










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.40 n.1 Stellenbosch 2019
ORIGINAL RESEARCH ARTICLES
Physiological Response of Three Grapevine Cultivars Grown in North-Western Poland to Mycorrhizal Fungi
G. MikiciukI, *; L. Sas-PasztII; M. MikiciukIII; E. DerkowskaII; P. TrzcinskiII; P. PtakIII; U. ChylewskaI; M. StatkiewiczI; A. LisekII
IDepartment of Horticulture, Faculty of Environmental Management and Agriculture, West Pomeranian University of Technology, Slowackiego 17, 71-434 Szczecin, Poland
IIDepartment of Microbiology, Research Institute of Horticulture, Pomologiczna 18, 96-100 Skierniewice, Poland
IIIDepartment of Plant Physiology and Biochemistry, Faculty of Environmental Management and Agriculture, West Pomeranian University of Technology, Slowackiego 17, 71-434 Szczecin, Poland
ABSTRACT
West Pomerania (Poland) is located near the northern boundary of the range of viticulture (the coldest zone A). Unfavourable weather conditions can pose a serious threat to the cultivated vines. One of the treatments used to increase the tolerance of plants to abiotic and biotic stresses is inoculation with symbiotic soil microorganisms. This paper focuses on the influence of mycorrhization on the changes in soil microbiology, the degree of colonization of roots by mycorrhizal fungi, and on selected physiological parameters of three grapevine cultivars ('Pinot Noir' on SO4 rootstock, 'Regent' on 5BB rootstock, and 'Rondo' on 125AA rootstock). The applied inoculation had a stimulating effect on the colonization of roots by arbuscular mycorrhizal (AM) fungi, as evidenced by higher mycorrhizal frequency and intensity in the mycorrhized plants. The mycorrhizal treatment increased the intensity of CO2 assimilation and transpiration. Mycorrhization reduced the efficiency of photosynthetic water use and increased stomatal conductance for water in the grapevines tested. The mycorrhizal treatment did not affect the concentration of assimilation pigments in vine leaves. The mycorrhization of grapevines had no effect on the values of initial fluorescence, maximum fluorescence, the maximum potential efficiency of photochemical reaction in PS II, the size of the pool of reduced electron acceptors in PS II, nor on the value of the PS II vitality index.
Key words: arbuscular mycorrhizal fungi, mycorrhizal frequency, physiological parameters, grapevine
INTRODUCTION
West Pomerania (Poland) is located near the northern boundary of the range of viticulture (the coldest zone A). On a global scale, Poland is currently an insignificant producer of wine. However, grape growing is becoming increasingly popular due to the good prices achieved by Polish wines (Stój et al., 2017). The most commonly grown cultivars of red grapes are 'Regent', 'Rondo' and 'Pinot Noir' (Wilk, 2011).
Mycorrhization of fruit plants is performed in order to improve vegetative growth, yield quality parameters, increase the tolerance of plants to abiotic and biotic stresses, and to reduce the use of chemicals in the environment. Because of these considerations, inoculation of plants with symbiotic soil microorganisms is a very important treatment used not only in organic farming, but also in integrated and conventional crop cultivation. The action of mycorrhizal fungi results in physical and chemical stabilization of the soil and practical possibility of limiting fertilization. Mycorrhizal mycelium increases the absorptive surface of roots and phosphorus availability to plants, which makes it a very important component of the rhizosphere of fruit plants (Sas Paszt et al., 2010). Mycorrhiza can indirectly affect the intensity of photosynthesis, and thus the productivity of plants, by increasing the stomatal conductance to CO2 and improving the efficiency of photochemical processes. This phenomenon is observed especially in the case of plant growth under stressful conditions (Borkowska, 2005; Wu and Zou, 2010).
Soil moisture is important for the mycorrhizal frequency in grapevine roots. The water content of the soil, weather conditions and its frequency or lack of irrigation significantly affects the mycorrhizal frequency in grapevine roots. Holland et al. (2014) had observed that irrigating every few days significantly increased the number and size of arbuscules found in the roots, while daily irrigation increase the number of vesicles. Donkó et al. (2014) studied the effect of soil moisture on the degree of colonization of grapevine roots by AM fungi. The most extensive colonization of roots by mycorrhizal fungi was shown by plants growing on the most elevated site, where there was never any stagnant water.
The fungal colonization there ranged from 64% to 81%, depending on the year and season. The lowest mycorrhizal frequency (from 46% to 76%) was found in the vines growing in a periodically flooded location. Water deficiency affects the mycorrhizal frequency in plant roots to a lesser extent than its excess, which is a factor that considerably restricts the colonization of roots (Deepika & Kothamasi, 2015). In addition, mycorrhizal frequency is also affected by soil pH, being lower at a low pH (Wang et al., 1993).
The aim of the study was to assess the influence of mycorrhization on the changes in soil microbiology, the degree of colonization of roots by mycorrhizal fungi, and on selected physiological parameters of three cultivars of red grapevines grafted onto different rootstocks, grown in the conditions of West Pomerania (Poland).
MATERIALS AND METHODS
The study was conducted in the Turnau vineyard near Baniewice (53°03'38'' N, 14°35'59" E) located in Northwestern Poland. A two-factor experiment was established in a random block design in three replications. One replication consisted of 5 plants planted at 2.5 χ 1 m. Grapevine plantlets were planted in a clayey-sandy soil in 2012. The soil pH in KCL was 6.10. Soil nutrient availabilities (mgT00g-1), determined in the soil samples collected from 0 to 60 cm of the soil depth, were: N-NH4 0.13, N-NO3 0.20, P 7.8, K 13.8 and Mg 5.5. In the first year, a multi-component fertilizer Suprofos 25 NPK (Ca, Mg, S) 5:10:25 (2.5:2:13) was applied at the dose of 400 kg/ha. In the second year, no mineral fertilization was used to fertilize the plants. The first experimental factor was the inoculation of plant roots with mycorrhizal fungi. Plants were treated with mycorrhizal substrate (produced by Mykoflor, Konskowola, Poland), containing 1000 propagules of arbuscular mycorrhizal fungi per 1 g of substrate: Rhizophagus irregularis, Glomus mosseae, Claroideoglomus etunicatum on an organic carrier medium, once with a special applicator for subsurface placement, in 2012, one month after planting. The inoculum vaccine was used in the form of an aqueous solution with a hydrogel, at a dose of 30 ml/plant (3000 propagules per plant). The following variants of the first factor were used: control, without mycorrhiza (variant M0), and with mycorrhiza (variant M1). The second experimental factor was the cultivars of red grapevines grafted onto different rootstocks. The following variants of the second factor were used: 'Pinot Noir' on SO4 rootstock (PNR variant), 'Regent' ('Diana' ('Silvaner' x 'Muller-Thurgau') x 'Chambourcin') on 5BB rootstock (REG variant), and 'Rondo' ('ZaryaSever' ('Seyanets Malengra' x V. amurensis) x 'St. Laureate') on 125AA rootstock (RON variant).
Microbiological analysis of soil
Soil samples were collected at the beginning of November. They were mixed thoroughly and 5 g of each sample was transferred to 100 mL Erlenmeyer flasks containing 45 g of sterile distilled water. The suspended samples were homogenized at 190 rpm for 45 minutes. A series of tenfold dilutions (10-2, 10-3 ... 10-5) were prepared from each suspension. Soil dry weight was determined by oven drying the soil samples at 90°C for four days.
Sterile Petri dishes were inoculated with 100 սL aliquots of each dilution prepared from the soil suspensions. The inoculated dishes were flooded with a liquid agar medium at a temperature of approx. 50°C. All analyses were made in triplicate. To estimate the number of microorganisms, the following microbial media were used: for estimation of the total number of culturable bacteria - 10% Tryptic Soy Agar (TSA) (Biocorp, cat. number: PS22); for estimation of the total number of spore-forming bacteria (to obtain the bacterial spores, the soil samples were incubated at 80°C for 30 minutes before being transferred into Petri dishes) - 10% TSA (Biocorp, cat. number: PS22); for estimation of the number of filamentous fungi - Rose Bengal Chloramphenicol Agar (Biocorp, cat. number: PS66); and for estimation of the number of fluorescent pseudomonades - S1 (Gould et al., 1984).
