










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.43 n.1 Stellenbosch 2022
http://dx.doi.org/10.21548/43-1-4589
ARTICLES
Insecticide Efficacy Against Trimen's False Tiger Moth, Agoma trimenii (Lepidoptera: Agaristidae)
C. MonisI; A.P. MalanI; J.Y. De WaalII; S.A. JohnsonI,*
IDepartment of Conservation Ecology and Entomology, Faculty of AgriSciences, Stellenbosch University, Private Bag X1 Matieland (Stellenbosch), South Africa
IICorteva Agriscience, Block A, 2nd Floor, Lakefield Office Park, 272 West Avenue De Howes, Centurion, Gauteng 0157, South Africa
ABSTRACT
Trimen's false tiger moth, Agoma trimenii (Lepidoptera: Agaristidae), has developed pest status in vineyards in the Northern Cape and Groblersdal areas of South Africa, and an integrated pest management system is required. The objective of this study was to test the susceptibility of A. trimenii larvae to three commercial products (Delegate®WG, Steward®150 EC and three doses of DiPel® DF). Bioassay tests using all three products, and semi-field trials to test the potential of DiPel® DF against A. trimenii larvae, applied at different water volumes (50 g/1 000 L/ha and 50 g/1 430 L/ha) were performed. The residual activity of DiPel® DF, when applied at different water volumes, was investigated daily. Delegate® WG, Steward®150 EC and the recommended dose of DiPel® DF showed 100% larval mortality within seven days. Delegate® WG and the recommended dose of DiPel® DF proved to be the fastest acting products. The product label recommended dose of DiPel® DF (0.25 g/500 mL distilled water) proved the most effective dose (in comparison to halved and doubled dosages) and showed 100% mortality five days after application. Increasing the water volume per ha of a spray application of DiPel® DF for the same application area, showed no significant increase in larval mortality. A reduction in insecticidal activity for DiPel® DF applied at both water volumes was seen between leaves picked four days after spraying and leaves picked five days after spraying, and no mortality was observed after day 6. To help improve efficacy, attention should be given to increasing spray coverage and residual activity of DiPel® DF, as well as using all tested products within an integrated pest management system.
Key words: Agoma trimenii, Trimen's false tiger moth, insecticide, persistence, spray coverage, grapevine
INTRODUCTION
Trimen's false tiger moth, Agoma trimenii Felder (Lepidoptera: Agaristidae), is an indigenous lepidopteran that is classified within the Noctuidae family and the subfamily Agaristidae (De Prins & De Prins, 2021). The moth recently developed pest status in parts of South Africa, after reports of moth infestations in vineyards in summer rainfall areas (Pretorius et al., 2012; Morris, 2019). The feeding of the larvae of A. trimenii is described as being host-specific, mainly on grapevine and wild host in the family Vitaceae (Picker et al., 2002; De Prins & De Prins, 2012). Agoma trimenii is an agricultural pest, targeting the young shoots and leaves of grapevine in the Northern Cape and Limpopo provinces of South Africa (Morris et al., 2020a). The voracious feeding of the larvae on the leaves of grapevine has previously been reported by Pretorius et al. (2012). As the prevalence of A. trimenii in these grape-producing regions of South Africa increases, so does its impact on production, by defoliation, and therefore the need to control it.
During October and March, adult moths, black in colour with striking cream and orange spots (± 54 mm) are found in vineyards. Females deposit their eggs singularly on the leaf surfaces, where the eggs hatch after three days. The newly hatched larvae (> 0.5 mm) display a cream to white colour, with the final instars (± 4 mm) being characterised by black and yellow markings (Pretorius et al, 2012). Morris et al. (2020a) reported that larvae feed on shoots and leaves throughout the growing season, causing extensive damage to the vines. At the end of the growing season, in March, final instar larvae move via a silken thread to the soil, where the pre-pupae develop into pupae (± 25 mm). The fully formed pupae have a hardened cuticle and are dark brown/red in colour. Open holes (± 3 mm) are visible in the soil (depth of ± 4 cm), which protect the pupae (Morris et al, 2020a).
Integrated pest management (IPM) strategies that incorporate various control measures are best practice for effective and sustainable pest management (Abrol & Shankar, 2012). Morris et al. (2020a) discuss potential cultural control measures that consider the pest's seasonal biology. Biological control agents such as entomopathogenic nematodes (EPNs) and fungi (EPF) have been tested against the larvae and pupae of A. trimenii. Results from these preliminary studies indicate that larvae are highly susceptible, whilst pupae are totally resistant to EPNs and EPF (Morris et al., 2020b) infection. Other potential biological control strategies, which are specifically pathogenic to the larval stages of Lepidoptera, include the application of formulated biologically-derived insecticides, such as Delegate® WG [containing the active ingredient, spinetoram, a derived from the fermentation of Saccharopolyspora spinosa Mertz & Yao (Actinomycetales: Pseudonocardiaceae), a naturally occurring soil bacterium] and DiPel® DF [a 5t-based insecticide formulated from Bacillus thuringiensis var. kurstaki (Bacillales: Bacillaceae)]. Biological insecticidal derivatives have beneficial traits, which include minimal risk to human health, as well as beneficial and other non-target insects, the absence of toxic residues in crops and host specificity (Inglis et al., 2001; Goettel et al., 2005).
