










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.35 n.2 Stellenbosch 2014
Environmental stress and aroma production during wine fermentation
S.C. Fairbairn; A.Y. Smit; D. Jacobson; B.A. Prior; F.F. Bauer*
Institute for Wine Biotechnology, Department of Viticulture and Enology, Private Bag X1, Stellenbosch University, Matieland 7602, South Africa
ABSTRACT
The sensory description of wine uses the widest range of descriptive terminology of all food products, reflecting the complex nature of a product whose character depends on the balance of hundreds of individual flavour-active compounds. There are many tools that can influence flavour profiles or wine styles, one of which is the choice of a specific yeast strain. Yeasts contribute to wine flavour by producing volatile metabolites with different flavour profiles. The impact of changing environmental conditions on the production of flavour compounds by yeast strains remains largely unexplored. This is the first study investigating the impact of two mild fermentation stresses, hyperosmotic and temperature stress, on aroma production in synthetic must by commercial Saccharomyces cerevisiae wine strains. Hyperosmotic stress was imposed by cultivation of the yeast for 21 days in the must containing either 0.3 or 0.5 M sorbitol. The transient temperature stresses were applied for 16 h at 8° or 37°C for either two or eight days after commencement of the fermentation. Greater glycerol and acetic acid levels were produced by most yeasts when only hyperosmotic stress was applied. Hyperosmotic and temperature stress conditions produced a limited number of significant changes to the profile of the esters, higher alcohols and volatile fatty acids. These changes differed significantly for each strain and stress treatment, suggesting that the fermentation conditions can significantly alter the aromatic profile of a wine, although these stress impacts cannot be predicted in general. The changes to the aromatic profile are specific to each individual wine yeast strain.
Key words: Wine aroma, hyperosmotic stress, temperature stress, synthetic grape must, fermentation
INTRODUCTION
Wine yeast strains mediate the near-complete conversion of high levels of sugar to ethanol and CO2 despite continuous exposure to various forms of stress (Bisson, 1999). The stresses may include temperature changes, hyperosmotic stress, ethanol stress, vitamin, mineral, nitrogen and oxygen deficiencies, and a low pH, among others (Attfield, 1997; Alexandre & Charpentier, 1998; Bisson, 1999; Bauer & Pretorius, 2000). There is evidence that such stresses are also often associated with problematic (stuck and sluggish) fermentations (Bisson, 1999; Gibson et al., 2007; Malherbe et al., 2007). The high initial sugar content of grape must results in hyperosmotic stress, which is counteracted by S. cerevisiae through the accumulation of intracellular glycerol in order to maintain a positive turgor pressure (Hohmann, 1997; Mager & Siderius, 2002; Tamás & Hohmann, 2003). Temperatures well below and above the growth optimum of yeast increase membrane permeability and disrupt transport systems into and out of the yeast cell (Bisson, 1999; Beales, 2004).
Yeast strains differ in their ability to sense and respond to stress. Most of these stresses have been studied extensively with regard to their impact on yeast growth and fermentation performance (Ivorra et al., 1999; Carrasco et al., 2001; Zuzuarregui & Del Olmo, 2004). Generally, yeast growth under stress is slowed, as its metabolism is adapting to meet the new environmental conditions. However, the literature provides few if any insights into the impact of fermentation-related stress on metabolite production that might influence wine flavour.
Wine flavour is defined as the cumulative effect of smell, taste and mouth-feel (Francis & Newton, 2005), and is a direct function of the chemical composition of the wine. The flavour- active metabolites that have an impact on wine perception are derived from the grapes and from microorganisms during fermentation, as well as from chemical processes during production and maturation (Rapp & Mandery, 1986; Rapp & Versini, 1996). Perceived flavour is the result of complex interactions between all the volatile and non-volatile compounds present in wine.
The volatile aromas produced de novo by wine yeast during fermentation include higher alcohols, esters and volatile fatty acids (Rapp & Versini, 1996). Higher alcohols are formed by the decarboxylation and reduction of α-keto acids, originating from either glycolysis or the Ehrlich pathway (Rapp & Mandery, 1986; Rapp & Versini, 1996; Swiegers et al., 2005; Styger et al., 2011). Volatile fatty acids are produced as a result of acetyl-CoA decarboxylation and condensation reactions (Lambrechts & Pretorius, 2000). Acetate esters are formed by enzyme-catalysed condensation reactions between acetyl-CoA and higher alcohols or ethanol (Lambrechts & Pretorius, 2000; Bell & Henschke, 2005; Bisson & Karpel, 2010), while ethyl esters are formed by the condensation reaction between ethanol and either a fusel acyl-CoA or a fatty acyl-CoA (Bisson & Karpel, 2010).
The metabolic pathways leading to the formation of these volatile aroma compounds are reasonably well mapped and established. However, the regulation of the metabolic flux through these networks is not well understood. Studies have shown that obvious factors, such as the availability of precursors, fermentation conditions (Henschke & Jiranek, 1993; Lambrechts & Pretorius, 2000; Saerens et al., 2008; Bisson & Karpel, 2010) and the genetic make-up of individual strains (Soles et al., 1982; Rossouw et al., 2008), play an important role in the modulation of aroma production. However, little data is available regarding the impact of environmental stress on the production of such metabolites.
