










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.36 n.2 Stellenbosch 2015
Effects of adding unsaturated fatty acids on fatty acid composition of saccharomyces cerevisiae and major volatile compounds in wine
L.L. Duan; Y. Shi; R. Jiang; Q. Yang; Y.Q. Wang; P.T. Liu; C.Q. Duan; G.L. Yan*
Centre for Viticulture and Oenology, College of Food Science and Nutritional Engineering, China Agricultural University, Beijing 100083, P.R. China
ABSTRACT
The synergistic influences of three unsaturated fatty acids (UFAs, namely linoleic acid, oleic acid and linolenic acid) on the fatty acid composition of Saccharomyces cerevisiae and major volatile compounds were investigated in synthetic grape juice. The addition of UFAs led to a corresponding increase in UFAs in the cellular lipid, which was accompanied by a reverse reduction in the content of medium-chain saturated fatty acids (C6:0 to C14:0) and little variation in that of long-chain saturated fatty acids (C16:0 to C24:0). The supplementation of UFAs considerably improved yeast growth and fermentation activity and, in particular, increased the concentrations of most volatile compounds in wine, including higher alcohols (2-phenylethanol, 2-methyl-1-propanol and 3-(methylthio)-1-propanol), medium-chain fatty acids (butanoic acid, hexanoic acid and octanoic acid), acetate esters (isoamyl acetate and 2-phenylethyl acetate) and all ethyl esters. Remarkable linear relationships were further found between ethyl esters and the concentration of the added UFAs (R2 from 0.909 to 0.996), which significantly intensified the fruity, flowery and sweet attributes of the final wine, as assessed by calculating the odour activity values. Our results suggest that rationally increasing the concentration of UFAs is not only a practical method to improve yeast fermentation activity, but also a potential approach to manipulating wine aroma.
Key words: Wine yeast, unsaturated fatty acids, fatty acid composition, aroma compounds
INTRODUCTION
Unsaturated fatty acids (UFAs) are required for Saccharomyces cerevisiae to grow under anaerobic conditions. They are essential not only for maintaining membrane integrity and function, but also for adapting to fermentation stresses, such as high sugar and ethanol toxicity (Holcberg & Margalith, 1981; You et al., 2003; RupCic & Juresic, 2010). A higher degree of unsaturation of the cell membrane can maintain higher membrane fluidity and guarantee the activity of membrane-associated enzymes and transporters, such as ATPase and general amino acid permease (GAP) (Calderbank et al., 1985; Rosa & Sa-Correia, 1992). The main UFAs of S. cerevisiae are oleic acid (18:1) and palmitoleic acid (16:1), which are derived from the desaturation of the saturated fatty acids (SFAs) palmitic acid (16:0) and stearic acid (18:0) by the oxygen-dependent enzyme Δ9 desaturase encoded by the OLE1 gene respectively (Yamada et al., 2005). Because it lacks Δ12-fatty acid desaturase and ω3-fatty acid desaturase, S. cerevisiae cannot produce polyunsaturated fatty acids (PUFAs), such as linoleic acid (18:2n6) and α-linolenic acid (18:3n3). The ability of PUFAs to maintain membrane fluidity is higher than that of monounsaturated fatty acids (MUFAs), and they thus offer better protection to cells under harsh conditions (Yazawa et al., 2009).
The lipid and fatty acid composition of yeast cell membranes can be modified by fermentation temperature, oxygen availability, growth rate and the presence of sterols, which in turn influence cell growth and fermentation activity (Fujii et al., 1997; Redon et al., 2009) and strongly affect the formation of volatile metabolites, including esters, acetate and ethyl esters, higher alcohols and medium-chain fatty acids (MCFAs), all of which are relevant to wine aroma (Thurston et al., 1982; Mauricio et al., 1997; Beltran et al.,, 2008; Varela et al., 2012). During wine fermentation, the absence of oxygen suppresses the fatty acid desaturation of yeast. An alternative to the biosynthesis is the direct uptake of UFAs from the grape juice, which can be used to avoid yeast stuck fermentation (Varela et al., 2012).
Grape berries contain 0.15% to 0.24% (wet weight basis) lipids (Gallander & Peng, 1980). UFAs represent the major components of the total lipids. Linoleic acid (C18:2n6) is the most abundant, followed by oleic acid (C18:1n9) and α-linolenic acid (C18:3n3) (Ancin et al., 1998; Santos et al., 2011). The concentrations of UFAs in grape must depend largely on the grape cultivar (Ancin et al., 1998) and technological fermentation procedures, such as grape must clarification (Varela et al., 1999) and grape-skin maceration (Valero et al., 1998). Several studies have investigated the effect of exogenous UFAs on intracellular fatty acid composition or wine aroma biosynthesis during wine fermentation. Redon et al. (2009) found that supplementation with palmitoleic acid (C16:1) can shorten the chain length and reduce total fatty acids (including UFAs and SFAs) compared with the control, which increases the viability of the yeast and reduces the duration of wine fermentation. The profile of aroma compounds was not available in this study. The influence of exogenous UFAs on aroma compounds is associated with the type and concentration of UFAs. The addition of oleic acid (31 mg/L) and ergosterol (25 mg/L) can increase the production of higher alcohols and acetate esters, except for 1-butanol and 1-pentanol, which exhibit the opposite trend (Mauricio et al., 1997). Thurston et al. (1982) found that the presence of linoleic acid (50 mg/L) can suppress the production of ethyl esters and acetate esters. In one study of a synergistic effect, Tween 80, containing 70% of oleic acid and 30% of palmic acid and stearic acid, was added and improved the esters, higher alcohols and volatile fatty acids of wine (Varela et al., 2012).
Because the three UFAs coexist in grape must and the concentrations vary with grape cultivar and fermentation procedure, it is essential from the wine production perspective to investigate their synergistic effect on the yeast fatty acid composition of yeast cells and yeast-derived aroma compounds during wine fermentation. To date, however, no completed information is available. Therefore, in this study we investigated the synergistic effects of UFAs, including linoleic acid, oleic acid and linolenic acid, on yeast fermentation activity and the fatty acid compositions of cells and volatile aroma compounds. To achieve this purpose, four series concentrations of UFA mixtures were set according to their concentration and ratio in real grape must, which were initially added to the medium (Gallander & Peng, 1980; Delfini et al., 1992; Ancin et al., 1998; Valero et al., 1998; Varela et al., 1999; Santos et al., 2011). To facilitate this investigation, a simplified, chemically defined medium (MS300) that resembles the nutrient composition of grape juice was used.
