










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.40 n.2 Stellenbosch 2019
http://dx.doi.org/10.21548/42-2-3224
ARTICLES
doi:https://doi.org/10.21548/42-2-3224
Hydric behaviour and gas exchange in different grapevine varieties (Vitis vinifera L.) from the Maule Valley (Chile)
G. Gutiérrez-GamboaI; A.G. Pérez-DonosoII; A. Pou-MirIII; C. Acevedo-OpazoIV; H. Valdés-GómeII, *
IUniversidad de Talca, Facultad de Ciências Agrárias, Centro Tecnológico de la Vid y del Vino (CTW), Av. Lircay S/N, Talca, Chile
IIPontifícia Universidad Católica de Chile, Departamento de Fruticultura y Enología, Vicuna Mackena 4860, Santiago, Chile
IIIInstituto de Ciências de la Vid y del Vino (CSIC, Gobierno de La Rioja, Universidad de La Rioja). Carretera de Burgos, Km. 6. 26007 Logrorio, Spain
IVUniversidad de Talca, Facultad de Ciências Agrárias, CITRA, 2 Norte, 685 Talca, Chile
ABSTRACT
In the near future, stomatal behaviour will be crucial to counteract conditions arising from climate change. Grapevine varieties are classified as either isohydric or anisohydric, depending on the sensitivity of sto-mata to water deficit and on their water potential homeostasis. However, the great variability observed in different studies indicates that a continuum exists in the range of stomatal sensitivity to water stress. Thus, more knowledge about the hydric behaviour and the gas exchange of isohydric and anisohydric grapevine varieties under different water conditions could lead to the development of irrigation strategies oriented at improving water-use efficiency, yield and berry composition. In this study, research was conducted in order to characterise the stomatal regulation of four different Vitis vinifera L. varieties, namely Pinot noir, Sauvignon blanc, Chardonnay and Merlot, according to soil water status. Measurements of leaf gas exchange, together with measurements of stem water potential (Ψ$) and leaf water potential (Ψ1), were taken during two seasons. Under conditions of water stress, Chardonnay and Merlot reached a minimum Ψ$ of -1.67 and -1.68 MPa respectively, and higher levels of water-use efficiency (AN/g), of 62.3 and 69.7 μmol C02/mol H20 respectively. In Sauvignon blanc and Pinot noir, the minimum Ψ$ was -1.26 and -1.40 MPa respectively, with lower levels of AN/gs (53.1 and 50.5 μmol C02/mol H20, respectively). Under conditions of water stress (Ψ1 < -0.9 MPa and *Fs < -0.6 MPa), all varieties had a significantly increased A^g1, despite a significant reduction in gas exchange. Therefore, the hydric behaviour and gas exchange observed in this study suggest that Chardonnay and Merlot could be characterised as anisohydric varieties, as they present less sensitive stomatal control, while Pinot noir can be classified as a near-anisohydric variety and Sauvignon blanc as an isohydric variety. New investigations should consider other characteristics of the varieties to classify them better.
Keywords: Anisohydric, intrinsic water-use efficiency, isohydric, leaf water potential, stem water potential
INTRODUCTION
Recently, viticultural management has experienced a series of modifications due to the increase in temperatures, which has had direct effects on grapevine ripening and, consequently, on berry and wine quality (Van Leeuwen & Darriet, 2016). The effects of climate change on winemaking can vary according to the style of wine produced and by geographical location, with milder effects expected for coastal regions (Jones et ah, 2005; Duchêne et ah, 2010; Verdugo-Vásquez et ah, 2019). Model outputs have predicted an average warming of 2°C in the next 50 years for global wine-producing regions (Jones et ah, 2005). Advances from eight to 11 days for budburst and from 16 to 24 days for véraison have been predicted for the end of the 21st century for white grapevine varieties cultivated in Alsace (Duchêne et ah, 2010). A decrease in the length of each phenological stage in grapevines may yield a precocity towards the end of the 21st century of 40 days earlier than the current timeframe (Ollat & Touzard, 2014; Sgubin et ah, 2018). Thus, grapevines have undergone modifications in their physiological behaviour that have affected vegetative growth (Gomez del Campo et ah, 2002; Lebon et ah, 2006), berry development (McCarthy, 2000; Ojeda et ah, 2001), and the maturity and organoleptic composition of the grape berry at harvest (Koundouras et ah, 1999; Ojeda et ah, 2002). Thus, the observed climatic effects would be modifying both the spectrum and the distribution of the currently used grapevine varieties (Popescu et ah, 2009), and would imply the implementation of adequate measures to reduce the negative effects through adjustments and changes in agricultural systems.
Soil water deficit and vapour-pressure deficit (VPD) are the most important environmental factors affecting stomatal closure in the Mediterranean area (Klein, 2014). Pou et al. (2008) reported that stomatal conductance (gs) and the rate of transpiration (E) were strongly affected by VPD in a manner that is dependent on irrigation treatment. Depending on the stomatal control strategy followed in response to water deficit, grapevine varieties can behave as isohydric or anisohydric plants (West et al., 2012). In isohydric varieties, gs responds to a greater extent to the changes in VPD. This high stomatal sensitivity of isohydric plants is usually associated with higher levels of abscisic acid in the xylem sap, and with hydraulic responses (Tardieu & Simonneau, 1998; Soaref al., 2006). In contrast, anisohydric grapevines have less control over Ψ1, and the magnitude of Ψ1 decreases as VPD increases, reaching much lower values of Ψ1 in stressed vines than in grapevines growing under favourable water conditions. In this case, stomatal closure in these grapevines is associated with root signals or with the anatomical architecture related to stress conditions (Tardieu & Simonneau, 1998; Collins et al., 2010; Rogiers et al., 2011; Gerzon et al., 2015). Anisohydric grapevines present a substantial depression of their Ψ during drought, also showing some degree of tolerance to water stress (Bucci etal, 2005; Pou et al., 2012). Pou et al. (2012) reported that anisohydric behaviour results in better performance under moderate water stress and recovery than isohydric-behaved grapevines (Bucci et al., 2005; Pou et al., 2012).