Here we investigate the impact of two of the most common stresses experienced during alcoholic fermentation, namely temperature shifts and hyperosmotic stress, on the aroma production capacity of five wine yeast strains. To our knowledge, this is the first study investigating the impact of hyperosmotic and transient temperature stresses on the production of fermentation-derived volatile aroma compounds in synthetic grape must. The addition of the non-metabolised sugar alcohol, sorbitol, to the synthetic grape must to impose a hyperosmotic stress was considered to be the most appropriate means to study the impact on aroma production without increasing metabolised sugar levels (Hohmann, 1997). This preliminary study provides information on how the exposure to stress affects wine aroma and whether the observed changes are conserved among different commercial Saccharomyces yeast strains.
MATERIALS AND METHODS
Yeast strains and growth conditions
Fermentations were conducted using synthetic grape must (100 g/L glucose and 100 g/L fructose), as previously described (Henschke & Jiranek, 1993), with the exception of the nitrogen additions. The nitrogen sources (amino acid and ammonium; Bely et al., 1990) were added to reach a level of 250 mg/L nitrogen. The medium was estimated to have an osmotic stress of 1.11 osm.
The commercial Saccharomyces yeast strains used in this study were Lalvin EC1118, Cross Evolution (referred to as 285 in this paper) (Lallemand Inc, Montreal, Canada), NT50, VIN7 and VIN13 (Anchor Yeast, Cape Town, South Africa). Strains VIN7, VIN 13 and NT50 are S. cerevisiae hybrids, whereas EC1118 is S. cerevisiae var. bayanus (also known as Prise de Mousse strain). The Cross Evolution strain is a natural cross between S. cerevisiae strains selected for specific winemaking properties. Strains were stored at -80°C, and were streaked out onto YPD plates, which subsequently were incubated at 30°C. A single colony was used to inoculate 5 mL of synthetic grape must, which was aerobically incubated at 30°C overnight. These cultures were used to inoculate the must at an initial OD600 of 0.1, corresponding to ~1 x 106 cells/mL.
Fermentation conditions
The fermentations were performed in 100 mL glass bottles, containing 70 mL of synthetic grape must. The bottles were sealed with rubber stoppers and a CO2 outlet was provided. Fermentations were conducted in triplicate at 20°C for 21 days, unless stated otherwise. To induce hyperosmotic stress, sorbitol was added upon inoculation to a final concentration of 0.3 M (S1; ~1.41 osm) or 0.5 M (S2; ~1.61 osm). To evaluate the impact of temperature stress, a set of fermentation bottles was exposed to 8°C (T8) or 37°C (T37) for 16 h either on day two (D2) or day eight (D8) of the fermentation, and subsequently returned to 20°C. Fermentations not subjected to any additional stresses served as the control data set. These stresses were chosen to represent conditions that might be experienced during must fermentation. Fermentation bottles were weighed to monitor the fermentation progress as CO2 weight loss.
Chemical analysis
Residual glucose and fructose, ethanol and glycerol
Synthetic wine samples were scanned using the Winescan FT120 instrument (FOSS Analytical A/S software version 2.2.1) equipped with a purpose-built Michelson interferometer (FOSS Analytical A/S, Hillered, Denmark), and Fourier transform mid-infrared (FT-MIR) spectra were generated. Quantified chemical data for residual glucose, fructose, ethanol and glycerol levels were predicted from infrared spectra by commercial calibrations or in-house adjustments using the Winescan FT120 2001 version 2.2.1 software.
Aroma compounds
The liquid-liquid extraction was conducted as previously described (Louw et al., 2009), except for the modification where the wine/ether mixture was centrifuged at 4 000 rpm for 3 min. A spatula of Na2SO4 was added to the mixture to aid extraction in the synthetic wine matrix and the centrifugation was repeated. The organic layer was removed and dried. The volatile higher alcohols, esters and fatty acids were quantified in duplicate using a Hewlett Packard 6890 Plus gas chromatograph (Agilent, Little Falls, Wilmington, USA). The gas chromatograph was equipped with a split/ splitless injector, set to a split ratio of 15:1, a split flow rate of 98.7 mL/min and a temperature of 200°C. The separation of compounds was achieved using a J & W DB-FFAP capillary GC column (Agilent, Little Falls, Wilmington, USA) with dimensions of 60 m length χ 0.32 mm internal diameter, with a 0.5 μm coating film thickness and using a hydrogen carrier gas flow rate of 6.6 mL/min. Three microlitres of the extracted sample were injected.
The initial oven temperature was maintained at 33°C for 8 min. Temperature increases occurred at a rate of 21°C/min. The oven temperature was maintained at 130° C for 1.3 min, at 170°C for 1 min and then at 240°C for 2.5 min. The flame ionization detector was operated at 250°C, with a hydrogen flow of 30 mL/min, oxygen of 350 mL/min and make-up gas flow of nitrogen at 30 mL/min. The oven temperature was maintained at 240°C for 5 min between runs. After 22 injections, the column was cleaned both thermally and chemically by a hexane injection at an oven temperature of 250°C, with a holding time of 10 min per injection (Louw et al., 2010). Authentic reference standards (Merck, Cape Town) were used to calibrate for each of the compounds using the internal standard compound 4-methyl-2-pentanol. Data collection and peak integration were performed manually using the HP Chemstation software.
