










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.38 n.2 Stellenbosch 2017
http://dx.doi.org/10.21548/38-2-1322
ORIGINAL RESEARCH ARTICLES
Characterisation of thiol-releasing and lower volatile acidity-forming intra-genus hybrid yeast strains for sauvignon blanc wine
R.S. HartI, II, *; B.K. NdimbaI; N.P. JollyI
IAgricultural Research Council Infruitec-Nietvoorbij, Private Bag X5026, Stellenbosch 7599, South Africa
IINational Agricultural Proteomics Research & Services Unit (NAPRSU), Department of Biotechnology, University of the Western Cape, Private Bag X17, Robert Sobukwe Road, Bellville, 7535, South Africa
ABSTRACT
A single Saccharomyces cerevisiae wine yeast strain produces a range of aroma and flavour metabolites (e.g. volatile thiols), as well as unfavourable metabolites (e.g. volatile acidity [VA]), during the alcoholic fermentation of white wine, especially Sauvignon blanc. The former contribute to the organoleptic quality of the final wine. Previous research showed that yeast-derived enzymes (proteins) are involved in the release of wine quality-enhancing or quality-reducing metabolites during fermentation. Small-scale winemaking trials were initiated to evaluate the protein expression and metabolite release of S. cerevisiae hybrid yeasts producing tropical fruit aroma. Commercial 'thiol-releasing' wine yeasts (TRWY) were included in winemaking trials as references. Improved hybrids were identified that showed enhanced thiol-releasing abilities, specifically 3-mercaptohexanol (3MH), and lower VA formation during the production of Sauvignon blanc wines compared to some commercial TRWY references. It is noteworthy that the hybrid NH 56 produced wines with the second highest 3MH levels after hybrid NH 84, and with the lowest acetic acid of all strains included in this study. This yeast was also the only strain to have downregulated proteins linked to amino acid biosynthesis, the pentose phosphate pathway, glycolysis, and fructose and galactose metabolism during the lag phase. Furthermore, differences in protein expression were reflected in the variation in metabolite release by different strains, thereby confirming that enzymes (proteins) are the final effectors of metabolite release.
Key words: Acetic acid, iTRAQ, metabolomic, Orbitrap LC-MS/MS, proteomics, SPE GC-MS/MS, volatile thiols
INTRODUCTION
During alcoholic fermentation of grape must, the metabolic activity of Saccharomyces cerevisiae leads to a range of compounds (metabolites) in wine (Pinu et al., 2015). These metabolites include, amongst others, monosaccharide sugars, organic acids, fatty acids, amino acids, esters and higher alcohols (Chambers et al., 2009). In addition, wine yeasts also mediate the release of metabolites that enhance tropical fruit aroma (e.g. volatile thiols) from their grape-derived bound aroma-inactive precursors (King, 2010; Bovo et al., 2015). Some wine yeast strains are implicated in the overproduction of undesirable metabolites (e.g. acetic acid). Acetic acid accounts for 90% of volatile acidity (VA) and results in vinegar-like off-flavours in wine (Du Toit & Pretorius, 2000; Swiegers et al., 2005; Ugliano et al., 2009). All these yeast-derived and/or yeast-mediated metabolites contribute to the organoleptic character of the wine, and it has been suggested that wine is the 'metabolic footprint' of the wine yeast strain used to carry out the fermentation (Howell et al., 2006; Mapelli et al., 2008). Excessive VA levels in wines will have a negative commercial impact, as these wines will not be marketable. Financial loss can also be incurred by commercial yeast manufacturers if a commercial wine yeast strain within their portfolio is implicated in excessive VA formation.
Strains of S. cerevisiae vary in their ability to produce and/or release volatile thiols, an important aroma compound in Sauvignon blanc, as well as the unfavourable metabolite, acetic acid (Swiegers et al., 2006a, 2007a; Holt et al., 2011). Therefore, the preferred wine yeast strain for Sauvignon blanc should produce varietal aromatic compounds without the production of undesirable off-flavours (Vilela-Moura et al., 2011; Coetzee & Du Toit, 2012).
Wine yeast strains suitable for the production of aromatic wine with enhanced tropical fruit aroma were identified as a wine industry priority (Swiegers et al, 2006b, 2007b; King, 2010; King et al, 2011). Previous laboratory-scale vinification trails with intra-genus hybrids showed differences in wine chemical and sensory properties (Hart et al., 2016). It has also been reported that yeast-derived enzymes (proteins) are involved in the release of wine quality-enhancing or reducing metabolites during fermentation (Holt et al, 2011; Roncoroni et al., 2011; Juega et al., 2012; Moreno-García et al., 2015). However, protein expression and metabolite production and release of the above-mentioned intra-genus hybrids during the fermentation of Sauvignon blanc must have never been investigated. This necessitated an in-depth study of wine yeast protein expression and metabolite release and the effect thereof on the organoleptic quality of wine, especially if the South African wine industry is to remain globally competitive. The Sauvignon blanc cultivar is ideally suited to this type of investigation, as the grapes contain bound aroma-inactive metabolites that can only be released by wine yeast during fermentation (Swiegers et al., 2006a; Holt et al., 2011). Therefore, this study was undertaken to evaluate a selection of yeasts for the improvement of Sauvignon blanc wine organoleptic quality with regard to tropical fruit aroma and low VA formation. A second objective was to investigate the expression of wine yeast protein and whether regulated proteins correlate with metabolites released and/or produced during fermentation. Isobaric tags for relative and absolute quantitation (iTRAQ) was used in conjunction with liquid chromatography-tandem mass spectrometry (LC-MS/ MS) to classify downregulated and overexpressed proteins originating from three commercial 'thiol-releasing' wine yeasts (TRWY) and five hybrid yeast strains.
MATERIALS AND METHODS
Yeast strains
A hybrid that was provisionally characterised as having the ability to produce wines with tropical fruit aroma will henceforth be referred to as TFPH. Subsequently, nine TFPH, viz. NH 48, NH 56, NH 57, NH 84, NH 88, NH 118, NH 140, NH 143 and NH 145 were used in this study. Likewise, a hybrid that was provisionally characterised as a low VA producer will henceforth be referred to as LVPH. Subsequently, five LVPH, viz. NH 18, NH 29, NH 34, NH 40 and NH 97 were used in this study. These hybrids are conserved in the ARC Infruitec-Nietvoorbij micro-organism culture collection (ARC Inf-Nvbij CC). Top TRWY strains, viz. VIN 7 and VIN 13 (Anchor yeast, Cape Town, South Africa), Zymaflore VL3, Zymaflore X5 (Laffort Oenologie, Bordeaux, France) and Fermicru 4F9 (DSM Oenology, Delft, The Netherlands) were included as references. The commercial hybrid strain N 96 (Anchor yeast, Cape Town, South Africa), and an additional experimental hybrid P 35, were also included in this study, as they have the ability to produce wine with tropical fruit aromas.
Matrix-assisted laser desorption/ionisation (MALDI) bio-typing
All yeast strains were characterised by matrix-assisted laser desorption/ionisation (MALDI) biotyping using a Bruker UltrafleXtreme MALDI-TOF/TOF MS (Bruker Daltonics, Bremen, Germany) as described by Hart et al. (2016).
