










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.40 n.2 Stellenbosch 2019
http://dx.doi.org/10.21548/40-2-3420
ARTICLES
doi:https://doi.org/10.21548/40-2-3420
Potential of South African entomopathogenic nematodes to control the leaf miner, Holocacista capensis (Lepidoptera: Heliozelidae)
L.A.I. Steyn; P. Addison; A.P. Malan*
Department of Conservation Ecology and Entomology, Faculty of AgriSciences, Stellenbosch University, Private Bag X1, Matieland 7602, Stellenbosch, South Africa
ABSTRACT
The Cape grapevine leafminer, Holocacista capensis, a sporadic pest of economic importance, is found in South African table and wine grape vineyards. The cocoon casings, constructed by the final instar larvae, can be found attached to grape bunches, posing a phytosanitary risk for table grape export markets. The current study aimed to determine the susceptibility of leaf-mining H. capensis larvae to seven entomopathogenic nematode (EPN) species belonging to Steinernematidae and Heterorhabditidae. To determine nematode virulence occupied leaf-mining galleries were extracted from infested leaves and inoculated with 200 infective juveniles (IJs) in 50 μ! of distilled water, for each of the EPNs screened. Concentration assays were conducted and and lethal dose was determined for the three most virulent species (Heterorhabditis baujardi, H. indica and H. noenieputensis), using concentrations of 0, 25, 50, 100, 200, and 400 IJs/leaf-mining larva. High mortality of leaf-mining larvae was obtained with H. baujardi (92%), H. noenieputensis (85%) and H. indica (83%). Almost double the number of H. noenieputensis (34 nematodes/insect) penetrated the insect larvae, in comparison with the other two EPNs. However, the relative potency of H. baujardi was 3.56 times higher than for H. indica, whilst that of H. indica was 2.57 times higher than it was for H. noenieputensis. The results obtained in the laboratory were found to be encouraging, especially with regard to the nematodes' ability to penetrate the leaf-mining galleries, and to infect the larvae successfully.
Key words: biological control, cocoon casing, EPN, Heterorhabditis, integrated pest management, leaf miner, Steinernema
INTRODUCTION
Holocacista capensis Van Nieukerken & Geertsema (Lepidoptera: Heliozelidae) is a sporadic, multivoltine pest that occurs on most ornamental and commercial varieties of Vitis vinifera L. in South Africa. The leaf-mining larvae were first reported in 2012 in a table grape vineyard close to Paarl in the Western Cape province(Van Nieukerken & Geertsema, 2015; Torrance, 2016). To date, no insecticide has been registered against the pest. This is attributed to the fact that an effective monitoring method for use in field trials is absent, as the male attractant is not commercially available.
To avoid control strategies relying solely on harmful insecticides, a variety of biological control strategies should be studied. Hagler (2000) describes biological control as "the deliberate exploitation of a natural enemy for pest control". A wide range of economically important insect pest populations have been suppressed through the deliberate application of insect-parasitic organisms known as entomopathogens, which include entomopathogenic nematodes (EPNs) of the genera Heterorhabditis (Rhabditida: Heterorhabditidae) and Steinernema (Rhabditida: Steinernematidae) (Stock & Hunt, 2005). In South Africa, a total of 17 Steinernema and seven Heterorhabditis species have been isolated from soil surveys conducted throughout the country (Malan & Hatting, 2015; Malan & Ferreira, 2017).
The ability of an EPN and its associated symbiotic bacteria to succeed as a control agent essentially depends on four factors: moisture requirements (or desiccation tolerance, with ± 85% relative humidity being required for optimum control); optimal temperature range; susceptibility of the targeted insect; and foraging strategy (Lacey & Georgis, 2012). EPNs, which occur naturally in soils throughout the world, are able to control foliar, soil-borne, cryptic, and subterranean pests from a variety of diverse habitats (Lacey & Georgis, 2012). They are suitable for including in an integrated pest management (IPM) programme, as no secondary effects of EPNs on non-target organisms are known to exist (Wright et al., 2005). The method of cover and placement of EPNs on open surfaces (like foliar habitats) is critical, as humidity, exposure to ultraviolet light and temperature are not buffered, in contrast to those that are experienced in soil environments (Wright et al., 2005; Dito et al., 2016). The use of EPNs has been successfully implemented as a control strategy for two leaf-mining lepidopteran pests, namely the tomato leaf miner, Tuta absoluta (Meyrick) (Gelechiidae) (Batalla-Carrera et al., 2010; Gözel & Kasap, 2015; Van Damme et al., 2015; Kamali et al., 2017; Mutegi et al., 2017) and the citrus leaf miner, Phyllocnistis citrella Stainton (Gracillariidae) (Beattie et al., 1995).