Statistical analysis
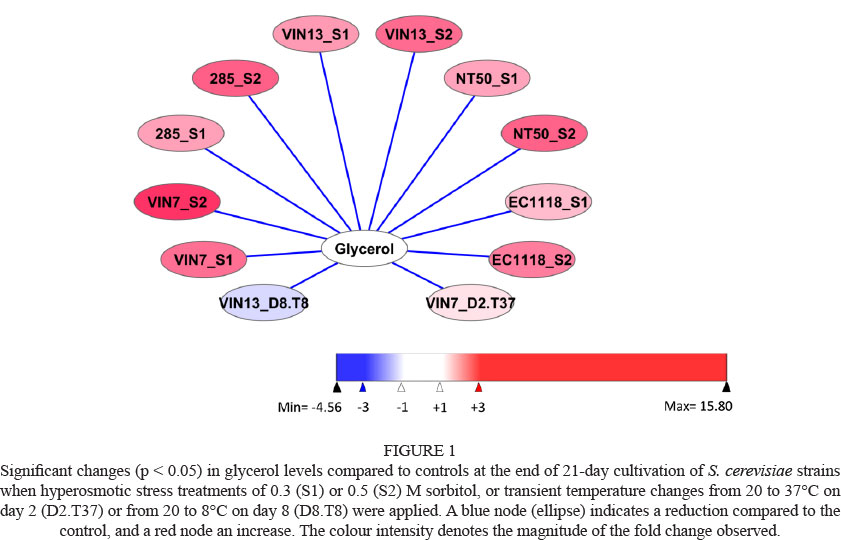
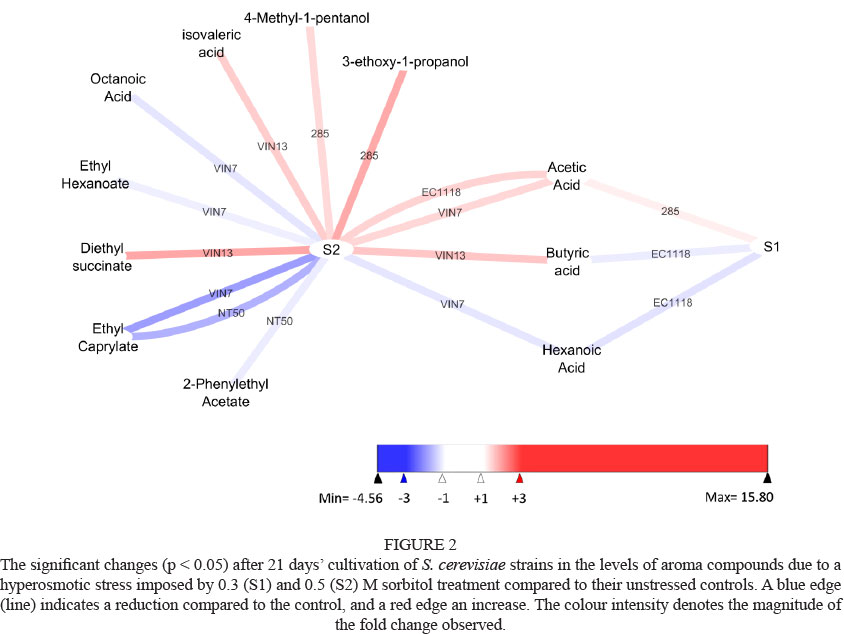
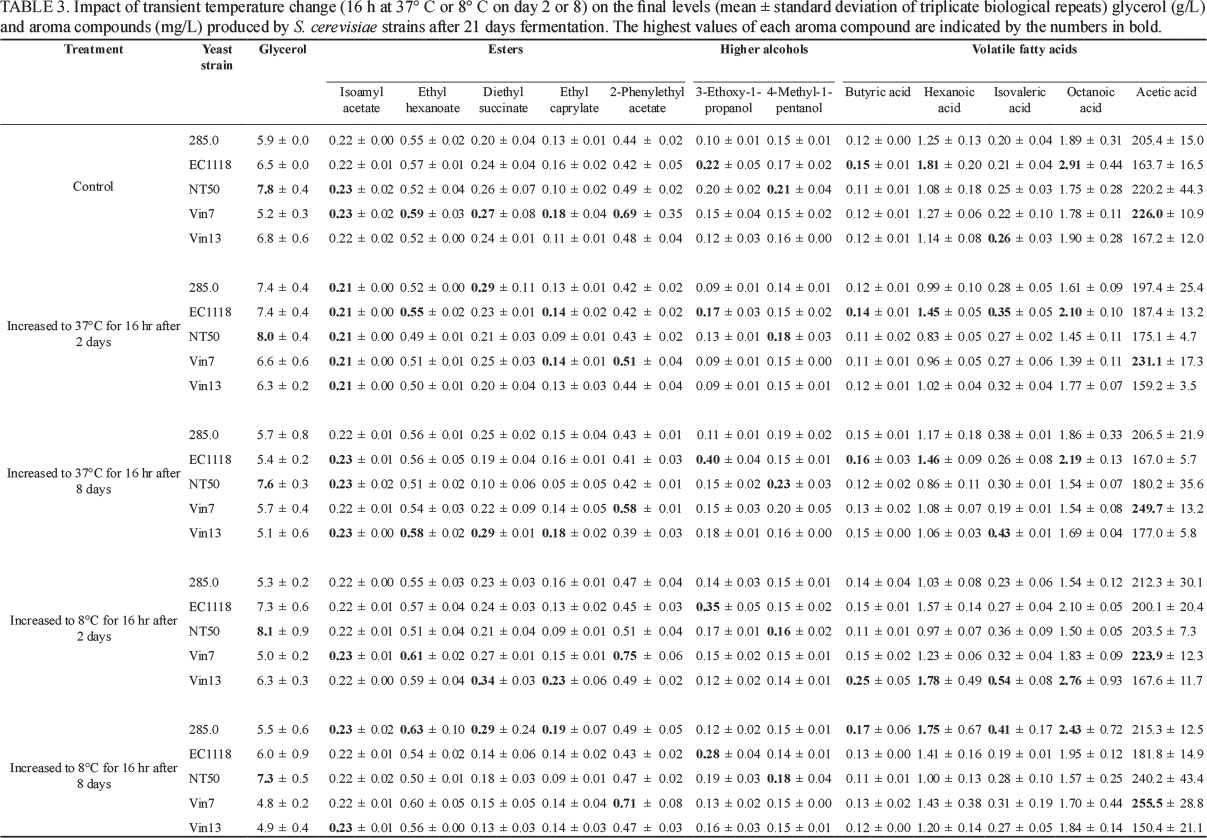
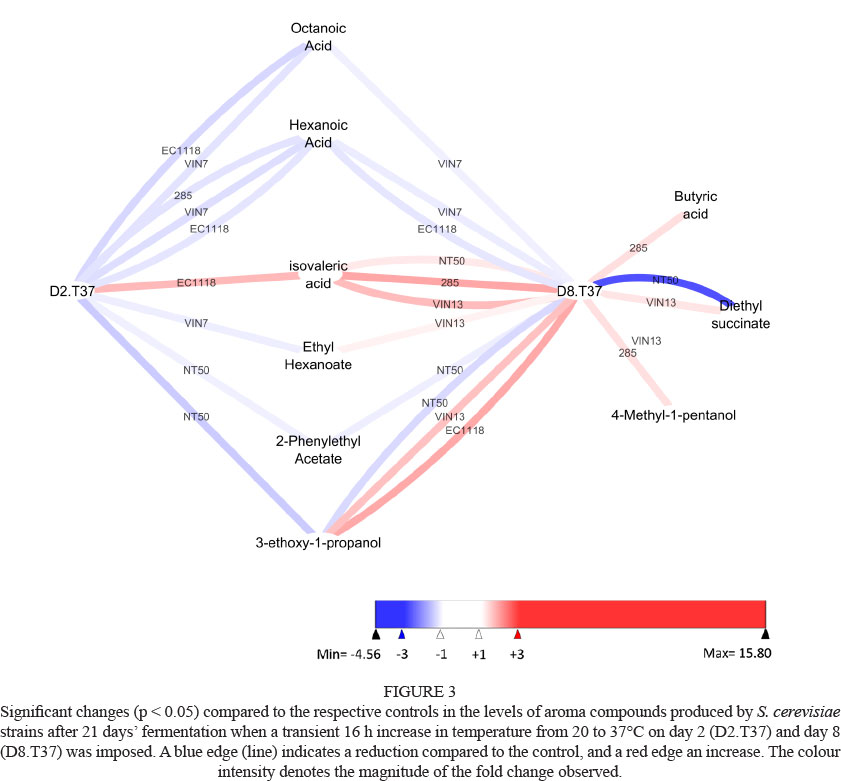
Fermentation data for stressed treatments were compared to that of the 'unstressed' control by creating pair-wise comparisons for each strain. These graphs were visualised using Cytoscape (version 2.8.2, http://www.cytoscape.org). These graphs only include information that differed significantly from the control, thus any omission was deemed to be statistically similar to its control. In all cases a significance level of 5% (p < 0.05) was used. A blue node (ellipse) or edge (line) indicates a statistically significant reduction compared to the control, whereas a red node or edge denotes an increase. The colour intensity of the node or edge is an indication of the magnitude of the fold change.
RESULTS AND DISCUSSION
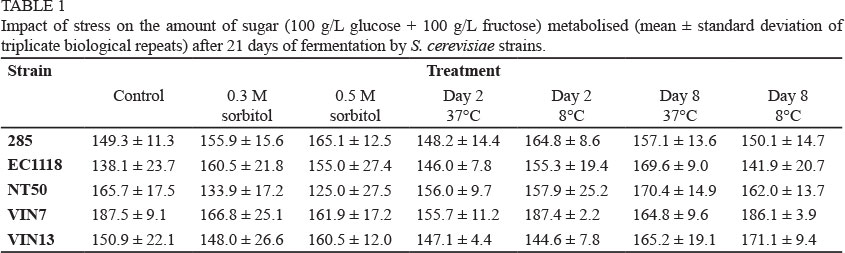
The hyperosmotic (0.3 and 0.5 M sorbitol) and transient temperature stresses (8°C and 37°C for 16 h on days 2 and 8) evaluated were relatively mild in order to mimic conditions that might be found during fermentations used in the wine industry. The synthetic grape must was not fermented to dryness (< 4 g/L residual sugar) within the 21 days from the commencement of the experiment. An imposition of a stress apparently had little or no impact on the sugar concentration fermented over 21 days (Table 1). Between 63% and 93% of the sugar was fermented, but there no consistent pattern in an effect of stress on sugar fermentation. Exposure of the five yeast strains to stress generally only resulted in small variations in ethanol concentrations when compared to control conditions (data not shown). The fermentation rates were only slightly affected by the application of temperature stress (data not shown). Hyperosmotic stress with 0.5 M sorbitol slightly reduced the fermentation rates of the NT50 and VIN7 strains, but surprisingly not that of the other strains (data not shown). VIN7 has previously been reported to ferment high-sugar grape musts poorly (Erasmus & Van Vuuren, 2009). The fermentation rates in the presence of 0.3 M sorbitol were similar to the controls of all strains.