For estimation of total number of spore forming bacteria and microscopic fungi, inoculated plates were incubated at 26°C for 5-7 days. For evaluation of bacteria population, the inoculated plates was incubated at 26°C for 10-14 days and for evaluation of fluorescent pseudomonads, the inoculated Petri plates were incubated at 26°C for 3 days.
To estimate the number of bacterial or fungal colonies, Petri dishes containing between 30 and 300 colonies were selected. The results were calculated as colony forming units per 1 g of dry weight of soil (Schinnerk et al., 1995).
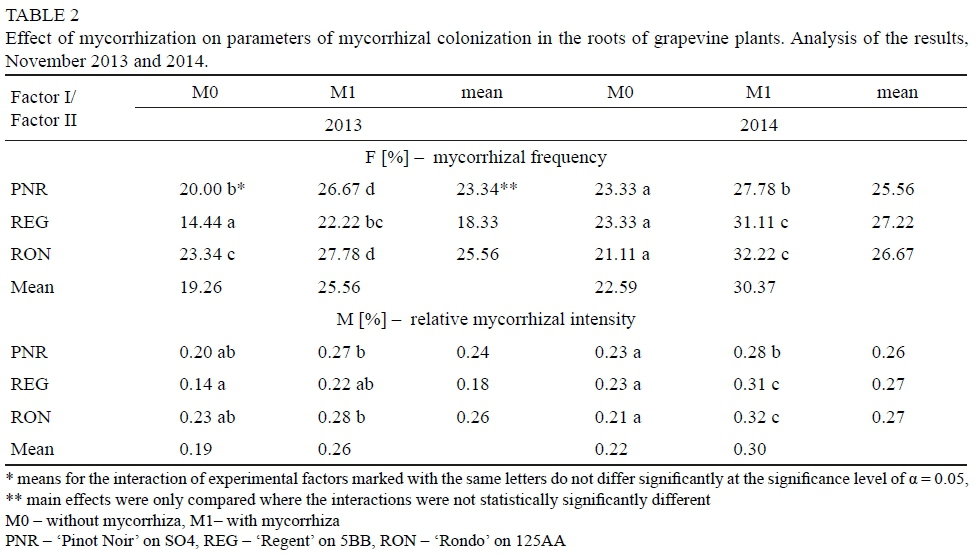
Assessment of root colonization by arbuscular mycorrhizal fungi The roots of grapevines (10 g from each replication), collected in November, were stained according to the method developed in the Department of Microbiology of the Research Institute of Horticulture (Derkowska et al., 2015). Microscopic specimens were prepared and examined with a Nikon 50i microscope (objectives with magnifications of 20χ, 40χ, 60χ, 100x), and photographic records of the observed mycorrhizal structures were produced. The assessment of the degree of colonization of the roots by arbuscular mycorrhizal fungi was performed by the Trouvelot method (Trouvelot et al., 1986). Based on the results, mycorrhizal frequency (F%), relative mycorrhizal intensity (M%) and absolute mycorrhizal intensity (m%) were calculated using the computer program MYCOCALC, available from the website: http://www2.dijon.inra.fr/mychintec/Mycocalcprg/MYCOCALC.EXE (Table 2).
Gas exchange parameters of plants The parameters of gas exchange of plants (CO2 assimilation intensity - A, transpiration - E, stomatal conductance for water - gs, and CO2 concentration in the intercellular spaces - ci) were measured twice in the growing season (at the veraison stage - 1st test date, and during fruit ripening - 2nd test date), with a TPS-2 (PP Systems) portable gas analyzer (with standard settings) equipped with a PLC4 measuring chamber operating in an open system. The results were read off the screen of the gas analyzer after the values had stabilized. The measurements were performed on healthy, fully grown vine leaves situated on the opposite side of the second or third cluster of grapes (counting from the tip of the shoot) in 12 replications (2 leaves per replication were analyzed, with each leaf coming from a different plant). On the basis of the results of CO2 assimilation intensity and transpiration, the photosynthetic water-use efficiency (ωW) was calculated, which was estimated by the ratio of assimilation intensity to transpiration (Candolfi-Vasconcelos & Koblet, 1991).
Concentrations of assimilation pigments in leaves
The amounts of chlorophyll 'a', 'b' and total chlorophyll in leaves were determined by the method of Arnon et al. (1956) modified by Lichtenthaler and Wellburn (1983), while the concentrations of carotenoids in leaves were determined by the method of Hager and Mayer-Berthenrath (1966). The concentration of assimilation pigments was determined in 6 replications on the same test dates and the same leaves on which gas exchange measurements were made. Extracts of the pigments were obtained by grinding samples of fresh leaf mass, about 0.05 g, in a mortar with 10 cm3 of 80% acetone. The homogenates were then centrifuged at 1500 rpm for 10 min. The optical density of samples was determined using a Marcel Mini spectrophotometer at wavelengths of λ = 440, 645 and 663 nm.
Chlorophyll fluorescence parameters
Chlorophyll fluorescence parameters were recorded in 2014, in the third year after mycorrhization, using a Handy PEA (Hansatech) spectrofluorometer, based on the standard apparatus procedure (3 x 650 nm LEDs, maximum actinic light intensity 3000 μmol. m-2 s-1). The measurements were made twice during the grapevine growing season, in 18 replications (3 leaves per replication were analyzed, each of them coming from a different plant), on the same test dates and the same leaves on which the other physiological characteristics were determined. Leaves were shaded 20 minutes before measurement with factory clips (illuminated area with a diameter of 4 mm). The following parameters of chlorophyll fluorescence induction were measured and calculated using the spectrofluorometer: F0 - initial fluorescence (zero), excitation energy loss index in power antennas; FM - maximum fluorescence, after reduction of acceptors in PS II and after dark adaptation; FV = FM - F0 - variable fluorescence, determined after dark adaptation, a parameter dependent on the maximum quantum yield of PS II; FV / FM - the maximum potential photochemical reaction efficiency in PS II determined after dark adaptation and after reduction of acceptors in PS II (Bolhár-Nordenkampf & Öquist, 1993); TFM - increase in time of chlorophyll fluorescence from the beginning of measurement to the maximum (FM); P I - PS II vitality index for the overall viability of this system; AM (Area) - surface area above the chlorophyll fluorescence curve and between F0 and FM points proportional to the size of the reduced plastoquinone acceptors in PS II (Kalaji & Loboda, 2007).
Statistical analysis of resultsThe results were statistically analyzed using multivariate analysis of variance in the system of random blocks. Multiple comparisons of the means for the combinations were performed with Tukey's test, at a significance level of α = 0.05, using STATISTICA v.10 software package. For the numerical data on the gas exchange parameters, concentration of assimilation pigments and chlorophyll fluorescence parameters, the analysis of variance was performed separately for each measurement date.
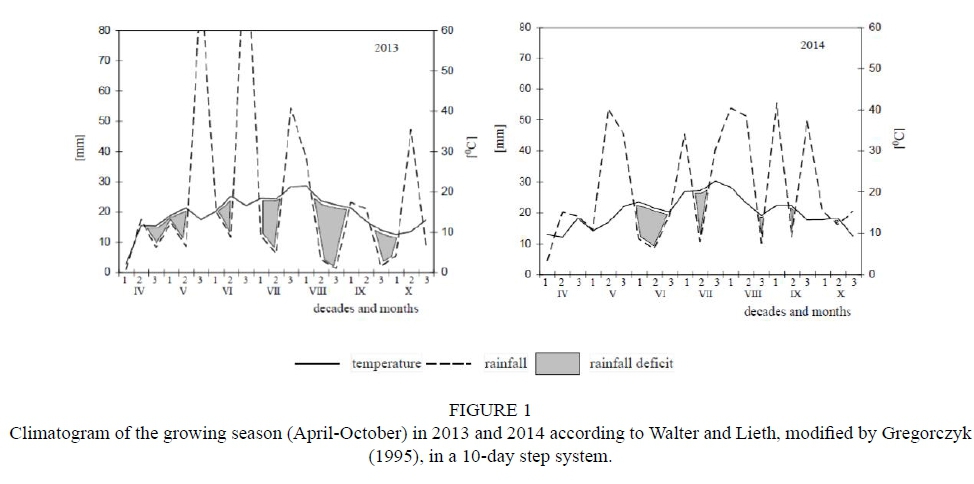
Meteorological conditions during the experiment During the growing season in 2013, the most rainfall was recorded in the period from around mid-May to the first 10 days of June and from around mid-June to the first 10 days of July. The greatest rainfall shortages occurred in the first 20 days of July, from around mid-August to the first 10 days of September, and from the last ten days of September to the first ten days of October. The weather conditions in 2013 were favourable for grapes to undergo veraison (colour change) and ripening. The year 2014 was characterized by considerable rainfall, practically throughout the entire growing season, with the exception of one prolonged period of rainfall deficit that occurred in June. Adverse weather conditions occurred both during the veraison and fruit ripening stages (Fig. 1). Meteorological data from the Szczecin-Dabie Meteorological Station (WMO 12205) were used.