Delegate®WG boasts a broad insecticidal spectrum, especially against all growth stages of tortricid pests, including codling moth, Cydia pomonella L., light brown apple moth, Epiphyas postvittana (Walker) and oriental fruit moth, Grapholita molesta (Busck) (Dripps et al., 2008; Magalhaes & Walgenbach, 2011; Sial et al., 2011). Its mode of action may be either directly through contact with the body surface (contact toxicity) or by ingestion (Shimkawatoko et al., 2012). Due to its unique mode of action, Delegate® WG could be an exceptional rotational product for use in an IPM programme (Bacci et al., 2016).
DiPel® DF is characterised by numerous crucial attributes that favour its application in a pest management programme, namely its nontoxic nature to both plants and vertebrates, and its relatively specific action on target insect species (Roditakis, 1986). Mortality of infected insects usually occurs within two to three days (Waites et al., 2009). DiPel® DF has proven to be a successful biological control agent against two lepidopteran pest species present in South Africa: the diamondback moth, Plutella xylostella L. (Plutellidae), which is highly resistant to conventional insecticides, as well as the cabbage moth, Mamestra brassicae L. (Noctuidae) (Tabashnik et al., 1990; Devetak et al., 2010).
The insect growth regulator, Steward® 150 EC, can be classified as an insecticide that disrupts crucial physiological functions associated with the life cycle of insect development and metamorphosis (Sanchez-Bayo, 2012). Steward®150 EC acts as a contact and stomach insecticide (Wing et al., 2000). Its novel mode of action of inhibiting sodium entry into nerve cells, results in paralysis (inhibited feeding) and death of the target pest within three to five days (Dinter & Wiles, 2000). With its unique mode of action and its minimal impact on beneficial parasitoid and predatory insects, it has great potential as a partner in IPM programmes (Nowak et al., 2001).
The use of the two environmentally-friendly biologically derived pesticides, as well as a 'softer' pure chemical product for the control of A. trimenii, requires knowledge of their susceptibility, prior to application on a large scale. Bioassays are the starting point for any control investigation in which virulence is important, as they allow for the removal of factors that could reduce virulence towards the target host (Coombes, 2012). This study investigated the potential of less harsh insecticides such as Delegate® WG, DiPel® DF and Steward®150 EC to control A. trimenii and determined their feasibility for use as part of an IPM program directed at the management of the pest. Bioassays were performed against the larval stages of A. trimenii, to determine the potential of each pesticide as a control agent against this stage of the insect. Furthermore, semi-field trials in table grape vineyards were performed to test the potential of DiPel® DF against A. trimenii larvae, applied at different water volumes per ha spray coverage, and the residual activity of DiPel® DF after application and ingestion by larvae.
MATERIALS AND METHODS
Source of larvae
Larvae of A. trimenii were collected from the untreated demarcated field site on a commercial table grape farm in the Northern Cape province, South Africa. Different larval instars were hand collected and used on the day of collection or soon after in November, 2017 and February, 2018. Larvae ranged from 1.5 - 4.5 cm in length, encompassing different larval instar stages. The larvae were stored in plastic 2 L containers, provided with a mesh-covered hole in the lid to ensure adequate ventilation. The treatment containers were stored in conditions similar to that of their natural environment.
Source of Delegate®WG, Steward®150 EC and DiPel® DF
The commercial products Delegate® WG (Dow AgroSciences), Steward® 150 EC (Du Pont) and DiPel® DF (Philagro) were obtained from South African distributers and applied at the recommended dosages for lepidopteran larvae, as prescribed on the product label (Table 1). Additionally, DiPel® DF was applied at half the recommended dose and double the recommended dose against A. trimenii.
Field sites
Semi-field trials were carried out on a commercial table grape production farm in the Northern Cape province of South Africa, with high infestation levels of A. trimenii, during the growing season, over a seven-day period in November, 2017 and February, 2018. The trials were conducted on two field sites (A and B), each site comprised of a netted block of the table grape, Thompson Seedless. The vines followed a double-gable trellising system, with plant spacing of 1.8 m, and row width of 3.3 m. For both field sites, four rows of 20 vines were demarcated and left untreated by pesticides.
Bioassays
Two bioassay trials with Delegate® WG, Steward®150 EC and DiPel® DF against the larval stages of A. trimenii were conducted, one during November, 2017 and another during February 2018. Three to five fresh, untreated, mature vine leaves still attached to their stems were lightly coated by using a hand-held spray bottle, until before drip, at the recommended concentration of Delegate® WG (0.05 g/500 mL) and Steward®150 EC (0.2 mL/ 500 mL), and three different doses of DiPel® DF [Treatment 1 (TMT1): half the recommended dosage = 0.125 g/500 mL, Treatment 2 (TMT2): recommended dosage = 0.25 g/500 mL and Treatment 3 (TMT3): double recommended dosage = 0.5 g/500 mL in distilled water]. The leaves were left to dry for ± 20 min and were placed into 2-L plastic ice cream containers according to the product tested and lined with moistenend paper towels to keep the humidity in the containers high. Larvae (12) were added to the respective containers, ventilated with a mesh-covered hole in the lid. For each treatment, a control with 12 larvae was also prepared on the day of screening. Leaves were sprayed with distilled water and dried before being placed into 2-L plastic containers with larvae. A total of six plastic containers were used for each insecticide, plus the control treatments for each, on the different test dates, with 30 containers and 72 A. trimenii larvae used for each trial. All plastic containers were kept outside under natural conditions. Effectivity was assessed after application by recording mortality of larvae every day, for a period of seven days and inspecting each larva for movement and change in physical appearance.
