










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
versão On-line ISSN 2224-7904
versão impressa ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.35 no.2 Stellenbosch 2014
Suitability of different PCR-DGGE primer sets for the monitoring of lactic acid bacteria in wine
S. Mayrhofer*; R. Filipp; D. Lehner; C. Reiterich; W. Kneifel; K.J. Domig
BOKU - University of Natural Resources and Life Sciences Vienna, Department of Food Science and Technology, Institute of Food Science, Muthgasse 18, A-1190 Vienna, Austria
ABSTRACT
Lactic acid bacteria (LAB) play a dual role in winemaking as they are the main effectors of malolactic fermentation, but some members can also cause wine spoilage. PCR-DGGE has proved to be a quick tool to study the LAB community and their fluctuation in wine. For detecting wine-associated LAB by PCR-DGGE, the primer sets WLAB1/WLAB2GC, WBAC1/WBAC2GC, Lac1/Lac1o/Lac2GC, 341fgC/518r and rpoBl/ rpoB1o/rpoB2GCwere tested and evaluated in this study. The primer systems were assessed by the separation of LAB reference strains on DGGE gels and by attributing the resulting amplicons to defined species. Subsequently, the detection of LAB in wine samples and enrichments thereof was compared. While the primer systems WBAC1/WBAC2GCand 341fGC/518r were not appropriate, the Lac1/Lac1o/Lac2GCprimer set performed well. However, multiple bands complicated the evaluation. The rpoB1/rpoB1o/rpoB2GCset seemed to be promising for the detection of LAB in wine, although further improvements in terms of the detection limit need to be done. Due to the pronounced sensitivity and the sufficient discrimination of LAB at species level, the WLAB1/WLAB2GCprimer system was found to be most suitable for studying the occurrence of LAB in wine.
Key words: Malolactic fermentation, lactic acid bacteria, wine, PCR-DGGE, primer-set
INTRODUCTION
Winemaking is a complex microbial process inwhich primarily yeasts, but also lactic acid bacteria (LAB), play pivotal roles (Lonvaud-Funel, 1999). Malolactic fermentation (MLF) can occur at the end of the alcoholic fermentation conducted by yeasts (Lonvaud-Funel, 1999). This fermentation is usually desirable in most of the red wines, some white cultivars, including Chardonnay, some sparkling wines and also in cool-climate Riesling wines (Lerm et al., 2010; Knoll et al., 2012). MLF is the bacterial conversion of L-malic acid to L-lactic acid and CO2 (Bousbouras & Kunkee, 1971) and exerts significant influences on wine acidity, flavour and microbiological stability (Moreno-Arribas & Polo, 2005). The main effectors of MLF are LAB. Of these, Oenococcus oeni is the species mainly responsible for MLF (Davis et al., 1986), as it is the species that has accommodated the best to the difficult fermentation conditions, such as low pH values and high ethanol concentrations (Wibowo et al., 1988). Due to its particular role, this species is commonly used as starter culture to promote MLF (Mills et al., 2005). Furthermore, it is not very frequently reported to be associated with off flavours like volatile acidity and mousiness, spoilage like ropiness, or the formation of undesirable metabolites such as ethyl carbamate and biogenic amines, which can be caused by other wine-related LAB (Mills et al., 2005).
Owing to these observations there is a need to control MLF to enhance the positive attributes or to reduce potential negative impacts on the particular wine (Mills et al., 2005). Traditional culture-based techniques are often used to detect LAB in wine samples (Cho et al., 2011) but, especially in case of the main effector O. oeni, up to 14 days are required to yield results. Such long cultivation periods, however, do not allow the carrying out of possible oenological prevention or operation in wine production (Pinzani et al., 2004). Thus, several culture-independent methods (e.g. PCR-DGGE, qPCR) have been developed because they overcome the problems described above.
PCR-denaturing gradient gel electrophoresis (DGGE) is a commonly used culture-independent fingerprinting technique for the rapid analysis of microbial communities and has been used to analyse LAB in food (Cocolin et al., 2001). This technique is applied to separate a mixture of PCR amplicons of the same size but of different sequences (Ercolini, 2004). Double-stranded PCR amplicons in the gel are subjected to an increasingly denaturing environment. The migration is stopped when the DNA fragments are completely denatured (Renouf et al., 2007), yielding patterns that visualise the genetic diversity of the investigated microbial community (Ercolini, 2004).