RESULTS AND DISCUSSION
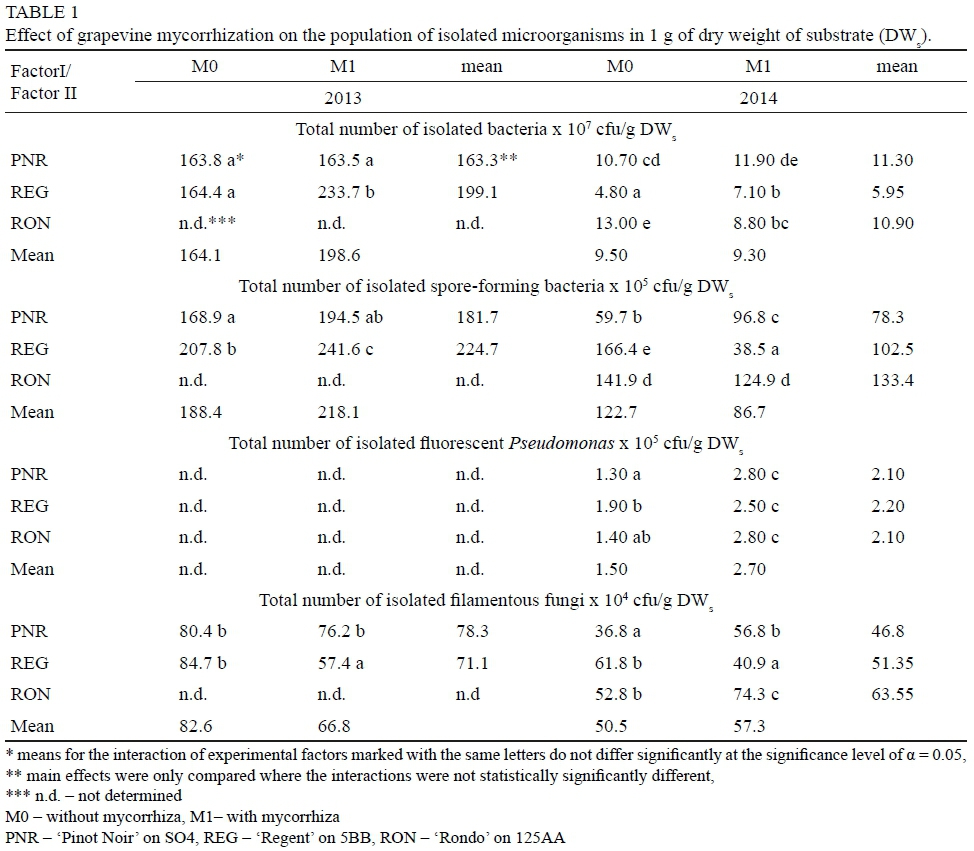
The applied treatments modified the bacterial population inhabiting the soil in the growing seasons of 2013 and 2014 (Table 1). In the cultivar 'Regent', mycorrhization was found to have an effect on increasing the total bacterial count in the soil in both years of the experiment. In the cultivar 'Rondo' in 2014, a reduction in the total number of bacteria in the soil was observed. In 2014, mycorrhization contributed to a significant reduction in the total number of spore-forming bacteria in the soil in the cultivar 'Regent'. In the case of the mycorrhized plants of the cultivar 'Pinot Noir', an increase in the number of spore-forming bacteria in the soil was observed in the second year of the experiment. Increased population of Pseudomonas bacteria in the mycorrhizal soil may indicate the existence of mutual interrelations between these two groups of microorganisms. The bacteria Pseudomonas fluoresceins are characterized by a high ability to colonize live mycorrhizal mycelium (Toljander et al., 2005). However, the explanation of the possible relationships between Pseudomonas bacteria and AM fungi requires further investigations (Table 1).
In the mycorrhized cultivar 'Regent', the total number of filamentous fungi in the soil was reduced in both years of the experiment. In 2014, an increase in the population of filamentous fungi was observed in the soil of the mycorrhized cultivars 'Pinot Noir' and 'Rondo'. In addition, there was a reduction in the total number of filamentous fungi in the 2014 growing season in comparison with the growing season of 2013 (Table 1). In 2014, a positive effect of mycorrhization on increasing the population of Pseudomonas bacteria was observed in all grapevines varieties. The obtained results indicate that the changes in soil microbiology were influenced by several factors, including mycorrhization, grapevine cultivar and weather conditions prevailing during the growing season. In 2014, a smaller overall number of isolated bacteria and filamentous fungi was observed (Table 1). This phenomenon could have been related to the meteorological conditions because in 2014 there was much more rainfall, compared with 2013, when rainfall shortages were observed.
In the soil environment, there are many interactions between arbuscular mycorrhizal fungi and bacteria, which can be of importance in agriculture, such as adherence of bacterial cells to fungal spores, production of volatile substances by bacteria and degradation of fungal cell walls (Miransari, 2011). Arbuscular mycorrhizal fungi and soil bacteria can interact synergistically and stimulate plant growth by facilitating the uptake of nutrients by plants and by reducing fungal soil pathogens. Such interactions are of great importance in sustainable agricultural systems based on the use of biological processes to maintain soil fertility and good plant health. Although there have been many studies on the interaction between arbuscular mycorrhizal fungi and bacteria, including fluorescent Pseudomonads, spore-forming bacteria and filamentous fungi, the mechanisms of interaction are not well understood and require further experimentation to optimize the composition of microbial consortia for use in agriculture (Artursson et al., 2006).
After inoculation with the biopreparation Mykoflor, more extensive colonization of roots by arbuscular mycorrhizal fungi was observed in all the cultivars tested, both in the first and second year of the study. The authors mainly observed numerous mycorrhizal vesicles in the grapevine plant roots, which are documented in the photos (Fig. 2-7). The stimulating effect of the biopreparation Mykoflor on the colonization of roots by AM fungi is evidenced by the higher mycorrhizal frequency and intensity observed in the mycorrhized plants during the two years of the experiment (Table 2). Absolute mycorrhizal intensity shows no significant differences between cultivars during the duration of the experiment. The degree of root colonization by mycorrhizal fungi after inoculation, in 2013 and 2014, was the highest for the cultivar 'Rondo' (combination M1RON) and was F% 27.78 and F% 32.22, respectively (Table 2). The lowest mycorrhizal frequency in the inoculated plants in the first year of the experiment was observed in the roots of 'Regent' grapevines (F% 22.22), and in the second year of the experiment in the roots of 'Pinot Noir' grapevines (F% 27.78). The degree of colonization of grapevine roots by arbuscular mycorrhizal fungi was higher in the second year of the study. Despite the stimulating effect of the biopreparation Mykoflor on the colonization of grapevine roots by arbuscular mycorrhizal fungi, the observed mycorrhizal frequency was low (22.22% to 32.22%) compared with higher values of mycorrhizal frequency for grapevine plants observed by Petit & Gubler (2006). These authors observed colonization of roots at 48.3% in Vitis rupestris grapevine plants inoculated with the fungus Rhizophagus intraradices, and at 54.5% in plants inoculated with R. intraradices and the pathogenic fungus Cylindrocarpon macrodidymum. A similar, low mycorrhizal frequency had been recorded for apple trees (6.6% to 36.7%) growing in the Pomological Orchard of the Research Institute of Horticulture in Skierniewice (Derkowska et al., 2013). A low mycorrhizal frequency had also been observed in the roots of apple trees (2.5% to 25.8%) and blackcurrant plants (0 to 11.1%) treated with organic mulches, growing in the same research facility (Sumorok et al., 2011). The degree of colonization of plant roots by AM fungi may be due to soil properties, such as phosphorus content and competition from other soil microorganisms (Treseder, 2013; van Overbeek & Saikkonen, 2016).