Most of the studies undertaken so far on leaf-mining Lepidoptera have focused on the use of steinernematids. These studies have identified Steinernema affine Wouts, Mrácek, Gerdin & Bedding (Gözel & Kasap, 2015), Steinernema carpocapsae Wouts, Mrácek, Gerdin & Bedding (Beattie et al., 1995; Batalla-Carrera et al., 2010; Gözel & Kasap, 2015; Van Damme et al., 2015; Kamali et al., 2017), Steinernema feltiae (Filipjev) Wouts, Mrácek, Gerdin & Bedding (Batalla-Carrera et al., 2010; Gözel & Kasap, 2015; Van Damme et al., 2015), and Steinernema karii Waturu, Hunt & Reid (Mutegi et al., 2017) as successful biological control agents against a variety of leaf-mining insects. Heterorhabditis bacteriophora Poinar has been used in an attempt to control T. absoluta pest populations, although success has only been achieved under greenhouse conditions (Batalla-Carrera et al., 2010; Gözel & Kasap, 2015; Kamali et al., 2017).
The use of EPNs to control leaf-mining insects through foliar applications using adjuvants seems promising, regardless of the concerns expressed regarding humidity and temperature requirements. It is hypothesized that the protection provided by the blotch mine created by the pest insect could potentially reduce the desiccation of infective juveniles (IJs) and the extent of exposure of the individuals to ultraviolet light (Gözel & Kasap, 2015). With the exception of those conducted by Beattie et al. (1995), using S. carpocapsae against P. citrella, EPNs have not yet been successful in adequately controlling T. absoluta, another major lepidopteran leaf-mining pest, in field trials. Gözel & Kasap (2015) speculated that the requirements of the sensitive relationships existing between the selected EPN and the target pest (in terms of virulence, host-seeking strategy, and ecological factors) must be satisfied, before considering field applications. Thereafter, additional knowledge of the appropriate application strategy (i.e. field dosage, volume, irrigation, and appropriate application methods) must be considered.
The preliminary evidence, based on findings associated with other leaf-mining foliar pests, indicates that EPNs could potentially be used to control H. capensis larvae in an IPM strategy. The aim of the current study was to determine whether H. capensis larvae are susceptible to EPNs. The main objective was, therefore, to screen different local EPN species, and to test their respective virulence against H. capensis larvae under laboratory conditions.
MATERIALS AND METHODS
Source of insects
Grapevine leaves infested with H. capensis larvae were collected from farms on the outskirts of Halfmanshof (33°08'48.7"S 18°59'18.0"E), Robertson (33°50'19.6"S 19°54'52.7"E), Klapmuts (33°49'30.3"S 18°55'36.8"E), and Paarl (33°40'20.4"S 18°56'25.0"E). Field-collected larvae were stored at 10 °C before being counted and used in EPN screening trials within 24 h after collection (pilot trials indicated high levels of mortality when the experiments were not carried out on the same day that collection took place).
Once collected from the field, each leaf was examined, with the aid of a stereomicroscope, for the presence of active/ feeding H. capensis larvae. Occupied mines (each containing a single larva) were carefully cut out from the collected leaves, so as to avoid any cuts or tears of the epidermal layers of the leaf surrounding the larva. A margin of 2 to 5 mm of unmined leaf tissue was maintained around the entire leaf-mining gallery. Approximately 2 340 feeding larvae were manually processed in this manner for the experiments conducted in this study (excluding mines that were damaged in the cutting process and leaves examined that contained deceased larvae). Larvae collected for the screening tests were of different ages, as it was not possible to determine the age or instar without damaging the larvae or their galleries.
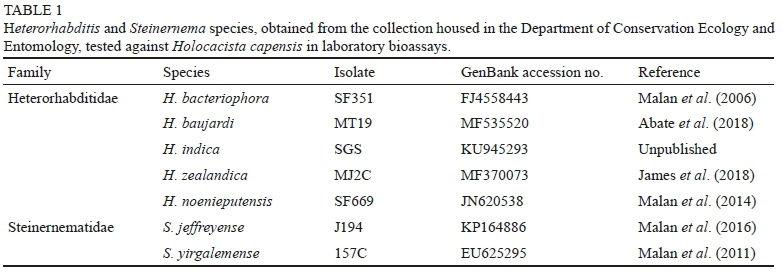
Source of EPNs
The seven EPN species that were tested for virulence against H. capensis larvae were obtained from the collection housed in the Department of Conservation Ecology and Entomology, Stellenbosch University (Table 1). Prior to screening, modified White's traps were used to collect the IJs cultured from EPN-inoculated larvae of Galleria mellonella L. (Lepidoptera: Pyralidae), otherwise known as greater wax moth (Kaya & Stock, 1997). More specifically, 10 G. mellonella larvae were placed in a Petri dish (90 mm diameter) lined with filter paper, and inoculated with 800 μl solution of an EPN suspension. The Petri dish was stored in a growth chamber at 25 °C. Transferral of the G. mellonella cadavers to clean Petri dishes, lined with moist filter paper, occurred 48 hours later. After 7 to 10 days, the Petri dish containing the dead larvae was placed on a modified White's trap to facilitate the collection of the emerging IJs (White, 1927). The IJ suspensions were harvested and transferred to vented culture flasks. The flasks were stored at 14 °C in a dark growth chamber, before being used in the screening tests.