Small-scale winemaking trials
Sauvignon blanc grapes were routinely sampled and submitted to the cellar laboratory for sugar (°B) and titratable acidity (TA) analyses. The grapes were harvested from vines grown on the ARC Nietvoorbij Research farm once the °B/TA ratio was ± 2.5. Subsequently, small-scale wines were made in triplicate according to the standard cellar method included in the ARC Infruitec-Nietvoorbij harvest programme 2014 (ARC Infruitec-Nietvoorbij experimental wine evaluation committee). Briefly, the harvested grapes were chilled to 15°C prior to mechanical destemming. Destemmed grape berries were crushed, followed directly by pressing the grape must and skins at 1 Bar. Free-run grape must (juice) was transferred to a sedimentation tank, after which Novozymes (0.5 g/hL) and SO2 (50 mg/L) were added to aid sedimentation and prevent oxidation respectively. The skins were discarded. A must sample was subjected to pH, TA, sugar and SO2 analyses. The total SO2 of the must was then adjusted to 50 mg/L and allowed to clarify overnight at 14°C. Subsequently, the clarified must was racked off the sediments by siphoning into a new fermentation container. Nine litres of Sauvignon blanc grape must (total sugar 21.9°B; TA 9.3 g/L; pH 3.28) were dispensed into stainless steel canisters and inoculated with the respective wine yeast starter cultures as described by Hart et al. (2016). The method was adjusted by having the respective yeast inoculums cultured for 24 hours in 800 mL yeast extract, peptone and dextrose (YPD) broth (Biolab, Merck, Johannesburg, South Africa). Subsequently, 180 mL of the 24-h cultures was used to inoculate clarified Sauvignon blanc grape must (2% [v/v] inoculum), after which 50 g/hL diammonium phosphate (DAP) was added. Ninety mL of bentonite solution (7.5 %) was added to the fermenting must after 48 h following inoculation. Samples of the fermenting must were taken every 48 h using food-grade CO2 to analyse residual glucose/fructose, ethanol, VA, total acidity and pH using an OenofossTM Fourier transform infrared (FTIR) spectrometer (FOSS Analytical A/S, Denmark) until the fermentations went to dryness (R/S < 5 g/L). The OenofossTM FTIR was calibrated using wine chemical parameters determined by wet chemistry and external accredited laboratories. The SO2 was analysed upon completion of the respective fermentations. The wines were racked off the yeast lees and the free SO2 was adjusted to 35 mg/L, followed by cold stabilisation at 0°C for at least two weeks.
Gas chromatography (GC) analysis
Wine aroma compounds (e.g. esters, total acids and higher alcohols [fusel oils]) were quantified by means of calibration mixtures in conjunction with gas chromatography (GC) as described by Louw (2007). The GC system used in this study comprised a HP 5890 Series II GC equipped with an HP 7673 Injector and HP 3396A Integrator (Hewlett Packard, Vienna, Austria). Aroma compounds were separated using a 60 m x 0.32 mm x 0.5 μηι Polyethylene Glycol column (Lab Alliance, State College, PA, USA).
Solid-phase extraction (SPE) and GC-MS/MS analysis
Solid-phase extraction was used to pre-concentrate key wine volatile thiols in the wines, viz. 3-mercaptohexanol (3MH) and 3-mercaptohexylacetate (3MHA), as described by Mateo-Vivaracho et al. (2009). Subsequently, GC coupled with tandem mass spectrometry (GC-MS/MS) was used to quantify thiol levels, as described by Mattivi et al. (2012). However, the GC-MS/MS system used in this study consisted of a GC Trace 1300/TSQ8000 mass selective detector equipped with an AI 1310 auto sampler (Thermo Scientific™ Inc, USA). Aroma compounds were separated using a 30 m x 0.25 mm x 0.25 μηι Zebron WAX plus column (Phenomenex Inc., Torrance, CA, USA). Volatile thiol analysis was conducted by an accredited laboratory.
Sensory evaluation
Descriptive sensory evaluation of the bottled wines was conducted by an experienced panel consisting of14 members who were requested to indicate the prominence of listed aroma descriptors on a unipolar six-point numerical scale (absent [0], very low [1], low [2], medium [3], high [4] and very high [5]). In addition, the panel members had to specify the listed aroma descriptors associated with Sauvignon blanc that they perceived. All wines were coded and served in a randomised order using international wine-tasting glasses (approximately 50 mL).
Quantitative LC-based iTRAQ proteomic analysis
Protein sample preparation
Eight yeast strains (VIN 7, Zymaflore X5, Zymaflore VL3, N 96, P 35, NH 56, NH 57 and NH 97) were subjected to protein extraction based on chemical (lower VA and total fatty acids) and sensory (tropical fruit aroma) analyses. Samples were taken during the lag phase (48 hours following inoculation) and stationary phase (end of fermentation). Protein quantification and characterisation were conducted by deploying an iTRAQ 8-plex reagent kit (AB Sciex, USA) in conjunction with LC-MS/MS at the Mass Spectroscopy Unit in the Proteomics Laboratory, at the Central Analytical Facility (CAF) of Stellenbosch University (SU) (protocol can be obtained from the CAF SU).
Protein alkylation, digestion and isobaric labelling
Cysteine residues of the proteins were alkylated using 200 mM methane methylthiosulphonate (MMTS), as described by Boutureira and Bernardes (2015). The proteins were then digested overnight at 37°C using 1 μg/μL trypsin solution (Promega, Madison WI, USA). Subsequently, tryptic digests were air-dried and dissolved in water containing 2% acetonitrile/0.1% formic acid (Sigma-Aldrich, South Africa). The solution was again air-dried, although it was now re-suspended in triethylammonium bicarbonate (TEAB). Tryptic digests extracted from the yeast strains were tagged with iTRAQ labels 113, 114, 115, 116, 117, 118, 119 and 121 respectively, as described by Kim et al. (2012).
Liquid chromatography-tandem mass spectrometry (LC-MS/MS)
Mass spectrometry for protein characterisation was performed using a mass spectrometer equipped with a Nanospray flex ionisation source (Thermo Scientific™ Inc, USA). Samples were introduced through a stainless steel emitter as described by Vehus et al. (2016). Data was generated using synchronised pre-cursor selection with MS3 reporter ion generation. Subsequently, the raw files were imported into Proteome Discoverer v. 1.4 and processed using the Mascot algorithm (Matrix Science, London, UK), as well as the SequestHT algorithm included in Proteome Discoverer. Protein quantitation was performed using the iTRAQ quantitation algorithm. Only proteins with more than two peptides, but less than 20% variation, and iTRAQ ratios below 0.5 and above 2 were considered downregulated and overexpressed respectively.
Bioinformatics analysis
Proteins that were shown to be differentially expressed using quantitative iTRAQ were subjected to Protein ANalysis THrough Evolutionary Relationships (PANTHER, www. pantherdb.org/) analysis to establish their involvement in biological processes, molecular function and protein classes (Sharma et al., 2014).
Statistical analyses
Data generated following the chemical, sensory and metabolomic analyses was subjected to analysis of variance (ANOVA) and principal component analysis (PCA), as described (Pearson, 1896, 1901; Zou et al., 2006). A Pearson's correlation was performed to study the linear relationship between the chemical, sensory and metabolomic variables and to standardise the data prior to performing the PCA using XLSTAT software (Addinsoft, 2013), with the principal components (PCs) as factors (i.e. F1 and F2).
RESULTS AND DISCUSSION
Matrix-assisted laser desorption/ionisation (MALDI) biotyping
All 21 yeast strains were shown to have distinctive mass spectra and to belong to the species S. cerevisiae (Hart et al., 2016).
