechi et al., 2007; Kodur et al., 2010). The trunk contained more dry material than the roots throughout the season. Similar results were reported for other field studies (Williams, 1991; Williams & Biscay, 1991, Mullins et al., 1992, Hanson & Howell, 1995; Schreiner & Scagel, 2006). Dry mass of roots showed a minor decline between budbreak and fruit-set, whereafter it increased gradually up to the end of the season. Following an initial increase between budbreak and fruit-set, the dry mass of the trunk stayed relatively constant throughout the rest of the growing season. Between fruit-set and harvest, the dry mass of vegetative growth (shoots & leaves) increased substantially from 0.74 kg per grapevine to 2.40 kg per grapevine. At harvest, total dry mass amounted to 11.32 kg per grapevine, with 17.8% in the roots, 35.4% in the trunk, 7.5% in cordon wood, 21.2% in shoots and leaves, and 18.1% in the clusters. Clusters (2.05 kg per grapevine) accounted for only 46% of the aerial dry mass of new growth (clusters, shoots & leaves) at harvest. Even though grapevines at Blouputs seemed to grow relatively slowly from budbreak to fruit-set in comparison to those at Augrabies, vigorous growth occurred between fruit-set and harvest. Active growth during the latter part of the pre-harvest period may have enhanced the allocation of nutrients to the growth of annual vegetative organs, at the expense of clusters (Keller & Koblet, 1995), resulting in yield being lower than that at Augrabies. Dry mass of roots and shoots continued to increase until the end of February, while roots showed an increasing trend until the end of leaf-fall. During the course of the growing season, dry mass increased by 2.28 kg per grapevine, from 6.00 kg to 8.28 kg.
Yield
Grapevines on the sandy soil further from the river at Augrabies yielded 14 kg per grapevine, amounting to 25.9 ton/ha (1852 grapevines/ha). Yield was marginally lower (13.5 kg per grapevine) on the alluvial soil at Blouputs, amounting to 22.7 ton/ha (1683 grapevines/ha). Seasonal uptake of mineral nutrients will be reported as kg/ha in the rest of the discussion.
Nitrogen
Budbreak to fruit-set
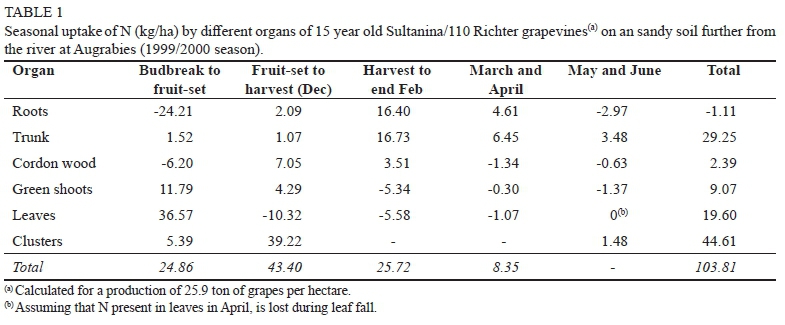
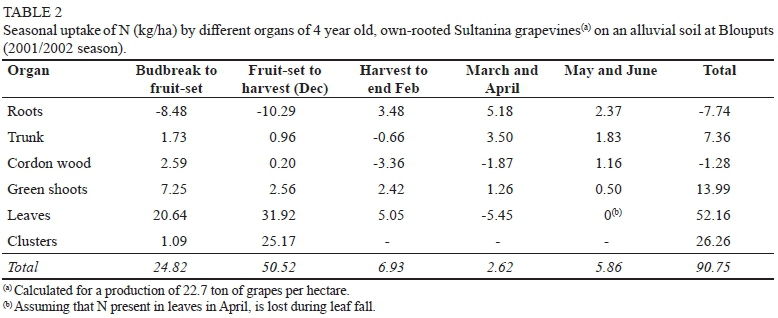
At Augrabies, new growth (green shoots, leaves & clusters) required 53.75 kg N/ha between budbreak and fruit-set (Table 1). Translocation from roots and cordon wood accounted for 30.41 kg/ha, suggesting that absorption of soil-N must have amounted to 24.86 kg/ha. At Blouputs, new growth required less N (28.98 kg/ha) between budbreak and fruit-set (Table 2). This may be ascribed to the possibly of "fruit-set" sampling being done too late at Augrabies, as indicated by the relatively high N content of clusters (5.39 kg/ha). From budbreak to fruit-set, however, reserve N played a relatively minor role at Blouputs. Active translocation from the permanent structure appeared to supply only 4.16 kg N/ha, resulting in the two vineyards absorbing an identical amount of soil-N (24.8 kg/ha). This is in broad agreement with other findings, that the amount of N remobilized from roots, cordons, and trunk between budbreak and fruit-set accounts for 20% to 40% of that needed by shoots, leaves and clusters (Conradie 1980; Williams 1991; Masclaux-Daubresse et al., 2010; Ferrara et al. 2018).