Implementing a new method in the own laboratory always requires intensive literature research. In terms of PCR-DGGE applied to study wine LAB microbiota, several different primer sets and PCR conditions have already been introduced by researchers. Of these, five primer sets were shortlisted, as they seemed to be appropriate (Lopez et al., 2003; Rantsiou et al., 2004; Endo & Okada, 2005; Bae et al., 2006; Renouf et al., 2006a; Spano et al., 2007). The objective of this study was to partly modify and extensively test and evaluate these primer systems regarding their suitability to monitor LAB in wine. The results of this study can be consulted to investigate the presence of LAB in wine by PCR-DGGE.
MATERIALS AND METHODS
Strains and growth conditions
Bacterial reference strains and their corresponding growth conditions (medium and temperature) used in this study are listed in Table 1. The strains were grown anaerobically (85% N2, 10% CO2, 5% H2) using a MACS VA 500 micro-aerophilic workstation (Don Whitley Scientific, Shipley, U.K.).
In addition, further LAB (Enterococcus faecalis LMG 7937T, Enterococcus faecium LMG 11423T; Streptococcus thermophilus LMG 6897T and Tetragenococcus muriaticus LMG 18498T) were included in the tests in order to evaluate the specificity of primer Laclo. Except for Tetragenococcus muriaticus, all strains were grown on MRS medium (deMan, Rogosa, Sharpe; Merck, Darmstadt, Germany): Streptococcus thermophilus anaerobically at 37°C and Enterococcus faecalis as well as Enterococcus faecium aerobically at 37°C. Tetragenococcus muriaticus was cultivated on GYP sodium acetate mineral salts medium with 5% sodium chloride (BCCM/LMG Bacteria Collection, Ghent University, Ghent, Belgium; medium 244) under aerobic conditions at 30°C.
Isolates of the accompanying bacterial flora (Bacillus coagulans MSB 29W, Gluconobacter japonicus MSB 32W, Gluconobacter oxydans MSB 107W and Acetobacter aceti MSB 109W) grown anaerobically on MLO medium (medium for Leuconostoc oeni; German Collection of Microorganisms and Cell Cultures (DSMZ); medium 59) at 30°C were also included.
Wine samples and corresponding microbial enrichment cultures
In addition to the reference strains described above, wine samples containing an unknown variety of microorganisms were investigated. However, as the detection limit of PCR-DGGE is around 104 cells/mL or even higher (Andorra et al., 2008), enrichment cultures of wine samples were also used for this investigation. For this purpose, 1 mL of wine sample was inoculated in 9 mL MRS and MLO medium and incubated for seven days at 30°C under anaerobic conditions to enhance naturally occurring wine LAB.
Initially, the DNA of 16 wine samples was isolated for the investigation. As the first PCR-DGGE results of these samples displayed no or only faint bands, their enrichments were used mainly to compare the performance of the different primer systems.
DNA extraction
DNA extraction from 2 mL of pure or enriched cultures was performed with the Archive Pure DNA Yeast & Gram+ Kit (5 Prime, Hamburg, Germany), according to the manufacturer's instructions.
The DNA from the wine sample was extracted directly using a protocol described by Renouf et al. (2009), with minor modifications. In brief, microbial cells were collected from 50 mL of wine by centrifugation (4 500 x g, 15 min, 4°C) and the pellet was washed in 600 μL TE buffer (10 mM Tris, 1 mM EDTA). After the next centrifugation step (10 000 x g, 7 min, 4°C), the supernatant was discarded and the pellet resuspended in 300 μL TE buffer. Furthermore, 300 μL of sterile glass beads were added and the samples were vortexed for 10 min at 4°C. The supernatant was mixed with 300 μL cell lysis solution (5 Prime). Subsequently, 200 μL of protein precipitation solution (5 Prime) were added and mixed. Precipitation of cellular fragments was done on ice for 5 min, followed by a centrifugation step at 10 000 x g for 3 min at 4°C. The supernatant was then transferred to a new 1.5 mL micro-centrifuge tube already containing 100 μL of 10% polyvinyl-pyrrolidone solution (PVP, Sigma-Aldrich, St. Louis, Missouri, USA) for the elimination of tannins. After vortexing and further centrifugation (10 000 x g, 10 min, 4°C), the supernatant was once more transferred to a new micro-centrifuge tube containing 300 μL of isopropanol. The tube was gently mixed by inversion and centrifuged at 10 000 x g for 3 min at 4°C. Subsequently, the supernatant was discarded, and 300 μL of 70% ethanol were added to the pellet and mixed by inversion. A final centrifugation step (10 000 x g, 3 min, 4°C) followed, and the ethanol was removed carefully. The tube was dried for 15 min. To rehydrate the DNA, 25 μL TE buffer and 0.5 μL RNase (4 mg/mL) were added to the sample overnight at 4°C. The DNA of the wine samples was stored at -20°C until use.