MATERIALS AND METHODS
Yeast strains and culture media
The S. cerevisiae strain EC1118 (Lallemand Inc. Denmark) was used in this study. The composition of the synthetic grape must MS300 was similar to that used in previous research (Bely et al., 1990), with a little modification. Because the purpose of the study was to investigate the effect of UFAs on yeast cell metabolism, Tween 80 and sodium oleate were not added. The components in the MS300 medium were as follows: main compounds (in g/L): glucose (240), citric acid (6), malic acid (6), KH2PO4 (0.75), K2SO4 (0.5), MgSO4-7H2O (0.25), CaCl2-2H2O (0.155), NaCl (0.2); mineral salts (mg/L): MnSO4-H2O (4), ZnSO4 (4), CuSO4-5H2O (1), KI (1), CoCl2-6H2O (0.4), H3BO3 (1), NaMoO4-H2O (1); vitamins (in mg/L): myo-inositol (20), nicotinic acid (2), calcium pantothenate (1.5), thiamine hydrochloride (0.25), pyridoxine hydrochloride (0.25), biotin (0.003); nitrogen compounds (in mg/L): proline (612.6), glutamine (505.3), arginine (374.4), tryptophan (179.3), alanine (145.3), glutamic acid (120.4), serine (78.5), threonine (759.2), leucine (48.4), aspartic (44.5), valine (44.5), phenylalanine (37.9), isoleucine (32.7), histidine (32.7), methionine (31.4), tyrosine (18.3), glycine (18.3), lysine (17), cysteine (13.1), ammonium chloride (460); lipids (in mg/L): ergosterol (15). The UFA mixture (including linoleic acid, oleic acid and α-linolenic acid, purchased from Sigma, purity > 98%) was added to the medium in the concentrations shown in Table 1. The minimum UFAs were required to ensure normal cell growth and fermentation under anaerobic conditions (Aceituno et al., 2012), which served as the control (Table 1, medium A). The concentration of UFAs in media B, C and D were four-, eight- and 12-fold higher than that in medium A respectively. The proportions of the three UFAs were determined according to the results of previous studies (Ancin et al., 1998; Santos et al., 2011).
Fermentation conditions
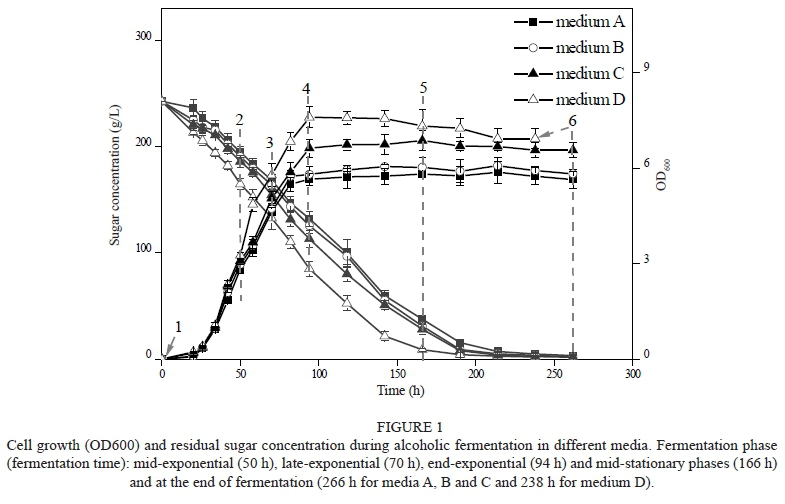
Yeast strains were pre-cultured aerobically in YEPD (1% yeast extract, 2% peptone, 2% glucose) at 120 rpm and 28 °C for approximately 15 h to the late exponential phase. Before the inoculums, nitrogen was sparged to eliminate the oxygen from the medium. The inoculations were in 500 mL flasks equipped with glass capillary stoppers containing 300 mL of MS300 medium. The initial yeast inoculums (1 x 106 CFU/mL) were cultivated statically at 25 °C. Fermentation was carried out in three biological repeats. The progress of fermentation was monitored daily by measuring cell density (OD600) and sugar consumption. Samples taken from the fermentation flasks were immediately centrifuged to collect the cell-free supernatants for the analysis of the main fermentation products and aroma compounds and the yeast cells for the analysis of intracellular fatty acids. UFA changes in the media and the intracellular fatty acids during fermentation were analysed at the pre-fermentation (0 h), mid-exponential (50 h), late-exponential (70 h), end-exponential (94 h) and mid-stationary phases (166 h), and at the end of fermentation (266 h for media A, B and C and 238 h for medium D) (Fig. 1). The analysis of the main fermentation products and aroma compounds were conducted at the end of fermentation. All fermentation samples were stored at -20 °C until analysis. Each analysis was repeated in triplicate for each biological repeat.
Analysis of main fermentation products
The concentrations of the main fermentation products, glucose, fructose, ethanol, glycerol, malic acid, citrate acid and acetic acid, were determined by high-performance liquid chromatography (HPLC, 1200 series, Agilent Technologies, Inc., Palo Alto, CA, USA) using an Aminex HPX-87H column (300 mm χ 7.8 mm, Bio-Rad, CA, USA), as described in a previous study but with a little modification (López & Gómez, 1996). The mobile phase flow rate of 0.6 ml/min was achieved with an elution gradient composed of 5 nmol/L H2SO4 in water: (1) The injection volume was 20 µL for the glucose, fructose, glycerol and alcohol analysis. The column temperature was 45°C with a refractive index detector (RID). The total analysis time was 30 min. (2) The injection volume was 10 μL for the malic acid, citric acid and acetic acid analysis. The column temperature was 60 °C, with a diode array detector (210 nm, DAD). The total analysis time was 30 min.
Analysis of fatty acids
The analysis of the fatty acids in the yeast cells was conducted according to the procedure of Redon et al. (2009). Yeast cell samples were placed in sealed tubes and saponified with 1 mL of 5% NaOH in 50% methanol/water. The saponification was conducted at 100 °C for 30 min. The saponified part then was cooled to room temperature, and 2 mL HCl 6 M was added to the tubes. Free fatty acids were extracted by adding 0.5 mL of hexane:methyl-tert-butyl ether (1:1, v/v). The tubes were vortexed twice for 30 sec each. The organic phase was collected after centrifugation (3 000 rpm, 3 min).