Fruit-set to harvest
At Augrabies, 43.4 kg N/ha was absorbed from the soil between fruit-set and harvest (Table 1). During this period, N was also remobilized from the leaves, while clusters were the largest sink for N. At Blouputs, 50.52 kg N/ha was absorbed from the soil, with N also being remobilized from the roots (Table 2). No N appeared to be remobilized from the leaves, with leaves being a stronger sink for N than clusters. The fact that grapevines at Blouputs were growing more vigorously, as indicated by the higher dry mass of shoots and leaves (Fig. 2), may have resulted in different partitioning patterns for Augrabies and Blouputs. At harvest, clusters at Augrabies contained 44.61 kg N/ha compared to only 26.26 kg N/ha at Blouputs, partly on account of the yield at Blouputs being 12.3% lower; however, clusters also contained lower levels of N (data not shown). Other studies have also indicated that levels of juice-N may be reduced in vineyards that grow too vigorously (Keller & Koblet, 1995; Wheeler & Pickering, 2003).
Post-harvest
At Augrabies, active absorption of N during January and February (25.72 kg/ha), resulted in the permanent structure (roots, stem, and hard wood) gaining N, while remobilization from shoots and leaves (10.92 kg/ha) also occurred (Table 1). Absorption continued during March and April, albeit at a slower rate (8.35 kg N/ha). During the leaf-fall period (May & June), little N (1.48 kg/ha) was absorbed. Over the course of the growing season (budbreak to June), the N content of the permanent structure (roots, trunk, and cordon wood) increased by 30.53 kg/ha, while 103.8 kg N/ha was absorbed.
The latter was equivalent to 4.01 kg N per ton of fresh grapes produced, being in good agreement with the value of 3.89 kg reported for Chenin blanc in sand culture (Conradie, 1980; Conradie, 2004). At Blouputs, little N (6.93 kg/ha) was absorbed from harvest to end February (Table 2), with leaves and shoots still accumulating N. Absorption of N decreased further during March and April, but appeared to accelerate during May and June. Despite an increase in dry mass, the permanent structure (roots, trunk, and hard wood) showed a small N loss (1.64 kg/ha) between budbreak and leaf-fall, mainly on account of the roots losing 7.74 kg N/ha. Fine-root death during the post-harvest root flush may have returned N to the soil, accounting for a proportion of the loss (Bates et al., 2002; Singha et al., 2020). Fine-root death may have occurred to a lesser extent for the better aerated and well-drained soil at Augrabies. Despite above-mentioned differences between the two localities, the amount of N absorbed at Blouputs for the production of one ton of fresh grapes (4.00 kg) was nearly identical to the value obtained at Augrabies.
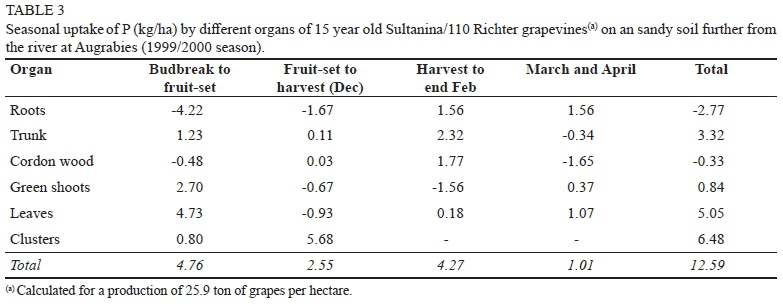
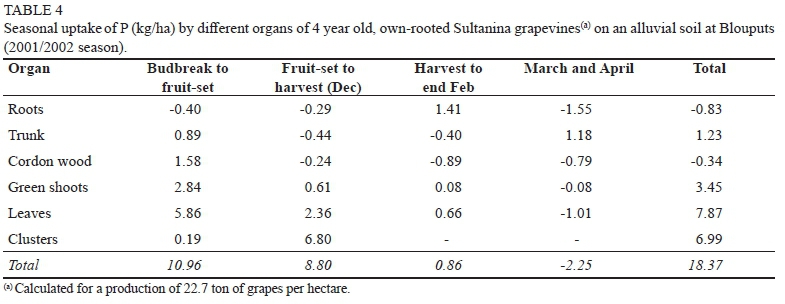
Phosphorus
As in the case of N, grapevines at Augrabies utilized more reserve-P between budbreak and fruit-set than those at Blouputs (Tables 3 & 4). From fruit-set to harvest, P was remobilized from roots, green shoots, and leaves in the Augrabies grapevines, while shoots and leaves still acted as P-sinks for the Blouputs grapevines. Consequently, the amount of soil-P absorbed between budbreak and harvest was considerably less at Augrabies (7.31 kg/ha) than at Blouputs (19.76 kg/ha). At harvest, leaves and shoots from the Blouputs grapevines contained twice as much P than Augrabies (Tables 3 & 4). This was not only on account