Virulence assays for Holocacista capensis larvae
The virulence of each EPN species (Table 1) to H. capensis larvae was tested at a concentration of 200 IJs/50 μl of distilled water per larva, while the controls were inoculated with 50 μl of distilled water only, using 24-well bioassay plates (Flat-bottom, Nunc™, Cat. no. 144530) as the test arena. Ten wells within each bioassay plate were lined with a circular piece of filter paper (13 mm diameter), placed in alternate wells, to obtain an even distribution throughout the plate. Each of the selected wells was inoculated with 200 IJs/50 μl of distilled water, using a separate Eppendorf® micropipette (www.eppendorf.com) for each EPN species. An occupied leaf mine (containing a single live/feeding H. capensis larva) was added to each inoculated well. The lid of the bioassay plate was secured in place with a rubber band. Three replicate plates were used for each of the EPN species tested (n = 30 occupied leaf mines/treatment). The plates were stored in 2 L plastic containers lined with moistened tissue paper (to maintain high humidity levels) and placed in a growth chamber at 25 °C for 48 h. Thereafter, the H. capensis larvae were removed from the leaf mines and larval mortality was determined. The experiment was repeated, using a fresh batch of cultured IJs and leaf-mining larvae, on a subsequent date.
Penetration analysis
Dead larvae from the virulence assays were placed in a clean Petri dish (90 mm diameter) lined with filter paper. The Petri dishes were placed in moistened 2 L plastic containers, and transferred back to the 25 °C growth chamber for a further 24 h. Thereafter, the number of IJs that had penetrated each larva was recorded by means of dissection with the aid of a stereomicroscope.
Lethal dose
The three most virulent EPN species from the previous experiments, namely Heterorhabditis baujardi Phan, Subbotin, Nguyen & Moens, Heterorhabditis indica Poinar, Karunakar & David, and Heterorhabditis noenieputensis Malan, Knoetze & Tiedt were used to determine their respective lethal concentrations. EPN concentrations of 0, 25, 50, 100, 200, and 400 IJs/larva were tested against H. capensis. Again, 24-well plates were used as the test arena, with each containing 10 alternately placed leaf mines enclosing a single live/feeding H. capensis larva. Three 24-well plates (n = 30) were used for each EPN concentration of each treatment.
Similar to the experimental procedure that was previously adopted (as described for the virulence assays), pieces of filter paper (13 mm diameter) were placed in 10 alternate wells, to which the occupied leaf mines were added. Thereafter, the leaf mines were inoculated with the various concentrations of IJs in 50 μ! of distilled water. Distilled water only was used for the untreated control treatments. The plates for each treatment were placed in a 2 L plastic container lined with moist tissue paper, and kept in a growth chamber at 25 °C. After 48 h, the larvae were removed from the mines and the percentage mortality of infected H. capensis larvae was determined visually, by dissection, and recorded.
Data analysis
All analyses were performed using STATISTICA 13.0 (Dell Inc., Headquarters in Round Rock, Texas, USA). If no significant difference in test date versus treatment interactions was recorded as the main effects in a two-way ANOVA, the data from the various test dates were pooled and analysed using a one-way ANOVA to identify the most virulent EPN species. A Fisher LSD post-hoc test was performed to determine significant differences between means. To confirm the differences in mortality between IJ doses within each species a one-way ANOVA and a Fisher LSD post-hoc test, by group (nematode species), was conducted. The recorded penetration data of IJs found within the H. capensis larvae 72 h after inoculation were analysed with a one-way ANOVA and a Fisher LSD post-hoc test. Correlation analyses, a factorial ANOVA, and a Fisher LSD post-hoc test were conducted to compare the effect of dose among the different EPNs tested. To determine the lethal doses of the three most virulent EPN species, a probit analysis (Finney, 1952) was carried out on mortality data, using Polo PC (LeOra Software, 1987).