A better knowledge of the hydric behaviour and gas exchange of isohydric and anisohydric grapevine varieties managed under different water conditions could lead to the development of irrigation strategies oriented to improving water productivity, yield and berry composition under the current effects of climate change. Therefore, the aim of this work was to characterise the vine water potential and gas exchange in four grapevines varieties (isohydric and anisohydric) managed under different water conditions in the Maule Valley, Chile over two consecutive seasons.
MATERIALS AND METHODS
Experimental site and plant material
The field trial was conducted in an experimental vineyard (cv. Chardonnay, Merlot, Pinot noir and Sauvignon blanc) belonging to the Vine and Wine Technological Centre of Talca University during the 2011/2012 and 2012/2013 seasons. The vineyard is located in Panguilemo, Talca, Maule Valley, Chile (35°22.2' south 71°35.39' west, and 121 metres above sea level). The ungrafted grapevines were planted in 2006, trained to a vertical shoot position trellis system and were pruned into two bilateral spur cordons. Planting density was about 5 000 vines ha1, with grapevine spacing between rows and within the row of 2.00 m χ 1.00 m and an east-west orientation. The soil texture was clay loam with a rooting depth of 150 cm, and the soil is classified as part of the Talca series (Thermic Ultic Haploxeralf). Bulk density, field capacity, wilting point and available water along the soil were 1.36 g cm3, 0.31 m3 m3, 0.13 m3 m3and 0.18 m3 m3 respectively. More information concerning soil and crop management carried out in the experimental field are available in Panitrur-De la Fuente et al. (2018).
Information about weather conditions was recorded by an automatic weather station installed 50 m from the trial plot. Maximum and minimum temperature in the 2011/2012 grapevine growth season (from September to April) was 34.6°C and 0.0°C respectively, while in the 2012/2013 season it was 34.5°C and 0.9°C respectively. Average temperature in the season was 16.9°C and 16.7°C for the 2011/2012 and 2012/2013 seasons respectively. The rainfall registered for the phenological cycle in the 2011/2012 season reached 31.8 mm, and was concentrated in the spring months. The rainfall registered for the 2012/2013 season reached 152.4 mm, was concentrated in November, and there was occasional rain throughout the summer, unlike the 2011/2012 season, when there was no precipitations during the summer. The reference evapotranspiration (ET) was estimated using the Penman-Monteith equation (Ahumada-Orellana et al., 2018). ET calculated for the 2011/2012 season was 1 037 mm, and for the 2012/2013 season it was 939 mm. In the 2011/2012 season, 1 438 chilling hours were accumulated, while in the 2012/2013 season 1 243 chilling hours were accumulated. During the 2011/2012 season, the accumulation of degree days was 1 375°C, while the accumulation of degree days during the 2012/2013 season was 1 311°C. Mean VPD during the experiment in the 2011/2012 season was 1.05 kPa, while in the 2012/2013 season it was 0.95 kPa. Additional information is presented in Figs 1 to 4 of the supplementary material.
Experimental design
The experimental design was a randomised complete block divided into two plots (treatments), in which the four Vitts vinifera L. varieties under study were arranged: Chardonnay, Merlot, Pinot noir and Sauvignon blanc. During the first season, one of the two blocks was managed under optimal water conditions (without irrigation restriction), while the other was managed with progressive water restrictions until reaching severe water deficit in the vines (leaf water potential < -1.4 MPa) (Van Leeuwen et al. (2009). During the second season, differential irrigation management was not carried out as in the first season; instead, all the plants were irrigated with the same water load. Despite this, plants presented differences in their water status in the second experimental year, thus presenting a wide range of water stress within the experiment. A total of eight treatments (variety χ water condition) were arranged in the vineyard, with five plants per treatment. The selected grapevines presented good phytosanitary conditions and were homogeneous in vegetative growth and productivity.
Gas exchange measurement
An infrared gas analyser, model LI 6400 (Li-cor, Lincoln, Nevada, USA), was used to measure stomatal conductance (gs), transpiration (E) and net C02 assimilation (AN). In addition, the intrinsic water-use efficiency (AN gs_1) was calculated as the ratio between AN and gs according to Medrano etal. (2014). The gas exchange measurements were carried out in the middle zone (6th leaf from the tip on fruit-bearing shoots) on the northern side of the canopy between 12:00 and 14:00 in five different plants per treatment on fully sunny days. The original position of the selected leaves in the canopy was not changed and the same light regime was maintained, thus ensuring that the leaves were exposed to full sunlight (PAR > 800 μmol m2 sec1). Measurements were taken every 15 days between December (one month before véraison) and April (near harvest) in each growing season.