of the higher dry mass (Figs. 1 & 2), but also on account of a higher soil-P level (data not shown). Clusters also contained less P at Augrabies (0.25 kg per ton of fresh grapes) than at Blouputs (0.31 kg per ton of fresh grapes). At Augrabies, P accumulated in the roots, trunk, and cordon wood between harvest and end February, resulting in a substantial amount of soil-P (4.27 kg/ha) being absorbed. Uptake of P was still noticeable during March and April for the Augrabies grapevines, even though rate of uptake slowed down considerably. In contrast to N, the permanent structure (roots, trunk, and hard wood) gained little P (0.22 kg/ha) over the course of the growing season. At Blouputs, little P (0.86 kg/ha) was absorbed from harvest to end February, while P was apparently lost during March and April. High P levels in the vegetative growth may have resulted in less P being absorbed during the post-harvest period. As already discussed, fine-root death during the post-harvest root flush may also have accounted for a proportion of the P loss. At Augrabies, 0.49 kg of P was utilized for the production of one ton of grapes, while a higher value of 0.81 kg was obtained at Blouputs. A value of 0.72 kg per ton of fresh grapes was obtained for pot-grown Chenin blanc (Conradie, 1981b, Conradie, 2004).
Potassium
Remobilization of K from the permanent structure appeared to be of minor consequence between budbreak and fruit-set at both localities (Tables 5 & 6). In the case of pot-grown grapevines, the contribution from K reserves was also relatively minor during this period (Conradie, 1981a; Schreiner & Scagel, 2006). At both localities, approximately 36 kg K/ha was absorbed between budbreak and fruit-set. From fruit-set to harvest, K was remobilized from the permanent structure (especially the trunk) at both localities. At Augrabies, K was also remobilized from shoots and leaves, being in agreement with results from some studies (Conradie, 1981a; Williams & Biscay, 1991; Schreiner & Scagel, 2006; Rogiers et al., 2017). At Blouputs, however, shoots and leaves were still accumulating K between fruit-set and harvest. This suggested that the re-translocation of K does not necessarily occur during the pre-harvest period, due to factors such as K availability, K uptake capacity of roots, continued vegetative growth, and canopy density (Mullins et al., 1992; Mpelasoka et al., 2003). Appreciably less soil-K (34.3 kg/ha) was absorbed between fruit-set and harvest at Augrabies than at Blouputs (45.6 kg/ha). Similar to P, clusters at Blouputs contained more K at harvest (2.31 kg per ton of fresh grapes) than at Augrabies (2.09 kg per ton of fresh grapes). At Augrabies, clusters accounted for 60.7% of the K in the current season's above-ground growth, in comparison to 54.0% at Blouputs. Comparable distributions have been reported (Lafon et al., 1965; Williams et al., 1987; Löhnertz, 1988; Mullins et al., 1992; Pradubsuk & Davenport, 2010). From harvest to end February, 16.78 kg K/ha was absorbed at Augrabies, mainly on account of a substantial amount of K gained by the trunk; however, at Blouputs, little soil-K (2.53 kg/ha) was absorbed. A loss of K occurred during March and April at both localities. At Augrabies, 3.20 kg of K was utilized for the production of one ton of grapes, while a higher value of 3.57 kg was obtained at Blouputs. This is in good agreement with the value of 3.05 kg reported for pot-grown Chenin blanc (Conradie, 1981b; Conradie, 2004).