Field application and efficacy assessment of DiPel® DF
Field trials with DiPel® DF, were performed in November, 2017 and repeated on February, 2018 at two untreated sites. DiPel® DF was applied in the field with a Cima T45 model centrifugal sprayer (1000 to 1 200 L) at the recommended concentration, 50 g/ha, as two separate treatments of different water volumes (Volume A - site A: 50 g/1 000 L/ha and Volume B - site B: 50 g /1 430 L/ha), i.e., increasing only the water volume, while keeping a constant concentration of DiPel® DF. Trials commenced in the early morning (07h00) to avoid too high levels of UV radiation. With a Cima centrifugal sprayer, treatments were applied to both sites, between each of the four rows of vines at a speed of 3.6 km/h.
After application, leaves from the top and bottom of the vines from the middle row of both treatment sites were collected daily, at the same time (08h00), for seven days. Ten young leaves were collected for each top and bottom container per treatment (320 leaves in total). The top and bottom leaves, for each day, were kept separate in two plastic 2-L containers, lined with wet paper towels to maintain high humidity. Fresh field-collected larvae (12) were then added to each container and left to feed on the treated leaves. A control in a 2-L plastic container was prepared on each day with leaves picked from untreated vines, sprayed with distilled water. Mesh-covered holes in the lids of the containers ensured adequate ventilation. All treatments and control plastic containers were kept outside under natural conditions. Each collection of leaves, picked daily for seven days, received 12 fresh larvae on the day of collection and larval mortality was recorded per container after 24 h of feeding for a period of seven days, i.e., the first collection of leaves with added larvae, had seven days of recorded observations. A total 336 larvae were used in the field trials.
Larval mortality was used to compare the effectiveness of the two water volumes, and the spray coverage between top and bottom leaves for each water volume application. Residual activity (activity of the product on leaves so many days after application) was presented by displaying the decrease in larval mortality for the two water volumes of DiPel® DF over time. Larval mortality was recorded after 24 h of feeding on the treated leaves for each day after spraying.
Statistical analyses
All statistical analyses were done using STATISTICA version 13 (StatSoft Inc. 2016). All data were normally distributed. Control mortality was zero in all cases, therefore there was no need for control correction of mortality data. The pathogenicity of Delegate® WG, Steward® 150 EC and DiPel® DF against larvae was analyzed by using repeated measures analysis of variance (ANOVA) of mortality over time, for each bioassay of Delegate® WG, Steward®150 EC and DiPel® DF. A post-hoc comparison of means was done by using the Fisher LSD test. Effectivity of DiPel® DF applied at different water volumes and subsequent spray coverage was analyzed using a two-way ANOVA for larval mortality due to the two water volumes of DiPel® DF, and between top and bottom leaves. A one-way ANOVA was conducted to compare the means of larval mortality of the two water volumes, as well as the means of larval mortality from top and bottom leaves. To determine the residual activity, DiPel® DF was applied at different water volumes of the three doses of DiPel® DF on picked leaves over the 7-day period, while repeated-measures ANOVA was conducted to compare larval mortality between consecutive days from which leaves were sprayed. Graphs depicted using descriptive statistics in Excel.
RESULTS
Pathogenicity of Delegate® WG, Steward® 150 EC and DiPel® DF against larvae
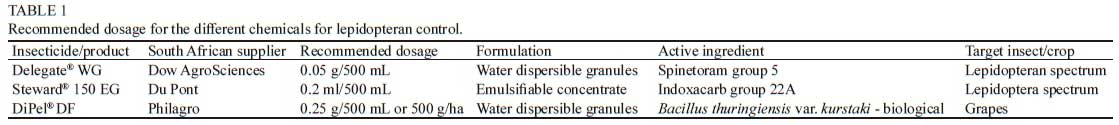
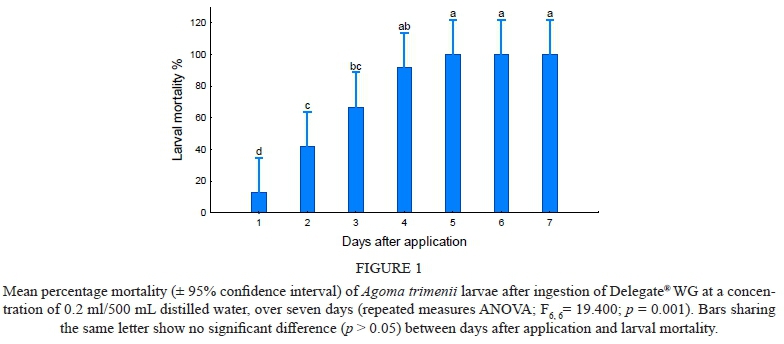
As no significant differences were found between the main effects of date and treatment, the data from the two trials were pooled before analysis. In the treated groups, mortality increased significantly over the seven-day period for Delegate® WG (F66 = 19.400; p = 0.001) with 100% mortality after seven days (Fig. 1 and 2). Larval mortality due to Delegate® WG increased at a faster rate than that of Steward®150 EC. By day four of the bioassays, larval mortality after treatment with Delegate® WG was 91.67% ± 9% and reached 100% by day five, compared to Steward®150 EC that resulted in 66.67% ± 20% by day four and reached 100% by day six. Mortality due to Delegate® WG, revealed that a significant increase in larval mortality occurred between the first and second day after ingestion (p = 0.04), compared to other consecutive days (Fig. 1). Data analysed showed no significant difference between consecutive day comparisons for mortality due to Steward®150 EC (p > 0.05).