Fatty acids were quantified with a 7890NGC gas chromatograph equipped with a 5975BMS mass spectrum system (GC-MS, Agilent, USA) on an HP-INNOWAX column (60 m χ 0.25 i.d., 0.25 μm film thickness, J&W Scientific, USA) under a helium carrier gas (1 ml/min). The temperature was programmed from 80 °C (held for 1 min) to 220 °C at 25 °C/min and from 220 °C to 250 °C (held for 20 min) at 5 °C/min. One microlitre of extract was injected in splitless mode. The temperature of the transfer line and the injector were 280 °C and 250 °C respectively. The mass spectrometer was operated in the electron impact (EI) mode at 70 eV, and in the selective ion mode (SIM) range of 20 to 350 m/z. The calibration was previously performed with pure standard chemicals.
Analysis of aroma compounds
The analysis of the final wine odour aromas was conducted according to previous research (Zhang et al., 2007), as follows. Five millilitres of the final fermentation sample, 1.00 g of NaCl and 10 μL of 4-methyl-2-pentanol (1.039 mg/mL water, internal standard) were blended in a 15 mL sample vial that was tightly capped with a PTFE silicon septum and contained a magnetic stirrer. Subsequently, the vial containing the sample was heated at 40°C for 30 min on a heating platform by agitation (80 rpm). A 7890NGC gas chromatograph equipped with a 5975BMS mass spectrum system (GC-MS, Agilent, USA) on an HP-INNOWAX column (60 m χ 0.25 i.d., 0.25 μm film thickness, J&W Scientific, USA) was applied for the analysis of the aroma compounds. The pre-treated (conditioned at 270 °C for 1 h) SPME fibre (50/30 μm DVB/Carboxen/PDMS, Supelco, Bellefonte, PA, USA) was then inserted into the headspace and extracted for 30 min with continued heating (40 °C) and agitation (80 rpm). The fibre was instantly desorbed in the GC injector for 8 min at 250 °C. The GC inlet was set in the splitless mode. The starting temperature of the oven was held at 50 °C for 1 min and then increased to 220 °C at a rate of 3 °C/min and held there for 5 min. The mass spectrometer was operated in the electron impact (EI) mode at 70 eV, and in the scan and selective ion mode (SIM) range of 35 to 350 m/z. The calibration was previously performed with pure standard chemicals based on previous studies (Howard et al., 2005).
Establishment of odour activity value and aromatic series
To evaluate the contribution of each aroma compound to the total odour profile, the odour activity value (OAV) was calculated by dividing the concentration of a compound by its odour threshold (OT, determined from the wine matrix) that was obtained from the literature cited in Table 4. To obtain the whole aroma profile of the wine, an aromatic series was established by grouping the OAVs of the volatiles exhibiting similar odour descriptions (Peinado et al., 2006; Wu et al., 2011).
Statistical analysis
Analysis of variance (ANOVA, least significant difference method at a significance level of P < 0.05) was used to evaluate differences between the volatile and non-volatile compounds in the fermentation samples by the SPSS statistical package version 19.0 (SPSS Inc., USA).
RESULTS AND DISCUSSION
Yeast growth and sugar consumption
Fig. 1 shows the progress of fermentation with different UFA concentrations. The addition of UFAs greatly improved yeast growth and sugar consumption. Cells reached the stationary phase after 100 h of fermentation in all cases. The highest biomass (OD600 values) was observed in medium D, followed by media C, B and A. The sugar concentration in medium D was reduced below 2 g/L after 238 h of fermentation and was shortened by 28 h compared with that of the three other cases. Correspondingly, a positive relationship between initial UFA concentration and the maximum biomass (OD600 values, R2 = 0.9514) and maximum glucose consumption rates (R2 = 0.9567) was established. These results confirmed the conclusion that the initial addition of UFAs can improve yeast growth and increase yeast fermentation activity (Redon et al., 2009).
Main fermentation products
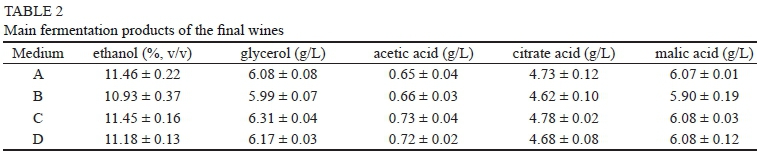
The final concentrations of ethanol, glycerol, acetic acid, citric acid and malic acid were further determined, and the results show that there was no significant difference among the different samples (Table 2). This finding suggests that the variation in the UFA concentrations did not have a considerable influence on the primary metabolites of the sugar.
Fatty acid composition of yeast cells
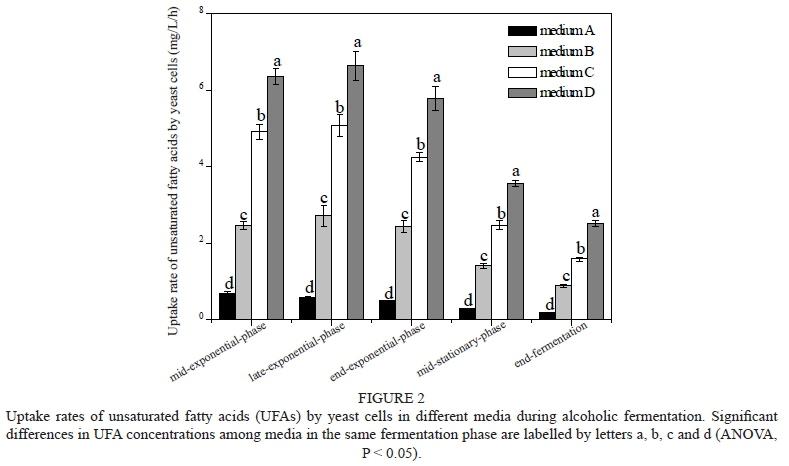
It generally is believed that the improvement in cell growth and fermentation performance that is caused by UFAs results from the modification of the composition of intracellular fatty acids. We thus determined the fatty acid content of the cells. First, the profiles of the supplemented UFA concentrations in the media during fermentation were investigated. The results showed that the three UFAs were rapidly taken up by yeast cells and that the UFA levels decreased rapidly. The highest UFA uptake rates were observed in medium D during fermentation, followed by media C, B and A (Fig. 2). In four cases, the UFAs were almost completely consumed after 94 h of fermentation when the yeast cells reached the end-exponential phase (data not shown). Correspondingly, the intracellular UFA content increased significantly and the level of particular UFAs was positive following the supplemented UFA concentration (Fig. 3a). The incorporation of UFAs led to an increase in UFAs in the total fatty acid content from 74.35% in medium A to 80.55% in medium C and 83.55% in medium D when the cells reached the mid-exponential phase; the highest UFA levels were obtained at this time. After the mid-exponential phase, the UFA content gradually decreased and finally was maintained at a constant level after the end-exponential phase in all samples, but the values with UFAs addition were all higher than those of the control (medium A). Our results were consistent with a previous study, which showed that the UFA concentration of yeast cells declined as the fermentation progressed due to cell growth under anaerobic fermentation (Venter et al., 1989). The incorporation of UFAs resulted in the highest total concentration of FAs observed in medium D, followed by yeast in media C, B and A (Fig. 3d, ANOVA, P < 0.05).