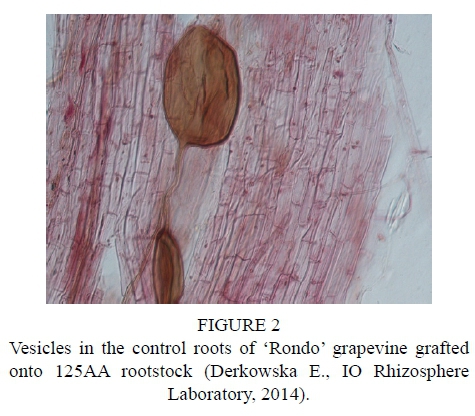
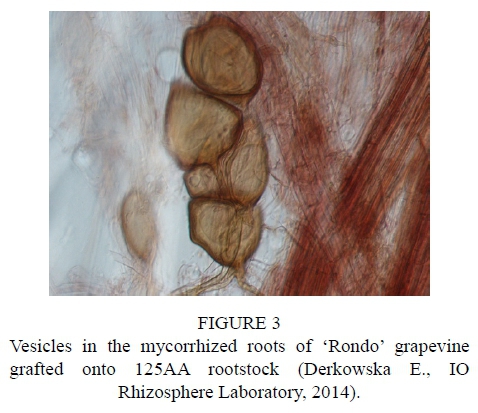
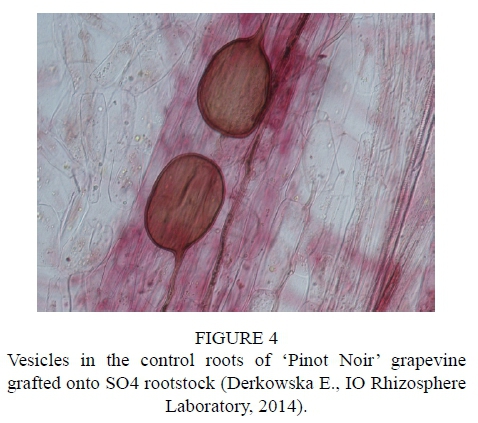
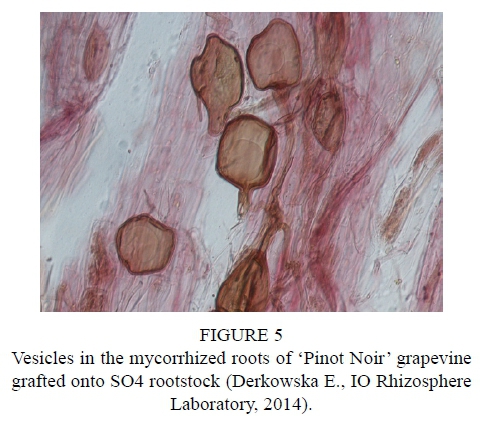
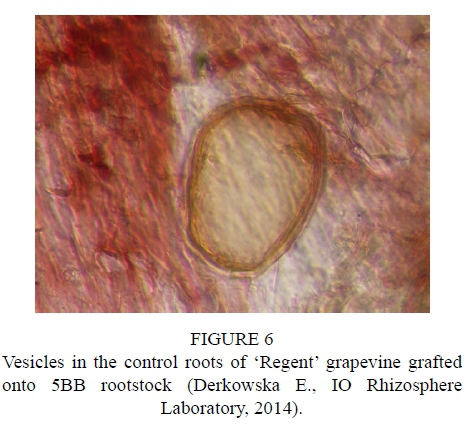
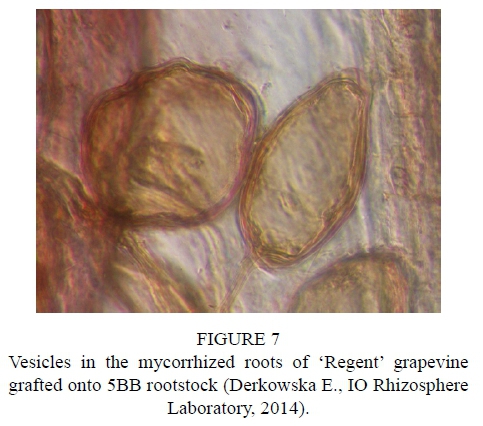
An important factor affecting mycorrhizal frequency in the roots of grapevine plants is the type of rootstock used. Karagiannidis et al. (1997) observed that mycorrhizal frequency, the number of mycorrhizal structures in the roots and the number of spores in the substrate depended on the rootstock used and on the cultivar grafted onto it. In their experiments, they examined 4 rootstocks onto which 9 grapevine cultivars were grafted. They found the highest mycorrhizal frequency, from 68% to 87%, in the roots of rootstock 1103 P, while the values determined for other rootstocks were in the ranges: 36% to 55% for 110 R, 41% to 62% for 140 Ru, and 57% to 74% for the 41B rootstock. Similar relations were noted in the number of observed spores, which occurred in the largest numbers in the rhizosphere of the 1103 P rootstock, with the lowest numbers found in the rhizosphere of the 110 R rootstock. In the tests conducted in 2013, the highest average mycorrhizal frequency was found for the cultivar 'Rondo' on the 125AA rootstock (25.56%) and for the cultivar 'Pinot Noir' on the SO4 rootstock (23.34%), while the lowest mycorrhizal frequency was observed for the cultivar 'Regent' on the 5BB rootstock (18.33%) (Table 2). In 2014, no statistically significant differences in the average mycorrhizal frequency were found among the cultivars.
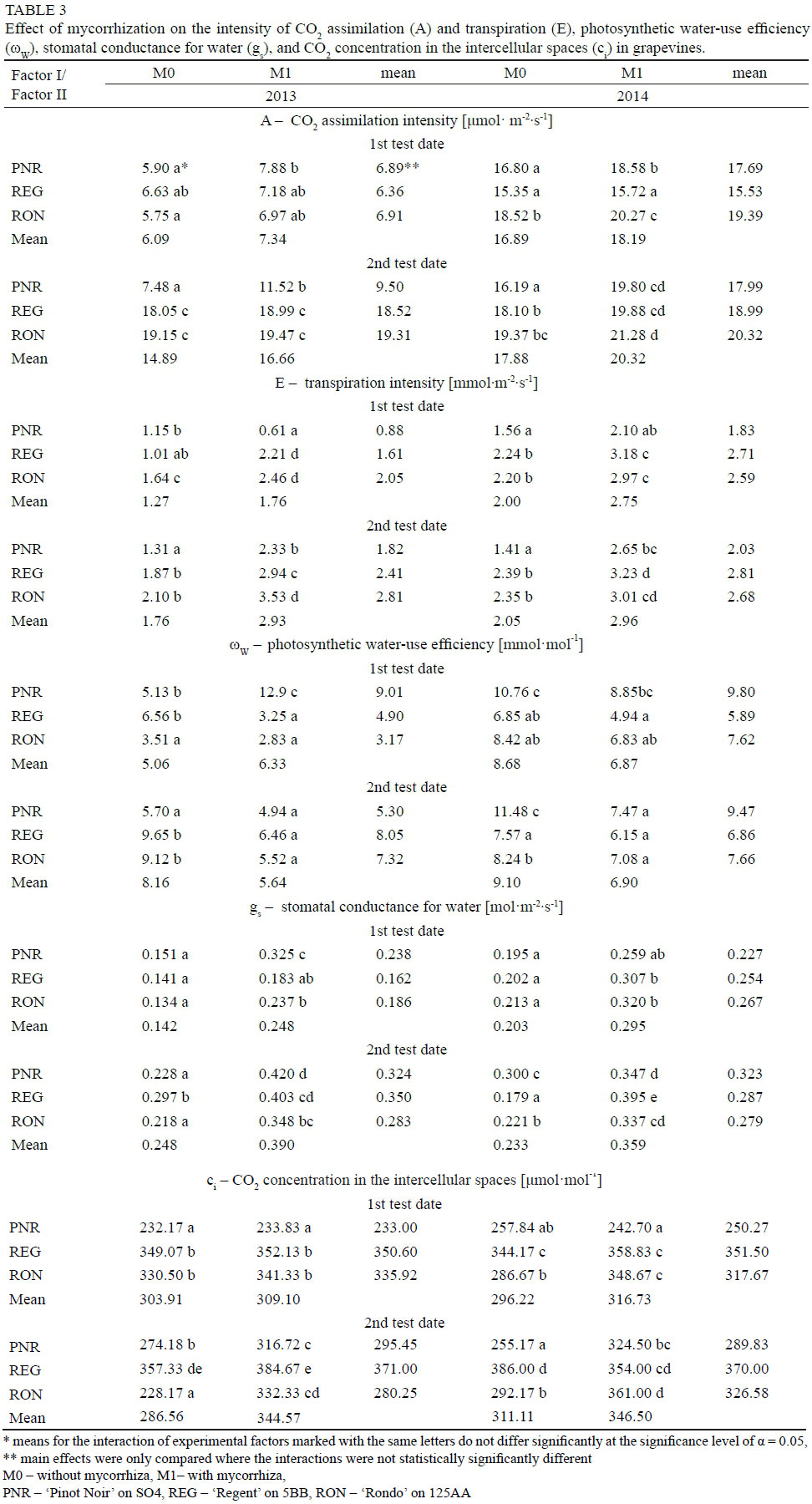
According to Borkowska (2010), the phenomenon of mycorrhiza does not directly affect the functioning of processes, such as electron transport during the light phase of photosynthesis and regeneration of the Rubisco enzyme involved in binding CO2, responsible for the activity of the photosynthetic apparatus of plants. Nevertheless, it can affect them indirectly by providing plants with water and a better supply of phosphorus necessary, inter alia, in photosynthetic phosphorylation. In addition, the plant receives nitrogen and magnesium through the fungus, and also microelements such as iron, copper and manganese - these elements perform important functions in photosynthesis (Kaschuk et al., 2009). The current study showed that the mycorrhized grapevine plants were characterized by a greater CO2 assimilation intensity - this relationship was found on one of the measurement dates in each year of the study. Similarly, according to Zhu et al. (2012), mycorrhization with the species Claroideoglomus etunicatum increased the rate of net photosynthesis in maize plants growing in the conditions of both drought and optimal soil moisture. An increase in the intensity of CO2 assimilation in grapevines of the cultivar 'Crimson' and Citrus tangerina under the influence of mycorrhization with fungi of the genus Glomus was also demonstrated by Nicolas et al. (2015) and Wu & Xia (2006), respectively.