For bioassays involving the three different doses of DiPel® DF, control groups for all three treatment bioassays showed no mortality over the seven-day period. Larval mortality due to the three doses of DiPel® DF over the seven-day period is presented in Fig. 3. By day four of the bioassay, mortality after applying the recommended dose (TMT2) was 83.3% ± 12% and reached 100% by day 5, compared to the half-recommended dose (TMT1), which resulted in 75% ± 12% by day 4, and remained unchanged. The double recommended dose (TMT3), resulted in 95.8% ± 12% mortality by day three and 100% by day four. Data analysed after the 7-day period showed no significant difference in total larval mortality between the means of each treatment (F12,18 = 0.843; p > 0.05). However, a pairwise comparison of larval mortality across all three treatments per day, using the Fisher LSD (minimum mean difference = 41.67), showed a significant difference between TMT1 and TMT3 on the third day of the bioassay trial (p = 0.025).
All tested insecticides resulted in 100% larval mortality within the 7-day test periods. Delegate® WG and the recommended dose of DiPel® DF achieved 100% larval mortality by day 5, while Steward®150 EC required one more day, and when the recommended dose of DiPel® DF was doubled, 100% mortality was achieved by day 4.
Field trials
Effectivity of DiPel® DF applied at different water volumes and subsequent spray coverage
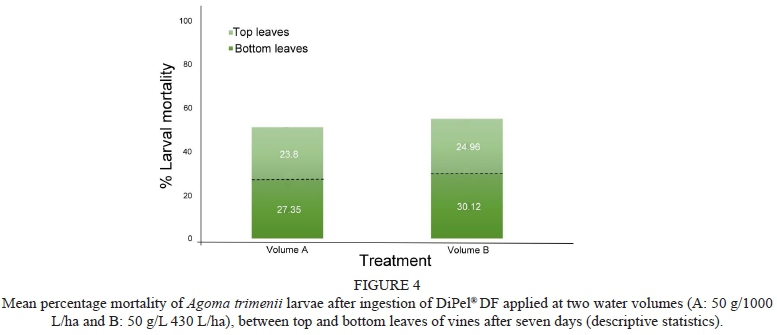
Total larval mortality (n = 168) was compared between the two water volume applications of DiPel® DF for the seven-day period, as well as larval mortality between the top and bottom leaves (Fig. 4). Water volume B (42 g/1 200 L/ha) resulted in higher larval mortality for combined top and bottom leaves (55% ± 5%), compared to larval mortality of water volume A (50 g/1 000 L/ha) for both top and bottom leaves (51% ± 5%), but not significantly so (F14 = 3.108, p > 0.05). For both water volumes A and B, higher larval mortality was seen on leaves picked from the bottom of the vine (27.35% ± 4% and 30.12% ± 4% respectively), compared to the top leaves (23.8% ± 4% and 24.96% ± 4% respectively). This was significantly higher for water volume B (p = 0.037), but not, however, statistically significant for water volume A.
Residual activity of DiPel® DF applied at different water volumes
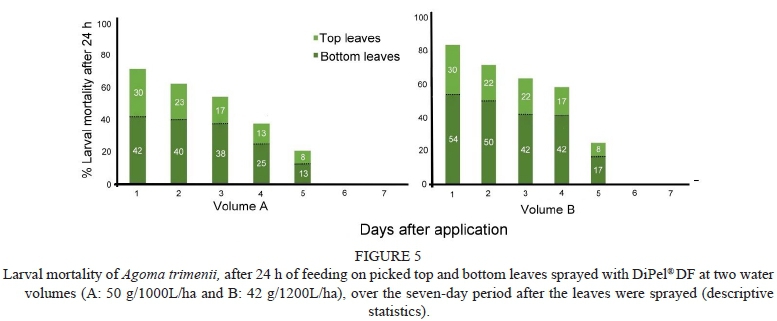
Residual activity of DiPel® DF applied at both water volumes, A and B, is presented by displaying the decrease in larval mortality after 24 h of feeding on picked top and bottom leaves, from one to seven days after the leaves were sprayed (Fig. 5). For both water volumes, larval mortality was higher on picked bottom leaves compared to top leaves. Larval mortality (n = 81) due to water volume B was higher than that of water volume A (n = 67), although not significantly different (p > 0.05). For both water volumes A and B, it appeared that larval mortality showed the notably decrease from day four (38% ± 5% and 59% ± 5%, respectively) to five (21% ± 5% and 25% ± 5%, respectively) after spraying, for combined top and bottom leaves and that by day 6, no larval mortality was recorded.
For water volume B, a pairwise comparison of larval mortality between consecutive days using the Fisher LSD (minimum mean difference = 37.50), revealed that notably decreased on the picked bottom leaves four to five days after spraying (p = 0.0054), however for water volume A, no significant differences (p > 0.05) were established between consecutive days for picked top leaves or picked bottom leaves.