It should be noted that palmitoleic acid (C16:1n-9), which is considered one of the main fatty acids in the composition of yeast cells, was not detected in our research (data not shown). This can be ascribed to the fact that fatty acid desaturation cannot occur in yeast cells under anaerobic conditions (Snoek & Steensma, 2007). Our results demonstrate that the enrichment of UFAs in cells is essential for yeast adaptation to environmental stress during alcoholic fermentation (You et al., 2003; RupCic & JuresiC, 2010).
Long-chain saturated fatty acids (LSFAs, C16:0 to C24:0, 15.54% to 39.38%) were the second largest group of fatty acids in the total fatty acids, followed by medium-chain fatty acids (MCFAs C6:0 to C14:0, 0.74% to 9.32%). Surprisingly, the variation in intracellular UFA concentration had no significant effect on LSFA synthesis, but it did have a considerable impact on the content of MCFAs. The highest concentration of MCFAs was observed in yeast cultivated in medium A, followed by media B, C and D (Fig. 3b, ANOVA, P < 0.05), which suggests that there is a negative relationship between the intracellular MCFAs and UFAs. At present we cannot fully explain the phenomenon; however, two deductions were proposed according to previous studies. First, Dufour et al. (2008) suggested that the increase in UFAs in yeast cells can lead to the release of MCFAs from the fatty acid synthase complex (FAS1 and FAS2) and further secrete them into the medium. This phenomenon was also observed in our study, as shown below. The secretion might be partially responsible for the reduction in MCFA content in the cells. An alternative explanation, proposed by Redon et al. (2011), states that, in addition to UFAs, MCFAs are functional compounds that help yeast cells adapt to stress conditions by increasing the membrane fluidity. The incorporation of exogenous UFAs into the cell membrane might be efficient for cells to handle stress conditions and may thus weaken the synthesis of MCFAs. The detailed mechanism needs to be investigated further.
Aroma compounds
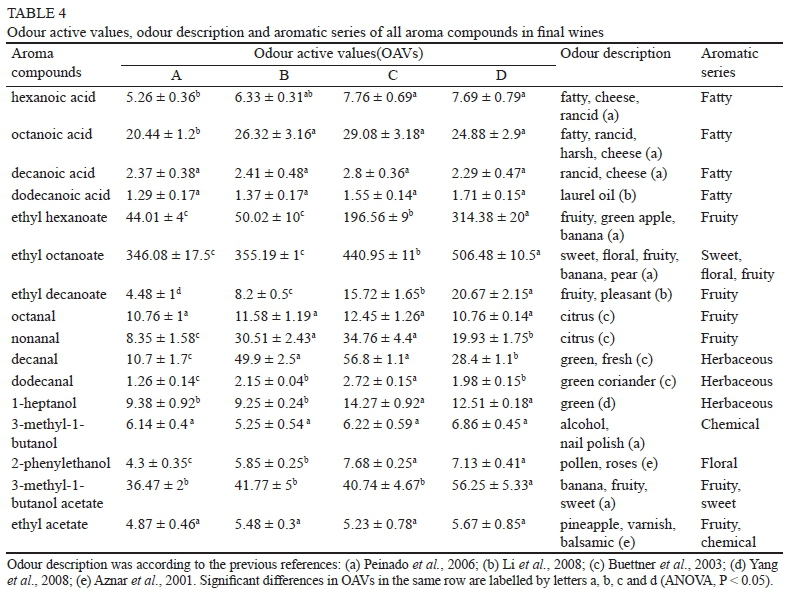
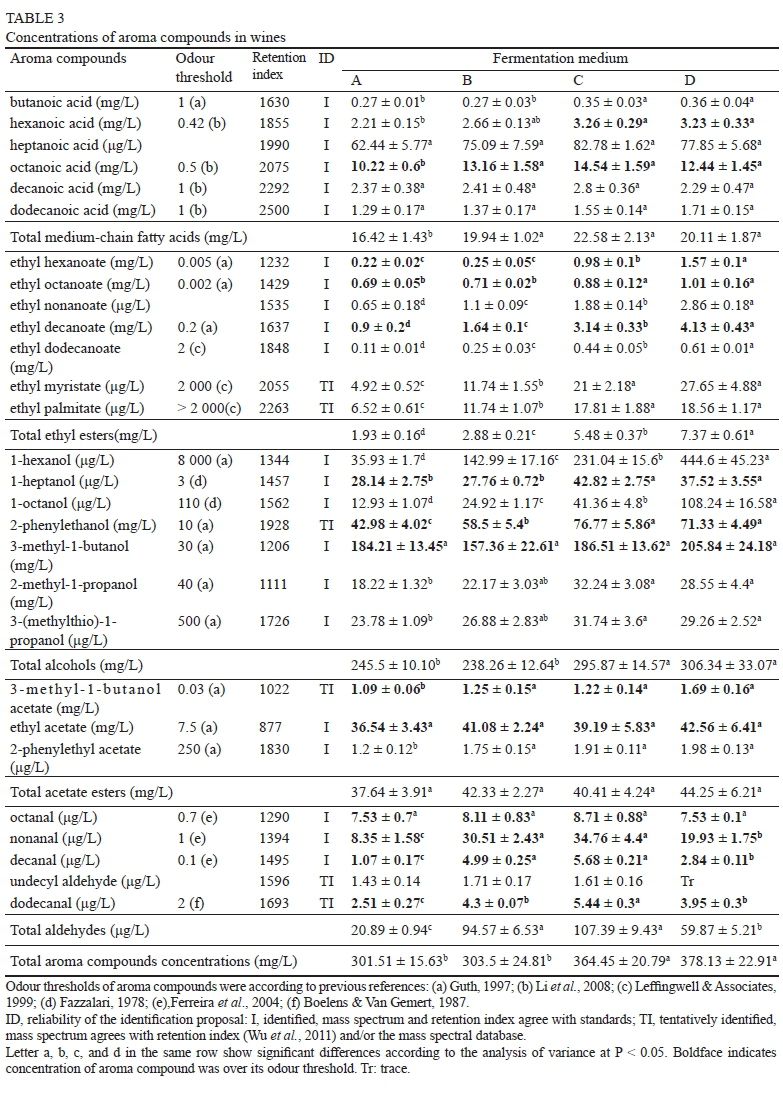
Although they had no considerable impact on the primary metabolic compounds of sugar, the variations in UFAs significantly affected the synthesis of wine aroma compounds. Generally, the volatile compounds were promoted by the addition of UFAs, and the total concentrations in the final wines of C and D were 1.21- and 1.25-fold that observed in wine A respectively (Table 3, ANOVA, P < 0.05). Moreover, the concentrations of ethyl esters, which are the main aroma-contributing compounds, in wines C and D were 1.84- and 2.82-fold higher than that in wine A respectively. Twenty-eight aroma compounds were detected in our study - six MCFAs, seven alcohols, five aldehydes, seven ethyl esters and three acetate esters. Of these, 22 compounds could be identified, and six compounds could be identified tentatively (Table 3). With the exception of undecyl aldehyde in wine D, all of the aroma compounds could be quantified in all the samples.