Effect of hyperosmotic and temperature stress on glycerol and acetic acid production
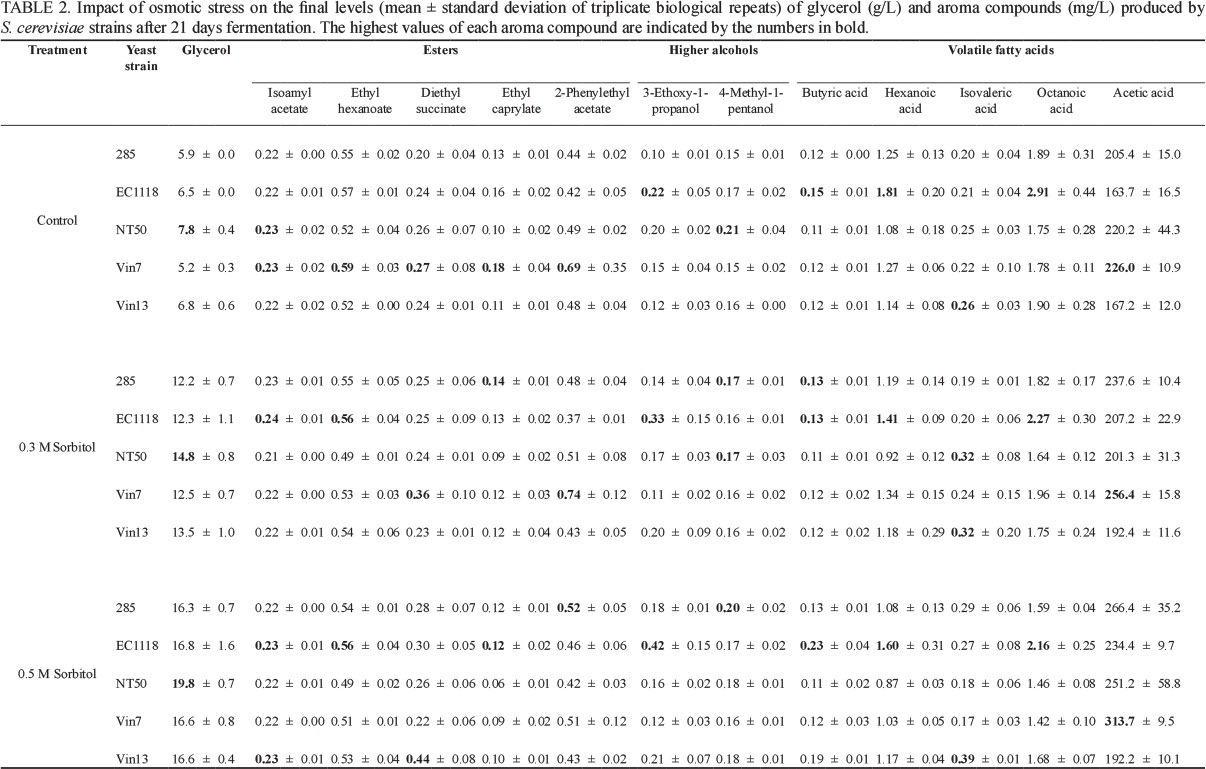
Hyperosmotic stress led to increased production of glycerol and acetic acid (Table 2). Compared to control conditions, significantly greater levels of glycerol were produced by all strains as the degree of hyperosmotic stress was increased (Fig. 1). Surprisingly, the levels of glycerol produced appeared to be much greater than the degree of hyperosmotic stress applied (Table 2), which might be related to the use of sorbitol to impose the hyperosmotic stress.
Additional glycerol is produced intracellularly to counter the hyperosmotic stress in terms of which the internal solute content of the yeast cells is increased and water uptake is promoted (Hohmann, 1997). Increased glycerol production is often accompanied by increased acetic acid production due to a redox imbalance created during glycerol production, which results in a surplus of NAD+ (Jain et al., 2012). This redox imbalance is corrected by the oxidation of acetaldehyde to acetic acid, with subsequent regeneration of NADH. Our results revealed that, in most instances, the acetic acid levels produced by the five strains increased together with the greater hyperosmotic stress imposed (Table 2), which is consistent with the creation of the redox imbalance. However, only strains 285 (0.3 M sorbitol), EC1118 (0.5 M sorbitol) and VIN7 (0.5 M sorbitol) displayed a statistically significant increase (Fig. 2). Under control and hyperosmotic stress conditions, strain NT50 was found to produce the highest levels of glycerol (Table 2), whereas strain 285 produced the lowest levels in most cases. On the other hand, VIN7 produced the highest acetic acid levels and continued to produce higher acetic acid concentrations when the yeast was placed under hyperosmotic stress (Table 2).
Temperature stress had little impact on the levels of glycerol and acetic acid produced (Table 3). A small but significant increase in glycerol was produced by VIN7 when a temperature stress of 37°C for 16 h was applied on day 2, whereas glycerol production by VIN13 was found to be significantly lower when the temperature was reduced to 8°C for 16 h on day 8 (Fig. 1). None of the small changes in acetic acid levels in response to temperature stress was statistically significant. As observed under hyperosmotic stress conditions, NT50 produced the highest glycerol levels, whereas the highest acetic acid levels were produced by VIN7 under all temperature stress conditions (Table 3). Other investigations of the impact of temperature stress on glycerol production reported an increase in levels by a commercial wine yeast (strain Uvaferm SLO) when temperature was rapidly increased from 18 to 34°C within 2 h (Kukec et al., 2003; Berovic et al., 2007). In a subsequent report, Berovic et al. (2007) reported that the longer the duration of the temperature shock (24 h compared to 4 h), the greater the amount of glycerol produced. However, the temperature stress was applied within the first 30 h of fermentation, whereas in our study the stress was only applied on day two or day eight of the fermentation, suggesting that the timing of stress application may play a role in glycerol production. When glycerol production by 13 wine yeast strains at 12° and 28°C was compared, no consistent effect of fermentation temperature on production could be established (Gamero et al., 2013). Other researchers have found the impact of reduced temperature fermentation on acetic acid levels to have variable results, pointing to an effect of strains and fermentation conditions. Molina et al. (2007) found minimal differences in the final acetic acid levels when strain EC1118 fermented a synthetic must at 15° and 28°C, whereas Beltran et al. (2008) reported a marked lower acetic acid level at 13°C compared to 25°C produced by their wine yeast (strain Lallemand QA23) in grape and synthetic musts.