Plant water status
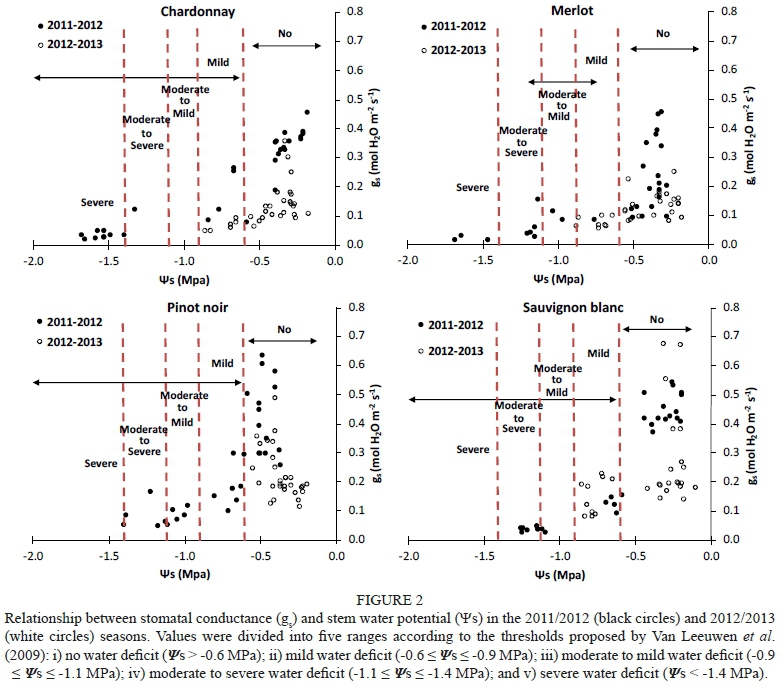
Stem water potential (Ψβ) and leaf water potential (Ψ1) were measured using a pressure chamber (PMS Instrument Co., model 600, Corvallis, Oregon, USA) according to the protocol stated by Acevedo-Opazo et al. (2013) and Jara-Rojas et al. (2015). Briefly, five fully expanded and sun-exposed leaves per treatment were wrapped in plastic transparent film and aluminium foil for at least 2 h, thus achieving an equilibrium between leaf and plant xylem. Ψ1 measurement were performed on uncovered leaves at the same time that and gas exchange measurements were taken (between 12:00 and 14:00). To define the level of water stress in this study, data were divided into five ranges according to the thresholds proposed by van Leeuwen et al. (2009): i) no water deficit (Ψ1 > -0.9 MPa; > -0.6 MPa); ii) mild water deficit (-0.9 < Ψ1 < -1.1 MPa; -0.6 < < -0.9 MPa; iii) moderate to mild water deficit (-1.1 < Ψ1 < -1.3 MPa; -0.9 < < -1.1 MPa; iv) moderate to severe water deficit (-1.3 < Ψ1< -1.4 MPa; -1.1 < < -1.4 MPa; v) severe water deficit (Ψ1 and < -1.4 MPa).
Statistical analysis
The statistical analysis in relation to the parameters analysed was performed by ANOVA, using Centurion XVI.I (Statgraphics Technologies; The Plains, Virginia, USA). Differences between treatments were compared using the Duncan test at the 95% confidence level. Non-linear regressions between gs and Ψ1, gs and Ψβ, AN and gs, Ε and gs, Ε and Ψβ, and \ gs' and gs were developed for different levels of vine water status by each grapevine variety. The coefficient of determination (r2) was used to evaluate how well the regression line represents the data. The relationship between net C02 assimilation (A^) and stomatal conductance (gs) in each variety was contrasted with that in the literature in order to compare how these varieties behave under different experimental conditions.
RESULTS AND DISCUSSION
Relationship between stomatal conductance and plant water status
To evaluate genotypic sensitivity to water deficit, the stomatal conductance (gs) was compared with the leaf water potential (Ψ1) and stem water potential (Ψβ) (Figs 1 and 2 respectively) in the 2011/2012 and 2012/2013 seasons. For similar values of Ψ1 and Ψβ, all varieties except Sauvignon blanc presented higher maximum values of gs during the first season (2011/2012) than during the second season (2012/2013) (Figs 1 and 2), as well as lower minimum gs values (Figs 1 and 2). During the 2011/2012 season, minimum gs values for Chardonnay, Merlot, Pinot noir and Sauvignon blanc were 0.02, 0.01, 0.05 and 0.03 mol H20 nr2 s1 respectively, whereas they were 0.05, 0.05, 0.11 and 0.08 mol H20 nr2 s"1 respectively in the 2012/2013 season. This higher sensitivity of the stomatal response to water deficit during the former season is probably related to the higher vapour-pressure deficit (VPD) experienced at the time of the measurements, reaching 1.05 kPa, compared to the 0.95 kPa reached during the second season. Prieto et al. (2010) reported that Syrah, Marselan, Mouvèdre and Ekigaïna grapevine varieties presented higher stomatal regulation in response to increased ambient VPD. Moreover, even under mild to moderate water stress (-0.9 < Ψ1 < -1.3 MPa), the variability of gs values was also higher in 2011/2012 than in 2012/2013 (Fig 1). Generally, when Ψ1 was used as an indicator of water status, higher gs magnitudes and more variability were observed at lower water potentials than when was used. Although Ψ1 varies according to vine water status, it is also dependent on the microclimatic environment of the leaf; due to this, midday Ψ1 probably is not a very accurate indicator of vine water status (Van Leeuwen et al., 2007). Moreover, a clear separation in gs magnitudes between stressed and non-stressed grapevines was only found when water status was characterised as (Fig. 2). The variability in gs decreased in most of the grapevine varieties in the 2012/2013 season, especially for values lower than -0.9 MPa (moderate water deficit) (Fig. 2).
Regarding gs sensitivity, Chardonnay and Merlot reached maximum g values of 0.45 mol HO m2 s1, and minimum g values of 0.02 mol HO m2 s1 and 0.01 mol H20 nr2 s1 respectively, while Pinot noir and Sauvignon blanc reached maximum g values of 0.63 mol HO m2 s1. and minimum g values of 0.05 mol HO m2 s1 and 0.03 mol
H20 m2 s1 respectively. Stomatal conductance is not only related to the availability of soil water and VPD, but also the interactions of internal and external leaf factors, such as hydraulic adjustment, root signals or anatomical architecture (Collins et al., 2010; Pou et al., 2012; Gerzon et al., 2015). Indeed, it has been shown that differences in the abscisic acid concentration [ABA] in the xylem sap may explain the more sensitive reaction to water deficit in isohydric grapevine varieties compared to anisohydric varieties (Chaves et al., 2016). ABA could be involved in the closure of aquaporins in bundle sheath cells, decreasing the water flow to the mesophyll cells and strengthening the implications of a hydraulic component in stomata closure (Chaves et al., 2016). Thus, according to the results presented in Figs 1 and 2, this might be the case in Sauvignon blanc, since there is rapid stomatal closure as the water content in the soil decreases. In this way, the leaf water status interacts strongly with gas exchange and, consequently, there is a well-defined correlation between Ψ1 and gs (Prieto et al., 2010).