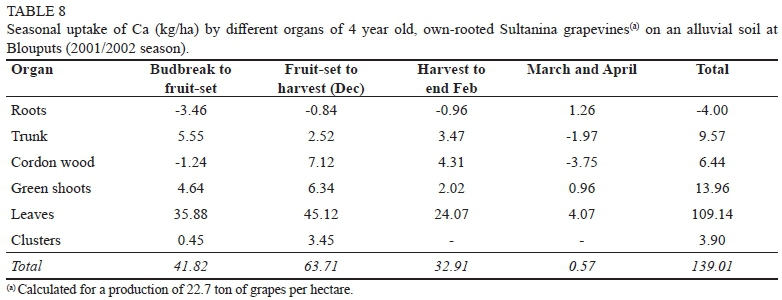
Calcium
At Augrabies, the Ca-content of roots showed a substantial increase from budbreak to harvest, followed by a major decrease from harvest to the end of April (Table 7). In view of Ca being a relatively immobile element (Mengel & Kirkby, 1979; Pradubsuk, 2008; Montanaro et al., 2010), this was unexpected. The high Ca-content of the specific soil (including the presence of free lime) most likely played a role as well. Even though roots were washed before chemical analysis, this could not be done too drastically, in order to prevent a loss in root mass. If root samples had been contaminated with a small soil fraction, the absolute values for N, P, K and Mg should not have been affected unduly, but this was not necessarily the case for Ca. The underground portion of the trunk may have been affected in the same way. Consequently, the total values in Table 7 have been calculated for above-ground organs only. At Blouputs, where grapevines were growing on a sandier soil, without free lime, the Ca-levels of under-ground organs did not show major declines over the course of the season (Table 8). At both localities, Ca was absorbed at the highest rate between fruit-set and harvest. The major share (56.6% & 76.9% for Augrabies and Blouputs, respectively) of the Ca absorbed up to harvest was incorporated into leaves. No remobilization of Ca from green shoots and leaves to clusters occurred. At harvest, leaves and shoots of the Augrabies grapevines contained only 33.4 kg Ca/ha, in comparison to 92.0 kg/ha at Blouputs. However, clusters from Augrabies and Blouputs contained a comparable amount of Ca (0.17 kg per ton of fresh grapes). An identical value was reported for Chenin blanc (Conradie, 1981a; Conradie, 1981b). Higher (Lafon et al., 1965; Mullins et al., 1992) and slightly lower (Schreiner & Scagel, 2006) Ca-levels have been reported for grapes from other regions or cultivars. Previous work has shown that cluster-Ca remains constant during the latter part of the pre-harvest period (Conradie, 1981a; Pradubsuk & Davenport, 2010). Since Ca is phloem immobile (Schreiner & Scagel, 2006; Montanaro et al., 2010), it is transported in the xylem only, and xylem in the berry is disrupted after veraison (Mpelasoka et al., 2003). Consequently, high Ca-levels in the leaves of the Blouputs grapevines appeared to have minimal effect on the Ca-content of clusters. During January and February, Ca was actively absorbed at both localities, with leaves still showing major gains. Absorption continued during March and April, albeit at a substantially slower rate. If it is assumed that the Ca-content of the roots and the trunk of the Augrabies grapevines remained relatively constant during the season, 2.07 kg Ca was utilized for the production of one ton of fresh grapes. This is in good agreement with the value (2.01 kg) obtained for young Chenin blanc (Conradie, 1981a, Conradie, 2004). At Blouputs, however, annual utilization was three times as high (6.12 kg/ton of grapes), largely on account of the high Ca-content of the leaves.
Magnesium
The seasonal uptake pattern of Mg was comparable at the two localities (Tables 9 & 10). It was absorbed at a fairly uniform rate between budbreak and harvest. In spite of Mg being a highly mobile element, re-translocation from the permanent structure appeared to be limited. At harvest, shoots and leaves at Augrabies contained less Mg (9.1 kg/ha) than at Blouputs (15.9 kg/ha). As with Ca, an identical amount of Mg was removed by the crop (0.09 kg/ton of grapes). Uptake continued at both localities during January and February, partly on account of leaves still gaining Mg. An apparent loss of Mg occurred at both localities during March and April, albeit of larger magnitude at Blouputs. The latter was largely on account of a substantial loss (3.08 kg/ha) from the roots. At Augrabies, the permanent organs gained 9.27 kg Mg/ha over the course of the season in comparison to negligible gain of 0.02 kg/ha at Blouputs. The amount of Mg utilized for the production of one ton of fresh grapes amounted to 0.82 kg and 0.85 kg for Augrabies and Blouputs, respectively. This is higher than the value (0.60 kg) obtained for young Chenin blanc (Conradie, 1981a; Conradie, 2004).