DISCUSSION
Results from bioassays showed that both Delegate® WG and Steward® 150 EC were effective against various larval stages of A. trimenii. The effect of Delegate® WG onA. trimenii larvae was faster than that of Steward®150 EC. In both bioassays, infected larvae stopped feeding on leaves, displayed paralysis and developed curved and discoloured bodies. Infected larvae also expelled a green liquid from the mouth. Other infected larvae stopped feeding and remained alive for several days before they died. The results from the bioassays are in line with a study conducted by Abbas et al. (2015), who reported that in semi-field trials, Delegate® WG showed relatively better persistence and contact mortality of cotton bollworm, Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae), compared to Steward® 150 EC, although the difference in larval mortality between the products was statistically not significant. Dobie (2010) reported Delegate® WG to be highly efficacious against potato tuberworm, Phthorimaea operculella Zeller (Lepidoptera: Celechiidae), due to its quick effect, however, Steward®150 EC provided longer residual activity. Liu et al. (2003) also reported on the longevity of Steward® 150 EC, where one application of Steward® 150 EC against diamondback moth, P. xylostella, suppressed larvae below the recommended economic threshold for 14 - 21 days in the field. However, this slow-acting property may affect the behaviour and physiology of target insects persistently at different levels, yet fails to cause immediate death (Wang et al., 2011). Therefore, a trade-off exists between the fast action of Delegate® WG and the longer persistence of activity of Steward® 150 EC. Nevertheless, both control options show excellent activity against larvae of A. trimenii and hold potential as exceptional rotational products for use in an IPM programme.
Bioassays testing the effectivity of different doses of DiPel® DF showed that the recommended dose (TMT2) and double the recommended dose (TMT3) give good control against larval stages of A. trimenii. Larval mortality resulting from TMT3 increased significantly on the third day after ingestion, compared to TMT2, which indicates that TMT3 caused a faster rate of mortality. However, this observation is only applicable to one day (day 3) with no significant differences in larval mortality between TMT2 and TMT3. Results show that the significant difference in larval mortality on the third day of ingestion had no significant effect on the rate of larval mortality between TMT2 and TMT3 over the seven-day period. This was confirmed, as no significant difference was observed between the means of each treatment after seven days. Treatment 2 successfully controlled 95% of A. trimenii larvae after three days and effectively killed 100% of larvae after seven days, however larval mortality was low (< 23%) 24 h after treatment. This finding is supported by Rowell (2005), who observed that B. thuringiensis var. kurstaki, did not work like most conventional insecticides in that it had no contact or partial paralysis effect (larvae were not killed instantly after application). Not only did TMT2 result in 100% larval mortality, but it is also more cost-effective than TMT3 as less amount of product is required to result in 100% larval mortality. Therefore, TMT2 (the recommended dose of DiPel® DF) should be used to control A. trimenii larvae.
Semi-field trials demonstrated the influence of water volume on the ability of a vineyard cima sprayer to effectively cover top and bottom leaves of vine canopies with DiPel® DF (Ebert et al., 1999). With field application, concerns such as coverage and application volume function through their effect on pesticide deposit structure (Ebert et al., 1999). Results showed that larval mortality was significantly higher on the bottom leaves when volume B (42 g/L 200 L/ha) was applied. This could be attributed to greater spray coverage on bottom leaves. This finding is supported by Brink et al. (2016), which observed that at higher spray volumes, lower leaf surfaces resulted in higher deposition quantity compared to upper leaf surfaces. Wise et al. (2010) reported that the volume of water used to carry the pesticide to the target, is one key factor of sprayer operation, which the grower can vary to improve the level of coverage of the targeted crop. Therefore, adjusting application volume influences coverage and affects the concentration of active ingredient per deposit given a constant application rate (Ebert et al., 1999). Additionally, it is suggested that spray coverage may be improved by increasing spray volume and decreasing tractor speed (Sial & Brunner, 2010; Cavalieri et al., 2015).
Results from semi-field trials are expected to differ in the real world under field conditions drastically. Agoma trimenii larvae appeared to feed on leaves high up in the vine canopy, as less damage was observed on bottom leaves during the visual scouting and observational studies conducted by Morris et al. (2020a). Therefore, coverage is a challenge due to the high positioning of the leaves and the need to spray the adaxial side of these leaves, as the most damaging stage of A. trimenii (early to mid-level instars) occurs at these sites. In the semi-field trials, larvae had no choice but to feed on bottom leaves; this may have resulted in higher larval mortality than would have occurred in the field under natural conditions.
Bioassays provide information on the relative toxicity of an insecticide, but it is the longevity of residual actives that informs growers about retreatment intervals. Prolonging the lifetimes of spray droplets on leaves increases the absorption and uptake of active ingredients (Knoche et al., 2000). Therefore, information on the evaporation time can help pesticide formulators develop better products to maximise uptake by leaves. It is important to establish the residual activity of DiPel® DF against the larvae of A. trimenii to incorporate appropriately timed spray applications into a control programme.
In this study, the loss of insecticidal activity over time was indicated by the decrease in larval mortality and an increase in the number of live larvae recorded. Results of semi-field trials showed a reduction in insecticidal activity for DiPel® DF applied at both water volumes, between leaves picked four days after spraying and leaves picked five days after spraying. This observation was similar to the product label recommendations of DiPel® DF, which states that after ingesting a lethal dose, larvae stop feeding within the hour and will die within several hours to three days, i.e. that after three days of ingestion, insecticidal activity will begin to decrease. This was observed for both water volumes, when larvae stopped feeding within hours of ingestion and mortality began after ± 24 hrs. Similar results were obtained in Copping & Menn (2000), who reported that feeding ceases within 2 h - 48 h of ingestion of B. thuringiensis. It can be confirmed from the results of this study that residual activity of the Bt toxins in DiPel® DF is prevalent for at least three days after spraying, regardless of an increase in water volume of applications. Therefore, water volume did not affect the longevity of DiPel® DF actives.