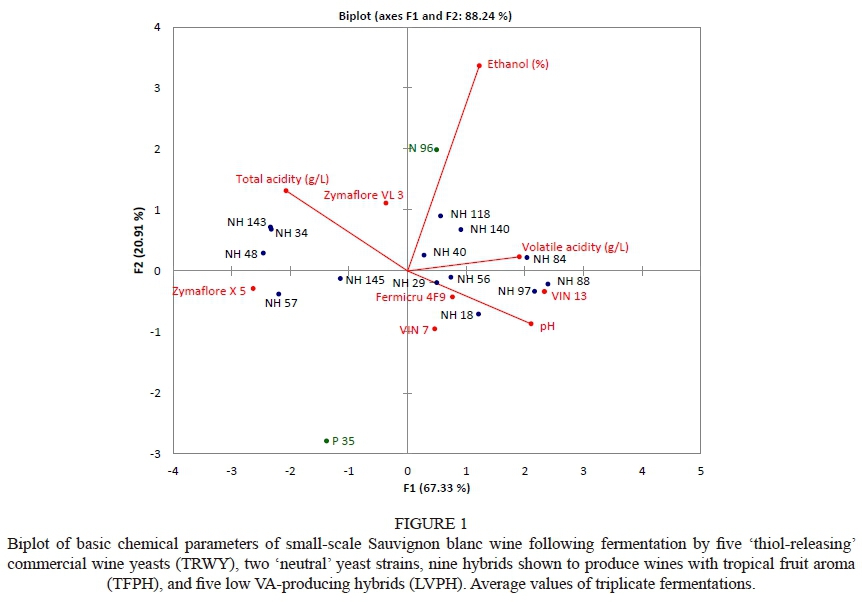
Fourier transform infra-red (FTIR) spectroscopy A principle component analysis (PCA) biplot of standard wine chemical data generated by FTIR spectroscopy showed that the LVPH NH 18, NH 29, NH 34 and NH 97 and the TFPH NH 48, NH 56, NH 57, NH 88, NH 143 and NH 145 were positioned in the left and bottom quadrants, and the wines had a negative association with VA (Fig. 1). The indications therefore are that these TFPH are also LVPH. The same observation was made with regard to commercial TRWY references, despite some producing wines with a more positive association with VA. However, Hart et al. (2016) reported that aforesaid yeast strains produced wines with VA levels that comply with legislation. None of the wines were perceived to be undesirable during descriptive sensory evaluation (Fig. 2).
Sensory evaluation
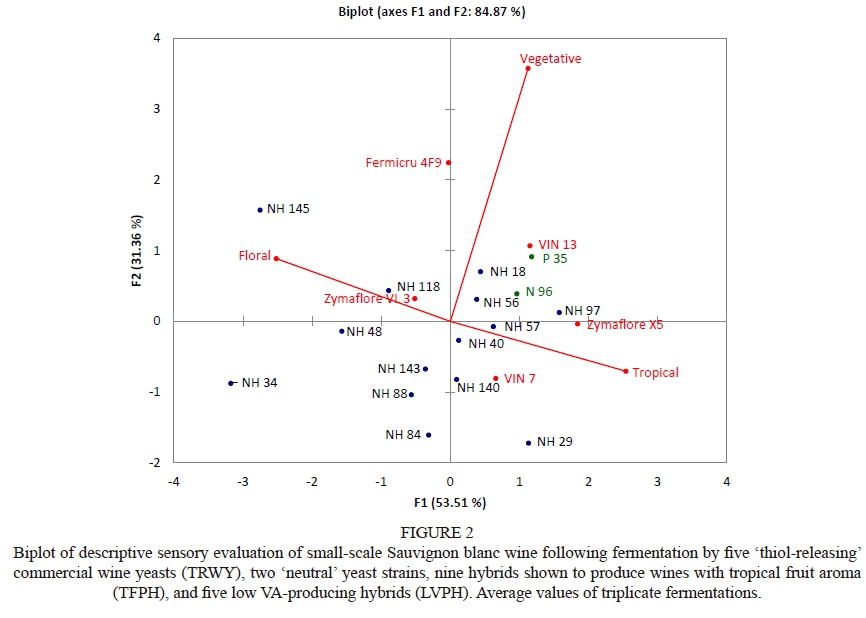
The PCA biplot of descriptive sensory evaluation showed that two commercial TRWY references, Zymaflore X5 and VIN 7, produced wines with a positive association with tropical fruit aromas, whilst Zymaflore VL3 and Fermicru 4F9 produced wines positively associated with floral and vegetative aromas (Fig. 2). The TRWY VIN 13 produced wines positively associated with both tropical fruit and vegetative aromas. Marais (1994) and Lapalus (2016) reported that green pepper and/or vegetative aromas can be attributed to Sauvignon blanc grape-derived aroma compounds (e.g. 2-isobutyl-3-methoxypyrazine [IBMP]). In this trial, these compounds could have masked the sought-after tropical fruit aroma of the wine produced with VIN 13 . Nonetheless, the production of wines with tropical fruit aromas is indicative of volatile thiol release and/or production, which supports yeast manufacturers' recommendations that the abovementioned TRWY strains are volatile "thiol-releasers" (Swiegers et al., 2009). This was further confirmed by Swiegers et al. (2006a) and Srisamatthakarn (2011), who reported that commercial TRWY references included in this study do in fact have enhanced thiol-releasing abilities.
Reference hybrid strains, namely N 96 and P 35, also produced wines with a positive association with tropical fruit and vegetative aromas respectively, as previously reported by Hart et al. (2016). Two TFPH, namely NH 57 and NH 140, produced wines that were perceived to have tropical fruit aromas, whilst NH 56 produced wines with almost equally intense tropical fruit and vegetative aromas. In contrast, three TFPH, namely NH 48, NH 118 and NH 145, produced wine with a positive association with floral aromas. The remainder of the TFPH, namely NH 84, NH 88 and NH 143, produced wines with a prominent tropical aroma with hints of floral aromas. Floral aroma can also be a characteristic of new-world Sauvignon blanc wines (Von Mollendorf, 2013). These hybrids produced wines that also had a negative association with vegetative aromas.
Sensorially, most TFPH included in this trial produced wine with equivalent and more pronounced tropical fruit and floral aromas compared to wines produced by TRWY references. The LVPH, namely NH 29 and NH 40, produced wines that had a positive association with tropical fruit aromas, whereas wines produced by NH 18 and NH 97 had a positive association with vegetative and tropical fruit aromas. On the other hand, the LVPH, namely NH 34, produced wines with floral aroma and tropical fruit aromas. Indications therefore are that some LVPH are also TFPH. Overall, most TFPH produced wines with more tropical fruit than vegetative aromas.
Gas chromatography (GC) analysis
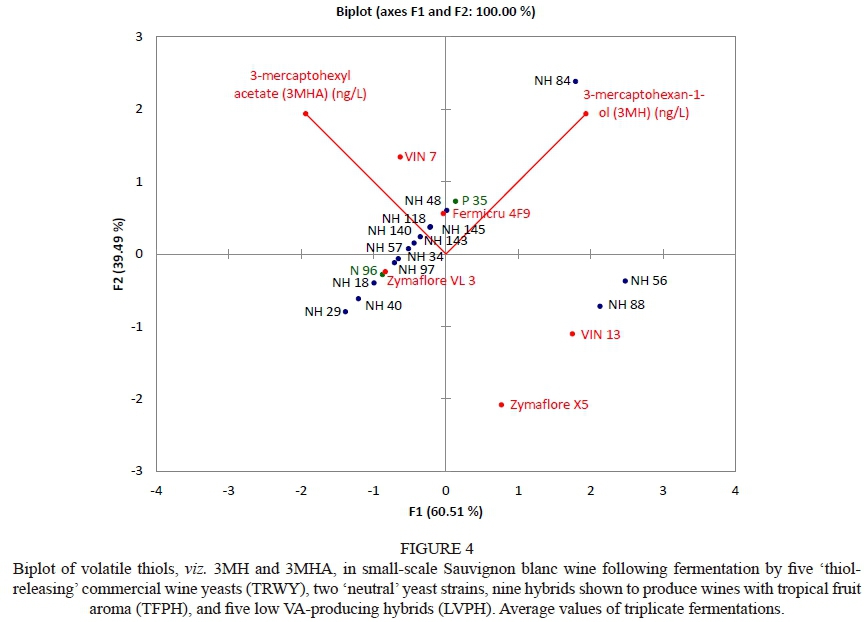
The ester, total fatty acid and higher alcohol levels of all small-scale wines were analysed, as these aroma compounds (metabolites) contribute positively to 'fermentation bouquet' when present in moderate concentration (Lambrechts & Pretorius, 2000; Coetzee & Du Toit, 2015). The PCA biplot showed that the TRWY reference, VIN 7, produced wines with the most positive association with esters compared to any other yeast strain included in this study (Fig. 3). The TRWY references, Zymaflore X5, Fermicru 4F9 and VIN 13, and the commercial hybrid yeast N 96, positioned in the upper quadrants, also produced wines with a positive association with esters. This observation complements the sensory evaluation results, where aforesaid yeast strains were shown to have a positive association with fruity aroma (Fig. 3). Wine esters are the main contributors to wine fruity aromas (Coetzee & Du Toit, 2015). In contrast, the TRWY reference, Zymaflore VL3, and the hybrid, P 35, produced wines with a negative association with esters, although P 35 produced wine that had a positive association with fruity aromas following sensory evaluation.