Nutrient uptake patterns of Augrabies and Blouputs
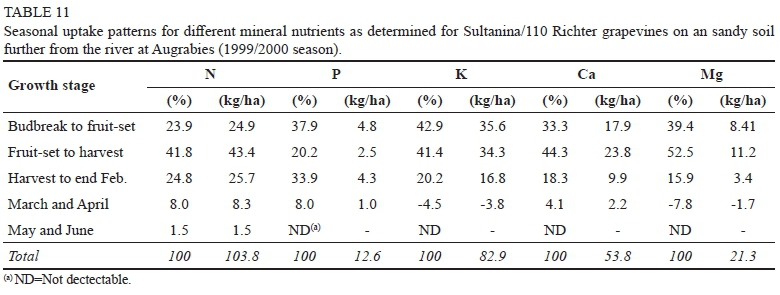
At Augrabies, where grapevines were not excessively vigorous, 23.9% of the annual N-requirement (Table 11) was absorbed between budbreak and fruit-set, with reserve-N playing a prominent role. The major fraction of the annual N-requirement (41.8%) was absorbed between fruit-set and harvest, with N also being relocated from the leaves. This pre-harvest pattern was largely comparable to previous results (Conradie, 2004; Schreiner, 2004; Masclaux-Daubresse et al., 2010; Ferrara et al. 2018). At Blouputs, where grapevines were growing more vigorously, reserve-N played a less prominent role between budbreak and fruit-set, resulting in the N-requirement (27.3% of seasonal total) mainly being supplied by means of soil uptake. Leaves were still a major sink for N between fruit-set and harvest, with 55.7% of the annual requirement being absorbed. Because entire grapevines were not scheduled for harvesting at veraison, N-uptake could not be quantified during the ripening period. At Augrabies, however, grapevines were erroneously sampled at veraison (data not shown). Results of the veraison analyses suggested that no more than 5% of the annual requirement was absorbed between veraison and harvest, being in agreement with previous studies (Conradie, 1980; Conradie, 1986; Araujo & Williams, 1988; Conradie, 2004; Ferrara et al. 2018). Because shoots and leaves of the Blouputs grapevines accumulated N up to the end of February (Table 2), it is unlikely that soil-uptake of N would have slowed down between veraison and harvest. This is in agreement with results from other studies (Hanson & Howell, 1995), where up to 33% of the annual N requirement was accumulated between veraison and harvest. The vigorous grapevines at Blouputs were possibly accumulating N in shoots and leaves during the ripening period, to the detriment of the clusters.
At Augrabies, 25.7% of the annual N-requirement was absorbed from harvest to end February (Table 1). From end February to the start of leaf senescence (end April), only 8.3% of the annual N requirement was accumulated, while absorption was even less (1.5%) during May and June (end leaf-fall). In spite of the grapevines at Augrabies being harvested in December already, the relative importance of N-uptake during the post-harvest period (34.3% of annual requirement) appeared to be nearly identical to the value (34.5% of annual requirement) obtained for young grapevines in the Western Cape (Conradie, 2004). Some previous studies (Kliewer, 1971; Zapata et al., 2001; Schreiner, 2004) suggested that results obtained for young grapevines, grown under a constant high supply of nutrients, will not necessarily be comparable to those from field surveys. The Augrabies results suggest that the post-harvest uptake of N is dictated by climate/length of the post-harvest period, and not by the N-supply of the growth medium. However, at Blouputs (more vigorous grapevines), only 7.6% of annual N-requirement was absorbed from harvest to the end of February (Table 2), decreasing to 2.9% during March and April, and increasing again to 6.5% during May and June (end leaf-fall). The N fraction accumulated during the post-harvest period (17.0% of annual requirement) was only half as high as at Augrabies. The fact that N was accumulated less effectively in the Blouputs grapevines during the post-harvest period may have been partly responsible for reserve-N being less readily utilized between budbreak and fruit-set (Table 2). On account of the root system at Blouputs being relatively small compared to Augrabies, fine-root death may also have played a more important role at Blouputs.
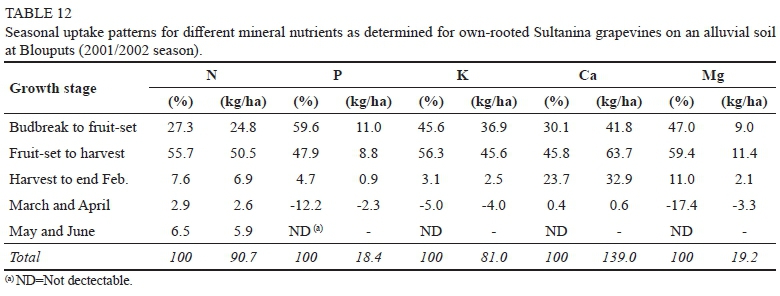
At Augrabies, 37.9% of the annual P-requirement was absorbed between budbreak and fruit-set, followed by 20.2% between fruit-set and harvest (Table 11). Values of 31.9% and 40.6% were reported for Chenin blanc in sand culture, respectively (Conradie, 1981; Conradie, 2004). Active uptake during January and February accounted for 33.9% of annual the P-requirement, followed by 8.0% during March and April. In comparison to the situation in the Western Cape, where 27.5% of the annual P-requirement is absorbed during the post-harvest period (Conradie, 2004), more P appeared to be absorbed after harvest at Augrabies. At Blouputs, the major portion of the annual P-requirement (59.6%) was absorbed between budbreak and fruit-set (Table 12). Even though P-uptake was noticeable (4.7%) during January and February, a substantial loss (12.2%) occurred during March and April.
