










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.37 n.2 Stellenbosch 2016
ORIGINAL RESEARCH ARTICLES
Relationships between water status, leaf chlorophyll content and photosynthetic performance in Tempranillo vineyards
G. HailemichaelI; A. CatalinaII; M.R GonzalezII; P. MartinII, *
IJimma Agricultural Research Centre, P.O. Box 192, Jimma, Ethiopia
IIDepartamento de Producción Vegetal y Recursos Forestales, Universidad de Valladolid, Avda. de Madrid 57, 34004 Palencia, Spain
ABSTRACT
The aims of this work were (i) to study the relationships between leaf water potential, foliar chlorophyll content and photosynthetic performance of grapevines under field conditions, and (ii) to evaluate the feasibility of chlorophyll fluorescence parameters to assess water status in commercial vineyards. During three consecutive seasons (2011 to 2013),20 non-irrigated "Tempranillo" vineyard subzones were monitored in Ribera del Duero, Spain. Data on foliar chlorophyll content, midday and predawn leaf water potential, net assimilation and chlorophyll fluorescence parameters were measured at fruit set, pre-véraison and véraison. The results show that the water potential indicated weak to moderate stress in the study years and decreased along the vegetative cycle, while foliar chlorophyll contents showed an increasing trend. The correlation between both variables was not consistent. Linear regression of the chlorophyll fluorescence parameters in dark-adapted leaves, Fo, Fv and Fv/Fm, over chlorophyll content was strong in all the study years (higher determination coefficients for Fv/Fm at fruit set in 2012 and 2013). In general, the water potential with low coefficients of variation showed a low effect on photosynthesis activity, although values in variables such as Fm, Fv, Fv/Fm, Fs/Fo and ΦPSII registered an increasing trend when midday leaf water potential decreased in the pre-véraison and véraison stages. These variables could be useful to estimate water status in commercial vineyards for application in precision viticulture.
Key words: chlorophyll fluorescence, chlorosis, photosynthesis, stress, Vitis vinifera L.
INTRODUCTION
Conditions such as extreme air temperature, changes in photoperiod, light intensity and quality, nutrient abundance and starvation, drought, flooding or excessive salts hinder the full expression of the genetic potential of plants and are limiting to yield and fruit quality (Ciais et al., 2005). There is a lot to research on the relationships between soil water availability to the grapevine, water loss through evapotranspiration, grapevine photosynthesis, and yield and grape composition (Medrano et al., 2003; Van Leeuwen et al., 2009; Alves et al., 2013). In temperate climates, water deficit conditions are necessary to produce high-quality red wine (Van Leeuwen et al., 2009), but stress as a result of excessive water deficit may lead to significant productivity and quality losses (Ojeda et al., 2002). Vine water status is dependent on soil and climate characteristics, and its spatial characterisation in vineyards is an important step in precision viticulture.
Drought limits photosynthesis, even in plants such as grapevines that are well adapted to this condition (Chaves, 1991; Adir et al., 2003). Under conditions of mild water stress, stomatal conductance (gs), internal CO2 concentration and/or net assimilation (An) decline and the An/gs ratio increases (Flexas et al., 2000; Van Leeuwen et al., 2009). When water stress is more severe, non-stomatal regulation of photosynthesis is observed, including decreased light-and CO2-saturated photosynthesis (Escalona et al., 1999), reduced photochemistry (Flexas et al., 1999; 2002a), depressed activity of photosynthetic enzymes (Maroco et al., 2002) and decreased mesophyll conductance (Flexas et al., 2002a).
Photosynthetic pigments allow plants to absorb energy from light, so foliar chlorophyll content is a key factor affecting the performance of plant photosynthesis (Taiz & Zeiger, 2006). Various reports have explained that drought stress significantly decreases the chlorophyll a, chlorophyll b and total chlorophyll content of different crops (Mafakheri et al., 2010; Gholamin & Khatnezhad, 2011). However, the presence of low levels of chlorophyll in leaves (chlorosis) is a general symptom of stress and might be poorly related to water status in the field when other environmental factors interact.