The Merlot, Pinot noir and Chardonnay grapevines progressively decreased their gs as stem water potential (Ψβ) became more negative due to high VPD and conditions of water stress. These varieties maintained moderately high gs levels under mild water deficit conditions (-0.9 MPa < < -0.6 MPa), as defined by Van Leeuwen et al. (2009). However, gs progressively declined with mild to moderate water stress (Ψβ < -0.6 MPa), and remained constant at a gs value close to 0.1 mol H20 m2 s1. This physiological behaviour is common in species described as having anisohydric responses to soil water deficit (Pou et ah, 2012). Thus, compared to isohydric behaviour, anisohydric behaviour involves the consumption of soil water resources by roots until lower water potentials are achieved (Chaves et ah, 2016). Merlot and Chardonnay presented lower values than Pinot noir and Sauvignon blanc (Fig. 2). In contrast, Sauvignon blanc grapevines managed under conditions of no water deficit (Ψε > -0.6 MPa). They presented high gs values (> 0.4 mol H20 m2 s1) and, after this point, the values dropped drastically (0.25 < mol H20 m2 s1) during the first season, leading to decreased transpiration and, consequently, decreased diffusion of C02 into the plant. During the second season, Pinot noir and Sauvignon blanc decreased their gs down to 0.2 mol H20 m2 s1 at values of -0.3 MPa (no water stress) (Fig. 2), thus avoiding a drastic fall in Ψβ. Pinor noir showed different hydric behaviour in the two experimental seasons. As a consequence, this variety was described as displaying anisohydric behaviour in the first season and isohydric behaviour in the second.
Relationship between net C02 assimilation and stomatal conductance
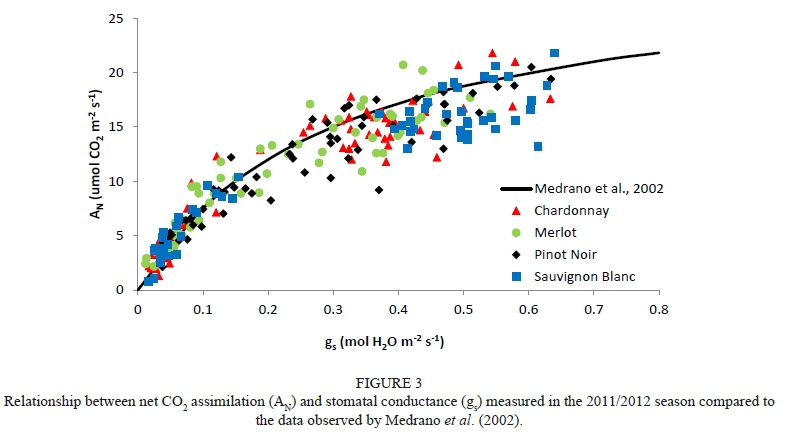
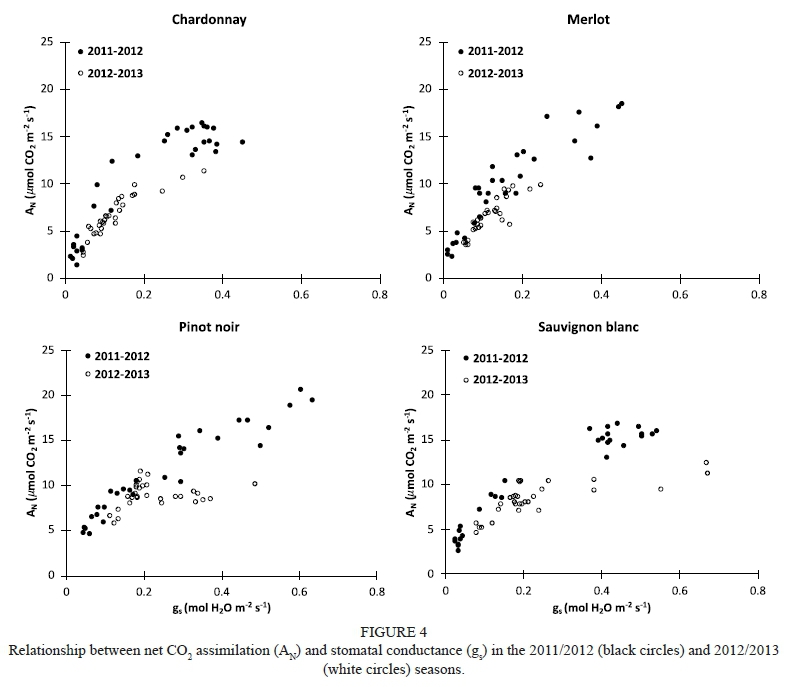
A typical exponential relationship for net C02 assimilation (AN) and stomatal conductance (gs) was found on the basis of the data measured during the field trial. Our data are located around the curve proposed by Medrano et al. (2002), which is considered representative for most grapevine varieties (Fig. 3). Moreover, different relationships between A^ and gs for each variety were obtained in both experimental years (Fig. 4).