Effect of hyperosmotic stress on production of aroma compounds
Hyperosmotic stress had either a small or no impact on the production of the 11 aroma compounds (acetic acid excluded) evaluated in this study (Table 2). Almost no impact of hyperosmotic stress was found in the levels of isoamyl acetate, ethyl hexanoate, diethyl succinate, 2-phenyl acetate, 3-ethoxy-1-propanol, 4-methyl-1-propanol, butyric acid and isovaleric acid produced by most strains (Table 2). However, statistical analysis revealed small but significant decreases in the respective levels of ethyl hexanoate and 2-phenyl acetate by VIN7 and NT50 (Fig. 2). Significant increases in levels of 3-ethoxy-1-propanol, 4-methyl-1-propanol were produced by 285 (Fig. 2). However, these significant changes were not found in other strains. Most strains showed small decreases of ethyl caprylate, hexanoic acid and octanoic acid when hyperosmotic stress was applied (Table 2). The decreases of ethyl caprylate (strains EC1118, VIN7 and NT50 for 0.5 M sorbitol), octanoic acid (VIN7, 0.5 M sorbitol) and hexanoic acid (VIN7, 0.5 M sorbitol and EC1118, 0.3 M sorbitol) were significant (Fig. 2). This suggests that the redox imbalance caused by glycerol production only influences the production of aroma compounds to a limited extent under the fermentation conditions evaluated. In the absence of hyperosmotic stress, strain VIN7 produced the highest levels of most aroma compounds (excluding acetic acid). However, when hyperosmotic stress was applied, strain EC1118 produced the highest levels of most aroma compounds (Table 2).
The redox balance in yeast was proposed to regulate the metabolic pathways responsible for the production of aroma compounds (Lambrechts & Pretorius, 2000). However, a recent study by Jain et al. (2012) found that the redox balance only seemed to affect some of the pathways responsible for aroma compound production. The deletion of glycerol-producing genes in a laboratory S. cerevisiae strain resulted in a significant increase in production of the higher alcohol isobutanol and a decrease in the ester ethyl acetate. The deletion strain would be expected to produce an excess of NADH, which in turn could promote the production of higher alcohols in an attempt to reduce NADH to NAD+. This redox imbalance would hinder the production of esters, which requires NAD+. However, the concentrations of the higher alcohols, isoamyl alcohol and 2-phenyl alcohol, were not affected by the deletion (Jain et al., 2012).
Effect of transient temperature increase on aroma compound production
The data showed that the influence of exposure to a transient temperature increase from 20° to 37°C for 16 h on the volatile aroma profile was time and strain dependent. A transient increase in temperature on either day 2 or day 8 resulted in a decrease in hexanoic acid and octanoic acid production and an increase in isovaleric acid production by most strains. Some of these aroma compound decreases or increases were significant (Fig. 3). For example, the decrease in hexanoic acid produced by strains 285, VIN7 and EC1118 on day 2 and strain VIN7 on day 8 was significant. In addition, octanoic acid produced by EC1118 and VIN7 on day 2 and VIN7 on day 8 decreased significantly (Fig. 3). The increase in isovaleric acid production by strain EC1118 on day 2 and strain VIN13 on day 8 was also significant.
Other significant increases or decreases in 4-methyl-1-pentanol, diethyl succinate, ethyl succinate and 3-ethoxy-1-propanol on day 2 or 8 were observed without a consistent pattern (Fig. 3). Similarly, Berovic et al. (2007) found that transient temperature increases from 18 to 34°C in S. cerevisiae resulted in one higher alcohol (isoamyl alcohol) concentration increasing and another (propanol) decreasing. The temperature increases on both days had no or little impact on the levels of isoamyl acetate, ethyl hexanoate, ethyl caprylate, 2-phenyl acetate and butyric acid produced by all strains (Table 3). Strain EC1118 produced the highest levels of eight of the 11 aroma compounds tested when the transient temperature was increased on day 2. However, on day 8, strains EC1118 and VIN13 both produced five of the highest levels of aroma compounds. Fermentation at higher temperature (28°C) is reported to yield greater concentrations of higher alcohols that give flowery aromas to wine (Molina et al., 2007). On the other hand, Gamero et al. (2013) found that the higher alcohol concentrations produced by 13 wine yeasts after fermentation at 28° compared to 12°C varied considerably between strains, pointing to an inconsistent response pattern to increased temperature.