DNA amplification and primers
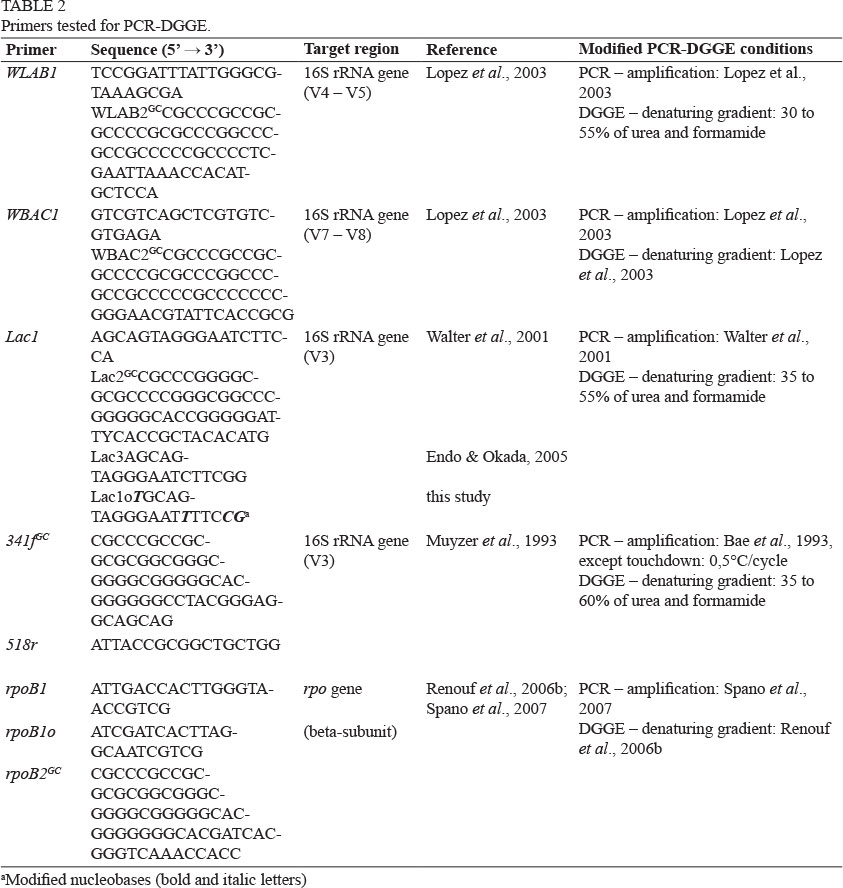
The primer systems WLAB1/WLAB2GC (Lopez et al., 2003), WBAC1/WBAC2GC(Lopez et al., 2003), Lac1/Lac2GC/Lac3 (Walter et al., 2001; Endo & Okada; 2005), Lac1/Lac1o/Lac2GC(Walter et al., 2001; this study) and 341fGC/518r (Bae et al., 2006; Muyzer et al., 1993) were used for the amplification of fragments of the bacterial 16S ribosomal RNA (rRNA) gene (Table 2). In addition, the primer system rpoB1/rpoB1o/rpoB2GCwas applied for the duplication of fragments of the RNA polymerase beta subunit, rpoB (Renouf et al., 2006b; Spano et al., 2007).
PCR amplification was performed at a final volume of 25 μL with a thermocycler (Mastercycler, Eppendorf, Hamburg, Germany) containing a combination of the corresponding primers and template DNA, as indicated by the authors (Muyzer et al., 1993; Walter et al., 2001; Lopez et al., 2003; Spano et al., 2007), and 2.5 μL 10 x PCR-buffer (Finnzymes, Vantaa, Finland), 0.5 μL dNTP-Mix (10 mM), and 0.5 μL DNA polymerase (2U/μL, Dynazyme II; Finnzymes). The remaining volume was filled up with sterile distilled water.
Amplicons were run on 2% agarose gels, stained with ethidium bromide and photographed under UV trans-illumination.
DGGE
The Dcode universal mutation detection systemTM (Bio-Rad, Hercules, California, USA) was used for the sequence-specific separation of PCR products. These were run on 8% (w/v) polyacrylamide gels in TAE-buffer (40 mM Tris-acetate; 2 mM Na2-EDTA x H2O, pH 8.5) and a denaturing gradient as described originally or modified according to Table 2. The electrophoresis was performed at 85 V for 16 h in 1 x TAE-buffer at a constant temperature of 60°C.