Hybrids NH 34 and NH 48 produced wines with a positive association with volatile fatty acids (e.g. octanoic acid, decanoic acid), which can be associated with faint fruity and citrus wine aroma (Lambrechts & Pretorius, 2000). However, wine fatty acids include acetic acid, the main acid responsible for vinegar-like off-flavours at higher concentrations (Vilela-Moura et al., 2011). Furthermore, Zoecklein et al. (1995) report that other steam-distillable carboxylic acids (e.g. lactic acid, formic acid, butyric acid and propionic acid) also contribute to the total acid content of wine. Volschenk et al. (2006) and Jackson (2014) report that, besides distillable acids, non-volatile acids (e.g. malic and tartaric acid) also contribute to TA, which was traditionally measured as an indicator of VA. Hybrids NH 34 and NH 48 are not necessarily high VA producers, since they were reported to be LVPH (Hart et al., 2016). The TRWY references, namely Zymaflore VL3, Fermicru 4F9, VIN 13 and hybrid P 35, produced wines with a negative association with total fatty acids. The same observation was made with regard to five hybrids, namely NH 29, NH 57, NH 84, NH 88, and NH 140, that also produced wines with prominent tropical fruit aromas (Fig. 2). Therefore, these strains comply with yeast selection criteria based on the ability to produce wine with tropical fruit aroma and low VA formation.
Overall, the level of higher alcohols in all wines were 242 ± 37.18 mg/L. Five hybrids, namely NH 34, NH 40, NH 48, NH 143 and NH 145, were shown to produce wines with a positive association with higher alcohols. Higher alcohol concentrations below 300 mg/L add complexity to wines (Munoz et al., 2006; Styger, 2011; Von Mollendorf, 2013; Moss, 2015). The complexity of the abovementioned wines is evident, as NH 48 and NH 145 produced wines with a positive association with floral aromas (Fig. 2), whilst NH 34, NH 40 and NH 143 produced wines associated with tropical fruit and floral aromas (Fig. 2). Lambrechts and Pretorius (2000) report that higher alcohols are key precursors involved in ester formation during alcoholic fermentation, which complements this study, as these hybrids produce wines with a positive association with tropical fruit aroma (Fig. 2) and esters (Fig. 3).
Solid-phase extraction (SPE) and GC-MS/MS analysis
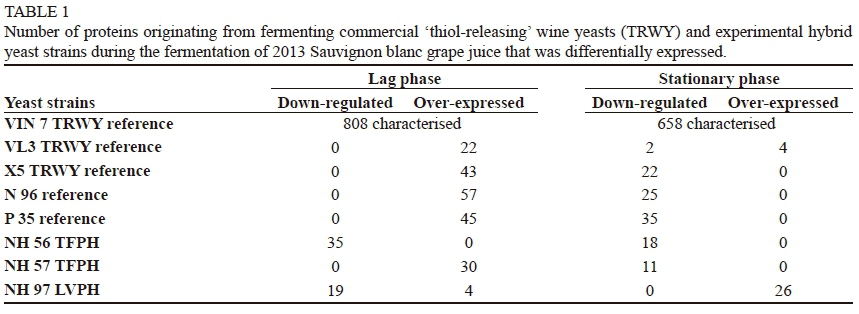
Volatile thiols contribute positively to the 'varietal aroma' of Sauvignon blanc wines (Fedrizzi et al., 2007; Pinu et al., 2012). Two thiols in particular, viz. 3-mercaptohexan-1-ol (3MH) and 3-mercaptohexyl acetate (3MHA), are the key aroma-enhancing metabolites in white wines, especially Sauvignon blanc, and are produced by wine yeasts during fermentation from odourless precursors present in grape juice (Swiegers et al., 2007b; Roland et al, 2011; Harsch et al., 2013). The SPE-GC-MS/MS data of the analysed wines is presented as a PCA biplot in Fig. 4. The TRWY references, namely VIN 7 and Fermicru 4F9, and hybrid P 35 produced wines with a positive association with both thiols. The TRWY, namely VIN 13 and Zymaflore VL3, produced wines positively associated with 3MH, whilst the TRWY Zymaflore X5 and the commercial hybrid N 96 produced wines positively associated with 3MHA (Fig. 4). Overall, the 3MH levels in these wines were 270, 258, 280, 266, 155, 140 and 150 ng/L. It is noteworthy that the 3MH levels in the VIN 7 produced wine was at least five times its sensory detection threshold (Van Wyngaard, 2013), explaining the positive association with tropical fruit aroma (Fig. 2). The TRWY references, namely Zymaflore X5 and Zymaflore VL3, produced wines with the lowest 3MH and 3MHA of all TRWY references. This is in contrast to results from Dubourdieu (2006) and Bowyer et al. (2008), who reported both yeasts to be high thiol releasers. Nonetheless, the 3MH levels in both wines were twice their sensory detection thresholds, and these wines had a positive association with tropical fruit and floral aromas (Fig. 2).
Seven hybrids (NH 48, NH 56, NH 57, NH 84, NH 118, NH 140 and NH 143) produced wines with varying 3MH levels (264, 360, 196, 493, 315, 234, 217 and 206 ng/L respectively). These hybrids also produced wines with the strongest association with 3MH and 3MHA compared to wines produced with Zymaflore X5, VIN 13 and Zymaflore VL3 (Fig. 3). The 3MH levels in the TFPH NH 84 produced wine was at least eight times its sensory detection threshold (Van Wyngaard, 2013), and twice that observed for the commercial TRWY reference VIN 7. Some of these hybrids were shown to produce wines with a more positive association with tropical fruit aromas (Fig. 2) and lower total fatty acids (Fig. 3) than wines produced with the commercial references, namely Zymaflore VL3 and Fermicru 4F9. Hybrids NH 56, NH 84 and NH 88, which produced wines with a positive association with esters (Fig. 3), also produced wines with the highest 3MH levels of all yeast strains included in this study. The remaining TFPH also produced wines with a stronger association with 3MH and 3MHA than some of the commercial references. The hybrid yeast P 35, which was also utilised as a parental yeast in the hybrid breeding programme, produced wines with higher 3MH levels than any of the commercial TRWY references. Therefore, TFPH that produced wines with a strong association with aforesaid thiols could have inherited this trait from P 35.
Quantitative LC-based iTRAQ proteomic analysis
Yeast-derived enzymes (proteins) are involved in the release of wine quality-enhancing or reducing metabolites during fermentation (Juega et al., 2012; Moreno-García et al., 2015). In this study, the selected TRWY references, VIN 7, Zymaflore VL3 and Zymaflore X5, the commercial yeast N 96, the experimental hybrid yeast P 35, promising TFPH, viz. NH 56 and NH 57, and one LVPH, NH 97, were selected for quantitative LC-based iTRAQ analysis to investigate protein expression during the lag phase (48 hours following inoculation) and stationary phase (end of fermentation). The TRWY VIN 7 was chosen as reference for iTRAQ quantification, as it is a known 'thiol-releaser' and producer of wines with tropical aroma (Swiegers et al., 2006b; Borneman et al., 2012; Howe, 2016).