Medium-chain fatty acids
MCFAs (C6 to C12) are produced primarily by yeasts as intermediates in the biosynthesis of long-chain fatty acids and are prematurely released from the fatty acid synthase (FAS) complex. These volatile fatty acids can directly contribute to the flavour of wine, or serve as substrates that participate in the formation of ethyl acetates. In contrast to the intracellular MCFAs, the addition of UFAs resulted in a greater accumulation of MCFAs in the medium. This result can be ascribed to the repression of the gene encoding acetyl-CoA carboxylase (ACC1) by exogenous UFAs, which causes the release of MCFAs from the fatty acid synthase complex (FAS1 and FAS2) and their secretion into the environment (Dufour et al., 2008). Among the six identified MCFAs, octanoic acid was the highest, followed by hexanoic acid, decanoic acid, dodecanoic acid, butanoic acid and heptanoic acid. The highest concentrations of octanoic acid, hexanoic acid and butanoic acid were found in wine C (14.59, 3.26 and 0.35 mg/L) and wine D (12.44, 3.23 and 0.36 mg/L), compared with those of wine A (10.22, 2.21 and 0.27 mg/L) (Table 3, ANOVA, P < 0.05).
Ethyl esters
The C6, C8 and C10 medium-chain fatty acid ethyl esters are important flavour-active metabolites that are responsible for the highly desired fruit aroma characters in wine. They are generated by an alcohol and the acyl group derived from activated MCFAs. Among the seven identified ethyl esters, ethyl decanoate was quantitatively the highest, followed by ethyl octanoate, ethyl hexanoate and ethyl dodecanoate. Higher contents of ethyl esters were formed in wines C and D, and the total concentrations were 1.84- and 2.82-fold higher than that in wine A (Table 3, ANOVA, P < 0.05)
respectively. In addition, significant positive relationships between ethyl esters, particularly ethyl hexanoate ('fruity' odour) and ethyl octanoate ('fruity', 'floral' and 'sweet' odour), and the added UFA concentrations were established (R2 from 0.909 to 0.996). Sufficient MCFAs are crucial for the higher biosynthesis of pleasant ethyl esters (Saerens et al., 2008); therefore, the improvement observed for the ethyl esters following fatty acid treatment by UFAs can be ascribed to the accumulation of the substrate MCFAs in the medium. This conclusion was supported further by the observation that linear correlations existed between MCFAs and the corresponding ethyl esters (data not shown).
Alcohols
Seven alcohols were identified in our research, including three fatty alcohols (1-hexanol, 1-heptanol and 1-octanol) and four higher alcohols (2-phenylethanol, 3-methyl-1-butanol, 2-methyl-1-propanol and 3-(methylthio)-1-propanol). The detailed mechanism of fatty alcohol biosynthesis by wine yeast is not fully understood. Our results indicate that treatment with UFAs can increase the production of fatty alcohols. The highest concentrations of 1-hexanol and 1-octanol were found in wine D (444.60 μg/L and 108.24 μg/L vs 35.93 and 12.93 μg/L in wine A respectively), whereas the highest concentration of 1-heptanol was in wine C (42.82 μg/L vs 28.14 μg/L in wine A) (Table 3, ANOVA, P < 0.05).
The higher alcohols are predominantly formed by yeast during fermentation from a-keto acids, a process that involves the degradation of an amino acid (i.e., valine, leucine, isoleucine, threonine and phenylalanine) via the so-called Ehrlich pathway (Hazelwood et al., 2008). High levels of 2-phenylethanol, 2-methyl-1-propanol and 3-(methylthio)-1-propanol were observed in wines C and D. Higher alcohols can have both a positive and negative impact on the aroma and flavour of wine, depending on their concentrations. Excessive concentrations (exceeding 400 mg/L) can result in a strong, pungent smell, whereas an optimal level (below 300 mg/L) imparts a pleasant character (Bartowsky & Pretorius, 2009). In this study, total concentrations of 295.9 mg/L and 306.3 mg/L were found in wines C and D respectively, which suggests that higher alcohols contributed positively to the aroma of the wine. At present, we cannot fully explain why the addition of exogenous UFAs promotes higher alcohol synthesis, but we inferred that it might be associated with the improvement of amino acid transportation into cells because it has been demonstrated that S. cerevisiae growing in medium enriched with UFAs (linoleic acid and oleic acid) can induce general amino acid permease (GAP) activity and increase the trans-shipment of amino acids into yeast cells (Calderbank et al., 1984, 1985), which is beneficial for the synthesis of higher alcohols (Hernândez-Orte et al., 2006). The detailed mechanism needs to be investigated further.