Band-matching analysis
Using the BioNumerics software, version 6.6 (Applied Maths, Sint-Martens-Latem, Belgium), wine-associated LAB species were identified by performing band matching. Accordingly, bands were automatically assigned to band classes defined by the program. Uncertain bands were ignored. The assignments were corrected manually, leading to an optimisation of 0% (WBAC, Lac, rpoB primer set) or 0.5% (WLAB, 341fGC/518r primer set) and a position tolerance of 0.5% (WLAB primer set) or 1% (WBAC, Lac, 341fGC/518r, rpoB primer set).
Sequence analysis
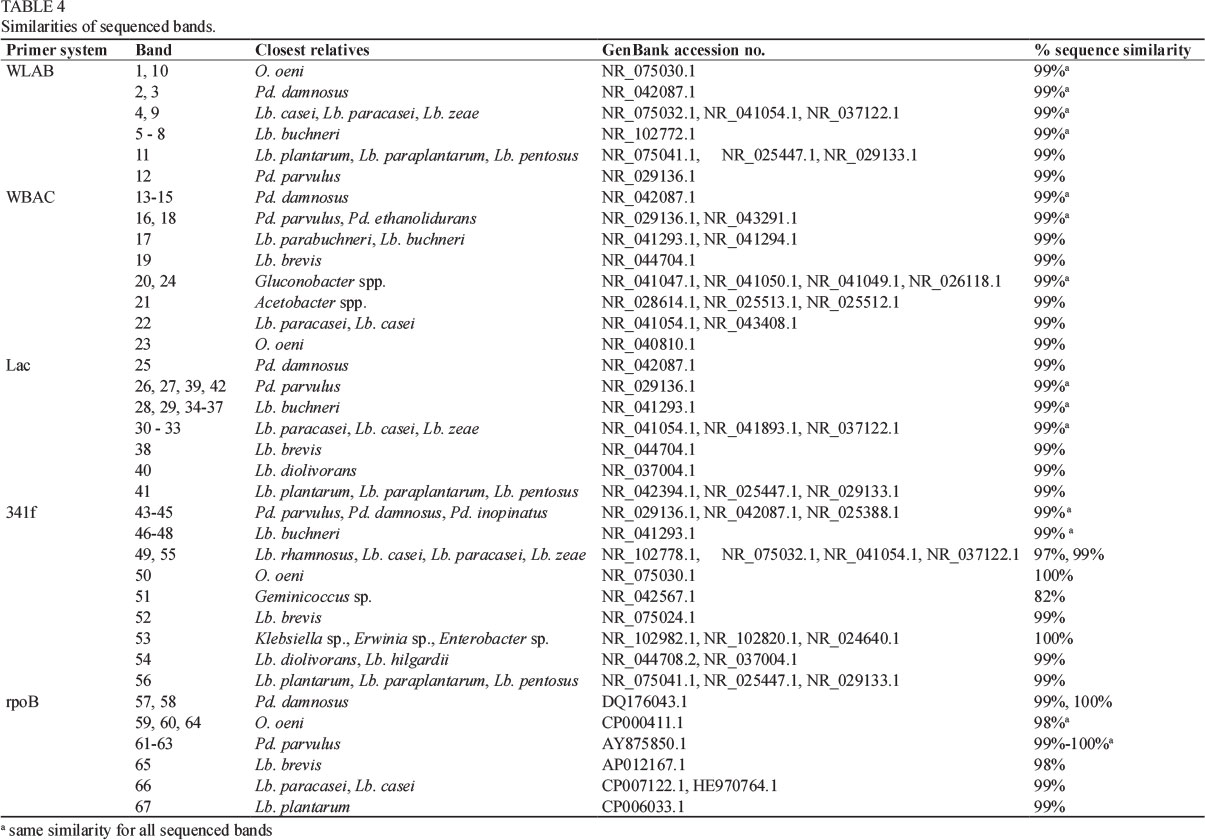
After staining the DGGE gel, bands of interest were excised directly from the gels with a scalpel, mixed with 100 μL of1x PCR buffer, and incubated overnight at 4°C. Two microlitres of this solution were used to re-amplify the PCR product. The PCR products were purified with the PCRExtract Mini Kit (5 Prime) and subjected to commercial sequencing (Eurofins MWG Operon, Ebersberg, Germany). Sequence compilation and comparison were performed with the BLASTn program.
RESULTS AND DISCUSSION
PCR-DGGE is a fast method for bacterial analysis, enabling the survey of LAB during winemaking (Renouf et al., 2006b). Therefore, a selection of primer systems proposed in the literature were checked against each other by testing a set of reference strains as well as wine samples and their enrichments.