In order to minimise false positives during relative iTRAQ quantification and the identification of proteins, cut-off ratios of 0.5 and 2.0 with less than 20% variation were applied for downregulated and overexpressed proteins respectively. The analysis of the combined datasets in conjunction with the Uniprot S. cerevisiae database (2015 edition) identified a total of 808 and 658 S. cerevisiae-derived proteins during the lag phase and stationary phase respectively (Table 1). All yeast strains showed variation in the number of up-and downregulated proteins in comparison to the VIN 7 reference-expressed proteins. Overall, 54 proteins (6.68%) were downregulated and 201 proteins (24.88%) were overexpressed during the lag phase of fermentation, whilst 113 proteins (17.17%) were downregulated and 28 proteins (4.25%) were overexpressed during the stationary phase of fermentation.
Non-Saccharomyces-expressed proteins originating from Schizosaccharomyces pombe and Candida tropicalis were observed during the lag phase, whilst none were detected at the end of fermentation. This observation complements previous research, since non-Saccharomyces genera are prominent in grape must but are vanquished by Saccharomyces spp. as the fermentation progresses (Varela et al., 2012; Jolly et al., 2014; Wang et al., 2016). Only three and 20 of the non-Saccharomyces derived-proteins were downregulated and overexpressed respectively. As the base must (juice) was homogenous and standard for all strains evaluated, the different reference and hybrid strain inoculums are the only variable (treatment).
Proteomic analyses showed that all yeast strains, except NH 56, overexpressed between four and 57 proteins during the lag phase, whilst NH 56 and NH 97 were the only strains to have downregulated 35 and 19 proteins respectively. The overexpression of proteins linked to the rate and process of glycolysis, nitrogen and carbohydrate metabolism during the lag phase of fermentation is to be expected, because the yeast is actively generating energy whilst adapting to the new environment prior to proliferation (Zuzuarregui et al., 2006; Salvado et al., 2008). The proteins downregulated by yeast NH 56 were linked to amino acid biosynthesis, the pentose phosphate pathway, glycolysis and the fructose and galactose metabolism pathways (e.g. dehydrogenases, reductases, synthetases, hydrolases, proteases, signalling molecules, dehydratases and transfer/carrier proteins). It therefore can be speculated that the yeast strain reduced its metabolic rate compared to the VIN 7 TRWY reference, as well as to other strains included in this trial. Anecdotal evidence suggests that this delay might be instrumental in volatile thiol release, since wines produced by NH 56 had the highest positive association with 3MH (Fig. 3). It is noteworthy that NH 56 did not regulate /?-lyases (carbon-sulphur lyases), enzymes previously reported to be responsible for the release of volatile thiols (Howell et al., 2005; Swiegers et al., 2007a).
Proteomic analyses showed that all strains downregulated proteins during the stationary phase, except for Zymaflore VL3 and NH 97, which overexpressed four and 28 proteins respectively. It was also observed that both strains produced wines with significantly lower 3MH compared to wines produced by NH 56. The remaining strains that did not overexpress any proteins during the stationary phase produced wines with similar 3MH levels. Indications therefore are that the downregulation of proteins linked to, amongst others, amino acid biosynthesis, the pentose phosphate pathway, glycolysis and fructose and galactose metabolism during the lag phase will enhance volatile thiol release. Furthermore, Holt et al. (2011) reported that amino acids and their linked metabolic pathways are directly linked to the release and production of wine aroma compounds (e.g. thiols).
Hybrid yeast NH 56 produced wines with the least association with total acids compared to wines produced with the remaining yeast strains. Vilela-Moura et al. (2011) and Vilela et al. (2013) reported that acetic acid, the main VA responsible for vinegar-like off-flavours, is a major contributor to total fatty acids. Therefore, the downregulation of proteins associated with, amongst others, the pentose phosphate pathway, glycolysis and fructose and galactose metabolism, and the lack of overexpression of any other proteins at the start of fermentation were instrumental in the lower VA levels observed in wines produced with NH 56. Varela et al. (2012) and Walkey et al., (2012) reported that dehydrogenase enzymes are involved in excessive acetic acid production. Yeast strains that produced wines with a stronger association with acetic acid than NH 56 overexpressed dehydrogenases at the start of fermentation. In contrast, hybrid yeast NH 56 downregulated dehydrogenase at the start of fermentation. It is evident that different yeast strains express proteins differentially in response to the same grape must matrix. Subsequently, the final wine organoleptic quality is affected by protein expression by the yeast starter culture.
Isobaric tags for relative and absolute quantification proved to be a practical approach to the identification of proteins extracted from eight different yeast strains simultaneously, as opposed to the laborious gel-based proteomics, which only makes provision for the visualisation of one sample at a time (May et al., 2012). Consequently, the molecular weights (MW) and isoelectric points (pI) of regulated proteins during the lag (88) and stationary (55) phases originating from NH 56, NH 57 and NH 97 were determined using two iTRAQ runs as opposed to six two-dimensional poly-acrylamide gel electrophoresis (2D-PAGE) runs. Blasco et al. (2011) reported that yeast-derived proteins present in must and/or wine are mainly glycoproteins. This study supports those observations, since regulated proteins include those associated with glycolysis. In general, regulated proteins during the different phases of fermentation were shown to be heterogeneous, since their MW and pI ranged from 7.10 to 351.40 kDa and pH 4.10 to 11.71 respectively.
Proteins differentially expressed by the yeast strains during the lag and stationary phases of Sauvignon blanc grape must fermentation were classified according to molecular function (Fig. 5a & 5b), biological process (Fig. 5c & 5d), and protein class (Fig. 5e & 5f) using PANTHER (Sharma et al., 2014). The classification of proteins showed that differentially expressed proteins during the lag phase related to seven molecular functions, viz. transporter, translation regulator, structural molecule, receptor, catalytic, binding and antioxidant activities (Fig. 5a), whilst differentially expressed proteins during the stationary phase are associated with the same molecular functions (Fig. 5b). Therefore, regulated proteins during the start and end of fermentation are associated with the same molecular functions.
The classification of proteins showed that differentially expressed proteins during the lag phase are associated with six biological processes, viz. response to stimulus, metabolic, localisation, cellular, biogenesis and biological regulation (Fig. 5c), whilst differentially expressed proteins during the stationary phase are associated with five biological processes, viz. metabolic, localisation, cellular, biogenesis and biological regulation (Fig. 5d). Therefore, no proteins associated with 'stimulated response' were regulated towards the end of fermentation. It can tentatively be speculated that aroma-inactive bound thiol precursors typically found in abundance in Sauvignon blanc grape must at the start of fermentation were depleted (Helwi et al., 2016; O'Kennedy, 2016), hence the fermenting yeast strains were not stimulated to express proteins associated with the release of volatile aromatic thiols. The classification of proteins showed that differentially expressed proteins during the lag phase clustered into sixteen protein classes (Fig. 5e), whilst differentially expressed proteins during the stationary phase were associated with twelve different protein classes (Fig. 5f).
It was observed that regulated proteins were mutual to eleven protein classes during the two phases of fermentation. However, five protein classes were observed exclusively during the lag phase, viz. transporter, transfer/carrier, transcription factors, signalling molecule and defence. Luy-ten et al. (2002), Henricsson et al. (2005) and Young et al. (2011) reported that transporter proteins that play a pivotal role during the transport of molecules (e.g. hexose sugars) from the grape must into the cell are overexpressed during the lag phase, as the cells are adapting to their environment prior to active proliferation. Perez et al. (2005) reported that carrier proteins responsible for the transport of glucose and fructose into cells were only expressed during the lag phase.