After ingestion of DiPel® DF treated leaves, larvae consumed little surface tissue before dying. This highlights the dual-action by which larvae are killed; inhibited feeding which leads to starvation and direct rapid action of the endotoxin (δ-endotoxins) (Copping & Menn, 2000; Bravo et al., 2007). Inhibited feeding may suggest that contamination of mouthparts by Bt crystals may be sufficient to kill those individuals, which carry some crystals from the leaf surface, either by chance or by initial feeding activity (Bailey et al., 1996). From the results of the DiPel® DF bioassays, it was expected to observe nearly 100% larval mortality on leaves picked on the first day in the semi-field trials. However, this was not the case, as larval mortality was lower than expected for both water volumes. The low levels of larval mortality for both water volumes can be attributed to several factors. The residual activity of DiPel® DF is dependent on its persistence and breakdown under field conditions (Duffield & Jordan, 2000). Such conditions include ultraviolet radiation (UV) exposure in the field, which adversely affects the insecticidal activity of B. thuringiensis (Van Es & Tautmann, 1990). Light energy destroys spore viability, degrades the toxic protein and reduces insecticidal activity (Navon 2000).
Semi-field trials commenced at 07h00 during the summer period, when sunrise was recorded at ± 05h30. Had the spray application commenced before sunrise, temperature and UV radiation would be significantly lower in comparison to post sunrise. Significant improvements are possible to extend the residual insecticidal activity of DiPel® DF, by the discovery of formulations that protect B. thuringiensis from degradation by sunlight (Behle et al., 1997). For the purposes of this study, such improvements may prove beneficial in enhancing the residual activity of DiPel® DF on the top leaves of the vine canopy. Higher larval mortality on picked bottom leaves for both water volumes over the seven day period, as well as the significantly higher larval mortality on picked bottom leaves for volume B after day 4 of spraying, suggests a more extended residual activity on the bottom treated leaves, regardless of water volume, which may be less effective for controlling A. trimenii populations, as the most damaging stages (early to mid-instars) feed on the highest leaves of the vine canopies which are exposed to significant UV radiation and subsequent high temperatures. Therefore, measures to enhance residual longevity of DiPel® DF are crucial for controlling the most aggressive larval stages of A. trimenii.
A prolonged residual activity of pesticides can be a useful tool for pest management, however, it is also a risk for resistance development in multivoltine species due to exposure to low-residue levels (Pavan et al., 2014). Insecticides that employ a long-lasting effect against A. trimenii may have a selective pressure on the next generations (Pavan et al., 2014). Additionally, the high reproductive ability of A. trimenii and its aggressive larval foliar damage make the evolution of pesticide resistance a serious threat to the sustainability of the chemical control of this pest (Morris et al. 2020a). The use of new pesticides for the control of A. trimenii requires knowledge of its current susceptibility under artificial conditions prior to application at a large scale.
The timing of application of pesticides as a means of pest control is becoming an increasingly more important issue (Morris et al., 2020a). Insect phenology models such as those based on the work of Riedl et al. (1986) on the codling moth have been used to forecast population events such as adult emergence, egg-laying and egg hatch. This information, together with knowledge of the toxicity of insecticides to different life stages of A. trimenii may be valuable in determining the appropriate timing of insecticide applications. The effectiveness of Bt application depends heavily on its timing. It is advised to spray early in the season, before high field populations of potential parasitoids and predators on the pest have been reduced by chemical pesticides and spraying after sunset instead of in the morning, which can increase the persistence of the product in warm countries, where activity of the microbe persists for only 2-3 days (Navon, 2000). Appropriate timing of spray application is crucial for A. trimenii, as this ensures a proper control method to be carried out and avoids overuse of DiPel® DF, which can increase costs. Knowledge of life stage specificity is essential for determining application timing in the field and devising bioassays in resistance monitoring programmes (Magalhaes & Welgenbach, 2011). One of the most significant economic aspects of pest management using DiPel® DF, is the application against young larvae, preferably neonates, as it has been confirmed in artificial and field bioassays that third instar larvae of Lepidoptera are less susceptible to the Bt products compared to younger larvae (Tan et al., 2008). Variations in susceptibility to B. thuringiensis during larval development would be of value in establishing the best management strategies in terms of the timing of DiPel® DF application for A. trimenii.