Apart from an apparent loss of K during March and April, the K-uptake pattern at Augrabies (Table 11), was comparable to that of pot-grown Chenin blanc (Conradie, 1981b; Conradie, 2004). The field-grown grapevines absorbed 15.7% of their annual K requirement during the post-harvest period, while a similar fraction (15.2%) was reported for Chenin blanc. At Blouputs, the major share of the annual K-requirement (56.3%), was absorbed from fruit-set to harvest (Table 12). As in the case of P, a net loss (1.9%) occurred during the post-harvest period.
Even though grapevines at the two localities utilized vastly different amounts of Ca (Tables 8 & 9), comparable fractions were absorbed during the different growth stages (Tables 11 & 12). Approximately 32% of the annual requirement was accumulated from budbreak to fruit-set, followed by a further 45% up to harvest and 23% after harvest. A comparable fraction (21.5%) was absorbed during the post-harvest period by Chenin blanc (Conradie, 2004).
The seasonal uptake pattern of Mg was comparable up to harvest (Tables 11 & 12). During the post-harvest period, however, grapevines at Augrabies showed a net gain (8.1% of annual requirement), in comparison to a net loss (6.4% of annual requirement) at Blouputs. A higher fraction (21.5%) was absorbed during the post-harvest period by pot-grown Chenin blanc (Conradie, 2004).
CONCLUSIONS
Seasonal uptake patterns of N, P, K and Ca are similar where grapevines are not growing too vigorously in a long-season growing area, such as the Orange River Region, compared to cooler growing areas. For Mg, uptake during post-harvest period appears to be less than in cooler areas. During the long post-harvest period, nutrient uptake occured at the fastest rate during the two months immediately after harvest. On account of nutrient reserves being replenished during the post-harvest period, grapevines should be fertilized (where necessary) shortly after harvest. The annual N, P, and K requirements of well-balanced grapevines should be comparable for long-season areas and areas with shorter growing seasons.
Seasonal uptake patterns may be different for overly vigorous grapevines (usually on alluvial soils) in long-season areas. Mineral nutrients continue to be accumulated in vegetative organs (shoots & leaves) during the period immediately after harvest. In comparison to more well-balanced plants, only half as much N may be absorbed during the post-harvest period, while P, K, and Mg may be lost. One should bear in mind, however, that amounts of N, P, K, and Mg absorbed during the post-harvest period may be underestimated in field trials on account of fine-root death. The latter seems to be a more important factor on sandier, alluvial soils than on gravelly, well-aerated soils. For Ca, where root-reserves play a less important role, the post-harvest uptake pattern seems to be unaffected by grapevine vigour/soil type. Even though the annual N, P, and K requirements of vigorous grapevines in long-season growing areas will be comparable to those of more well-balanced plants, fertilizer applications should be timed differently. In order to prevent vigorous growth during the post-harvest period, N should not be applied during the first two months after harvest.
LITERATURE CITED
Alexander, D.M., 1958. Seasonal fluctuations in the nitrogen content of the Sultana vine. Aust. J. Agric. Res. 8, 162-178. [ Links ]
Araujo, F.J. & Williams, L.E., 1988. Dry matter and nitrogen partitioning and root growth of young field-grown Thompson Seedless grapevines. Vitis 27, 21-32. [ Links ]
Barnuud, N.N., Zerihun, A., Gibberd, M. & Bates, B., 2014. Berry composition and climate: responses and empirical models. Int. J. Biometeorol. 58, 1207-1223. [ Links ]
Bates, T.R., Dunst, R.M. & Joy, P., 2002. Seasonal dry matter, starch and nutrient distribution in 'Concord' grapevine roots. Hort. Sci. 37, 313-316. [ Links ]
Campbell, C.R., 1991. Determination of total nitrogen in plant tissue by combustion. In: Plant analysis reference procedures for the southern region of the United States. C.O. Plank (Ed.). pp. 21-23. Southern Coop. Ser. Bul. 368. Univ. of Georgia, Athens. [ Links ]