RESULTS
Virulence assays for Holocacista capensis larvae
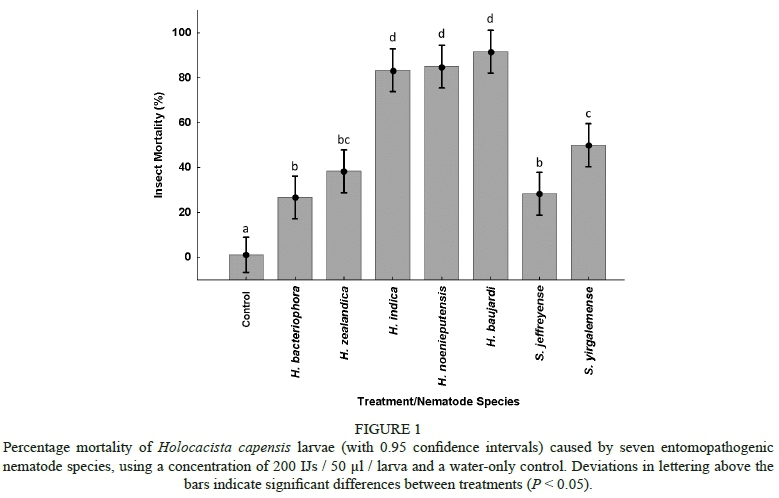
Screening trials showed that the IJs were capable of penetrating occupied, undamaged leaf-mining galleries and of infesting the leaf-mining H. capensis larvae within 48 h after inoculation (Fig. 1). Analysis of the pooled data, and the subsequent multiple comparisons test, showed that all of the screened EPN species caused significantly higher mortality of the H. capensis larvae than the untreated control (F743 = 56.099; P < 0.001). The mortality (infection) associated with H. bacteriophora (26.67% ± 4.94%) and Steinernema jeffreyense Malan, Knoetze & Tiedt (28.33% ± 4.77%) was the lowest and did not differ significantly (P = 0.804). The natural mortality of the control treatment was close to zero (Fig. 1). Heterorhabditis baujardi (91.67% ± 3.07%), H. indica (83.33% ± 7.60%) and H. noenieputensis (85.00% ± 4.28%) caused the highest H. capensis larval mortality. The three treatments did not differ significantly from each other (P > 0.2). The virulence of Heterorhabditis zealandica Poinar (38.33% ± 7.03%) did not differ significantly from that of H. bacteriophora (P = 0.088), S. jeffreyense (P = 0.142), or Steinernema yirgalemense Nguyen, Tesfamariam, Gozel, Gaugler & Adams (50.00% ± 2.58%) (P = 0.088).
Penetration analysis
The mean number of IJs that penetrated H. capensis larvae differed significantly between the screened EPN species (F635 = 13.157; P < 0.001) (Fig. 2). The highest mean penetration was achieved by H. noenieputensis (34.27 ± 6.94), which differed significantly from that of H. baujardi (18.23 ± 2.90) (P = 0.002) and H. indica (17.02 ± 3.98) (P = 0.001). The lowest mean penetration of 1.45 (± 0.33) was achieved by S. jeffreyense, which did not differ significantly from H. bacteriophora (2.72 ± 0.82) (P = 0.789), H. zealandica (4.13 ± 1.61) (P = 0.571), and S. yirgalemense (4.97 ± 1.10) (P = 0.458).
Lethal dose
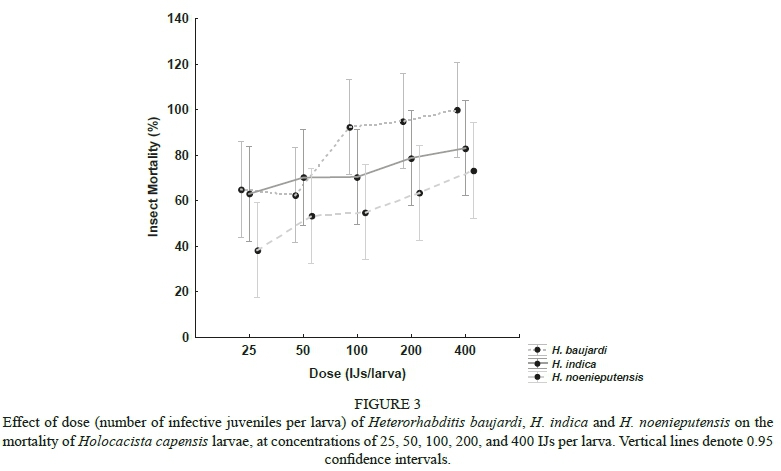
A positive correlation between the dosage and lethal capacity of the EPN species was recorded (r = 0.783) (Fig. 3). As the natural mortality in the untreated control (< 6.67%) was significantly different from all other doses (P < 0.01), the control treatment was removed from the analysis to appropriately gauge differences in infection between the treatments and the respective doses. The mortality recorded for the 25, 50 and 400 IJ/larva treatments did not differ significantly between the EPN species tested (P > 0.05). The 100 IJ and 200 IJ treatments, however, differed between H. baujardi and H. noenieputensis (P = 0.016 and P = 0.0375, respectively) (Fig. 3). No significant effect of dose was recorded for the H. indica treatments (P > 0.05).
For all of the EPN species tested, the highest insecticidal activity was reached at 400 IJs/larva, although mortality did not differ significantly between the 100, 200 and 400 IJ/larva treatments (P > 0.5) for each respective species.