The analysis of chlorophyll fluorescence parameters is considered as an important approach to evaluating the health or integrity of the photosynthetic apparatus within a leaf, and provides a rapid and accurate technique for detecting and quantifying the interaction among and tolerance of plants to stress (Maxwell & Johnson, 2000; Lichtenthaler et al., 2005). The intensity of water stress in vines has been significantly correlated with non-photochemical quenching and minimal, maximum, variable and steady-state fluorescence in light-adapted leaves (Souza et al., 2004; Catalina et al., 2011). The ratio of steady-state to minimal fluorescence in dark-adapted leaves (Fs/Fo) has also been reported to be useful in the early detection of water stress (Flexas et al., 2002b). On the other hand, the fluorescence variables measured in dark-adapted leaves may be strongly affected by other factors, such as iron deficiency chlorosis (Bavaresco et al., 2006; Martín et al., 2007; Catalina et al., 2011; Zarco-Tejada et al., 2013).
Most of the studies that have linked photosynthesis and water status in grapevines, mentioned above, were carried out in single irrigation experiments, applying different water regimes to vines under controlled conditions. It was regarded as interesting for precision viticulture to contrast the impact of water status on photosynthetic activity in commercial vineyards, where variables such as soil characteristics or cultivation practices may modify its effects. The objectives of this work therefore were (i) to study the relationships between water status, foliar chlorophyll content and photosynthetic performance of grapevines under field conditions, and (ii) to evaluate the feasibility of chlorophyll fluorescence parameters to assess water status in vineyards for application in precision viticulture.
MATERIALS AND METHODS
Description of study site
The study was conducted in 2011, 2012 and 2013 on 24 non-irrigated vineyard subzones in Pesquera de Duero, Valladolid (Ribera del Duero Designation of Origin, North-Central Spain), at an altitude of 800 m above sea level. The subzones (10 m x 10 m each) were selected with different soil depths, soil texture and topography to assure maximum variability in water availability across the sites, in terms of the purposes of the study. The vineyards were planted with the 'Tempranillo' cultivar, were 10 to 14 years old and grafted on 110-Richter rootstock. Vines were spaced at 3 m x 1.5 m (2 222 plants per hectare), pruned on a double cordon Royat system, trellis trained and with vertically positioned shoots.
The soils in the study area are calcareous, very basic and poor in organic matter (Martín et al., 2008). Texture ranged from medium to medium-weighted. Concentrations of active carbonate (3.3 to 15.5%) and DPTA extractable iron (2.3 to 6.4 mg/kg) were highly heterogeneous within the area. These soil properties, along with the presence of a lime-sensitive rootstock (110-Richter), led to different levels of iron deficiency chlorosis in the vineyards of the area. The subzones had a high variability of soil concentrations of extractable potassium, phosphorus and magnesium (Zarco-Tejada et al., 2013).
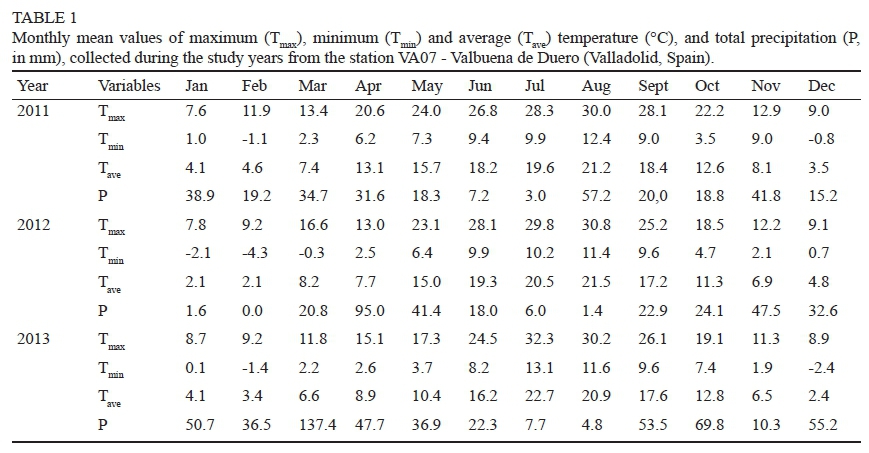
The study area has a Mediterranean climate, with low temperatures in the winter and hot and dry summers. Mean annual temperatures were 11.2°C, 11.4°C and 11.1°C in 2011, 2012 and 2013 respectively. The rainfall registered from 1 April to 30 September was 137 mm, 185 mm and 173 mm in the first, second and third year respectively (Table 1). Without irrigation, this insufficient water supply in soils varying in texture and root explorable depth ensured a broad variability in soil water content in the study zones.