Conradie, W.J., 1980. Seasonal uptake of nutrients by Chenin blanc in sand culture. I. Nitrogen. S. Afr. J. Enol. Vitic. 1, 59-65. [ Links ]
Conradie, W.J., 1981a. Seasonal uptake of nutrients by Chenin blanc in sand culture. II. Phosphorus, Potassium, Calcium and Magnesium. S. Afr. J. Enol. Vitic. 2, 7-13. [ Links ]
Conradie, W.J., 1981b. Nutrient consumption by Chenin blanc grown in sand culture and seasonal changes in the chemical composition of leaf blades and petioles. S. Afr. J. Enol. Vitic. 2, 15-18. [ Links ]
Conradie, W.J., 1986. Utilisation of nitrogen by the grapevine as affected by time of application and soil type. S. Afr. J. Enol. Vitic. 7, 76-82. [ Links ]
Conradie, W.J., 1990. Distribution and translocation of nitrogen absorbed during late spring by two-year-old grapevines grown in sand culture. Am. J. Enol. Vitic. 41, 241-250. [ Links ]
Conradie, W.J., 1991a. Translocation and storage of nitrogen by grapevines as affected by time of application. In: Proc. Int. Symposium on Nitrogen in Grapes and Wine. J.M. Rantz (Ed.), pp. 32-42. American Society of Enology and Viticulture, Davis, CA. [ Links ]
Conradie, W.J., 1991b. Distribution and translocation of nitrogen absorbed during early summer by two-year-old grapevines in sand culture. Am. J. Enol. Vitic. 42, 180-190. [ Links ]
Conradie, W.J., 1992. Partitioning of nitrogen in grapevines during autumn and the utilisation of nitrogen reserves during the following growing season. S. Afr. J. Enol. Vitic. 13, 45-51. [ Links ]
Conradie, W.J. 2004. Partitioning of mineral nutrients and timing of fertilizer applications for optimum efficiency. In: Proc. Symposium on Soil Environment and Vine Mineral Nutrition. [ Links ]
Ferrara, G., Malerba, A.D., Matarrese, A.M.S., Mondelli, D. & Mazzeo, A., 2018. Nitrogen distribution in annual growth of 'Italia' table grape vines. Front. Plant Sci. 9, 1-18. [ Links ]
Grechi, I, Vivin, Ph., Hilbert, G., Milin, S., Robert, T. & Gaudillere, J.-P., 2007. Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Env. Exp. Bot. 59, 139-149. [ Links ]
Hanson, E.J. & Howell, G.S., 1995. Nitrogen accumulation and fertilizer use efficiency by grapevines in short-season growing areas. Hort. Sci. 30, 504-507. [ Links ]
Holzapfel, B.P. & Treeby, M.T., 2007. Effects of timing and rate of N supply on leaf nitrogen status, grape yield and juice composition from Shiraz grapevines grafted to one of three different rootstocks. Aust. J. Grape Wine Res. 13, 14-22. [ Links ]
Keller, M. & Koblet, W., 1995. Dry matter and leaf area partitioning, bud fertility and second-season growth of Vitis vinifera L.: Responses to nitrogen supply and limiting irradiance. Vitis 34, 77-83. [ Links ]
Kliewer, W.M., 1971. Effect of nitrogen on growth and composition of fruits from 'Thompson Seedless' grapevines. J. Am. Soc. Hortic. Sci. 96, 816-819. [ Links ]
Kodur, S., Tisdall, J.M., Tang, C. & Walker, R.R., 2010. Accumulation of potassium in grapevine rootstocks (Vitis) as affected by dry matter partitioning, root traits and transpiration. Aust. Soc. Vitic. Oenol. Inc. 16, 273-282. [ Links ]
Lafon, J., Couillard, P. & Gay-Bellile, F., 1965. Rythme de l'absorption minérale de la vigne au cours d'un cycle vegetatif. Vignes Vins 140, 17-21. [ Links ]
Leão, P.C. & Chaves, A.R.. 2020. Agronomic responses of grapevine 'Chenin Blanc' as a function of training systems and rootstocks. Sci. Agric. 78, 1-7. [ Links ]
Lõhnertz, O. 1988., Nãhrstoffelementaufname von Reben im Verlauf eines Vegetationszyklus. Mitt. Klosterneuburg 38, 124-129. [ Links ]
Lõhnertz, O. 1991., Soil nitrogen and the uptake of nitrogen in grapevines. In: Proceedings of the International Symposium on Nitrogen in Grapes and Wines. Rantz, J.M. (Ed.). pp. 1-11. American Society of Enology and Viticulture, Davis, CA. [ Links ]
Marocke, R., Balthazard, J. & Correge, G., 1976. Exportations en éléments fertilisants des principaux cépages cultivés en Alsace. Acad. d'Agiculture de France. pp. 420-429. [ Links ]
Masclaux-Daubresse, C., Daniel-Vedele, F., Dechorgnat, J., Chardon, F., Gaufichon, L. & Suzuki, A., 2010. Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Ann. Bot. 105, 1141-1157. [ Links ]
Mártensson, A., 2020. Nutrient Balance and Fertilizer Use Efficiency in Swedish Vineyards. Int. J. Wine Res. 12, 17-21. [ Links ]