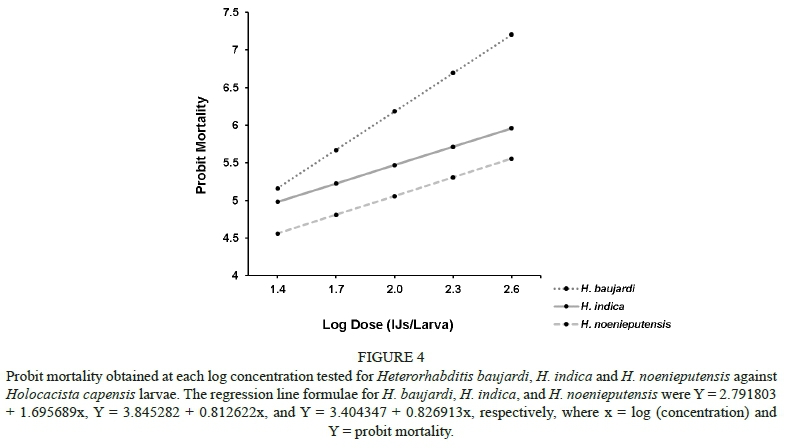
The probit analysis indicated that the slopes and the intercepts of the regression lines for the three EPN species, H. baujardi, H. indica and H. noenieputensis, differed significantly (χ2= 36.835, P < 0.01, df = 4). Their slopes were, however, constrained (i.e. parallel) (χ2 = 4.451; P = 0.108, df = 2). Trends in lethal capacity differed slightly for each of the EPN species, as indicated by the probit regression lines for the three heterorhabditids (y = 1.6957x + 2.7918, y = 0.8126x + 3.8453, and y = 0.8269x + 3.4043, respectively) (Fig. 4).
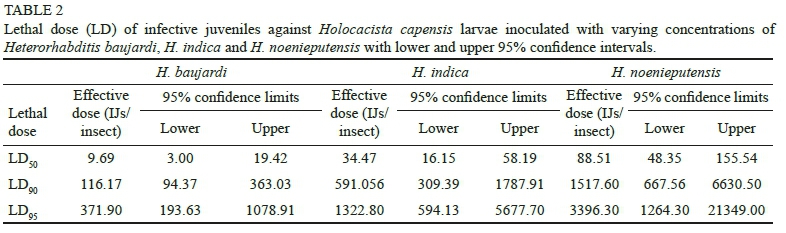
Heterorhabditis baujardi exhibited the lowest LD50 and LD90 values (9.69 and 94.37 IJs/larva, respectively), whereas H. noenieputensis had the highest LD50 and LD90 values C88.51 and 1517.60 IJs/larva, respectively) (Table 2). The relative potency of H. baujardi was 3.56 times higher than that of H. indica, whereas H. indica was 2.57 times more potent than H. noenieputensis (Table 3).
DISCUSSION
EPNs regulate insect populations by using an insect as a host for reproduction and breeding purposes (Griffin et al., 2005). The application of different obligate, parasitic EPN species in conjunction with other chemical, cultural and physical control strategies, can be used to control leaf-mining insects. The current study aimed to determine whether local EPNs could potentially be used as biological control agents to aid in the control of H. capensis, a foliar pest of commercial table grapes in South Africa. High virulence would allow for the inclusion of EPN applications (after the necessary field trials) in an integrated approach for the control of this phytosanitary pest. The present study is the first to consider the use of biological control strategies against H. capensis, using locally isolated EPNs.
All of the local EPN species tested caused a significant increase in the larval mortality of H. capensis, compared to the untreated control. Three of the EPN species, namely H. indica, H. noenieputensis and H. baujardi, yielded excellent results (83%, 85% and 91% larval mortality, respectively) in relation to virulence and the ability to penetrate leaf-mining galleries. Assessing these isolates in field trials to confirm virulence under field conditions is, therefore, highly recommended. The other EPN species tested caused between 26% and 50% larval mortality. Virulence did not differ significantly between the EPN species tested at lower doses, with differences only becoming apparent at doses of 100 IJs/larva and more. In a similar study by Van Damme et al. (2015), S. feltiae and S. carpocapsae caused the highest mortality of T. absoluta for all larval stadia pooled (79% and 80%, respectively), whereas H. bacteriophora (60%) did not perform as well.
Bastidas et al. (2014) identified the importance of the size of the target insect, as well as that of the EPN species used against it. They found that although all the steinernematids screened were able to invade small insect hosts, the invasion potential decreased as the target insect's size decreased and the EPN species' size increased. This was confirmed in a study by Katumanyane et al. (2018), where large South African EPN species (> 1000 μm), like Steinernema khoisanae Nguyen, Malan & Gozel, S. jeffreyense and Steinernema litchii Steyn, Knoetze, Tiedt & Malan caused almost zero control of another micro-insect, Bradysia impatiens Johannsen (Diptera: Sciaridae). The effect of the size of the nematodes compared to that of the host was confirmed in Dlamini's (2018) study on Frankliniella occidentalis (Thysanoptera: Thripidae). The high virulence and relatively high penetration potential recorded for H. indica, H. noenieputensis, and H. baujardi in the present study might be attributed to their relatively small body size. Comparatively small hosts are speculated to limit the development of the invading nematodes, while the presence of relatively large hosts is necessary for the long-term persistence ofthe nematodes (Bastidas et al., 2014). This raises concerns regarding adequate and persistent control of H. capensis under field conditions. As the three most virulent species in the current study were heterorhabditids, the possession of a dorsal tooth (which is generally absent in steinernematids) might possibly also have facilitated the penetration of the leaf-mining larvae (Griffin et al., 2005).