Field data collection
Data on foliar chlorophyll content, midday and predawn leaf water potential, net assimilation and chlorophyll fluorescence variables were measured in the fruit set, pre-véraison (approximately 20 to 28 days after fruit set) and véraison growth stages.
Foliar chlorophyll content was recorded with a CL-01 portable colorimeter (Hansatech Instruments Ltd., Norfolk, UK). In each subzone, 30 leaves were selected by choosing the fourth or fifth leaf, counting from the first sheet of the apex. Chlorophyll content (Chl, in μg/cm2 leaf) was calculated from the CL-01 meter readings (R) by the regression line obtained for cv. Tempranillo by González et al. (2005), as Chl = 6.0817R + 7.6084.
Net assimilation (An: μmol CO2/m2/s) and chlorophyll fluorescence parameters were monitored using a LI-Cor 6400 portable infrared gas analyser (IRGA) equipped with a 640040 leaf chamber pulse amplitude modulation fluorometer (Li-Cor, Inc. Lincoln, Nebr., USA). Fluorescence parameters included minimum (Fo) and maximum (Fm) fluorescence in dark-adapted leaves (Fo' and Fm' under light conditions), variable (Fv) and steady-state fluorescence (Fs), efficiency (ΦPSII) and maximum efficiency (Fv/Fm and Fv'/Fm') of photosystem II, apparent rate of electron transport (ETR), and photochemical (qP) and non-photochemical quenching (NPQ).
Nine fully expanded leaves exposed to direct solar radiation and located at the fourth or fifth node from the apex were selected in each subzone to conduct the photosynthesis measurements. The chlorophyll fluorescence parameters of the dark-adapted leaves were obtained before dawn (01:00 to 03:00 local time), while the levels of gas exchange and fluorescence parameters of the light-adapted leaves were obtained at midday (11:00 to 13:00 local time). Leaf chamber conditions were set constant for all measurements conducted: incident photosynthetic photon flux density on the leaves was set to 1 500 μmol photons/m2/s photosynthetic saturation in field-grown grapevines (Escalona et al., 1999); and CO2 levels in the analyser were held constant at 370 μmol CO2/mol air. Temperature, air pressure and relative humidity were maintained at ambient levels. Air flow rate through the leaf chamber was maintained at 500 μmol/s. Leaf absorbances measured at 635 nm and 465 nm were entered into the IRGA for photosynthetic measurements taken at each study site. These values were estimated from leaf greenness data, previously obtained with a CL-01 portable colorimeter as described above (Gonzalez et al., 2005).
Measurements of predawn leaf water potential (ψPd) were taken during the two hours before dawn. The vine is in equilibrium with the water potential of the soil at that time (Winkel & Rambal, 1993). Measurements of midday leaf water potential (ψMd) were performed within two hours of solar noon (13:00 to 15:00 local time). The leaf water potential was monitored with a Scholander pressure chamber (Solfranc Technologies SL, Spain). Values were measured in six exposed leaves in each subzone, on the same shoot/branch that was utilised for the photosynthesis and chlorophyll fluorescence measurements.