CONCLUSIONS
Results obtained in the current study have provided useful information on whether Delegate® WG, Steward® 150 EC and DiPel® DF can be used to control A. trimenii. From the findings, both Delegate® WG and Steward® 150 EC prove effective against the larval stages of A. trimenii, however, a trade-off exists between the fast action of Delegate®WG and the longer persistence of activity of Steward®150 EC. According to the product label, results from bioassays showed that the recommended dose of DiPel® DF is the most appropriate and cost-effective dose for the control of A. trimenii larvae compared to halving or doubling the recommended dose. Results from semi-field trials suggested that increasing the water volume of a spray application of DiPel® DF will result in greater spray coverage and subsequent larval mortality, highlighting the importance of achieving maximum spray coverage. Efforts to increase the residual longevity of DiPel® DF are imperative for effective application on top leaves of the vine canopy, as the most damaging stages of A. trimenii are located at these sites. It is suggested that knowledge of pesticide resistance, susceptibility of different life stages to all tested products and the subsequent appropriate timing of spray application, will prove advantageous in developing control programmes. Future research should be directed at investigating the effects of all products against A. trimenii under field conditions, either simultaneously or in rotation, as part of an IPM system for this pest in South African vineyards.
LITERATURE CITED
Abbas, G., Hassan, N., Farhan, M., Haq, I. & Karar, H., 2015. Effect of selected insecticides on Helicoverpa armigera Hubner (Lepidoptera: Noctuidae) on tomato (Lycopersicon esculentum Miller) and their successful management. Adv. Entomol. 3, 16. [ Links ]
Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18(2), 265-267. [ Links ]
Abrol, D. & Shankar, 2012. Integrated Pest Management: Principles and Practice. CAB International, Wallingford. [ Links ]
Bacci, L., Lupi, D., Savoldelli, S. & Rossaro, B., 2016. A review of Spinosyns, a derivative of biological acting substances as a class of insecticides with a broad range of action against many insect pests. J. Entomol. Acarol. Res. 48, 40-52. [ Links ]
Bailey, P., Baker, G. & Caon, G., 1996. Field efficacy and persistence of Bacillus thuringiensis var. kurstaki against Epiphyas postvittana (Walker) (Lepidoptera: Tortricidae) in relation to larval behaviour on grapevine leaves. Aust. J. Entomol. 35, 297-302. [ Links ]
Behle, R.W., McGuire, M.R. & Shasha, B. S., 1997. Effects of sunlight and simulated rain on residual activity of Bacillus thuringiensis formulations. J. Econ. Entomol. 90, 1560-1566. [ Links ]
Bravo, A., Gill, S.S. & Soberon, M., 2007. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 49(4), 423-435. [ Links ]
Brink, J.C., Calitz, F. J. & Fourie, P.H., 2016. Spray deposition and control of Botrytis cinerea on grape leaves and bunches: part 1 (Table Grapes). S. Afr. J. Enol. Vitic. 37, 93-103. [ Links ]
Cavalieri, J.D., Raetano, C.G., Madureira, R. P. & Moreira, L. L., 2015. Spraying systems and traveling speed in the deposit and spectrum of droplets in cotton plant. Eng. Agrícola. 35, 1042-1052. [ Links ]
Coombes, C.A., 2012. Entomopathogenic fungi for control of soil-borne life stages of false codling moth, Thaumatotibia leucotreta (Meyrick) (1912) (Lepidoptera: Tortricidae). Thesis, Rhodes University, Drostdy Road, Grahamstown 6139, South Africa. [ Links ]
Copping, L.G. & Menn, J.J., 2000. Biopesticides: A review of their action, application and efficacy. Pest Manag. Sci. 56, 655-676. [ Links ]
De Prins, J. & De Prins, W. 2021. Afromoths, online database of Afrotropical moth species (Lepidoptera). http://www.afromoths.net. [May 2021] [ Links ]
Devetak, M., Vidrih, M. & Trdan, S., 2010. Cabbage moth (Mamestra brassicae [L.]) and bright-line brown-eyes moth (Mamestra oleracea [L.])-presentation of the species, their monitoring and control measures. Acta Agric. Slov. 95, 149. [ Links ]
Dinter, A. & Wiles, J.A., 2000. Safety of the new DuPont insecticide, "Indoxacarb" to beneficial arthropods: an overview. IOBC WPRS Bulletin. 23, 149-156. [ Links ]
Dobie, C.H., 2010. Pesticide susceptibility of potato tuberworm in the Pacific Northwest (Doctoral dissertation, Washington State University). [ Links ]
Dripps, J., Olson, B., Sparks, T. & Crouse, G., 2008. Spinetoram: how artificial intelligence combined natural fermentation with synthetic chemistry to produce a new spinosyn insecticide. Plant Health Progress August. [ Links ]
Duffield, S.J. & Jordan, S.L., 2000. Evaluation of insecticides for the control of Helicoverpa armigera (Hübner) and Helicoverpa punctigera (Wallengren) (Lepidoptera: Noctuidae) on soybean, and the implications for field adoption. Aust. J. Entomol. 39, 322-327. [ Links ]
Ebert, T.A., Taylor, R.A.J., Downer, R.A. & Hall, F.R., 1999. Deposit structure and efficacy of pesticide application: Interactions between deposit size, toxicant concentration and deposit number. Pestic. Sci. 55, 783-792. [ Links ]