Rossignol et al. (2006) reported that many genes associated with transcription and/or protein synthesis were regulated during the initiation of fermentation. It can be concluded that the resultant protein expression will be higher. In accordance with Rossignol et al. (2006), the transcription factors (proteins) in this study were also regulated during the lag phase. The wine yeast S. cerevisiae is known to secrete signalling molecules (proteins) to communicate to surrounding haploid S. cerevisiae with the intention to mate whilst growing in a rich medium (Lodish et al., 2000; Merlini et al., 2013). Proteomic analyses of fermenting must during this study showed that signalling molecules were also regulated during the lag phase (Fig. 5e). Wine yeasts were also shown to undergo oxidative stress during fermentation, resulting in the upregulation of defence-related genes during the fermentation of a high sugar-containing medium (Gómez-Pastor et al, 2010; Navarro-Tapia et al., 2016). The Sauvignon blanc base must used in this study had an initial sugar content of 221 g/L. It therefore can be concluded that this physiological condition also induced oxidative stress in the yeast investigated, as the data showed that defence proteins were only expressed during the lag phase.
CONCLUSIONS
In conclusion, seven LVPH (NH 29, NH 56, NH 57, NH 84, NH 88, NH 118 and NH 140) were identified, of which all but strain NH 29 were also identified as TFPH. These strains therefore conform to the initial yeast selection criteria set forth in the aims of study, viz. enhanced thiol-releasing or tropical fruit aroma wine-producing abilities and lower VA formation. The TFPH were shown to produce wines with a more positive association with 3MH and 3MHA compared to wines produced with the commercial TRWY references. The LVPH and TFPH therefore have the potential to play a commercial role in the production of varietal aromatic white wine. The TFPH NH 56 produced wines with the second highest 3MH levels after NH 84, but with the lowest acetic acid of all strains included in this study. This yeast was also the only strain to have downregulated proteins during the lag phase; these were linked to amino acid biosynthesis, the pentose phosphate pathway, glycolysis and fructose and galactose metabolism. Differences in protein expression were reflected in the variation of metabolite release by the different strains, thereby confirming that proteins are the final effectors of metabolite release. This study highlights the importance of said protein classes, besides //-lyases, in the release of volatile thiols.
LITERATURE CITED
Addinsoft (2013). XLSTAT software, version 2013. Paris, France. [ Links ]
Blasco, L., Vinas, M. & Villa, T.G., 2011. Proteins influencing foam formation in wine and beer: The role of yeast. Intl. Microbiol. 14, 61-71. doi:10.2436/20.1501.01.136 [ Links ]
Borneman, A.R., Desany, B.A., Riches, D., Affourtit, J.P., Forgan, A.H., Pretorius, I.S., Egholm, M. & Chambers, P.J., 2012. The genome sequence of the wine yeast VIN 7 reveals an allotriploid hybrid genome with Saccharomyces cerevisiae and Saccharomyces kudriavzevii origins. FEMS Yeast Res. 12, 88-96. [ Links ]
Boutureira, O. & Bernardes, G.J.L., 2015. Advances in chemical protein modification Chem. Rev. 115, 2174-2195. [ Links ]
Bovo, B., Carlot, M., Fontana, F., Lombardi, A., Soligo, S., Giacomini, A. & Corich, V., 2015. Outlining a selection procedure for Saccharomyces cerevisiae isolated from grape marc to improve fermentation process and distillate quality. Food Microbiol. 46, 573-581. [ Links ]
Bowyer, P., Gourraud, C., Murat, M. & Van der Westhuizen, T., 2008. Modulation of Sauvignon blanc aromas through yeast strain, nutrition and seasonal variation [Online]: http://wineland.archive.shapeshift.co.za/archive/index.php?option =com_ zine&view=article&id=156:modulation-of-sauvignon-blanc-aromas-through-yeast-strain-nutrition-and-seasonal-variation [accessed on 1st of July 2016]. [ Links ]
Chambers, P.J., Borneman, A.R., Schmidt, S.A., Hack. J.C., Varela, C., Mercurio, M., Curtin, C.D., Cozzolino, D., Ugliano, M., Herderich, M.J. & Pretorius, I.S., 2009. The dawn of a new paradigm for wine yeast strain development. Aust. N. Z. Wine Indus. J. 24(3), 16-18. [ Links ]
Coetzee, C. & Du Toit, W.J., 2012. A comprehensive review on Sauvignon blanc aroma with a focus on certain positive volatile thiols. Food Res. Int. 45, 287-298. [ Links ]
Coetzee, C. & Du Toit, W.J., 2015. Sauvignon blanc wine: Contribution of ageing and oxygen on aromatic and non-aromatic compounds and sensory composition - A review. S. Afr. J. Enol. Vitic. 36, 347-365. [ Links ]
Dubourdieu, D., Tominaga, T., Masneuf, I., Des Gachons, C.P. & Murat, M.L., 2006. The role of yeasts in grape flavor development during fermentation: The example of Sauvignon blanc. Am. J. Enol. Vitic. 57, 81-88. [ Links ]
Du Toit, M. & Pretorius, I.S., 2000. Microbial spoilage and preservation of wine: Using weapons from nature's own arsenal - A review. S. Afr. J. Enol. Vitic. 21, 74-96. [ Links ]
Fedrizzi, B., Versini, G., Lavagnini, I., Nicolini, G. & Magno, F., 2007. Gas chromatography-mass spectrometry determination of 3-mercaptohexan-1-ol and 3-mercaptohexyl acetate in wine: A comparison of headspace solid phase microextraction and solid phase extraction methods. Anal. Chim. Acta. 596, 291-297. [ Links ]
Gómez-Pastor, R., Pérez-Torrado, R., Cabiscol, E., Ros, J. & Matallana, E., 2010. Reduction of oxidative cellular damage by overexpression of the thioredoxin TRX2 gene improves yield and quality of wine yeast dry active biomass. Microb. Cell Fact. 9, 9. doi:10.1186/1475-2859-9-9. [ Links ]
Hart, R.S., Jolly, N.P., Mohamed, G., Booyse, M. & Ndimba, B.K., 2016. Characterisation of Saccharomyces cerevisiae hybrid yeasts selected for low volatile acidity formation and the production of aromatic Sauvignon blanc wine. Afr. J. Biotech. 15, 2068-2081. [ Links ]
Harsch, M.J., Benkwitz, F., Frost, A., Colonna-Ceccaldi, B., Garner, R.C. & Salmon, J.M., 2013. New precursor of 3-mercaptohexan-1-ol in grape juice: Thiol-forming potential and kinetics during early stages of must fermentation. J. Agr. Food Chem. 61, 3703. [ Links ]
Helwi, P., Guillaumie, S., Thibon, C., Keime, C., Habran, A., Hilbert, G., Gomes, E., Darriet, P., Delrot, S. & Van Leeuwen, C., 2016. Vine nitrogen status and volatile thiols and their precursors from plot to transcriptome level. BMC Plant Biol. 16, 173. doi:10.1186/s12870-016-0836-y [ Links ]
Henricsson, C., De Jesus Ferreira, M.C., Hedfalk, K., Elbing, K., Larsson, C., Bill, R.M., Norbeck, J., Hohmann, S. & Gustafsson, L., 2005. Engineering of a novel Saccharomyces cerevisiae wine strain with a respiratory phenotype at high external glucose concentrations. Appl. Environ. Microbiol. 71, 6185-6192. [ Links ]