Statistical analysis
The relationships of the leaf water potential, foliar chlorophyll content and photosynthesis variables were tested with linear regression analysis. Analysis of variance (ANOVA) and Tukey's test were performed to test mean differences between years. Data analysis was performed with version 9.2 of the SAS software package (SAS Institute Inc., Cary NC, USA).
RESULTS AND DISCUSSION
Water status and foliar chlorophyll content
Table 2 shows the variation in the ranges of leaf water potential in the study zones in each phenological stage and year studied. It is interesting to note that predawn leaf water potential (Ψpd) always registered higher variation coefficients than midday leaf water potential (ΨΜd). The variability of water status between subzones was clearly greater in 2011 than in 2012 and 2013 (Table 2), when the precipitation registered during the vegetative cycle was lower (Table 1).
In general, the records of midday and predawn leaf water potential at fruit set and véraison in the study years indicated moderate to weak water deficit, according to the ranges from Van Leeuwen et al. (2009), although in some stages, such as pre-véraison in 2013 (Table 2), the ΨΜd values correspond to moderate to severe water deficit ranges. Normally, values of leaf water potential decrease through the vegetative cycle (Table 2). ΨΜd and ΨΜd decreased as the precipitation became very scanty, the soil water reserves dropped and the temperature increased from spring to summer (Medrano et al., 2003).
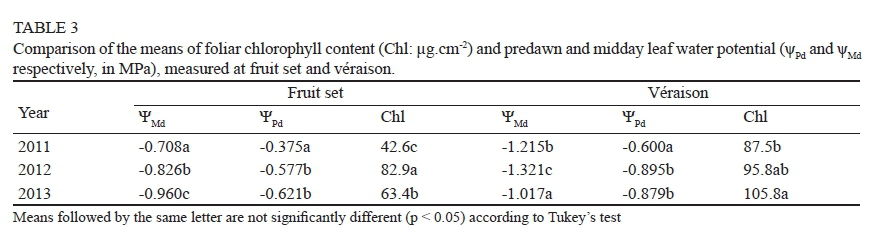
The ANOVA of leaf water potential and foliar chlorophyll content (Chl) at fruit set and véraison was significant according to year from the explanatory variables (data not shown), evidence of the existence of a high variability in meteorological conditions in the study years. From a comparison of the means, the values of both and ΨPd at fruit set decreased from 2011 to 2013 (Table 3), while more negative values were registered at véraison in 2012.
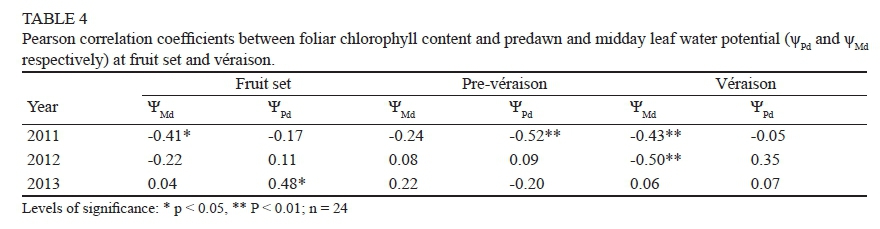
The mean values of Chl at fruit set in 2012 were higher than in 2013, but at véraison the result was reversed (Table 3). Lower levels of leaf water potential (more water-stressed vines) corresponded to lower values of Chl at fruit set and véraison in 2011; however, this relationship was not repeated in other years. These results gave a hint that water status and foliar chlorophyll content appear to be independent. In fact, no consistent correlations were obtained between these variables at different growth stages and in different years of study (Table 4). Negative correlations were observed at fruit set, pre-véraison and véraison in 2011, and at véraison in 2012, but the correlation coefficient of Chl with ΨPd at fruit set in 2013 was positive.