The probit analysis indicated a positive relationship between total larval mortality and IJ concentration/dose for all EPN species tested (i.e. the larval mortality increased with the increase in dose). When considering H. baujardi, H. indica, and H. noenieputensis as potential biological control agents, it is important to note that H. baujardi proved to be just over three times more potent than H. indica, with a lower number of IJs required to attain a lethal dose of 90% of larvae than for H. indica. In turn, H. indica was just more than twice as potent as H. noenieputensis. Based on the results of this study, these three species should be tested under field conditions for the control of H. capensis. In addition, they should also be tested against other economically important leaf-mining insects, like T. absoluta and P. citrella.
To enable as many IJs as possible to locate the mine entrance of a leaf-mining insect, Wright et al. (2005) emphasised the need to maximise the density and distribution of EPNs on the leaf surfaces. As a nematode's residual infectivity generally lasts only a few hours, appropriate cover and placement is critical in foliar applications of EPNs that are targeted at controlling leaf-mining insects (Wright et al., 2005). The importance of sufficient cover and placement is further underscored by the fact that limited migration opportunities exist to compensate for suboptimal placement, which is made even more acute in the dry summer conditions of the Western Cape province. The need to apply foliar and aerial EPN treatments for a variety of pests that persist above the soil surface has necessitated the study of various adjuvants for maintaining suitable conditions, to ensure the longevity of the IJs on field-treated crops. The addition of adjuvants to various EPN species solutions proved to be positive against codling moth (De Waal et al., 2013), citrus mealybug (Van Niekerk & Malan, 2015), wheat stem sawfly (Portman et al., 2016), and vine mealybug (Platt et al., 2018; 2019).
Several other South African table grape pests (Allsopp et al., 2015) have been shown to be susceptible to EPNs, including Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae) (Le Vieux & Malan, 2013), Phlyctinus callosus (Schönherr) (Coleoptera: Curculionidae) (Ferreira & Malan, 2014), and Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) (Malan & Moore, 2016; Steyn, 2019). It would be ideal if the same EPN species could be used against all of these pests. Alternatively, a combination of EPNs could be developed for foliar application to target the leaf-miner and these other pests simultaneously.
Appropriate mass-rearing techniques and formulations with suitable adjuvants still need to be developed for the area-wide use of local EPNs in commercial vineyards. Advances in improving the method of mass-rearing heterorhabditid colonies (by means of improving solid-state and liquid cultures) should facilitate the future use of the successful candidates identified in the present study on a commercial scale (Ehlers et al., 2000; Ferreira & Malan, 2014).
CONCLUSIONS
This study identified three locally sourced EPNs, namely H. baujardi, H. indica, and H. noenieputensis, as potential biological control agents for the Cape grapevine leafminer, H. capensis, in South Africa. It provides a baseline for further research regarding the targeted control of H. capensis larvae with EPNs. Field trials should aim to determine the frequency of EPN applications required under field conditions, as well as optimal EPN concentrations and appropriate application techniques. The adoption of biological control measures for the integrated control of H. capensis would serve not only to minimise the risk of developing insecticide resistance in pest populations, but it would also pose less of a risk to human well-being and longevity.
LITERATURE CITED
Abate, B.A., Slippers, B., Wingfield, M.J., Malan, A.P. & Hurley, B.P., 2018. Diversity of entomopathogenic nematodes and their symbiotic bacteria in South African plantations and indigenous forests. Nematology 20, 355 -371. doi:10.1163/15685411-00003144 [ Links ]
Allsopp, E., Barnes, B.N., Blomefield, T.L. & Pringle, K.L., 2015. Grapevine. In: Prinsloo, G.L. & Uys, V.M. (eds). Insects of Cultivated Plants and Natural Pastures in Southern Africa, Entomological Society of Southern Africa, Pretoria. pp. 420 - 437. [ Links ]
Bastidas, B., Portillo, E. & San-Blas, E., 2014. Size does matter: the life cycle of Steinernema spp. in micro-insect hosts. J. Invertebr. Pathol. 121, 46 - 55. doi:10.1016/j.jip.2014.06.010 [ Links ]