Holt, S., Cordente, A.G., Williams, S.J., Capone, D.L., Jitjaroen, W., Menz, I.R., Curtin, C. & Anderson, P.A., 2011. Engineering Saccharomyces cerevisiae to release 3-mercaptohexan-1-ol during fermentation through overexpression of an S. cerevisiae gene, STR3, for improvement of wine aroma. App. Environ. Microbiol. 77, 3626-3632. [ Links ]
Howe, G., 2016. The new wine speak of Sauvignon blanc [Online]: http://www.durbanvillewine.co.za/blog/the-new-wine-speak-of-sauvignon-blanc [accessed on 07th of Mar 2016]. [ Links ]
Howell, K.S., Cozzolino, D., Bartowsky, E., Fleet, G.H. & Henschke, P.A., 2006. Metabolic profiling as a tool for revealing Saccharomyces interactions during wine fermentation. FEMS Yeast Res. 6, 91-101. [ Links ]
Howell, K.S., Klein, M., Swiegers, J.H., Hayasaka, Y., Elsey, G.M., Fleet, G.H., H0j, P.B., Pretorius, I.S. & De Barros Lopes, M.A., 2005. Genetic determinants of volatile-thiol release by Saccharomyces cerevisiae during wine fermentation. Appl. Environ. Microbiol. 71, 5420-5426. doi:10.1128/AEM.71.9.5420-5426.2005 [ Links ]
Jackson, R.S., 2014 (4th ed). Wine science: Principles and applications (Food Science and Technology). Academic Press, an imprint of Elsevier, Amsterdam. [ Links ]
Jolly, N.P., Varela, C. & Pretorius, I.S., 2014. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 14, 215-237 doi:10.1111/1567-1364.12111 [ Links ]
Juega, M., Nunez, Y.P., Carrascosa, A.V. & Martinez-Rodriguez, A.J., 2012. Influence of yeast mannoproteins in the aroma improvement of white wines. J. Food Sci. 77, 499-504. [ Links ]
Kim, P.D., Patel, B.B. & Yeung, A.T., 2012. Isobaric labeling and data normalization without requiring protein quantitation. J. Biomol. Tech. 23, 11-23. [ Links ]
King, E.S., 2010. The modulation of Sauvignon blanc wine aroma through control of primary fermentation. PhD thesis, The School of Agriculture, Food and Wine, University of Adelaide, Adelaide SA, 5005, Australia. [ Links ]
King, E.S., Francis, I.L., Swiegers, J.H. & Curtin, C., 2011. Yeast strain-derived sensory differences retained in Sauvignon blanc wines after extended bottle storage Am. J. Enol. Vitic. 62, 366-370. [ Links ]
Lambrechts, M.G. & Pretorius, I.S., 2000. Yeast and its importance to wine aroma - A review. S. Afr. J. Enol. Vitic. 21, 97-129. [ Links ]
Lapalus, E., 2016. Linking sensory attributes to selected aroma compounds in South African Cabernet Sauvignon wines. MSc thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Lodish, H.B., Berk, A., Zipursky, S.L., Matsudaira, P., Baltimore, D. & Darnell, J.E., 2000 (4th ed). Molecular cell biology. W.H. Freeman, New York. [ Links ]
Louw, L., 2007. Chemical characterisation of South African young wines. MSc thesis, Stellenbosch University, Private Bag X1, 7602, Matieland (Stellenbosch), South Africa. [ Links ]
Luyten, K., Riou, C. & Blondin, B., 2002. The hexose transporters of Saccharomyces cerevisiae play different roles during enological fermentation. Yeast. 19, 713-726. [ Links ]
Mapelli, V., Olsson, L. & Nielsen, J., 2008. Metabolic footprinting in microbiology: Methods and applications in functional genomics and biotechnology. Trends Biotechnol. 26, 490-497. [ Links ]
Marais, J., 1994. Sauvignon blanc cultivar aroma - A review. S. Afr. J. Enol. Vitic. 15, 41-45. [ Links ]
Mateo-Vivaracho, L., Cacho, J. & Ferreira, V., 2009. Selective preconcentration of volatile mercaptans in small SPE cartridges: Quantitative determination of trace odor-active polyfunctional mercaptans in wine. J. Sep. Sci. 32, 3845-3853. [ Links ]
Mattivi, F., Fedrizzi, B., Zenatob, A., Tiefenthalerb, P., Tempestab, S., Perenzonia, D., Cantarella, P., Simeoni, F. & Vrhovseka, U., 2012. Development of reliable analytical tools for evaluating the influence of reductive winemaking on the quality of Lugana wines. Anal. Chim. Acta. 732, 194-202. [ Links ]
May, C., Brosseron, F., Chartowski, P., Meyer, H.E. & Marcus, K., 2012. Differential proteome analysis using 2D-DIGE. Methods Mol. Biol. 893, 75-82. [ Links ]
Merlini, L., Dudin, O. & Martin, S.G., 2013. Mate and fuse: How yeast cells do it. Open Biol. doi:10.1098/rsob.130008 [ Links ]
Moreno-García, J., García-Martínez, T., Millán, M.C., Mauricio, J.C. & Moreno, J., 2015. Proteins involved in wine aroma compounds metabolism by a Saccharomyces cerevisiae flor-velum yeast strain grown in two conditions. Food Microbiol. 51, 1-9. [ Links ]
Moss, R., 2015. How volatile fatty acids and sulphurous compounds impact on key aromas [Online]: http://www.wineland.co.za/technical/how-volatile-fatty-acids-and-sulphurous-compounds-impact-on-key-aromas [accessed on 27th of May 2016]. [ Links ]
Munoz, D., Peinado, R., Medina, M. & Moreno, J., 2006 Higher alcohols concentration and its relation with the biological aging evolution. Eur. Food. Res. Technol. 222, 629-635. [ Links ]
Navarro-Tapia, E., Nana, R.K., Querol, A. & Pérez-Torrado, R., 2016. Ethanol cellular defense induce unfolded protein response in yeast. Front. Microbiol. 7, 189. doi:10.3389/fmicb.2016.00189 [ Links ]
O'Kennedy, K., 2016. Increasing Sauvignon blanc aroma with H2S [Online]: http://igws.co.za/article/blog/increasing-sauvignon-blanc-aroma-with-h2s [accessed on 06th of Sep 2016]. [ Links ]
Pearson, K., 1896. Mathematical contributions to the theory of evolution. III. Regression, heredity and panmixia. Philos. Trans. Royal Soc. London Ser. A. 187, 253-318. [ Links ]
Pearson, K., 1901. On lines and planes of closest fit to systems of points in space. Phil. Mag. 2, 559-572. [ Links ]
Perez, M., Luyten, K., Michel, R., Riou, C. & Blondin, B., 2005. Analysis of Saccharomyces cerevisiae hexose carrier expression during wine fermentation: Both low- and high-affinity Hxt transporters are expressed. FEMS Yeast Res. 5, 351-361. [ Links ]
Pinu, F.R., Edwards, P.J.B., Gardner, R.C. & Villas-Boas, S.G., 2015. Nitrogen and carbon assimilation by Saccharomyces cerevisiae during Sauvignon blanc juice fermentation. FEMS Yeast Res. 14, 1206-1222. [ Links ]
Pinu, F.R., Jouanneau, S., Nicolau, L., Gardner, R.C. & Villas-Boas, S.G., 2012. Concentrations of the volatile thiol 3-mercaptohexanol in Sauvignon blanc wines: No correlation with juice precursors. Am. J. Enol. Vitic. 63, 407-412. [ Links ]
Roland, A., Schneider, R., Razungles, A. & Cavelier, F., 2011. Varietal thiols in wine: Discovery, analysis and applications. Chem. Rev. 22, 7355-7376. doi:10.1021/cr100205b [ Links ]
Roncoroni, M., Santiago, M., Hooks, D.O., Moroney, S., Harsch, M.J. & Lee, S.A., 2011. The yeast IRC7 gene encodes a β-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 28, 926-935. [ Links ]