The obtained \ data was homogeneously distributed throughout the gs data spectra for the Chardonnay, Pinot noir and Merlot grapevines (Fig. 4). Thus, as the gs increased, there was greater AN. In contrast, there were no \ values in Sauvignon blanc when the gs values were between 0.25 and 0.4 mol H20 m2 s1. At one extreme, when well-watered conditions were applied, this variety assimilated C02 at a high rate. However, under water-stressed conditions, gs fell considerably in comparison to the rest of the grapevine varieties, probably due its strong stomatal regulation, leading to a fast decrease in AN. This protective physiological response might have costs in terms of lower C02 assimilation rates during water stress, leading to a reduction of growth, and under severe abiotic stress this could reach a critical threshold for leaf damage (Chaves et al, 2010, 2016; Pou et al, 2012). One the other hand, Pinot noir grapevines growing under severe water-stress conditions did not completely close their stomata, maintaining a higher C02 assimilation rate than the other studied varieties. This could imply that, in this variety, lower gs magnitudes could be less limiting for C02 assimilation than for the transpiration rate compared to other varieties. Additionally, under well-watered conditions, this grapevine variety presented the highest maximum AN value compared with the rest of the studied varieties (Fig. 4), suggesting near-anisohydric behaviour in this case. Accordingly, Chaves et al. (2016) report that anisohydric varieties present cooler leaves and higher photosynthetic rates than isohydric ones. However, anisohydric grapevines may suffer accelerated dehydration under severe drought stress due to their high transpiration rates, which are not compensated by soil water uptake (Chaves et al., 2016). With respect to stomatal control strategy, certain authors have reported that Pinot noir behaved as an anisohydric variety when water stress was applied at the pre-véraison stage and as an isohydric variety when it was applied at the post-véraison stage (Poni et al.. 1993;Lovisoloefa/.,2010).
Transpiration according to stomatal conductance and plant water status
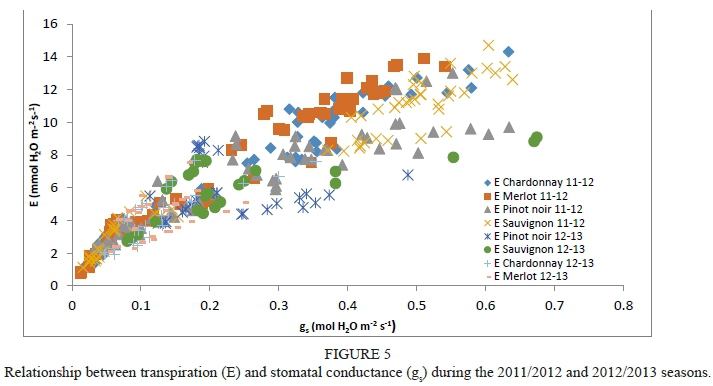
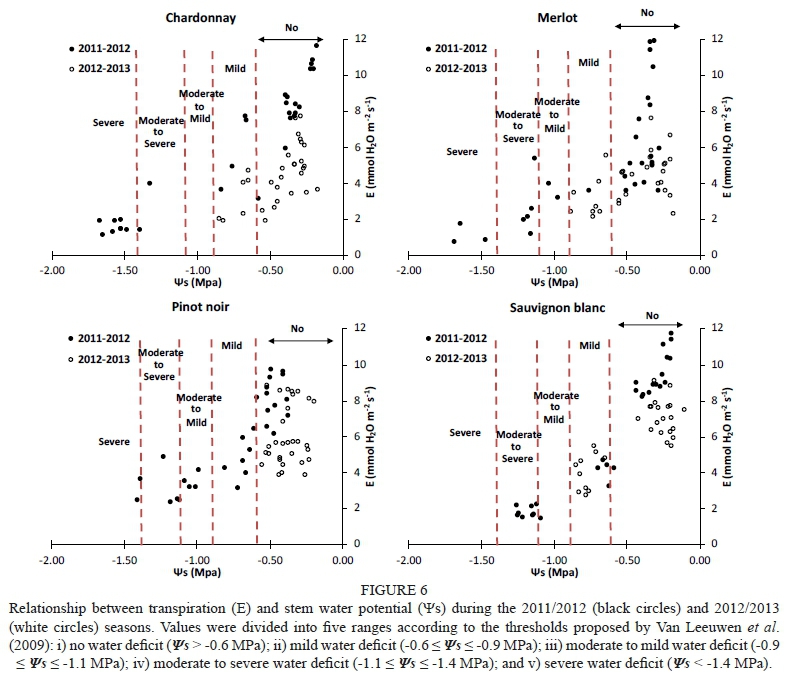
Grapevines close their stomata under conditions of water stress, leading to a decrease in transpiration (E). Therefore, gs has a great influence on Ε (Fig. 5). Correlation coefficients (r2) between Ε and gs were higher than 0.83 for the 2011/2012 season (data not shown). However, the obtained r2 in the 2012/2013 season were only significant for Chardonnay (r2 = 0.71). This was probably because the treatments performed during the second season did not generate a severe water deficit, which mean there were no extreme values.
The 2011/2012 season was warmer than the 2012/2013 season and presented a higher reference evapotranspiration (ETo) and vapour-pressure deficit (VPD). This had a direct effect on Ε and stomatal conductance (gs) rates, which were greater in the 2011/2012 season, favouring more negative water potentials.