Foliar chlorophyll content, which is part and parcel of the vigour of plants, is influenced by soil and environmental factors. Various reports have explained that drought stress significantly decreases the chlorophyll content in crops such as chickpeas (Mafakheri et al., 2010) or maize (Gholamin & Khatnezhad, 2011). In the present study, Chl was not a good indicator of the water status of grapevines. This might be due to both the existence of narrow ranges of variation in leaf water potential (slight to moderate stress), and the influence of nutrient status (iron deficiency chlorosis), which could strongly affect the foliar chlorophyll content in the study area (Martín et al., 2008).
Chlorophyll content and photosynthesis activity
Foliar chlorophyll content per unit area of leaf is considered to be an indicator of the photosynthetic capacity of plants (Taiz & Zeiger, 2006; Ling et al., 2011). In the present study, the linear regression of An over Chl was significant and had a positive slope at the pre-véraison stage in 2011 (Table 5), but no significant regressions were obtained in any of the other years and at any of the other measuring times.
As Table 5 shows, the linear regressions of chlorophyll fluorescence parameters in dark-adapted leaves, Fo, Fv and Fv/Fm, over Chl were significant in all the study years (higher determination coefficients for Fv/Fm at fruit set in 2012 and 2013). The positive relationship between Fv/Fm and Chl confirms the results of Palliotti et al. (2009), who showed that high levels of leaf photosynthetic pigments might enhance light absorption, thus increasing the maximum quantum yield of photosystem II. In relation to this, Mandal et al. (2009) observed that Fv/Fm was higher in healthy leaves than in plants infected by downy mildew (with chlorosis).
Fo measured at fruit set resulted in a significant and negative regression in all study years, while the relationship between both variables was not significant at more advanced phenological stages (Table 5). Significant and positive linear regressions were obtained between Fs/Fo and Chl in all vegetative stages in 2011, and at fruit set in 2013. The regressions of qP, NPQ and ETR over Chl was not statistically significant in most years and growth stages. Fluorescence variables in light-adapted leaves, Fo', Fm', Fs and Fv'/Fm', showed significant positive regression over Chl only at pre-véraison and véraison in 2011, and at pre-véraison in 2013.
The significant relationships observed between Fo, Fv/ Fm and ΦPSII and Chl, repeated over three years, might be associated with the high variability in Chl generated within the study zone by the incidence of iron stress (Martín et al., 2008). A low chlorophyll concentration (chlorosis) in young leaves is a visible symptom of iron deficiency, which has a significant negative effect on photosynthesis activity (Maxwell & Johnson, 2000). A shortage of physiologically active iron leads to a decrease in the electron transport rate, as well as to a lowering of the efficiency of photosystem II (Bavaresco et al., 2006; Martín et al., 2007).
Water status and photosynthetic activity
It is well known that the net assimilation by leaves decreases as water potential decreases (Chaves, 1991; Lawlor, 2002). Stomatal closure is one of the first responses of plants to the lack of water in the soil (Chaves, 1991; Flexas et al., 2002a). After stomatal closure, water deficit induces a decline in photosynthetic activity by reducing the availability of CO2 in the leaf mesophyll (Chaves, 1991; Flexas et al., 2002a). In the present study, linear regression relations of An over ψMd and ψΜd were not consistent (Table 6). However, the relationships between water potential and some chlorophyll fluorescence parameters reflect that the water status had a significant influence on photosynthetic performance, as explained below.
Minimum fluorescence (Fo) showed a significant and positive relationship with midday leaf water potential at fruit set in 2011, while no other significant relations were obtained in the rest of the phenological stages and years (Table 6). Other chlorophyll fluorescence variables measured in dark-adapted leaves, such as Fm, Fv and Fv/Fm, showed a significant and negative relationship with midday leaf water potential at fruit set and véraison in 2011 and at pre-véraison and véraison in 2012. On the other hand, relationships between Fm, Fv and Fv/Fm and predawn leaf water potential were weak, except for values registered at véraison in 2013.