Batalla-Carrera, L., Morton, A. & García-Del-Pino, F., 2010. Efficacy of entomopathogenic nematodes against the tomato leafminer Tuta absoluta in laboratory and greenhouse conditions. BioControl 55, 523 - 530. doi:10.1007/s10526-010-9284-z [ Links ]
Beattie, G.A.C., Somsook, V., Watson, D.M., Clift, A.D. & Jiang, L., 1995. Field evaluation of Steinernema carpocapsae (Weiser) (Rhabditida: Steinernematidae) and selected pesticides and enhancers for control of Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae). Aust. J. Entomol. 34, 335 - 342. doi:10.1111/j.1440-6055.1995.tb01351.x [ Links ]
De Waal, J.Y., Malan, A.P. & Addison, M.F., 2013. Effect of humidity and a superabsorbent polymer formulation on the efficacy of Heterorhabditis zealandica (Rhabditida: Heterorhabditidae) to control codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Biocontrol Sci. Techn. 23, 62 -78. doi:10.1080/09583157.2012.736472 [ Links ]
Dito, D.F., Shapiro-Ilan, D.I., Dunlap, C.A., Behle, R.W. & Lewis, E.E., 2016. Enhanced biological control potential of the entomopathogenic nematode, Steinernema carpocapsae, applied with a protective gel formulation. Biocontrol Sci. Techn. 26: 835 - 848. doi: 10.1080/09583157.2016.1159659 [ Links ]
Dlamini, T.M., 2018. Prospects for using entomopathogenic nematodes as a biocontrol agent against western flower thrips Frankliniella occidentalis (Thysanoptera: Thripidae). MSc thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Ehlers, R.U., Niemann, I., Hollmer, S., Strauch, O., Jende, D., Shanmugasundaram, M., Mehta, U.K., Easwaramoorthy, S.K. & Burnell, A., 2000. Mass production potential of the bacto-helminthic biocontrol complex Heterorhabditis indica - Photorhabdus luminescens. Biocontrol Sci. Techn. 10: 607 - 616. doi:10.1080/095831500750016406 [ Links ]
Ferreira, T. & Malan, A.P., 2014. Potential of entomopathogenic nematodes for the control of the banded fruit weevil, Phlyctinus callosus (Schönherr) (Coleoptera: Curculionidae). J. Helminthol. 88, 293 - 301. doi:10.1017/ S0022149X13000175 [ Links ]
Finney D.J., 1952. Probit Analysis. Cambridge University Press, London. [ Links ] Gözel, Ç. & Kasap, 1., 2015. Efficacy of entomopathogenic nematodes against the tomato leafminer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in tomato field. Turk. Entomol. Derg. 39, 229 - 237. doi:10.16970/ted.84972 [ Links ]
Griffin, C.T., Boemare, N.E. & Lewis, E.E., 2005. Biology and behaviour. In: Grewal, P.S., Ehlers, R.-U. & Shapiro-Ilan, D.I. (eds). Nematodes as Biocontrol Agents, CABI Publishing, Wallingford. pp. 47 - 64. [ Links ]
Hagler, J.R., 2000. Biological control of insects. In: Rechcigl, J.E. & Rechcigl, N.A. (eds). Insect Pest Management: Techniques for Environmental Protection, CRC Press LLC, Florida. pp. 207 - 242. [ Links ]
James, M., Addison, P. & Malan, A.P., 2018. Surveying and screening South African entomopathogenic nematodes for the control of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann). Crop Prot. 105, 41 - 48. doi:10.1016/j.cropro.2017.11.008 [ Links ]
Kamali, S., Karimi, J. & Koppenhöfer, A.M., 2017. New insight into the management of the tomato leaf miner, Tuta absoluta (Lepidoptera: Gelechiidae) with entomopathogenic nematodes. J. Econ. Entomol. 111, 112 - 119. doi:10.1093/jee/tox332 [ Links ]
Katumanyane, A., Ferreira, T. & Malan, A.P., 2018. Potential use of local entomopathogenic nematodes to control Bradysia impatiens (Diptera: Sciaridae) under laboratory conditions. Afr. Entomol. 26, 337 - 349. doi:10.4001/003.026.0337 [ Links ]
Kaya, H.K. & Stock, S.P., 1997. Techniques in insect nematology. In: Lacey, L.A. (ed.). Manual of Techniques in Invertebrate Pathology, Academic Press, San Diego, California. pp. 281 - 324. [ Links ]
Lacey, L.A. & Georgis, R., 2012. Entomopathogenic nematodes for control of insect pests above and below ground with comments on commercial production. J. Nematol. 44, 218 - 225. [ Links ]
LeOra Software. 1987. POLO-PC: A user's guide to probit or logit analysis. LeOra Software. Berkeley, CA. [ Links ]
Le Vieux, P.D. & Malan, A.P., 2013. An overview of the vine mealybug (Planococcus ficus) in South African vineyards and the use of entomopathogenic nematodes as potential biocontrol agent. S. Afr. J. Enol. Vitic. 34, 108 - 118. [ Links ]
Malan, A.P. & Ferreira, T., 2017. Entomopathogenic nematodes. In: Fourie, H., Spaull, V.W., Jones, R.K., Daneel, M.S. & De Wale, D. (eds). Nematology in South Africa: a view from the 21st Century, Springer International, Berlin. pp. 459 - 480. [ Links ]
Malan, A.P. & Hatting, J.L., 2015. Entomopathogenic nematode exploitation: case studies in laboratory and field applications from South Africa. In: Campos-Herrera, R. (ed.). Nematode Pathogenesis of Insects and Other Pests, Springer International Publishing, Basel. pp. 477 - 508. [ Links ]