In the first year of the study, on the second measurement date, the highest intensity of CO2 assimilation was shown by the grapevines 'Rondo' and 'Regent'. In the second year of the study, the highest intensity of CO2 assimilation was also observed in the cultivar 'Rondo' on the second measurement date, compared to 'Pinor Noir'. After analyzing the interaction of the experimental factors, it can be stated that in the first year of the study, on the second measurement date, the highest intensity of CO2 assimilation was shown by the grapevines of the cultivars 'Rondo' and 'Regent', both the mycorrhized and control ones. In the second year of the study, the highest intensity of this process was exhibited by the grapevines from the combination M1RON (on the second measurement date it was significantly different from the intensity found in the combinations M0RON, M0REG and M0PNR) (Table 3).
The vines of the cultivars 'Regent' and 'Rondo' showed the highest intensity of transpiration. The cultivar 'Pinot Noir' conducted transpiration at a level similar to that of the cultivar 'Regent', but only on the second measurement date of the first year of the study. In both years of the study, the highest intensity of transpiration was exhibited by vines of the cultivars 'Rondo' and 'Regent' subjected to mycorrhization (Table 3). The inoculation with mycorrhizal fungi increased the intensity of transpiration in the vines. Similar results had also been obtained by Wu & Xia (2006) and Zhu et al. (2012) in studies on the impact of mycorrhization with fungi of the genus Glomus on the intensity of transpiration in species such as Citrus tangerina and maize, respectively.
The plants subjected to mycorrhization were characterized by lower photosynthetic water-use efficiency - this correlation was proved on the second test date of the first year and on both test dates of the second year of the study. Comparing the tested grapevine cultivars with respect to this trait, it was found that the highest value of this index on one of the measurement dates of both years of the study was shown by the cultivar 'Pinot Noir'. On the second test date of the first year of the study, this was shown by the cultivars 'Regent' and 'Rondo', while on the first test date of the second year by the cultivars 'Pinot Noir' and 'Rondo'. The interaction of the experimental factors exerted a multidirectional effect, depending on the measurement date, on the efficiency of water use in photosynthesis in the tested grapevine cultivars (Table 3).
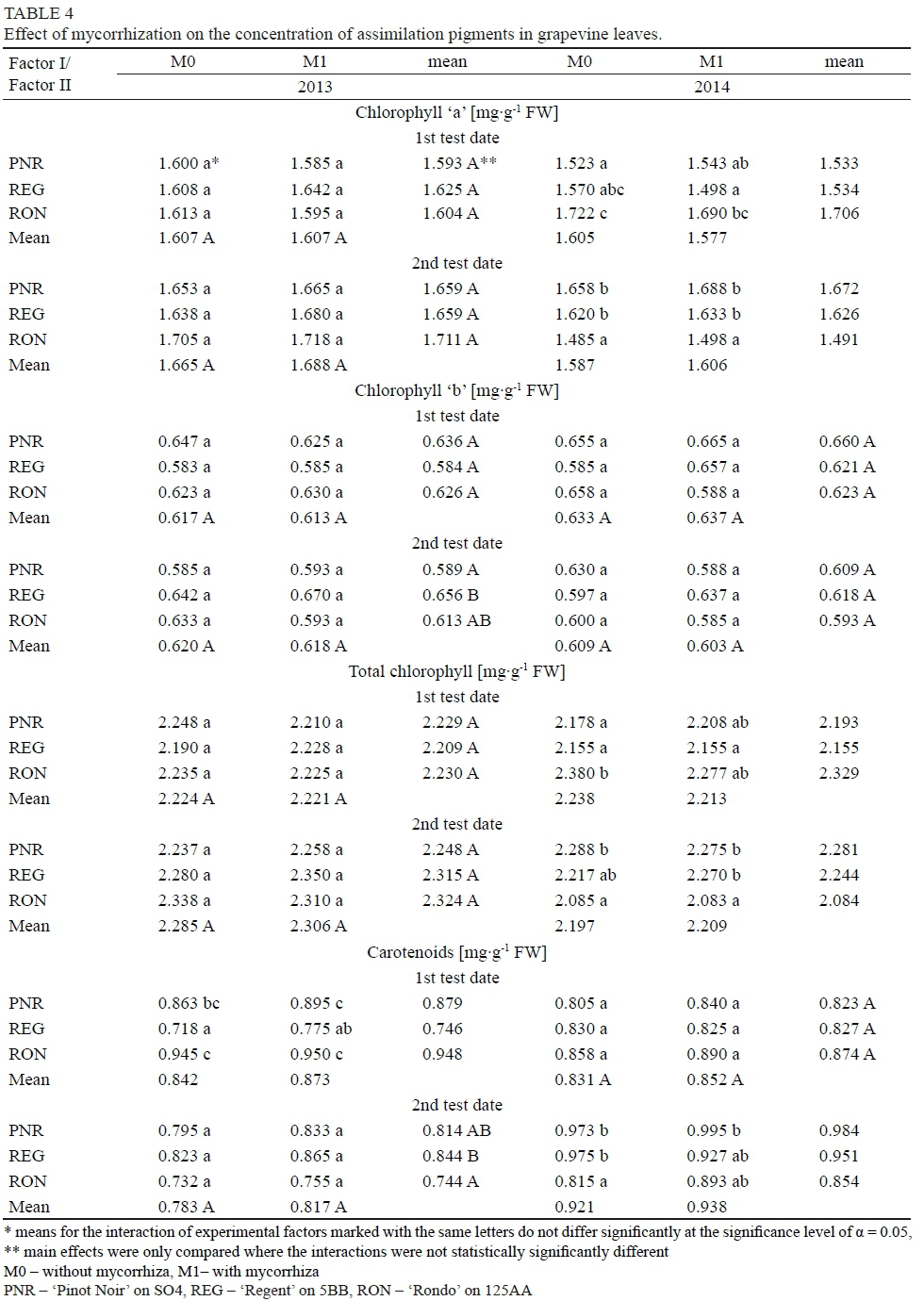
The study proved that mycorrhization of grapevine plants increased stomatal conductance for water - this correlation was recorded on both measurement dates in the first year of the study and on the first test date in the second year. The grapevine cultivars under comparison did not differ in terms of this physiological characteristic. After analyzing the interaction of the experimental factors, it was found that the inoculation with mycorrhizal fungi increased the stomatal conductance for water, determined in the two consecutive years of the study, on the first measurement date, in the grapevine cultivars 'Pinot Noir' and 'Rondo', and 'Regent' and 'Rondo', respectively. On the second test date, this relationship was demonstrated in both years of the study, for all the cultivars tested (Table 3). Similar results had been obtained by Wu & Xia (2006), Nicolas et al. (2015) and Zhu et al. (2012), according to which mycorrhization with fungi of the genus Glomus increased the stomatal conductance for water in the leaves of Citrus tangerine, the grapevine cultivar 'Crimson', and maize plants growing in drought conditions (no such relationship was found for maize grown under optimal conditions). On the first measurement date of testing, the concentration of CO2 in the intercellular spaces of leaves was demonstrated to be higher in the grapevines subjected to inoculation with mycorrhizal fungi than in the control. Comparing the tested grapevine cultivars, it was found that the highest concentration of intercellular CO2 was shown by the vine leaves of the cultivars 'Regent' and 'Rondo' (on the first test date of the first year of the study) and the cultivar 'Regent' (on the second test date of the first year and in the second year of the study). The smallest value of this trait, however, was shown by the grapevine cultivar 'Pinot Noir' on the first test date of the first year of the study and on both test dates of the second year. After analyzing the interaction of the experimental factors, it was found that the highest concentration of CO2 in the intercellular spaces of leaves in the first year of the study was shown by the combinations M1REG, M0REG, M1R0N and M0RON (1st date), and M1REG (2nd date). In the second year of the study, however, it was the combinations M1REG, M1RON and M0REG (1st date), and M0REG and M1RON (2nd date) - Table 3. According to Zhu et al. (2012), inoculation with the mycelium of Claroideoglomus etunicatum reduced the CO2 concentration in the intercellular