Mengel, K. & Kirkby, E.A., 1979. Principles of Plant Nutrition (2nd ed.). International Potash Institute, Worblaufen-Bern, Switzerland. [ Links ]
Montanaro, G., Dichio, B. & Xiloyannis, C., 2010. Significance of fruit transpiration on calcium nutrition in developing apricot fruit. J. Plant Nutr. Soil Sci. 173, 618-622. [ Links ]
Mpelasoka, B.S., Schachtmann, D.P., Treeby, M.T. & Thomas, M.R., 2003. A review of potassium nutrition in grapevines with special emphasis on berry accumulation. Aust. J. Grape Wine Res. 9, 154-168. [ Links ]
Mullins, M.G., Bouquet, A. & Williams, L.E., 1992. Biology of the grapevine. Cambridge University Press, Cambridge, UK. [ Links ]
Pradubsuk, S., 2008. Uptake and partitioning of mineral nutrients in concord grape. Thesis, Washington State University, 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, MI 48106-1346. [ Links ]
Pradubsuk, S. & Davenport, J.R., 2010. Seasonal uptake and partitioning of macronutrients in mature 'Concord' grape. J. Am. Soc. Hortic. Sci.135, 474-483. [ Links ]
Rogiers, S., Coetzee. Z., Walker, R., Deloire, A. & Teyerman, S.D., 2017. Potassium in the grape (Vitis vinifera L.) berry: Transport and function. Front. Plant Sci. 8, 1-19. [ Links ]
Schreiner, P.S., 2004. Mycorrhizas and mineral acquisition in grapevines. In: Proc. Symposium on Soil Environment and Vine Mineral Nutrition. Christensen, L.P. & Smart, D.R. (Eds.). pp. 49-60. American Society of Enology and Viticulture, Davis, CA. [ Links ]
Schreiner, R.P. & Scagel, C.F., 2006. Nutrient uptake and distribution in a mature 'Pinot noir' vineyard. Hort. Sci. 41, 336-345. [ Links ]
Singha, D., Brearley. F.Q. & Tripathi, S.K., 2020. Fine root and soil nitrogen dynamics during stand development following shifting agriculture in Northeast India. Forests. 11, 1-12. [ Links ]
Soil Classification Working Group., 1991. Soil classification - A taxonomic system for South Africa. Memoirs on natural resources of South Africa No. 15. Dept. Agric. Development., Pretoria. [ Links ]
Tomasi, N., Monte, R., Varanini, Z., Cesco, S. & Pinton, R., 2015. Induction of nitrate uptake in Sauvignon Blanc and Chardonnay grapevines depends on the scion and is affected by the rootstock. Aust. J. Grape Wine Res. 21, 331-338. [ Links ]
Vos, R.J., 2003. Effect of the timing of nitrogen application on soil nitrogen and use efficiency of Vitis labrusca in a short-season region. Thesis, Michigan State University, 300 North Zeeb Road P.O. Box 1346, Ann Arbor, Ml 48106-1346. [ Links ]
Wheeler, S.J. & Pickering, G.J., 2003. Optimizing grape quality through soil management practices. Food, Agric. Environ. 1, 190-197. [ Links ]
Williams, L.E., 1991. Vine nitrogen requirements - utilisation of N sources from soils, fertilisers, and reserves. In: Proc. Int. Symposium on Nitrogen in Grapes and Wine. Rantz, J.M. (Ed.). pp. 62-66. American Society of Enology and Viticulture, Davis, CA. [ Links ]
Wiliams, L.E. & Biscay, P.J., 1991. Partitioning of dry weight, nitrogen and potassium in Cabernet Sauvignon grapevines from anthesis until harvest. Am. J. Enol. Vitic. 42, 113-117. [ Links ]
Williams, L.E., Biscay, P. & Smith, R.J., 1987. Effect of interior canopy defoliation on berry composition and potassium distribution in Thompson Seedless grapevines. Am. J. Enol. Vitic. 38, 287-292. [ Links ]
Winkler, A.J., 1962. General Viticulture. University of California Press, Los Angeles. [ Links ]
Zapata, C., Magne, C., Deleens, E., Brun, O., Audran, J.C. & Chaillou, S., 2001. Grapevine culture in trenches: root growth and dry matter partitioning. Aust. J. Grape Wine Res. 7, 127-131. [ Links ]
Zeeman, A.S. 1981. Oplei. In: Wingerdbou in Suid-Afrika. Burger, J.D. & Deist, J. (eds). Nietvoorbij, South Africa. pp. 185-201. [ Links ]
Submitted for publication: November 2021
Accepted for publication: March 2022
# Fruit, Vine & Wine Institute of the Agricultural Research Council
* Corresponding author: pjraath@sun.ac.za
Acknowledgements: The ARC for infrastructure and other resources, Hortgro Science for partial funding as well as the Soil and Water Science personnel at ARC Infruitec-Nietvoorbij for technical support


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}