A sustained decrease in dark-adapted Fv/Fm and increase in Fo indicate the occurrence of damage to photosystem II, which reduces the efficiency of absorbed light energy transfer from the light-harvesting complex. For Maxwell and Johnson (2000), this may result from various environmental stresses, including high temperature or water deficit. Nevertheless, Baker and Rosenqvist (2004) have stated that water stress has no major impact on the efficiency of photosystem II. In this regard, Medrano et al. (2003) did not detect significant differences in Fv/Fm between irrigated and non-irrigated grapevines of cv. Tempranillo. Souza et al. (2004) observed a tendency for higher Fo values and lower Fm in water-stressed versus non-stressed plants, but a slight decrease in Fv/Fm in stressed plants was confirmed only with intense water stress.
Water status showed a better relationship with chlorophyll fluorescence variables measured in the dark-adapted than in the light-adapted leaves (Table 6). Both types of variables showed a negative relationship with leaf water potential in the most cases, disagreeing with Alves et al. (2013), who demonstrated that ΦPSII, ETR and qP decreased significantly in non-irrigated in comparison to irrigated grapevines. On the other hand, the current results show that water status was not linked with steady-state fluorescence, as has been explained by other authors (Maxwell & Johnson, 2000; Flexas et al., 2002b).
Although most of the regressions were not significant (Table 6), it can be seen that midday leaf water potential was related more than predawn leaf water potential to chlorophyll fluorescence variables in light-adapted leaves. These results confirm previous findings obtained in the same study area in 2010 (Catalina et al., 2011). Flexas et al. (2002a) showed that light-adapted chlorophyll fluorescence variables, such as Fo', Fm', Fv', Fv'/Fm' and Fs, had a strong relationship with midday leaf water potential.
The regressions of Fs/Fo with ψMd at fruit set in 2011 and at pre-véraison in 2012 were significant and had a negative slope (Table 6), and slightly increased the coefficients of determination obtained with Fs. Flexas et al. (2002b) reported that Fs/Fo is the best modulated test for the early detection of water stress in plants. The relationship could be negative or positive, depending on the stress level. A positive correlation is observed between Fs/Fo and photosynthetic photon flux density under irrigation, but an inverse correlation develops as drought progresses (Flexas et al., 1999).
Non-photochemical quenching (NPQ) resulted in a significant relationship only with ψPd at pre-véraison in 2012 and at véraison in 2013 (Table 6). NPQ reflects the thermal dissipation of excessive excitation energy in the chloroplasts, and has a strong link with water status, but not with Fv/Fm (Maxwell & Johnson, 2000). The present results disagree with those of Flexas et al. (1999; 2000) and Alves et al. (2013), who showed a strong increase in NPQ as drought progressed. It is possible that, in the current study, there were no subzones with sufficiently high levels of water stress for this.
CONCLUSIONS
The presented results have revealed that foliar chlorophyll content cannot be considered an indicator of water stress in vineyards when the spatial variation of water status is limited and/or overlapped by a variation in iron or other nutrient status. Nevertheless, a linear regression of the chlorophyll fluorescence variables on foliar chlorophyll content was significant and positive in more of the growth stages and study years.
Linear regression relationships of chlorophyll fluorescence parameters with leaf water potential showed little significance in all the study years. However, variables such as Fm, Fv, Fv/Fm, Fs/Fo and ΦPSII showed an increasing trend when midday leaf water potential decreased at pre-véraison and véraison. These variables could help assess the water status of vineyards for application in precision viticulture. In future studies it would be interesting to study the use of chlorophyll fluorescence variables as indicators of yield and grape quality in vineyards with significant spatial variability in water status.