spaces of leaves of maize plants growing both under the conditions of optimal soil moisture content and in drought conditions. The study presented here did not find a significant effect of mycorrhization on the concentration of the estimated assimilation pigments, i.e. chlorophyll 'a', 'b', total chlorophyll and carotenoids (Table 4). In the first year of the study, no differences were found in chlorophyll 'a' content and total chlorophyll in the leaves of the grapevine cultivars under comparison. On the second test date, a difference in chlorophyll 'b' content was found between the cultivar 'Regent', in which it was the highest, and the cultivar 'Pinot Noir'. The tested grapevine cultivars were characterized by a varied concentration of carotenoids in the leaves, which depended on the test date and year of the study. On the first test date in the first year of the study, the lowest concentration of these pigments was shown by the cultivar 'Regent', and the highest by 'Rondo'. On the second test date in the second year of the study, the cultivars 'Regent' and 'Pinot Noir' surpassed the cultivar 'Rondo' in terms of the carotenoid content. In the first year of the study, there was no significant effect of the interaction of the experimental factors on the amounts of chlorophyll 'a', 'b' and total chlorophyll in grapevine leaves. On the first test date in the second year of the study, the effect on the amount of chlorophyll 'a' and total chlorophyll was significant, but ambiguous. On the second test date of that year, the highest chlorophyll 'a' content was recorded in the mycorrhized and control plants of the cultivars 'Pinot Noir' and 'Regent'. In the case of total chlorophyll, there was a proven difference between the amount of this pigment in plant leaves from the combinations M0PNR, M1PNR, M1REG and M0PNR, where it was the largest, and the combinations M0RON and M1RON. On that test date, however, the highest amounts of carotenoids were found in the combinations M1PNR, M0REG and M0PNR. Different results had been obtained by Aslanpour et al. (2016), according to which the inoculation of white grapevines with the fungus Glomus mosseae increased the total chlorophyll content in the leaves. However, the authors did not demonstrate any differences in the chlorophyll content in the leaves of the tested grapevine cultivar depending on the species of fungi (Glomus mosseae, Glomus fasciculatum, Glomus intraradies) used for mycorrhization. Increased amounts of chlorophyll 'a', 'b' and total chlorophyll as a result of mycorrhization with the species Claroideoglomus etunicatum had also been found by Zhu et al. (2012) in the leaves of maize plants growing under the conditions of drought stress - such correlations were not observed in plants growing in a soil with an optimal moisture content. According to Smith & Read (2008), the increase in the chlorophyll content of the leaves of plants inoculated with mycorrhizal fungi may be caused by the increased uptake of phosphorus.
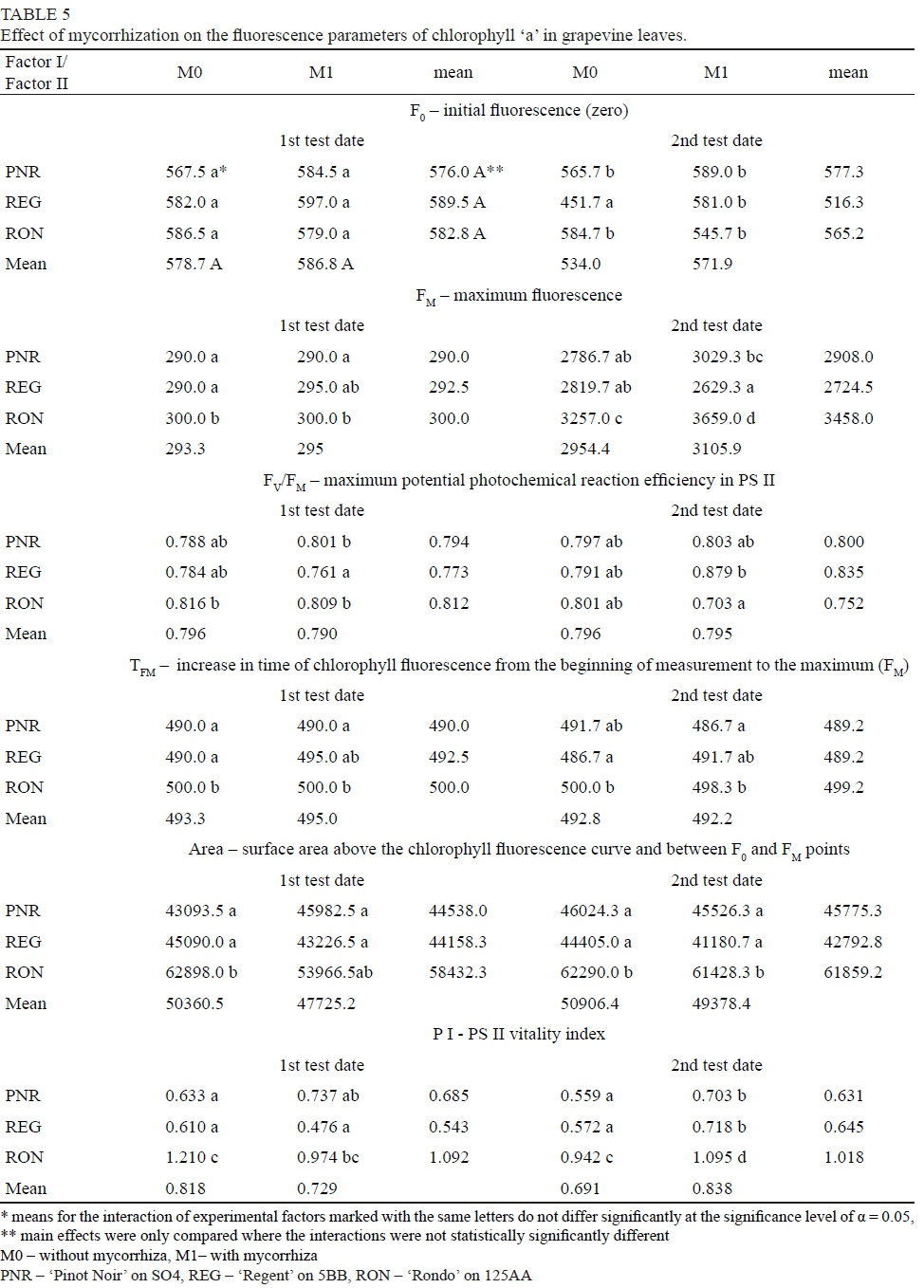
On both test dates, there was no effect of the mycorrhizal treatment on the size of the F0 parameter (initial fluorescence after dark adaptation) in the leaves of the vines, and also the studied grapevine cultivars did not differ in respect of this trait. On the second test date, the smallest F0 value was shown by the cultivar 'Regent' not subjected to mycorrhization, which may indicate that its photosynthetic apparatus, in comparison with plants from other combinations, was characterized by a higher efficiency of excitation energy transfer between chlorophyll molecules (Baker & Risenquist, 2004). The inoculation with mycorrhizal fungi did not affect the value of the FM parameter, i.e. the maximum fluorescence after dark adaptation. After comparing the tested grapevine cultivars in respect of this trait, it was found that the highest FM value was shown by the leaves of the cultivar 'Rondo', which may indicate that in the other cultivars, due to stress, not all electron acceptors in PS II had been completely reduced (Kalaji & Loboda, 2010). On the first measurement date, the highest FM values (significantly different from all the combinations, except M1REG) were characterized by the vines of the cultivar 'Rondo', both the mycorrhized and control ones. On the second test date, however, it was the cultivar 'Rondo' subjected to fungal inoculation (Table 5). The maximum potential efficiency of photochemical reaction in PS II determined after dark adaptation (FV/FM) is a parameter considered to be a measure of the photochemical activity of the photosynthetic apparatus of plants and describes the efficiency with which PS II absorbs light energy. Under optimal growth conditions, it takes on values of approximately 0.85 relative units, according to Angelini et al. (2001) and Bjorkman & Demmig (1987). The tested grapevine cultivars, both those under mycorrhization and the untreated control, were characterized, on both measurement dates, by lower than optimal values of this index, which may have been due to the