Ε declined as became more negative. Similar Ε magnitudes close to 12 mmol H20 m2 s1 were observed in Chardonnay, Merlot and Sauvignon blanc grapevines for the 2011/2012 season (Fig. 6). Pinot noir grapevines reached maximum Ε values of slightly below the others. Chardonnay grapevines presented a progressive drop in Ε from 10.3 to 1.1 mmol H20 m2 s1, and values lower than -0.2 MPa (non-stressed vines). This behaviour was similar to that found in Merlot, Pinot noir and Sauvignon blanc grapevines, which presented a progressive fall in Ε as became more negative. In Chardonnay and Merlot, levels of below -1.6 MPa (severe water stress) gave rise to minimum Ε (< 1.1 mmol H20 m2 s1). However, the minimum value of Ε (1.45 mmol H20 m2 s1) in Sauvignon blanc was reached at -1.1 MPa, defined as moderate to severe water stress by Van Leeuwen et al. (2009). In addition, high levels of Ε were reached under non-stress conditions in Sauvignon blanc grapevines, and subsequently Ε fell drastically to Ψε values lower than -0.6 MPa, with an average value of 3.1 mmol H20 m2 s1. Moreover, stressed Pinot noir grapevines presented higher levels of Ε at very negative (-1.4 MPa) than those obtained in Sauvignon blanc, Chardonnay and Merlot. In Pinot noir, Ε fluctuated slightly at values lower than -0.9 MPa, ranging from 4.8 to 2.3 mmol H20 m2 s1, with a minimum value of 2.33 mmol H20 m2 s1 at -1.2 MPa. Then, under moderate to severe water-stress conditions, Ε fluctuated between 1.45 and 2.26 mmol H20 m2 s1. Therefore, Ε followed the same behaviour as g and contributed to the explanation of the isohydric (Sauvignon blanc) or anisohydric (Merlot and Chardonnay) behaviour of the varieties.
Intrinsic water-use efficiency
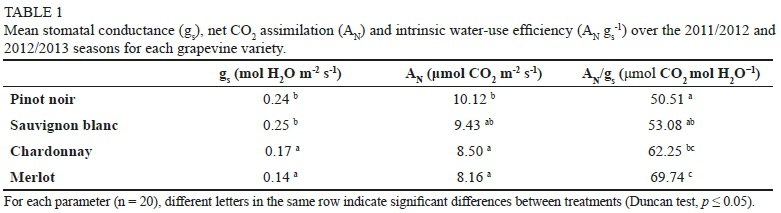
Merlot and Chardonnay grapevines showed higher levels of ANgs-1 than Pinot noir, mainly due to the low levels of gs obtained; therefore, it is likely that these varieties partially owe their higher efficiency in the use of water to the fact that their stomata were not completely open (Table 1). Sauvignon blanc and Pinot noir reached a lower \ gjlthan Merlot, together with high levels of gs, which could affect their productive potential under conditions of unfavourable water availability. Based on the literature, isohydric varieties under high evaporative demand have been considered as experiencing a more pronounced increase in \ gs-', thus being better adapted to drought stress than anisohydric varieties (Schultz 2003; Vandeleur et al., 2009). However, other reports in the literature show that the same variety could behave as iso- or anisohydric, depending on the experimental conditions (Chaves et al., 2010; Lovisolo et al., 2010). Moreover, Pou et al. (2012) showed that Chardonnay (considered an anisohydric variety) displayed higher water-use efficiency at the leaf level than two other isohydric varieties.
ANgs-1 is independent of atmospheric conditions, since it measures the ability of the leaf to regulate photosynthesis and gs (Medrano et al., 2007). Fig. 7 shows the different values of gs measured during the first and second experimental seasons and their respective estimated water-use efficiency (WUE) values. Based on our results for gs values between 0.70 and 0.14 mol H20 m2 s1 (slight stress) with adequate water availability, \ gs' gradually increased as the water status of the grapevines decreased. Under these conditions, AN decreased slightly and stomatal closure limited photosynthesis. Thereafter, as gs decreased from 0.14 mol H20 m2 s1 to 0.05 mol H20 m2 s1 (moderate water stress), \ decreased and AN gs-1 increased significantly. Under these conditions, stomatal limitations dominated and photosynthesis was reversible. Finally, at gs levels lower than 0.05 molH20 m2 s1 (severe water stress). \ gs' decreased drastically again, as has been reported by Cifre et al. (2005). Therefore, grapevines become less efficient in the use of water when there is severe water stress represented by low gs levels (Fig. 7). The depicted results displayed under moderate water stress differ from those reported by Douthe et al. (2018) when measuring WUE at the whole-plant scale. These authors showed that carbon and water fluxes were drastically reduced, while estimated WUE was not improved but decreased. So, in this case, scaling up WUE readings from leaves to the whole plant leads to some discrepancies among single-leaf and whole-canopy results. However, the results reported by these authors are in agreement with those shown in Fig. 7 for grapevines growing under severe water stress. Grapevine leaves from all locations in the canopy except those located in the central part, showed a similar radiation-use efficiency, suggesting that light interception considerably affects variations in photosynthesis within the grapevine canopy (Escalona et al., 2003). Thus, it is possible that, under severe water stress, variations between leaves within the canopy disappear and any single leaf may reflect what is happening at the whole-plant level. Additionally other factors, such as nocturnal water loss, changes in dry matter, partitioning among the sinks, or harvest load respiration rates, could also explain the lack of correlation usually reported between instantaneous WUE and intrinsic water-use efficiency (WUEi) (Douthe et al., 2018).
CONCLUSIONS
Chardonnay and Merlot decreased their leaf and stem water potential in accordance with the applied water stress and reached high levels of intrinsic water-use efficiency (WUEi) (ANg;1)(62.25and69.74μmol C02molΗ20' respectively). Thereby, Chardonnay and Merlot could be characterised in this trial as anisohydric varieties. In contrast, Sauvignon blanc drastically decreased gs (0.67 to 0.03 mol H20 m2 s1) and Ε (11.7 to 1.45 mmol H20 m2 s1) under conditions of water stress, leading to a strong decrease in \ (2.53 μmol C02 m2 s1), thus displaying isohydric behaviour. In Pinot noir, gs dropped (0.63 to 0.05 mol H20 m2 s1) in relation to the water deficit, and Ψ1 changed in association with the irrigation treatments, probably due to the availability of water in the soil. However, under stress conditions, gs' was lower in this variety (50.51 μmol C02 mol H20-1)-Accordingly, this variety could be characterised as either an isohydric or anisohydric variety. Consequently, these results suggest that Pinot noir may be considered as a near-anisohydric variety. In general, physiological responses of the varieties are directly related to the climate and water content in the soil and may vary from one area to another. For this reason, other physiological parameters could be measured to characterise more accurately the isohydric or anisohydric behaviour of the varieties of this study.