Goettel, M.S., Eilenberg, J. & Glare, T., 2005. Entomopathogenic fungi and their role in regulation of insect populations. In: Gilbert, L.I., Iatrou, K. & Gill, S.S. (eds.). Comprehensive molecular insect science. Elsevier, Amsterdam. pp. 361 - 406. [ Links ]
Inglis, G.D., Goettel, M., Butt, T. & Strasser, H., 2001. Use of hyphomycetous fungi for managing insect pests. In: Butt, T.M., Jackson, C.W. & Magan, N. (eds.). Fungi as biocontrol agents: Progress, problems and potential. CABI, Wallingford. pp. 23 - 70. [ Links ]
Knoche, M., Lownds, N. K. & Bukovac, M. J., 2000. Spray application factors and plant growth regulator performance: IV. Dose response relationships. J. Am. Soc. Hortic. Sci. 12, 195-199. [ Links ]
Liu, T.X., Sparks, A.N. & Chen, W., 2003. Toxicity, persistence and efficacy of indoxacarb and two other insecticides on Plutella xylostella (Lepidoptera: Plutellidae) immatures in cabbage. Int. J. Pest Manag. 49, 235-241. [ Links ]
Morris, C.A., 2019. Trimen's false tiger moth, Agoma trimenii (Lepidoptera: Agaristidae): Biology and potential control options. Thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Morris, C.A., Johnson, S.A., De Waal J.Y. & Malan A.P., 2020a. Review of Trimen's false tiger moth, Agoma trimenii (Lepidoptera: Agaristidae): Seasonal biology, potential monitoring and control techniques. S. Afr. J. Enol. Vitic. 42(2), 128-132. [ Links ]
Morris, C.A., Malan A.P., De Waal, J.Y. & Johnson, S., 2020b. Laboratory bioassays on the susceptibility of Trimen's false tiger moth, Agoma trimenii (Lepidoptera: Agaristidae), to entomopathogenic nematodes and fungi. S. Afr. J. Enol. Vitic. 41(2), 183-188. [ Links ]
Magalhaes, L.C. & Walgenbach, J.F., 2011. Life stage toxicity and residual activity of insecticides to codling moth and oriental fruit moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 104, 1950-1959. [ Links ]
Nowak, J.T., McCravy, K.W., Fettig, C.J. & Berisford, C.W., 2001. Susceptibility of adult hymenopteran parasitoids of the Nantucket pine tip moth (Lepidoptera: Tortricidae) to broad-spectrum and biorational insecticides in a laboratory study. J. Econ. Entomol. 94, 1122-1127. [ Links ]
Pavan, F., Cargnus, E., Bigot, G. & Zandigiacomo, P., 2014. Residual activity of insecticides applied against Lobesia botrana and its influence on resistance management strategies. Bull. Insectology 67, 273-280. [ Links ]
Pretorius, J.D., Zaayman, J.L. & Van den Berg, J., 2012. Confirming the pest status of Trimen's false tiger moth, Agoma trimenii (Felder) (Lepidoptera: Agaristidae), on grapevines in South Africa. Afr. Entomol. 20, 198-200. [ Links ]
Riedl, H., Croft, B.A. & Howitt, A.J., 1986. Forecasting codling moth phenology based on pheromone trap catches and physiological-time models. Can. Entomol. 108(5), 449-460. [ Links ]
Roditakis, N.E., 1986. Effectiveness of Bacillus thuringiensis Berliner var. kurstaki on the grape berry moth Lobesia botrana Den. and Schiff. (Lepidoptera, Tortricidae) under field and laboratory conditions in Crete. Entomol. Hell. 4, 31-35. [ Links ]
Rowell, B., 2005. Bt basic for Vegetables Integrated Pest Management. IPM Newslettter # 11. Dept. of Horticulture, University of Kentucky. pp 8. [ Links ]
Sanchez-Bayo, F.P., 2012. Insecticides mode of action in relation to their toxicity to non-target organisms. J. Environ. Anal. Toxicol. S4, S4-002. [ Links ]
Shimokawatoko, Y., Sato, N., Yamaguchi, Y. & Tanaka, H., 2012. Development of the novel insecticide spinetoram (Diana®). - Sumitomo Chemical Co., Ltd., Tokyo. [ Links ]
Sial, A.A. & Brunner, J.F. 2010. Lethal and sublethal effects of an insect growth regulator, pyriproxyfen, on oblique banded leafroller (Lepidoptera: Tortricidae). J. Econ. Entomol. 103(2), 340-347. [ Links ]
Sial, A.A., Brunner, J.F. & Garczynski, S.F., 2011. Biochemical characterization of chlorantraniliprole and spinetoram resistance in laboratory-selected oblique banded leafroller, Choristoneura rosaceana (Harris) (Lepidoptera: Tortricidae). Pestic. Biochem. Physiol. 99, 274-279. [ Links ]
Statsoft inc, 2016. STATISTICA (Data Analysis Software System). Version 14.0. [ Links ]
Tabashnik, B.E., Cushing, N.L., Finson, N. & Johnson, M.W., 1990. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 83, 1671-1676. [ Links ]
Wang, G., Huang, X., Wei, H. & Fadamiro, H.Y., 2011. Sublethal effects of larval exposure to indoxacarb on reproductive activities of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae). Pestic. Biochem. Phys. 101(3), 227-231. [ Links ]
Wing, K.D., Sacher, M., Kagaya, Y., Tsurubuchi, Y., Mulderig, L., Connair, M. & Schnee, M., 2000. Bioactivation and mode of action of the oxadiazine indoxacarb in insects. Crop Protect. 19, 537-545. [ Links ]
Submitted for publication: March 2021
Accepted for publication: December 2021
* Corresponding author: S. Johnson sjohnson@sun.ac.za
Acknowledgements: The authors would like to thank the South African Table Grape Industry (SATI) and the National Research Foundation (NRF-THRIP: TP2011060100026) for funding of the project.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}