Effect of transient decrease in temperature on aroma compound production
Similar to the temperature increase effect, the effect of exposure to a transient temperature decrease from 20°C to 8°C for 16 h on the volatile aroma profile was also time and strain dependent (Table 3). Most of the diethyl succinate levels produced on day 8 was lower than the control levels, although none were significant. Most strains produced higher levels of butyric and isovaleric acids when the temperature was decreased on day 2 and 8 (Table 3), but only the higher levels produced by VIN13 were statistically significant (Fig. 4). On the other hand, some strains produced lower levels of octanoic acid at the lower temperature compared to their respective controls, and the lower levels produced by EC1118 on day 2 and 8 were significant. The significant increases in 3-ethoxy-1-propanol and ethyl caprylate levels by respectively EC1118 and 285 (Fig. 4) were not found consistently in the other strains (Table 3). A transient decrease in temperature on both days had no or little impact on the levels of isoamyl acetate, ethyl hexanoate, 2-phenyl acetate, 4-methyl-1-pentanol and hexanoic acid produced by all strains (Table 3). Strain VIN13 produced the highest levels of six of the 11 aroma compounds tested when the transient temperature was decreased on day 2, but on day 8 eight of the highest levels were produced by strain 285 (Table 3). In contrast to our studies, for which a transient temperature decrease was applied, other studies have compared the impact of yeast fermentation temperature at 25 to 28°C and 12 to 15°C on the levels of volatile aroma compounds (Molina et al., 2007; Beltran et al., 2008; Gamero et al., 2013). The reduction in the yeast fermentation temperature had a major impact on the levels of volatile aroma compounds produced, and the levels produced varied between yeast strains. However, the differences in the levels of aroma compounds produced at the ambient and lower temperatures did not concur in the response to lower temperature stress observed in this study. For example, in contrast to observations that our strains produced lower levels of octanoic acid during decreased transient temperature stress (Table 3). Molina et al. (2007) and Beltran et al. (2008) found that their strains produced higher levels when the fermentation was conducted at 13 to 15°C compared to 25 to 28°C, respectively. Furthermore, Molina et al. (2007) reported that fermentation by strain EC1118 at 15°C resulted in higher levels of esters compared to fermentation at 28°C, whereas decreased temperature stress in this study had little impact. However, a comparison of levels of aroma compounds produced by 13 commercial wine yeast strains at fermentation temperatures of 12 and 28°C showed wide ranges (Gamero et al., 2013), pointing to a strain-specific response to reduced temperature.
CONCLUSIONS
This is the first study to investigate the influence of environmental stress on the volatile aroma profile in a synthetic grape must. The preliminary results demonstrate that the evaluated stresses caused a limited number of significant changes in the levels of 11 aroma compounds. Most changes occurred when the transient temperature was increased (25 changes of 110 measurements), whereas a temperature decrease resulted in nine significant changes, and hyperosmotic stress resulted in 16 changes of 110 measurements. The changes observed also differed for each of the strains, but changes could not be related to whether the strains were hybrids or S. cerevisiae strains. Hyperosmotic stress resulted in a greater production of glycerol and acetic acid, but the impact on volatile aroma compound production was inconsistent. This stress, together with the resultant glycerol production, would lead to a redox imbalance, but with an apparent limited impact on volatile aroma compound production. Additionally, significant changes may have occurred in the levels of compounds not detected by the method used in this study.
In conclusion, exposure to mild environmental stress caused a relatively small number of specific changes to individual compounds. These changes differed significantly for each strain and stress treatment, suggesting that the fermentation conditions can significantly alter the aromatic profile of a wine, although these impacts cannot be predicted in general. The changes to the aromatic profile appear to be specific to each individual wine yeast strain. Furthermore, as aroma compounds interact with each other, they may cause significant changes in the overall aroma profile of the resultant wine. Further analysis on the impact of environmental stress during grape must fermentation, and the duration of the stress application, would provide information regarding the extent to which environmental stress affects the aroma profile of wine.
LITERATURE CITED
Alexandre, H. & Charpentier, C., 1998. Biochemical aspects of stuck and sluggish fermentation in grape must. J. Ind. Microbiol. Biotechnol. 20, 20-27. [ Links ]
Attfield, P. V., 1997. Stress tolerance: The key to effective strains of industrial baker's yeast. Nat. Biotechnol. 15, 1351-1357. [ Links ]
Bauer, F.F. & Pretorius, I.S., 2000. Yeast stress response and fermentation efficiency: How to survive the making of wine - A review. S. Afr. J. Enol. Vitic. 21, 27-51. [ Links ]
Beales, N., 2004. Adaptation of microorganisms to cold temperatures, weak acid preservatives, low pH, and hyperosmotic stress: A review. Compr. Rev. Food Sci. Food Safety 3, 1-20. [ Links ]
Bell, S.J. & Henschke, P.A., 2005. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 11, 242-295. [ Links ]