LITERATURE CITED
Acevedo-Opazo, C, Valdés-Gómez, Η., Taylor, J.A., Avalo, Α., Verdugo-Vásquez, Ν., Araya, Μ., Jara-Rojas, F. & Tisseyre, Β., 2013. Assessment of an empirical spatial prediction model of vine water status for irrigation management in a grapevine field. Agric. Water Manag. 124, 58-68. [ Links ]
Ahumada-Orellana, L.E., Ortega-Farias, S. & Searles, P.S., 2018. Olive oil quality response to irrigation cut-off strategies in a super-high density orchard. Agric. Water Manag. 202, 81-88. [ Links ]
Bucci, S.J., Goldstein, G., Meinzer, F.C., Franco, A.C., Campanello, P. & Scholz, F.G., 2005. Mechanisms contributing to seasonal homeostasis of minimum leaf water potential and predawn disequilibrium between soil and plant water potential in neotropical savanna trees. Trees Struct. Funct. 19. 296-304. [ Links ]
Chaves, M.M., Costa, J.M., Zarrouk, O., Pinheiro, C, Lopes, CM. & Pereira, J.S., 2016. Controlling stomatal aperture in semi-arid regions - The dilemma of saving water or being cool? Plant Sei. 251, 54-64. [ Links ]
Chaves, Μ.Μ., Zarrouk, Ο., Francisco, R., Costa, J.M., Santos, T.. Regalado, A.R, Rodrigues, M.L. & Lopes, CM., 2010. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 105(5), 661-676. [ Links ]
Cifre, J., Bota, J., Escalona, J., Medrano, H. & Flexas, J., 2005. Physiological tools for irrigation scheduling in grapevine (Vitts vinifera L.). An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 106, 159-170. [ Links ]
Collins, M.J., Fuentes, S. & Barlow, E.W.R., 2010. Partial rootzone drying and deficit irrigation increase stomatal sensitivity to vapour pressure deficit in anisohydric grapevines. Funct. Plant Biol. 37(2), 128-138. [ Links ]
Douthe, C, Medrano, H., Tortosa, 1., Escalona, J.M., Hernández-Montes, Ε. & Pou, Α., 2018. Whole-plant water use in field grown grapevine: Seasonal and environmental effects on water and carbon balance. Front. Plant Sei. 9, 1540. [ Links ]
Duchêne, Ε., Huard, F., Dumas, V, Schneider, C. & Merdinoglu, D., 2010. The challenge of adapting grapevine varieties to climate change. Clim. Res. 41, 193-204. [ Links ]
Escalona, J.M., Flexas, J., Bota, J. & Medrano, I., 2003. Distribution of leaf photosynthesis and transpiration within grapevine canopies under different drought conditions. Vitis 42(2), 57-64. [ Links ]
Gerzon, E., Biton, I., Yaniv, Y., Zemach, H., Netzer, Y, Schwartz, Α., Fait. A. & Ben-Ari, G., 2015. Grapevine anatomy as a possible determinant of isohydric or anisohydric behavior. Am. J. Enol. Vitic. 66(3), 340-347. [ Links ]
Gomez del Campo, Μ., Ruiz, C. & Lissaguirre, J.R., 2002. Effect of water stress on leaf area development, photosynthesis and productivity in Chardonnay and Airen grapevines. Am. J. Enol. Vitic. 53, 138-143 [ Links ]
Jara-Rojas, F., Ortega-Farias, S., Valdéz-Gómez, Η. & Acevedo-Opazo. C, 2015. Gas exchange relations of ungrafted grapevines (cv. Carménère) growing under irrigated field conditions. S. Afr. J. Enol. Vitic. 36, 231-242. [ Links ]
Jones, G.V., White, M.A., Cooper, O.R. & Storchmann, Κ., 2005. Climate change and global wine quality. Clim. Change 73(3), 319-343. [ Links ]
Klein, T., 2014. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 28(6), 1313-1320. [ Links ]
Koundouras, S., Van Leeuwen, C, Seguin, G. & Gloires, Y, 1999. Influence de l'alimentation en eau sur la croissance de la vigne, la maturation des raisins et les caractéristiques des vins en zone méditerranéenne (Exemple de Némée, Grèce, cépage Saint Georges, 1997). J. Int. Sei. Vigne Vin 33, 149-160. [ Links ]
Lebon, E., Pellegrino, Α., Louarn, G. & Lecoeur, J., 2006. Branch development controls leaf area dynamics in grapevine (Vitis viniferá) growing in drying soil. Ann. Bot. 98, 175-185. [ Links ]
Lovisolo, C, Perrone, I., Carra, Α., Ferrandino, Α., Flexas, J., Medrano, H. & Schubert, Α., 2010. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 37(2), 98-116. [ Links ]
McCarthy, M., 2000. Developmental variation in sensitivity of Vitis vinifera L. (Shiraz) berries to soil water deficit. Aust. J. Grape Wine Res. 6, 136-140. [ Links ]
Medrano, H., Bota, J., Cifre, J., Flexas, J., Ribas-Carbó, Μ. & Gulías. J., 2007. Eficiência del uso del agua por las plantas. Grupo de Biologia de plantas en condiciones mediterrâneas. Departamento de Biologia. Universität de les Illes Balears- IMEDEA. Investigaciones Geográficas 43. 63-84. [ Links ]
Medrano, H., Escalona, J., Bota, J., Gulías, J. & Flexas, J., 2002. Regulation of photosynthesis of C-3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 89(7), 895-905. [ Links ]
Medrano, H., Tomás, Μ., Martorell, S., Escalona, J.-M., Pou, Α., Fuentes. S., Flexas, J. & Bota, J., 2014. Improving water use efficiency of vineyards in semi-arid regions. Areview. Agron. Sustain. Dev. 35(2), 499-517. [ Links ]