Malan, A.P., Knoetze, R. & Moore, S.D., 2011. Isolation and identification of entomopathogenic nematodes from citrus orchards in South Africa and their biocontrol potential against false codling moth. J. Invertebr. Pathol 108, 115 - 125. doi:10.1016/j.jip.2011.07.006 [ Links ]
Malan, A.P., Knoetze, R. & Tiedt, L.R., 2014. Heterorhabditis noenieputensis n. sp. (Rhabditida: Heterorhabditidae), a new entomopathogenic nematode from South Africa. J. Helminthol. 88, 139 - 151. doi:10.1017/ S0022149X12000806 [ Links ]
Malan, A.P., Knoetze, R. & Tiedt, L.R., 2016. Steinernema jeffreyense n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from South Africa. J. Helminthol. 90, 262 - 278. doi:10.1017/ S0022149X15000097 [ Links ]
Malan, A.P. & Moore, S.D., 2016. Evaluation of local entomopathogenic nematodes for the control of false codling moth, Thaumatotibia leucotreta (Meyrick, 1913), in a citrus orchard in South Africa. Afr. Entomol. 24, 489 - 501. doi:10.4001/003.024.0489 [ Links ]
Malan, A.P., Nguyen, K. & Addison, M., 2006. Entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) from the southwestern parts of South Africa. Afr. Plant Prot. 12, 65 - 69. [ Links ]
Mutegi, D.M., Kilalo, D., Kimenju, J.W. & Waturu, C., 2017. Pathogenicity of selected native entomopathogenic nematodes against tomato leaf miner (Tuta absoluta) in Kenya. World J. Agric. Res. 5, 233 - 239. doi:10.12691/ wjar-5-4-5 [ Links ]
Platt, T., Stokwe, N.F. & Malan, A.P., 2018. Potential of local entomopathogenic nematodes for control of the vine mealybug, Planococcus ficus. S. Afr. J. Enol. Vitic. 39, 208 - 215. doi: 10.21548/39-2-3158 [ Links ]
Platt, T., Stokwe, N.F. & Malan, A.P., 2019. Foliar application ofSteinernema yirgalemense to control Planococcus ficus: assessing adjuvants to improve efficacy. S. Afr. J. Enol. Vitic. 40, 13 - 19. doi:10.21548/40-1-2920 [ Links ]
Portman, S.L., Krishnankutty, S.M. & Reddy, G.V.P., 2016. Entomopathogenic nematodes combined with adjuvants presents a new potential biological control method for managing the wheat stem sawfly, Cephus cinctus (Hymenoptera: Cephidae). PLoS ONE 11. doi:10.1371journal.pone.0169022. [ Links ]
Steyn, V.M., 2019. Integrated control of false codling moth, Thaumatotibia leucotreta, on stone fruit and table grapes. PhD thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Stock, S.P. & Hunt, D.J., 2005. Morphology and systematics of nematodes used in biocontrol. In: Grewal, P.S., Ehlers, R.-U. & Shapiro-Ilan, D.I. (eds). Nematodes as Biocontrol Agents, CABI, Wallingford. pp. 3 - 43. [ Links ]
Torrance, L.A.I., 2016. The bio-ecology of the Cape grapevine leafminer, Holocacista capensis (Lepidoptera: Heliozelidae), in the Western Cape. MSc thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Van Damme, V.M., Beck, B.K.E.G., Berckmoes, E., Moerkens, R., Wittemans, L., De Vis, R., Nuyttens, D., Casteels, H.F., Maes, M., Tirry, L. & De Clercq, P., 2015. Efficacy of entomopathogenic nematodes against larvae of Tuta absoluta in the laboratory. Pest Manag. Sci. 72, 1702 - 1709. doi:10.1002/ps.4195 [ Links ]
Van Niekerk, S. & Malan, A.P., 2015. Adjuvants to improve aerial control of the citrus mealybug Planococcus citri (Hemiptera: Pseudococcidae) using entomopathogenic nematodes. J. Helminthol. 89, 189 - 195. doi:10.1017/ S0022149X13000771 [ Links ]
Van Nieukerken, E.J. & Geertsema, H., 2015. A new leafminer on grapevine and Rhoicissus (Vitacea) in South Africa within an expanded generic concept of Holocacista (Insecta, Lepidoptera, Heliozelidae). ZooKeys 507, 41 - 97. doi:10.3897/zookeys.507.9536 [ Links ]
White, G.F., 1927. A method for obtaining infective nematode larvae from cultures. Science 66, 302 - 303. [ Links ]
Wright, D., Peters, A., Schroer, S. & Fife, J., 2005. Application technology. In: Grewal, P.S., Ehlers, R.-U. & Shapiro-Ilan, D.I. (eds). Nematodes as Biocontrol Agents, CABI, Wallingford. pp. 91 - 106. [ Links ]
Submitted for publication: March 2019
Accepted for publication: July 2019
* Corresponding author: E-mail address: apm@sun.ac.za
Acknowledgements: The authors wish to thank D.G. Nel (Centre for Statistical Consultation, Stellenbosch University) and K.L. Pringle (Department of Conservation Ecology and Entomology, Stellenbosch University) for their assistance with the statistical analyses. Financial support was provided by the National Research Foundation (NRF), the South African Table Grape Industry (SATI), and the Technology and Human Resources for Industry Programme (THRIP-TP14062571871)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}