Beltran, G., Novo, M., Guillamón, J.M., Mas, A. & Rozès, N., 2008. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 121, 169-177. [ Links ]
Bely, M., Sablayrolles, J.M. & Barre, P., 1990. Automatic detection of assimilable nitrogen deficiencies during alcoholic fermentation in oenological conditions. J. Ferment. Bioeng. 70, 246-252. [ Links ]
Berovic, M., Pivec, A., Kosmerl, T., Wondra, M. & Celan, S., 2007. Influence of heat shock on glycerol production in alcohol fermentation. J. Biosci. Bioeng. 103, 135-139. [ Links ]
Bisson, L.F., 1999. Stuck and sluggish fermentations. Amer. J. Enol. Vitic. 50, 107-119. [ Links ]
Bisson, L.F. & Karpel, J.E., 2010. Genetics of yeast impacting wine quality. Food Sci. Technol. 1, 139-162. [ Links ]
Carrasco, P., Querol, A. & Del Olmo, M., 2001. Analysis of the stress resistance of commercial wine yeast strains. Arch. Microbiol. 175, 450-457. [ Links ]
Erasmus, D.J. & Van Vuuren, H.J.J., 2009. Genetic basis for osmosensitivity and genetic instability of the wine yeast Saccharomyces cerevisiae VIN7. Amer. J. Enol. Vitic. 60, 145-154. [ Links ]
Francis. I. & Newton, J., 2005. Determining wine aroma from compositional data. Aust. J. Grape Wine Res. 11, 114-126. [ Links ]
Gamero, A., Tronchoni, J., Querol, A. & Belloch, C., 2013. Production of aroma compounds by cryotolerant Saccharomyces species and hybrids at low and moderate fermentation temperatures. J. Appl. Microbiol. 114, 1405-1414. [ Links ]
Gibson, B.R., Lawrence, S.J., Leclaire, J.P.R., Powell, C.D. & Smart, K.A., 2007. Yeast responses to stresses associated with industrial brewery handling. FEMS Microbiol. Rev. 31, 535-569. [ Links ]
Henschke, P. & Jiranek, V., 1993. Yeasts - metabolism of nitrogen compounds. In: Fleet, G.H. (ed). Wine Microbiology and Biotechnology. Harwood Academic Publishers, Switzerland. pp. 77 - 164. [ Links ]
Hohmann, S., 1997. Shaping up: The response of yeast to hyperosmotic stress. In: Hohmann. S. & Mager, W.H. (eds). Yeast stress responses. Springer, New York. pp. 101 - 145. [ Links ]
Ivorra, C., Pérez Ortín, J.E. & Del Olmo, M., 1999. An inverse correlation between stress resistance and stuck fermentations in wine yeasts. A molecular study. Biotechnol. Bioeng. 64, 698-708. [ Links ]
Jain, V.K., Divol, B., Prior, B.A. & Bauer, F.F., 2012. Effect of alternative NAD+-regenerating pathways on the formation of primary and secondary aroma compounds in a Saccharomyces cerevisiae glycerol-defective mutant. Appl. Microbiol. Biotechnol. 93, 131-141. [ Links ]
Kukec, A., Berovic, M., Wondra, M., Celan, S. & Kosmerl, T., 2003. Influence of temperature shock on the glycerol production in cv. Sauvignon blanc fermentation. Vitis 42, 205-206. [ Links ]
Lambrechts, M. & Pretorius, I.S., 2000. Yeast and its importance to wine aroma - A review. S. Afr. J. Enol. Vitic. 21, 97-129. [ Links ]
Louw, L., Roux, K., Tredoux, A., Tomic, O., Naes, T, Nieuwoudt, H.H. & Van Rensburg, P. 2009. Characterization of selected South African young cultivar wines using FTMIR spectroscopy, gas chromatography, and multivariate data analysis. J. Agric. Food Chem. 57, 2623-2632. [ Links ]
Louw, L., Tredoux, A., Van Rensburg, P., Kidd, M., Naes, T. & Nieuwoudt, H.H., 2010. Fermentation-derived aroma compounds in varietal young wines from South Africa. S. Afr. J. Enol. Vitic. 31, 213-225. [ Links ]
Mager, W.H. & Siderius, M., 2002. Novel insights into the hyperosmotic stress response of yeast. FEMS Yeast Res. 2, 251-257. [ Links ]
Malherbe, S., Bauer, F.F. & Du Toit, M., 2007. Understanding problem fermentations - A review. S. Afr. J. Enol. Vitic. 28, 169-186. [ Links ]
Molina, A.M., Swiegers, J.H., Varela, C., Pretorius, I.S. & Agosin, E., 2007. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 77, 675-687. [ Links ]
Rapp, A. & Mandery, H., 1986. Wine aroma. Experientia 42, 873-884. [ Links ]
Rapp, A. & Versini, G. 1996. Influence of nitrogen compounds in grapes on aroma compounds of wines. Wein-Wiss. 51, 1659-1694. [ Links ]
Rossouw, D., Nas, T. & Bauer, F.F., 2008. Linking gene regulation and the exo-metabolome: A comparative transcriptomics approach to identify genes that impact on the production of volatile aroma compounds in yeast. BMC Genomics 9, 530-547. [ Links ]
Saerens, S.M.G., Delvaux, F., Verstrepen, K.J., Van Dijck P., Thevelein J.M & Delvaux, F.R., 2008. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 74, 454-461. [ Links ]
Soles, R., Ough, C. & Kunkee, R., 1982. Ester concentration differences in wine fermented by various species and strains of yeasts. Amer. J. Enol. Vitic. 33, 94-98. [ Links ]
Styger, G., Prior, B.A. & Bauer, F.F., 2011. Wine flavor and aroma: A review. J. Ind. Microbiol. Biotechnol. 38, 1145-1159. [ Links ]
Swiegers, J., Bartowsky, E.J., Henschke, P. & Pretorius, I.S., 2005. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 11, 139-173. [ Links ]
Tamás, M.J. & Hohmann, S., 2003. The hyperosmotic stress response of Saccharomyces cerevisiae. In: Hohmann, S. & Mager, P.W.H. (eds). Yeast stress responses. Springer-Verlag, Berlin. pp. 121 - 200. [ Links ]
Zuzuarregui, A. & Del Olmo, M., 2004. Analyses of stress resistance under laboratory conditions constitute a suitable criterion for wine yeast selection. Antonie van Leeuwenhoek 85, 271-280. [ Links ]
Submitted for publication: July 2013
Accepted for publication: March 2014
Acknowledgements: We thank Candice Stilwaney for laboratory assistance. The National Research Foundation and Anchor Yeast are thanked for financial support
* Corresponding author: Florian F. Bauer: fb2@sun.ac.za [Tel.: +27 21 808 4346; Fax: +27 21 808 3771]
Other authors: Samantha C. Fairbairn: fairbairn@sun.ac.za, Anita Y. Smit: aysmit@distel.co.za, Dan Jacobson: jacobson@sun.ac.za, Bernard A. Prior: bap@sun.ac.za

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}