Ojeda, H., Andary, C, Kraeva, E., Carbonneau, A. & Deloire, Α.. 2002. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vitic. 53, 261-267. [ Links ]
Ojeda, H., Deloire, A. & Carbonneau, Α., 2001. Influence of water stress deficits on grape berry growth. Vitis 40, 141-145. [ Links ]
Ollat, N. & Touzard, J.-M., 2014. Long-term adaptation to climate change in viticulture and enology: The LACCAVE project. Special Laccave. J. Int. Sei. Vigne Vin 1-7. [ Links ]
Panitrur-De la Fuente, C, Valdés-Gómez, Η., Roudet, J., Acevedo-Opazo. C, Verdugo-Vásquez, Ν, Araya-Alman, Μ., Lolas, Μ., Moreno, Y. & Fermaud, M., 2018. Classification of wine grape cultivars in Chile and France according to their susceptibility to Botrytis cinerea related to fruit maturity. Aust. J. Grape Wine Res. 24, 145-157. [ Links ]
Poni, S., Lakso, A.N., Turner, J.R. & Melious, R.E., 1993. The effects of pre- and post-veraison water stress on growth and physiology of potted Pinot noir grapevines at varying crop levels. Vitis 32, 207-214. [ Links ]
Popescu, Α., Enache, V, Simion, C, Donici, A. & Tabaranu, G., 2009. Research concerning the economic impact of climate change upon grape production. Bulletin UASVM 66, 366-372. [ Links ]
Pou, Α., Flexas, J., Alsina, M. del M., Bota, J., Carambula, C, De Herralde. F., Galmés, J., Lovisolo, C, Jimenez, M., Ribas-Carbó, Μ., Rusjan, D.. Secchi, F., Tomás, Μ., Zsófi, Ζ. & Medrano, Η., 2008. Adjustments of water use efficiency by stomatal regulation during drought and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri χ V. rupestris). Physiol. Plant 134(2), 313-323. [ Links ]
Pou, Α., Medrano, 1., Tomás, Μ., Martorell, S., Ribas-Carbó, Μ. & Flexas. J., 2012. Anisohydric behaviour in grapevines results in better performance under moderate water stress and recovery than isohydric behaviour. Plant Soil 359, 335-349. [ Links ]
Prieto, J.A., Lebon, E. & Ojeda, H., 2010. Stomatal behavior of different grapevine cultivars in response to soil water status and air water vapor pressure deficit. J. Int. Sei. Vigne Vin. 44, 9-20. [ Links ]
Rogiers, S., Greer, D.H., Hutton, R.J., Clarke, S.J. (2011) Transpiration efficiency of the grapevine cv. Semillon is tied to VPD in warm climates. AnnAppl Biol 158:106-114. doi:10.1111/j.l744-7348.2010.00446.x [ Links ]
Schultz, Η., 2003. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 26, 1393-1405. [ Links ]
Sgubin, G., Swingedouw, D., Dayon, G., Garcia de Cortázar-Atauri, I.. Ollat, N, Page, C. & Van Leeuwen, C, 2018. The risk of tardive frost damage in French vineyards in a changing climate. Agric. For. Meteorol. 250-251, 226-242. [ Links ]
Soar, C.J., Speirs, J., Maffel, S.M., Penrose, A.B., McCarthy, M.G. & Loveys, B.R., 2006. Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: Apparent links with ABA physiology and gene expression in leaf tissue. Aust. J. Grape Wine Res. 12, 2-12. [ Links ]
Tardieu, F. & Simonneau, T, 1998. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 49, 419-432. [ Links ]
Vandeleur, R.K., Mayo, G., Shelden, M.C., Gilliham, M., Kaiser, B.N. & Tyerman, S.D., 2008. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: Diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol. 149(1), 445-460. [ Links ]
Van Leeuwen, C. & Darriet, P., 2016. The impact of climate change on viticulture and wine quality. J. Wine Econ. 11, 150-167. [ Links ]
Van Leeuwen, C, Tregoat, O., Choné, X., Bois, Β., Pemet, D. & Gaudillère. J.P, 2009. Vine water status is a key factor in grape ripening and vintage quality for red Bordeaux wine. How can it be assessed for vineyard management purposes? J. Int. Sei. Vigne Vin. 43, 121-134. [ Links ]
Van Leeuwen, C, Trégoat, O., Choné, O., Gaudillère, J.-P. & Pernet, D., 2007. Different environmental conditions, different results: The role of controlled environmental stress on grape quality and the way to monitor it. In: Proc. XHIth Aust. Wine Ind. Tech. Conf, 28 July-2 August, Adelaide, Australia. [ Links ]
Verdugo-Vásquez, Ν., Acevedo-Opazo, C, Valdés-Gómez, Η., Ingram, Β.. Garcia de Cortazar, I. & Tisseyre, B., 2019. Towards an empirical model to estimate the spatial variability of grapevine phenology at the within field scale. Precis. Agric. 1-24. [ Links ]
West, A.G., Dawson, T.E., February, E.C., Midgley, G.F., Bond, W.J. & Aston, TL., 2012. Diverse functional responses to drought in a Mediterranean-type shrubland in South Africa. New Phytol. 195, 396-407. [ Links ]
Submitted for publication: September 2018
Accepted for publication: June 2019
* Corresponding author: E-mail address: hevaldes@uc.cl
Acknowledgements:


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}