Acetate esters
Acetate esters are another important aroma compound that are generated by the reaction of acetyl-CoA with higher alcohols that are formed from the degradation of amino acids or carbohydrates and have a significant effect on the fruity flavour of wine. The acetate esters detected in our study included ethyl acetate ('pineapple', 'varnish' and 'balsamic' odour, Table 3), isoamyl acetate ('banana' and 'sweet' odour, Table 3) and 2-phenylethyl acetate ('honey', 'roses' and 'flowery' odour). No significant difference was found in ethyl acetate in all the samples, although higher concentrations of 3-methyl-1-butanol acetate (isoamyl acetate) and 2-phenylethyl acetate accumulated in wines C and D (Table 3, ANOVA, p < 0.05). This was in contrast to the results of a previous study, which showed that acetate esters were inhibited by exogenous UFAs due to the repression of ATF1 transcription, the gene encoding the enzyme (alcohol acetyltransferase) that catalyses the synthesis of acetate esters (Fujii et al., 1997). Our results indicate that, despite the negative regulation, increased UFA concentrations can enhance the production of acetate esters by increasing yeast growth and substrate production (higher alcohol).
Aldehydes
High levels of decanal and dodecanal can cause an unpleasant, intense, 'green' odour in wine, whereas a lower content of C8 to C11 aldehydes can contribute to a citrus fruit aroma (Culleré et al., 2011). The effects of UFAs on the production of aldehydes were different from other aroma compounds, which were largely dependent on the UFA concentration. Relatively high UFA contents (wines B and C) resulted in higher concentrations of nonanal, decanal and dodecanal compared with wine A, whereas further increasing the UFA content (wine D) led to a great reduction in the aldehyde level (Table 3, ANOVA, P < 0.05). The OAVs decreased from 34.8 , 56.8- and 2.7-fold in wine C to 19.9-, 28.4- and 2.0-fold in wine D respectively, as shown below. This finding implies that manipulating the initial UFA concentration potentially can regulate the influence of aldehydes on wine aroma.
Influence of adding UFAs on odour profile of final wine
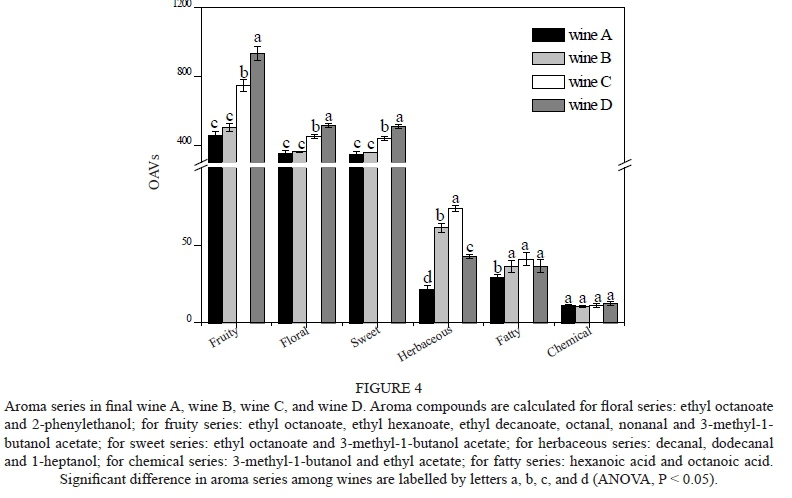
The OAV can assess the extent of the potential sensory aroma contribution for each volatile compound. An aroma compound with an OAV over one is considered to make a flavour contribution to wine (Guth, 1997). As shown in Table 4, 16 aroma compounds had an OAV above 1 in our study. According to previous research (Aznar et al., 2001; Buettner et al., 2003; Peinado et al., 2006; Li et al., 2008; Yang et al., 2008), these compounds are mainly associated with the attributes 'banana', 'apple', 'pear', 'strawberry', 'citrus' 'sweet', 'roses', 'fruity', 'fatty', 'cheese', 'rancid', 'nail polish', 'balsamic' and 'green' (Table 4). The OVAs of ethyl hexanoate (fruity, green apple, banana attributes) and ethyl decanoate (fruity attribute) in wine D were 6.15- and 3.62-fold higher than those of wine A. Other important volatile aroma compounds, including ethyl octanoate, 2-phenylethanol and 3-methyl-1-butanol acetate, also had higher OAVs in wines C and D than in wine A. They certainly are responsible for the increment of fruity and flowery attributes in wines C and D, as shown below.
The odour of a compound is described in terms of several descriptors that are agreed upon by experts, and an aromatic series can be defined as a group of volatile compounds with similar odour descriptors (Wu et al., 2011). To better understand the influence of UFAs on the wine odour profile and the contribution of various volatile compounds to the olfactory impression of wine, an aromatic series was further established in our study by combining the OAVs of a group of volatiles with similar odour descriptions (Peinado et al., 2006; Wu et al., 2011). Six aromatic series of volatile compounds were established: fruity, floral, sweet, herbaceous, chemical and fatty (Fig. 4). Of these, the fruity series was prominent, followed by the flowery, sweet, herbaceous, fatty and chemical series. The higher OAVs of ethyl esters, higher alcohol and acetate esters (such as ethyl octanoate, ethyl hexanoate, 2-phenylethanol and 3-methyl-1-butanol acetate) led to wines C and D featuring the fruity, flowery and sweet series (Fig. 4, ANOVA, P < 0.05). Although wines C and D also had relatively higher OAVs in the herbaceous and fatty series, their influence on the total odour profile was limited due to the low concentration or high odour threshold of a particular compound, such as dodecanal, 1-heptanol and 1-hexanol (Table 3). As a result, there was no considerable discrepancy between the herbaceous and fatty series among the wines. Similarly, no significant difference was observed among the samples for the chemical series. Based on these results, it can be concluded that an increased initial UFA concentration in the medium can intensify the fruity, floral and sweet attributes of wine and thus improve the odour profile of the wine.
The purpose of the present study was to evaluate the synergistic impact of three UFAs on volatile compounds from the point of view of practical production. Clearly, to fully understand the mechanism of how UFAs regulate aroma compounds it is essential to investigate the separate influence of individual UFAs. In addition, the influence of yeast strain specificity should be considered. These related experiments are being conducted in our laboratory. In this study, we used the synthetic grape juice MS300 medium to eliminate the influence of grape cultivar aroma compounds and aroma precursors on the production of yeast-derived aroma compounds; however, the utilisation of natural grape must may change the effect of the variation in UFA concentration on the types and levels of volatile compounds, because complex flavour precursors (such as glycoside aroma molecules) (Loscos et al., 2007) and other enzyme-catalysed reactions (such as a 1ipoxygenase-catalysed reactions) (Deluc et al., 2009) may occur. Therefore, further work is needed to evaluate the extent to which changing the initial UFA concentration affects the production of wine aroma compounds in real grape must.