excessive rainfall that was recorded in that year of the study (Table 5). Lowering of the FV/FM value under the influence of excessive soil water content and root flooding had been demonstrated by Bertolde et al. (2012) and Yu et al. (2015) in one of the cultivars of the cocoa tree (Theobroma cacao) and the Euphrates poplar (Populus euphratica), respectively. On the first measurement date, higher values of the FV/FM parameter were shown by the cultivars 'Rondo' and 'Pinot Noir'. In the case of all the cultivars tested, no significant effect of mycorrhization on the maximum potential efficiency of the photochemical reaction in PS II was demonstrated. Comparing the values of this index determined on the first measurement date for the individual grapevine cultivars subjected to mycorrhization, it was found that they were the smallest for the cultivar 'Regent' (Table 5).
During the veraison stage in the grapevine cultivars tested, a higher value of the TFM index was found in the plants under mycorrhization, which may indicate that they were in a state of stress for longer in comparison with the control. Similarly, when comparing the cultivars, a longer time to reach the maximum chlorophyll fluorescence was recorded on both measurement dates in the cultivar 'Rondo'. Analyzing the influence of the interaction of the experimental factors on the TFM parameter, its highest values, significantly different from all the experimental combinations except M1REG (in the first measurement date) and M0PNR, M1REG (in the second measurement date), were found in combinations M0RON and M1RON (Table 5).
The mycorrhizal treatment did not affect the value of the Area index, i.e. the size of the pool of reduced electron acceptors in PS II, which is, according to Kalaji & Loboda (2007), one of the best performance indicators of the photosynthetic apparatus of plants; nor on the PS II (P I) vitality index describing the overall vitality of this photosystem. The highest values of these parameters of chlorophyll fluorescence, on both measurement dates, were shown by the cultivar 'Rondo', which, together with the TFM values determined for this cultivar, may indicate a high efficiency of its repair mechanisms under stressful conditions. On the first measurement date, the highest value of Area, significantly different from all the combinations except M1RON, was found in the leaves of the vines from the combination M0RON. On the second test day, however, it was the combinations M0RON and M1RON. According to Krause & Weiss (1991), the Area parameter informs about the number of acceptors available in PS II. On the first measurement date, the highest value of the P I index (significantly different from all the combinations except M1RON) was found in the cultivar 'Rondo' not subjected to mycorrhization. On the second measurement date, however, it was the mycorrhized cultivar 'Rondo'. On that date, the measurements also showed higher values, for all the cultivars, of the PS II vitality index after inoculation with mycorrhizal fungi (Table 5).
CONCLUSIONS
The number of isolated microorganisms in the soil depended mainly on the weather conditions prevailing during the growing season. The considerably greater amount of rainfall in the second year of the study limited the numbers of all the soil microorganisms determined in both growing seasons. In the individual years of the study, the changes in soil microbiology were also influenced by the mycorrhizal treatment, the type of rootstock and the cultivar grafted onto it. The applied inoculation had a stimulating effect on the colonization of roots by AM fungi, as evidenced by higher mycorrhizal frequency and intensity in the mycorrhized plants. The influence of the applied rootstock and the cultivar grafted onto it on mycorrhizal frequency was apparent only in the first year of the study.
The inoculation of the roots of the tested grapevine cultivars with mycorrhizal fungi increased the intensity of CO2 assimilation (on one of the measurement dates, in each year of testing), transpiration and stomatal conductance for water in leaves. Mycorrhization reduced the efficiency of water use in photosynthesis and did not affect the concentration of assimilation pigments and such chlorophyll fluorescence parameters as F0, FM FV/FM and PI in grapevine leaves. The highest values of the Area index and the PI index were shown by the cultivar 'Rondo', which, together with TFM values (increase in time to achieve the maximum chlorophyll fluorescence) determined for this cultivar, may indicate a high efficiency of its repair mechanisms within its photosynthetic apparatus under stressful conditions. The obtained research results indicate that, among the tested cultivars, 'Rondo' is the most adapted to growing in the coldest zone (A) of viticulture. Beneficial effects of the mycorrhization on some physiological features of the examined grapevine varieties, including, inter alia, intensity of CO2 assimilation, which determine plant productivity, may indicate usefulness of mycorrhizal inoculation in viticulture.