Rossignol, T., Postaire, O., Storaï, J. & Blondin, B., 2006. Analysis of the genomic response of a wine yeast to rehydration and inoculation. Appl. Microbiol. Biotechnol. 71, 699-712. [ Links ]
Salvado, Z., Chiva, R., Rodríguez-Vargas, S., Rández-Gil, F., Mas, A. & Guillamo, J.M., 2008. Proteomic evolution of a wine yeast during the first hours of fermentation. FEMS Yeast Res 8, 1137-1146. [ Links ]
Sharma, S., Ray, S., Moiyadi, A., Sridhar, E. & Srivastava, S., 2014. Quantitative proteomic analysis of meningiomas for the identification of surrogate protein markers. Sci. Rep. 4, 71-40. doi:10.1038/srep07140 [ Links ]
Srisamatthakarn, P., 2011. Improvement of varietal aroma in grape and tropical fruit wines by optimal choice of yeasts and nutrient supplements. Dr. Agr. thesis, Justus-Liebig-University Giessen and Geisenheim Research Centre, Germany. [ Links ]
Styger, G., 2011. Elucidating the metabolic pathways responsible for higher alcohol production in Saccharomyces cerevisiae. PhD thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Swiegers, J., Bartowsky, E., Henschke, P. & Pretorius, I., 2005. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 11, 139-173. [ Links ]
Swiegers, J.H., Capone, D.L., Pardon, K.H., Elsey, G.M., Sefton, M.A., Francis, I.L. & Pretorius, I.S., 2007a. Engineering volatile thiol release in Saccharomyces cerevisiae for improved wine aroma. Yeast 24, 561-574. [ Links ]
Swiegers, J.H., Francis, I.L., Herderich, M.J. & Pretorius, I.S., 2006a. Meeting consumer expectations through management in vineyard and winery: The choice of yeast for fermentation offers great potential to adjust the aroma of Sauvignon blanc wine. Aust. N. Z. Wine Indus. J. 21, 34-42. [ Links ]
Swiegers, J.H., Kievit, R.L., Siebert, T., Lattey, K.A., Bramley, B.R., Francis, I.L., King, E.S. & Pretorius, I.S., 2009. The influence of yeast on the aroma of Sauvignon blanc wine. Food Microbiol. 26, 204-211. [ Links ]
Swiegers, J.H., King, E., Travis, B., Francis, L. & Pretorius, I.S., 2007b. Enhancement of Sauvignon blanc wine aroma through yeast combinations [Online]: http://www.wineland.co.za/technical/enhancement-of-sauvignon-blanc-wine-aroma-through-yeast-combinations [accessed on 27th of May 2016]. [ Links ]
Swiegers, J.H., Willmott, R., Hill-Ling, A., Capone, D.L., Pardon, K.H., Elsey, G.M., Howell, K.S., De Barros Lopes, M.A., Sefton, M.A., Lilly, M. & Pretorius, I.S., 2006b. Modulation of volatile thiol and ester aromas by modified wine yeast. Developments in Food Sci. 43, 113-116. [ Links ]
Ugliano, M., Kwiatkowski, M.J., Travis, B., Francis, I.L., Waters, E.J., Herderich, M.J. & Pretorius, I.S., 2009. Post-bottling management of oxygen to reduce off-flavour formation and optimise wine style [Online]: http://www.newworldwinemaker.com/pdf/AWRI_report_post-bottling_management.pdf [accessed on 23rd of Aug 2016]. [ Links ]
Van Wyngaard, E., 2013. Volatiles playing an important role in South African Sauvignon blanc wines. MSc thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Varela, C., Kutyna, D.R., Solomon, M.R., Black, C.A., Borneman, A., Henschke, P.A., Pretorius, I.S. & Chambers, P.J. 2012. Evaluation of gene modification strategies for the development of low-alcohol-wine yeasts. Appl. Environ. Microbiol. 78, 6068-6077. [ Links ]
Vehus, T., Seterdal, K.E., Krauss, S., Lundanes, E. & Wilson, S.R., 2016. Comparison of commercial nanoliquid chromatography columns for fast, targeted mass spectrometry-based proteomics. Future Sci. OA. doi:10.4155/ fsoa-2016-0014 [ Links ]
Vilela, A., Schuller, D., Mendes-Faia, A. & Cörte-Real, M., 2013. Reduction of volatile acidity of acidic wines by immobilized Saccharomyces cerevisiae cells. Appl. Microbiol. Biotechnol. 97, 4991-5000. [ Links ]
Vilela-Moura, A., Schuller, D., Mendes-Faia, A., Silva, R.D., Chaves, S.R., Sousa, M.J. & Cörte-Real, M., 2011. The impact of acetate metabolism on yeast fermentative performance and wine quality: Reduction of volatile acidity of grape musts and wines. Appl. Microbiol. Biotechnol. 89, 271-280. [ Links ]
Volschenk, H., Van Vuuren, H.J.J. & Viljoen-Bloom, M., 2006. Malic acid in wine: Origin, function and metabolism during vinification. S. Afr. J. Enol. Vitic. 27, 123-136. [ Links ]
Von Mollendorf, A., 2013. The impact of wine yeast strains on the aromatic profiles of Sauvignon blanc wines derived from characterized viticultural treatments. MSc thesis, Stellenbosch University, Private Bag X1, 7602 Matieland (Stellenbosch), South Africa. [ Links ]
Walkey, C.J., Luo. Z., Madilao, L.L. & Van Vuuren, H.J.J., 2012. The fermentation stress response protein Aaf1p/Yml081Wp regulates acetate production in Saccharomyces cerevisiae. PLoS ONE 7, e51551. doi:10.1371/journal.pone.0051551 [ Links ]
Wang, C., Mas, A. & Esteve-Zarzoso, B., 2016. The interaction between Saccharomyces cerevisiae and non-Saccharomyces yeast during alcoholic fermentation is species and strain specific. Front Microbiol. 7, 502. [ Links ]
Young, E., Poucher, A., Comer, A., Bailey, A. & Alper, H., 2011. Functional survey for heterologous sugar transport proteins, using Saccharomyces cerevisiae as a host. Appl. Environ. Microbiol. 77, 3311-3319. [ Links ]
Zoecklein, B.W., Fugelsang, K.C., Gump, B.H. & Nury, F.S., 1995. Wine analysis and production. Chapman & Hall, New York. [ Links ]
Zou, H., Hastie, T. & Tibshirani, R., 2006. Sparse principal component analysis. J. Comput. Graph. Stat. 15, 265-286. [ Links ]
Zuzuarregui, A., Monteoliva, L., Gil, C. & Del Olmo, M., 2006. Transcriptomic and proteomic approach for understanding the molecular basis of adaptation of Saccharomyces cerevisiae to wine fermentation. Appl. Environ. Microbiol. 72, 836-847. [ Links ]
Submitted for publication: November 2016
Accepted for publication: April 2017
Acknowledgements: The authors wish to thank the Agricultural Research Council (ARC) for the infrastructure and/or financial support; Anchor Yeast (Rymco) for financial support; the National Research Foundation/Research and Technology Fund (NRF/RTF) (RTF grant UID 98693); the National Research Foundation/Technology and Human Resources for Industry Programme (NRF/THRIP) (THRIP grant numbers UID 71526 and 86086); Drs Mardé Booyse and Maré Vlok, for the statistical and quantitative LC-based iTRAQ proteomic analysis; and Mr C.D. Abrahams, for technical assistance. The opinions, findings and conclusions expressed in this publication are those of the authors, and the National Research Foundation accepts no liability in this regard
* Corresponding author: E-mail address: hartr@arc.agric.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}