CONCLUSIONS
The present results indicate that supplementation with mixed UFAs (including linoleic acid, oleic acid and linolenic acid) can correspondingly increase the intracellular UFA content, which simultaneously leads to the reverse reduction of MCFA content but does not influence LSFA synthesis. In addition to improving yeast cell growth and fermentation activity, the additions of mixed UFAs promoted the most volatile aroma compounds, including MCFAs, higher alcohols, acetate esters and all ethyl esters. A linear relationship between UFA concentrations and all ethyl esters was established, which significantly intensified the fruity, floral and sweet attributes of the final wine. Our results suggest that rationally increasing the UFA concentration in grape must could not only improve yeast fermentation activity, but also manipulate wine aroma.
LITERATURE CITED
Aceituno, F.F., Orellana, M., Torres, J., Mendoza, S., Slater, A.W., Melo, F. & Agosin, E., 2012. Oxygen response of the wine yeast Saccharomyces cerevisiae EC1118 grown under carbon-sufficient, nitrogen-limited enological conditions. Appl. Environ. Microbiol. 78, 8340-8352. [ Links ]
Ancin, C., Ayestaran, B., Garcia, A. & Garrido, J., 1998. Evolution of fatty acid contents in Garnacha and Viura musts during fermentation and the aging of wine. Z. Lebensm. Unters. F. A. 206, 143-147. [ Links ]
Aznar, M., López, R., Cacho, J.F. & Ferreira, V., 2001. Identification and quantification of impact odorants of aged red wines from Rioja. GC- olfactometry, quantitative GC-MS, and odor evaluation of HPLC fractions. J. Agric. Food Chem. 49, 2924-2929. [ Links ]
Bartowsky, E.J. & Pretorius, I.S., 2009. Microbial formation and modification of flavor and off-flavor compounds in wine. In: Biology of Microorganisms on Grapes, in Must and in Wine (1st ed). Springer-Verlag Berlin, Germany. pp. 209 - 231. [ Links ]
Beltran, G., Novo, M., Guillamón, J.M., Mas, A. & Rozès, N., 2008. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 121, 169-177. [ Links ]
Bely, M., Sablayrolles, J.M. & Barre, P., 1990. Automatic detection of assimilable nitrogen deficiencies during alcoholic fermentation in oenological conditions. J. Ferment. Bioeng. 70, 246-252. [ Links ]
Boelens, M. & Van Gemert, L., 1987. Organoleptic properties of aliphatic aldehydes. Perfumer & Flavorist 12, 31-43. [ Links ]
Buettner, A., Mestres, M., Fischer, A., Guasch, J. & Schieberle, P., 2003. Evaluation of the most odour-active compounds in the peel oil of clementines (Citrus reticulata Blanco cv. clementine). Eur. Food. Res. Technol. 216, 11-14. [ Links ]
Calderbank, J., Keenan, M.H. & Rose, A.H., 1985. Plasma-membrane phospholipid unsaturation affects expression of the general amino-acid permease in Saccharomyces cerevisiae Y185. J. Gen. Microbiol. 131, 57-65. [ Links ]
Calderbank, J., Keenan, M.H., Rose, A.H. & Holman, G.D., 1984. Accumulation of amino acids by Saccharomyces cerevisiae Y185 with phospholipids enriched in different fatty-acyl residues: A statistical analysis of data. J. Gen. Microbiol. 130, 2817-2824. [ Links ]
Culleré, L., Ferreira, V. & Cacho, J., 2011. Analysis, occurrence and potential sensory significance of aliphatic aldehydes in white wines. Food Chem. 127, 1397-1403. [ Links ]
Delfini, C., Conterno, L., Giacosa, D., Cocito, C., Ravaglia, S. & Bardi, L., 1992. Influence of clarification and suspended solid contact on the oxygen demand and long-chain fatty acid contents of free run, macerated and pressed grape musts, in relation to acetic acid production. Wein Wissenschaft 47, 69-75. [ Links ]
Deluc, L., Quilici, D., Decendit, A., Grimplet, J., Wheatley, M., Schlauch, K., Mérillon, J.-M., Cushman, J. & Cramer, G., 2009. Water deficit alters differentially metabolic pathways affecting important flavor and quality traits in grape berries of Cabernet Sauvignon and Chardonnay. BMC Genomics 10, 212. [ Links ]
Dufour, J., Malcorps, P. & Silcock, P., 2008. 21 Control of ester synthesis during brewery fermentation. In: Smart, K. (ed). Brewing yeast fermentation performance (2nd ed). Blackwell Science, Oxford, UK. pp. 213 - 233. [ Links ]
Fazzalari, F., 1978. Compilation of odor and taste threshold values data. ASTM data series. American Society for Testing and Materials, Philadelphia, USA. [ Links ]
Ferreira, V., Culleré, L., López, R. & Cacho, J., 2004. Determination of important odor-active aldehydes of wine through gas chromatography-mass spectrometry of their 0-(2, 3, 4, 5, 6-pentafluorobenzyl) oximes formed directly in the solid phase extraction cartridge used for selective isolation. J. Chromatogr. A. 1028, 339-345. [ Links ]
Fujii, T., Kobayashi, O., Yoshimoto, H., Furukawa, S. & Tamai, Y., 1997. Effect of aeration and unsaturated fatty acids on expression of the Saccharomyces cerevisiae alcohol acetyltransferase gene. Appl. Environ. Microbiol. 63, 910-915. [ Links ]
Gallander, J.F. & Peng, A.C., 1980. Lipid and fatty acid compositions of different grape types. Am. J. Enol. Vitic. 31, 24-27. [ Links ]
Guth, H., 1997. Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 45, 3027-3032. [ Links ]
Hazelwood, L.A., Daran, J.-M., Van Maris, A.J., Pronk, J.T. & Dickinson, J.R., 2008. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 74, 2259-2266. [ Links ]
Hernández-Orte, P., Ibarz, M., Cacho, J. & Ferreira, V., 2006. Addition of amino acids to grape juice of the Merlot variety: Effect on amino acid uptake and aroma generation during alcoholic fermentation. Food Chem. 98, 300-310. [ Links ]