LITERATURE CITED
Angelini, G., Ragni, P., Esposito, D., Giardi, P., Pompili, M.L., Moscardelli, R. & Giardi, M.T., 2001. A device to study the effect of space radiation on photosynthetic organisms. Phys. Med. 17, 267-268. [ Links ]
Arnon, D.J., Allen, M.B. & Whatley, F., 1956. Photosynthesis by isolated chloroplast. Biochim. Biophys. Acta. 20, 449-461. [ Links ]
Artursson, V., Finlay, R.D. & Jansson, J.K., 2006. Interactions between arbuscular mycorrhizal fungi and bacteria and their potential for stimulating plant growth. Environ. Microbiol. 8, 1-10. [ Links ]
Aslanpour, M., Baneh, H.D., Tehranifar, A. & Shoor, M., 2016. The effect of micorrhized fungi on the amount of glycine betaine, soluble sugar, proline, leaf water content and leaf chlorophyll of the white seedless grape under drought stress conditions. Inter. J. Adv. Biotechnol. Res. 7, 1119-1133. [ Links ]
Baker, N.R. & Resenquist, E., 2004. Application of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 55, 1607-1621. [ Links ]
Bertolde, F.Z., Almeida A.A., Pirovani C.P., Gomes F.P., Ahnert D., Baligar V.C. & Valle R.R., 2012. Physiological and biochemical responses of Theobroma cacao L. genotypes to flooding. Photos. 50, 447-457. [ Links ]
Bjorkman, O. & Demmig, B., 1987. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta. 170, 489-504. [ Links ]
Bolhár-Nordenkampf, H.R. & Öquist, G., 1993. Chlorophyll fluorescene as a total in photosynthesis research. In: Hall, D.O., et al. (eds). Photosynthesis and production in a changing environment. Chapman and Hall, London. 193-206. [ Links ]
Borkowska, B., 2005. The photosynthetic activity of micropropagated strawberry plants inoculated with endomycorrhizal fungi (AMF) and growing under drought stress. Acta Physiol. Plant. 24, 365-370. [ Links ]
Borkowska, B., 2010. Fizjologia Roslin sadowniczych strefy umiarkowanej t.1. In Jankiewicz, L. & Lipecki, J. (eds). Mikoryza, PWN Warszawa. 224250. [ Links ]
Candolfi-Vasconcelos, M.C. & Koblet, W., 1991. Influence of partial defoliation on gas exchange parameters and chlorophyll content of field-grown grapevines - Mechanisms and limitations of the compensation capacity. Vitis. 30, 129-141. [ Links ]
Deepika, S. & Kothamasi, D., 2015. Soil moisture - a regulator of arbuscular mycorrhizal fungal community assembly and symbiotic phosphorus uptake. Mycorrhiza. 25, 67-75. [ Links ]
Derkowska E., Sas Paszt, L., Dyki, B. & Sumorok, B., 2015. Assessment of mycorrhizal frequency in the roots of fruit plants using different dyes. Adv. Microbiol. 5, 54-64. [ Links ]
Derkowska, E., Sas Paszt, L., Sumorok, B. & Dyki, B., 2013. Colonisation of apple and blackcurrant roots by arbuscular mycorrhizal fungi following mycorrhisation and the use of organic mulches. Folia Hort. 25, 117-122. [ Links ]
Donkó, Á., Zanathy, G., Èros-Honti, Z., Villangó, S. & Bisztray, G.D., 2014. Changes of mycorrhizal colonization along moist gradient in a vineyard of Eger (Hungary). Acta Universitatis Sapientiae Agric. Env. 6, 13. [ Links ]
Gould, W.D., Hagedorn, C., Bardinelli, T.R. & Zablotowicz, R.M., 1984. New selective media for enumeration and recovery of fluorescent Pseudomonads from various habitats. Applied Environ. Microbiol. 49, 28-32. [ Links ]
Gregorczyk, A., 1995. O modyfikacji klimatogramów Waltera i Lietha. Zesz. Nauk. AR Szczec. Ser. Rolnictwo. 167, 29-33. [ Links ]
Hager, A. & Mayer-Berthenrath, T., 1966. Die Isolierung und quanttaive Bestimung der Carotenoide und Chlorophyll von Blatern, Algea und isolierten Chloroplasten mit Hilfe Dunnschichtchromatographischer Methoden. Planta. 69, 198-217. [ Links ]
Holland, T.C., Bowen, P., Bogdanoff, C. & Hart, M., 2014. Arbuscular mycorrhizal fungal communities associated with Vitis vinifera vines under different frequencies of irrigation. Am. J. Enol. Vitic. 65, 222-229. [ Links ]
Kalaji, H.M. & Loboda T., 2007. Photosystem II of barley seedlings under cadmium and lead stress. Plant Soil Environ. 53, 511-516. [ Links ]
Kalaji, H.M. & Loboda, T., 2010. Fluorescencja chlorofilu w badaniach stanu fizjologicznego roslin. Wydawnictwo SGGW, Warszawa. [ Links ]
Karagiannidis, N., Velemis, D. & Stavropoulos, N., 1997. Root colonization and spore population by VA-mycorrhizal fungi in four grapevine rootstocks. Vitis. 36, 57-60. [ Links ]
Kaschuk, G., Kuyper, T.W., Leffelaar, P.A., Hungria, M. & Giller, K.E., 2009. Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses? Soil Biol. Bioch. 41, 1233-1244. [ Links ]
Krause, G.H. & Weiss, E., 1991. Chlorophyll fluorescence and photosynthesis: the basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 42, 313-349. [ Links ]
Lichtenthaler, H.K. & Wellburn, A.R., 1983. Determinations of total carotenoids and chlorophyll a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 11, 591-592. [ Links ]
Miransari, M., 2011. Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl. Microbiol. Biotechnol. 89, 917-930. [ Links ]
Nicolas, E., Maestre-Valero, J.F., Alarcón, J.J., Pedrero, F., Vincente-Sánchez, J., Bernabé, A., Gómez-Montiel, J., Hernandez, J.A. & Fernandez, F., 2015. Effectiveness and persistence of arbuscular mycorrhizal fungi on the physiology, nutrient uptake and yield of Crimson seedless grapevine. J. Agric. Sci. 153, 1084-1096. [ Links ]
Petit, E. & Gubler, W.D., 2006. Influence of Glomus intraradices on Black Foot disease caused by Cylindrocarpon macrodidymum on Vitis rupestris under controlled conditions. Plant Dis. 90, 1481-1484. [ Links ]
Sas Paszt, L., Malusa, E., Grzyb, Z., Rozpara, E., Wawrzynczak, P., Rutkowski, K.P., Zmarlicki, K., Michalczuk, B., Podlaska, B. & Nowak, D., 2010. Srodowiskowe i zdrowotne znaczenie ekologicznej produkcji owoców. Post. Nauk Rol. 1, 109-121. [ Links ]
Schinnerk, F., Ohlingerk, R., Kandelerk, E. & Margesin, R., 1995. Methods in Soil Biology. Springer-Verlag, New York. [ Links ]
Smith, S. E. & Read, D.J., 2008. (3rd ed). Mycorrhizal symbiosis. Academic Press, Elsevier. [ Links ]
Stój, A., Czernecki, T., Domagala, D. & Targonski, Z., 2017. Application of volatile compound analysis for distinguishing between red wines from Poland and from other European countries. S. Afr. J. Enol. Vitic. 38, 245263. [ Links ]
Sumorok, B., Sas Paszt, L., Gluszek, S., Derkowska, E. & Zurawicz, E., 2011. The effect of mycorrhization and mulching of apple trees 'Gold Milenium' and blackcurrant bushes 'Tiben' on the occurrence of arbuscular mycorrhizal fungi. J. Fruit Ornam. Plant Res. 19, 35-49. [ Links ]
Toljander, J.F., Artursson, V., Paul, L.R., Jansson, J.K. & Finlay, R.D., 2005. Attachment of different soil bacteria to arbuscular mycorrhizal fungal extraradical hyphae is determined by hyphal vitality and fungal species. FEMS Microbiology Letters. 254, 34-40. [ Links ]
Treseder, K.K., 2013. The extent of mycorrhizal colonization of roots and its influence on plant growth and phosphorus content. Plant and Soil. 371, 1-13. [ Links ]
Trouvelot, A., Kough, J.L. & Gianinazzi-Pearson, V., 1986. Mesure du taux de mycorhization VA d'un systeme radiculaire. Recherche de methods d'estimation ayant une signification fonctionnelle. In: Gianinazzi-Pearson, V. & Gianinazzi, S. (eds). Physiological and Genetical Aspects of Mycorrhizae. INRA, Paris. [ Links ]
Van Overbeek, L.S. & Saikkonen, K., 2016. Impact of bacterial-fungal interactions on the colonization of the endosphere. Trends Plant Sci. 21, 230-242. [ Links ]
Wang, G.M., Stribley, D.P., Tinker, P.B. & Walker, C., 1993. Effects of pH on arbuscular mycorrhiza I. Field observations on the long-term liming experiments at Rothamsted and Woburn. New Phytol. 124, 465-472. [ Links ]
Wilk, K., 2011. Polski rynek win w swietle zmian w krajowych i wspólnotowych uregulowaniach prawnych. Studia i Prace Wydzialu Nauk Ekonomicznych i Zarzadzania. 22, 135-148. [ Links ]
Wu, Q.S. & Xia, R.X., 2006. Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well-watered and water stress conditions. J. Plant. Physiol. 163, 417-425. [ Links ]
Wu, Q.S. & Zou, Y.N., 2010. Beneficial roles of arbuscular mycorrhizal in citrus seedlings at temperature stress. Scientia Hort. 125, 289-293. [ Links ]
Yu, B., Zhao, C.Y., Li, J., Li, J.Y. & Peng, G., 2015. Morphological, physiological, and biochemical responses of Populus euphratica to soil flooding. Photosynth. 53, 110-117. [ Links ]
Zhu, X.C., Song, F.B., Liu, S.Q., Liu, T.D. & Zhou, X., 2012. Arbuscular mycorrhizae improves photosynthesis and water status of Zea mays L. under drought stress. Plant Soil Environ. 58, 186-191. [ Links ]
Submitted for publication: July 2018
Accepted for publication: November 2018
* Corresponding author: E-mail address: grzegorz.mikiciuk@zut.edu.pl


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}