LITERATURE CITED
Adir, N., Zer, H., Shochat, S. & Ohad, I., 2003. Photoinhibition - A historical perspective. Photosynth. Res. 76, 343-370. [ Links ]
Alves, F., Costa, J., Costa, P., Correia, C., Gonçalves, B., Soares, R. & Pereira, J.M., 2013. Grapevine water stress management in Douro Region: Long-term physiology, yield and quality studies in cv. Touriga Nacional. In: Group of International Experts of Vitivinicultural Systems for Co-Operation (ed.). Proc. 18th Int. Symp. GiESCO, July 2013, Porto, Portugal. [ Links ]
Baker, N.R. & Rosenqvist, E., 2004. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 55, 1607-1621. [ Links ]
Bavaresco, L., Bertamini, M. & Iacono, F., 2006. Lime-induced chlorosis and physiological responses in grapevine (Vitis vinifera L. cv. Pinot blanc) leaves. Vitis 45, 45-46. [ Links ]
Catalina, A., Gonzalez, R., Gonzalez, M.R., Zarco-Tejada P.J. & Martín, P., 2011. Iron and water stress, photosynthetic efficiency differently affects vine photosynthetic efficiency and grape composition. Proc. 34th World Congress of Vine and Wine, June 2011, Porto, Portugal. [ Links ]
Chaves, M.M., 1991. Effects of water deficits on carbon assimilation. J. Exp. Bot. 42, 1-16. [ Links ]
Ciais, P., Reichstein, M., Viovy, N., Granier, A., Ogée, J., Allard, V., Aubinet, M., Buchmann, N., Bernhofer, Chr., Carrara, A., Chevallier, F., De Noblet, N., Friend, A.D., Friedlingstein, P., Grünwald, T., Heinesch, B., Keronen, P., Knohl, A., Krinner, G., Loustau, D., Manca, G., Matteucci, G., Miglietta, F., Ourcival, J.M., Papale, D., Pilegaard, K., Rambal, S., Seufert, G., Soussana, J.F., Sanz, M.J., Schulze, E.D., Vesala, T. & Valentini, R., 2005. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 472, 529-533. [ Links ]
Escalona, J.M., Flexas, J. & Medrano, H., 1999. Stomatal and non-stomatal limitations of photosynthesis under water stress in field grown grapevines. Aust. J. Plant Physiol. 26, 421-433. [ Links ]
Flexas, J., Bota, J., Escalona, J.M., Sampol, B. & Medrano, H., 2002a. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 29, 461-471. [ Links ]
Flexas, J., Briantais, J.M., Cerovic, Z., Medrano, H. & Moya, I., 2000. Steady-state and maximum chlorophyll fluorescence responses to water stress in grapevine leaves. A new remote sensing system. Remote Sens. Environ. 73, 282-297. [ Links ]
Flexas, J., Escalona, J.M. & Medrano, H., 1999. Water stress induces different photosynthesis and electron transport rate regulation in grapevine. Plant Cell & Environ. 22, 39-48. [ Links ]
Flexas, J., Escalona, J.M., Evain, S., Gulías, J., Moya, I., Osmond, C.B. & Medrano, H., 2002b. Steady-state chlorophyll fluorescence (Fs) measurements as a tool to follow variations of net CO2 assimilation and stomatal conductance during water-stress in C3 plants. Physiol. Plant. 114, 231-240. [ Links ]
Gholamin, R. & Khayatnezhad, M., 2011. The effect of end season drought stress on the chlorophyll content, chlorophyll fluorescence parameters and yield in maize cultivars. Sci. Res. Essay 6, 5351-5357. [ Links ]
González, R., Nunez, L.C., Martin, P., Berjon, A. & Zarco-Tejada, P. J., 2005. Estimación de la absorbancia de radiación PAR en hojas de vid a partir de su contenido en clorofila. In: Asociación Portuguesa de Ciencias Hortícolas (ed.). Proc. 5th Congreso Ibérico de Ciencias Hortícolas, Actas Portuguesas de Horticultura, Vol. 2, May 2005, Oporto, Portugal. pp. 384 -389. [ Links ]
Lawlor, D.W., 2002. Limitation to photosynthesis in water-stressed leaves: Stomata vs. metabolism and the role of ATP. Ann. Bot. 89, 871-885. [ Links ]