Holcberg, I. & Margalith, P., 1981. Alcoholic fermentation by immobilized yeast at high sugar concentrations. Euro. J. Appl. Microbiol. Biotech. 13, 133-140. [ Links ]
Howard, K.L., Mike, J.H. & Riesen, R., 2005. Validation of a solid-phase microextraction method for headspace analysis of wine aroma components. Am. J. Enol. Vitic. 56, 37-45. [ Links ]
Leffingwell & Associates, 1999. Available at http://www.leffingwell.com./esters1.htm [ Links ]
Li, H., Tao, Y.S., Wang, H. & Zhang, L., 2008. Impact odorants of Chardonnay dry white wine from Changli County (China). Eur. Food Res. Technol. 227, 287-292. [ Links ]
López, E.F. & Gómez, E.F., 1996. Simultaneous determination of the major organic acids, sugars, glycerol, and ethanol by HPLC in grape musts and white wines. J. Chromatogr. Sci. 34, 254-257. [ Links ]
Loscos, N., Hernandez-Orte, P., Cacho, J. & Ferreira, V., 2007. Release and formation of varietal aroma compounds during alcoholic fermentation from nonfloral grape odorless flavor precursors fractions. J. Agric. Food Chem. 55, 6674-6684. [ Links ]
Mauricio, J.C., Moreno, J., Zea, L., Ortega, J.M. & Medina, M., 1997. The effects of grape must fermentation conditions on volatile alcohols and esters formed by Saccharomyces cerevisiae. J. Sci. Food Agric. 75, 155-160. [ Links ]
Peinado, R.A., Mauricio, J.C. & Moreno, J., 2006. Aromatic series in sherry wines with gluconic acid subjected to different biological aging conditions by Saccharomyces cerevisiae var. capensis. Food Chem. 94, 232-239. [ Links ]
Redon, M., Guillamon, J.M., Mas, A. & Rozes, N., 2009. Effect of lipid supplementation upon Saccharomyces cerevisiae lipid composition and fermentation performance at low temperature. Eur. Food Res. Technol. 228, 833-840. [ Links ]
Redon, M., Guillamon, J.M., Mas, A. & Rozes, N., 2011. Effect of growth temperature on yeast lipid composition and alcoholic fermentation at low temperature. Eur. Food Res. Technol. 232, 517-527. [ Links ]
Rosa, M.F. & Sa-Correia, I., 1992. Ethanol tolerance and activity of plasma membrane ATPase in Kluyveromyces marxianus and Saccharomyces cerevisiae. Enzyme Microb. Tech. 14, 23-27. [ Links ]
Rupĕié, J. & Jurešié, G.Č., 2010. Influence of stressful fermentation conditions on neutral lipids of a Saccharomyces cerevisiae brewing strain. World J. Microb. Biotechnol. 26, 1331-1336. [ Links ]
Saerens, S., Delvaux, F., Verstrepen, K., Van Dijck, P., Thevelein, J. & Delvaux, F., 2008. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 74, 454-461. [ Links ]
Santos, L.P., Morais, D.R., Souza, N.E., Cottica, S.M., Boroski, M. & Visentainer, J.V., 2011. Phenolic compounds and fatty acids in different parts of Vitis labrusca and V. vinifera grapes. Food Res. Int. 44, 1414-1418. [ Links ]
Snoek, I.S.I. & Steensma, H.Y., 2007. Factors involved in anaerobic growth of Saccharomyces cerevisiae. Yeast 24, 1-10. [ Links ]
Thurston, P., Quain, D. & Tubb, R., 1982. Lipid metabolism and the regulation of volatile ester synthesis in Saccharomyces J. Instcerevisiae. . Brew. 88, 90-94. [ Links ]
Valero, E., Millan, M.C., Mauricio, J.C. & Ortega, J., 1998. Effect of grape skin maceration on sterol, phospholipid, and fatty acid contents of Saccharomyces cerevisiae during alcoholic fermentation. Am. J. Enol. Vitic. 49, 119-124. [ Links ]
Varela, C., Torrea, D., Schmidt, S., Ancin-Azpilicueta, C. & Henschke, P., 2012. Effect of oxygen and lipid supplementation on the volatile composition of chemically defined medium and Chardonnay wine fermented with Saccharomyces cerevisiae. Food Chem. 135, 2863-2871. [ Links ]
Varela, F., Calderon, F., Gonzalez, M.C., Colomo, B. & Suarez, J.A., 1999. Effect of clarification on the fatty acid composition of grape must and the fermentation kinetics of white wines. Eur. Food Res. Technol. 209, 439-444. [ Links ]
Venter, J.J., Vuuren, H.J.J., Tromp, A. & Randall, J.H., 1989. Relationship between fatty acid concentrations in wine yeasts and sugar fermentation at different temperatures. S. Afr. J. Enol. Vitic. 10, 44-48. [ Links ]
Wu, Y., Zhu, B., Tu, C., Duan, C. & Pan, Q., 2011. Generation of volatile compounds in litchi wine during winemaking and short-term bottle storage. J. Agric. Food Chem. 59, 4923-4931. [ Links ]
Yamada, T., Shimoi, H. & Ito, K., 2005. High expression of unsaturated fatty acid synthesis gene OLE1 in sake yeasts. J. Biosci. Bioeng. 99, 512-516. [ Links ]
Yang, D.S., Shewfelt R.L., Lee K.S. & Kays S.J., 2008. Comparison of odor-active compounds from six distinctly different rice flavor types. J. Agric. Food Chem. 56, 2780-2787. [ Links ]
Yazawa, H., Iwahashi, H., Kamisaka, Y., Kimura, K. & Uemura, H., 2009. Production of polyunsaturated fatty acids in yeast Saccharomyces cerevisiae and its relation to alkaline pH tolerance. Yeast 26, 167-184. [ Links ]
You, K.M., Rosenfield, C.L. & Knipple, D.C., 2003. Ethanol tolerance in the yeast Saccharomyces cerevisiae is dependent on cellular oleic acid content. Appl. Environ. Microbiol. 69, 1499-1503. [ Links ]
Zhang, M., Xu Q., Duan, C., Qu, W. & Wu, Y., 2007. Comparative study of aromatic compounds in young red wines from Cabernet Sauvignon, Cabernet Franc, and Cabernet Gernischet varieties in China. J. Food Sci. 72, C248-C252. [ Links ]
Submitted for publication: February 2015
Accepted for publication: May 2015
* Corresponding author: E-mail: glyan@cau.edu.cn
Acknowledgements: This work was supported financially by the China Agriculture Research System (CARS-30)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}