Lichtenthaler, H.K., Buschmann, C. & Knapp, M., 2005. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica 43, 379-393. [ Links ]
Ling, Q., Huang, W. & Jarvis, P., 2011. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 107, 209-214. [ Links ]
Mafakheri, A.B., Siosemardeh, P.C., Bahramnejad, Y., Struik, T. & Sohrabi, S., 2010. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 4, 580-585. [ Links ]
Mandal, K., Saravanan, R., Maiti, S. & Kothari, I.L., 2009. Effect of downy mildew disease on photosynthesis and chlorophyll fluorescence in Plantago ovata Forsk. J. Plant Dis. Prot. 116, 164-168. [ Links ]
Maroco, J.P., Rodrigues, M.L., Lopes, C. & Chaves, M.M., 2002. Limitations to leaf photosynthesis in field-grown grapevine under drought-metabolic and modelling approaches. Funct. Plant Biol. 29, 451-459. [ Links ]
Martín, P., Gonzalez, M.R., Núnez, L.C., Gonzalez, R. & Zarco-Tejada P.J., 2007. Influencia de la clorosis férrica sobre el rendimiento fotosintético del vinedo cv. Tempranillo. In: GIENOL (ed). Actas de las IX Jornadas de los Grupos de Investigación Enológica, May 2007, Badajoz, Spain. pp. 437 - 439. [ Links ]
Martín, P., Zarco-Tejada, P.J. & González, M.R., 2008. Diagnostico nutricional y recomendaciones de abonado en suelos calizos de la Ribera de Duero. Vida Rural 270, 26-32. [ Links ]
Maxwell, K. & Johnson, G.N., 2000. Chlorophyll fluorescence - A practical guide. J. Exp. Bot. 51, 659-668. [ Links ]
Medrano, H., Escalona, J.M., Cifre, J., Bota, J. & Flexas, J., 2003. A ten-year study on the physiology of two Spanish grapevine cultivars under field conditions: Effects of water availability from leaf photosynthesis to grape yield and quality. Funct. Plant Biol. 30, 607-619. [ Links ]
Ojeda, H., Andara, C., Kraeva, E., Carbonneau, A. & Deloire, A., 2002. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Syrah. Am. J. Enol. Vitic. 53, 261-267. [ Links ]
Palliotti, A., Silvestroni, O. & Petoumenou, D., 2009. Photosynthetic and photoinhibition behavior of two field grown grapevine cultivars under multiple summer stresses. Am. J. Enol. Vitic. 60, 189-198. [ Links ]
Souza, R.P., Machado, E.C., Silva, J.A.B., Lagöa, A.M.M.A. & Silveria, J.A.G., 2004. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exper. Bot. 51, 45-56. [ Links ]
Taiz, L. & Zeiger, E., 2006. Plant physiology. Sinauer Associates Inc., Massachusetts, USA. [ Links ]
Van Leeuwen, C., Tregoat, O., Choné, X., Bois, B., Pernet, D. & Gaudillère, J.-P., 2009. Vine water status is a key factor in grape ripening and vintage quality for red Bordeaux wine. How can it be assessed for vineyard management purposes? J. Int. Sci. Vigne Vin 43, 121-134. [ Links ]
Winkel, T. & Rambal, S., 1993. Influence of water stress on grapevines growing in the field: From leaf to whole-plant response. Aust. J. Plant Physiol. 20, 143-157. [ Links ]
Zarco-Tejada, P.J., Catalina, A., Gonzalez, M.R. & Martín, P., 2013. Relationships between net photosynthesis and steady-state chlorophyll fluorescence retrieved from airborne hyperspectral imagery. Remote Sens. Environ. 136, 247-258. [ Links ]
Submitted for publication: April 2016
Accepted for publication: July 2016
Acknowledgements: The authors acknowledge the financial support of the Spanish Ministry of Science and Education (MEC) for project RTA2014-00077-C02-02, and of the Regional Government of Castilla y Leon for project VA011A10-2. The authors are grateful to Bodegas Emilio Moro S.L., for their collaboration in conducting this research.
* Corresponding author: pmartinp@pvs.uva.es

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}