Lopez et al. (2003) have already shown that a number of primers are not suited, as they also amplify nonbacterial DNA, resulting in a masking of bacterial populations in DGGE profiles. They therefore developed two new primer sets specifically for the amplification of bacterial 16S rRNA gene in wine fermentation samples. One primer set, termed WLAB1/WLAB2GC, amplifies LAB, while the other one, termed WBAC1/WBAC2GC, amplifies LAB and acetic acid bacteria (AAB). The primer set WLAB1/WLAB2GCtargets the V4 and V5 regions of the 16S rRNA gene and produces a fragment of approximately 400 bp (Lopez et al., 2003). Pure reference cultures were examined using this primer system. It was found that several LAB species exhibited similar electrophoretic mobilities, because all amplicons were only displayed in a small range of the denaturant concentration. Varying the concentration of denaturant of the electrophoresis gel did not improve the separation of the tested reference strains. Due to several copies of the targeted gene, some species even resulted in multiple bands, complicating the allocation of bands to certain LAB species. However, all of the tested LAB could be detected and differentiated, except for Lb. casei and Lb. paracasei (Fig. 1).
Primer set WBAC1/WBAC2GCtargets the V7 to V8 regions of the 16S rRNA gene and produced an approximately 320 bp amplicon with all tested reference strains. With reference to Lopez et al. (2003), this primer system works particularly well to resolve AAB strains on DGGE. Anyhow, according to our results, this primer pair was not capable for analysing the LAB diversity in wine, as the separation of the tested reference strains was not sufficient and many species migrated to the same position (Fig. 1). Nevertheless, it was possible to discriminate between Lb. casei and Lb. paracasei. Compared to the WLAB primer system, more multiple and stronger bands were obtained with the reference strains, except for Lb. buchneri and Lb. fructiovorans, which resulted in weak bands. Along with all the LAB reference strains, the Bacillus sp. isolate was also detected. This isolate, as well as the AAB, produced amplicons at the same gel positions as LAB.
The primer pair Lac1/Lac2GCwas designed for analysing the diversity of faecal or vaginal LAB and is specific for the genera Lactobacillus, Pediococcus, Weissella and Leuconostoc. The primer pair forms a 340 bp fragment of the V3 region of the 16S rRNA gene (Walter et al., 2001). An additional primer was constructed by Endo and Okada (2005) to extend the range of detectable LAB for the investigation of fermented foods. This Lac3 primer attaches at the same position as Lac1 and amplifies the 16S rRNA gene of Lactococcus spp., Streptococcus spp., Enterococcus spp., Vagococcus spp. and Tetragenococcus spp. Testing different primer combinations, Endo and Okada (2005) observed that the use of all three primers in a PCR at the same time was useful to analyse LAB diversity. Applying the primer mixture Lac1/Lac2GC/Lac3, the most relevant wine LAB, O. oeni, was not amplified (data not shown). To overcome this problem, the primer Lac1 or Lac3 was modified in this work (Lac1o, Table 2). The specificity of the new primer set Lac1/Lac1o/Lac2GCwas analysed using BLASTn and evaluated by performing PCR-DGGE with reference strains and isolates of the unwanted, accompanying bacterial wine micro-flora (for details see Materials and Methods). DGGE bands were obtained for all strains of the genera Lactobacillus, Pediococcus, Leuconostoc, Weissella and O. oeni. In contrast, no bands were achieved for the genera Enterococcus, Streptococcus, Tetragenococcus (data not shown) and Lactococcus, for which the Lac3 primer was designed by Endo and Okada (2005). However, these genera generally are not relevant for the fermentation processes of wines. Due to the generation of multiple bands for many reference strains, the evaluation of the results was difficult. Except for Lb. paracasein the identification of species of the Lactobacillus casei and Lb. plantarum group, as well as the Pediococcus genus, was often only possible at genus or species group level (Fig. 1), although a good separation was achieved for all other reference strains. No bands on DGGE gel were obtained for the non-LAB bacteria Bacillus coagulans, Acetobacter aceti and Gluconobacter spp. (Fig. 1)
The universal primer set 341fGC/518r, designed by Muyzer et al. (1993), was applied successfully by Bae et al. (2006) to detect LAB associated with wine grapes. It amplifies a fragment of the V3 region of the 16S rRNA gene, forming a 233 bp product. The PCR product from the Lb. fructivorans reference strain was weak when using this primer set without GC-clamp, and resulted in no visible band on the DGGE gel. However, the distribution of the bands of all other LAB reference strains was good, although multiple bands per strain appeared (Fig. 1). In addition, different LAB of one genus or species group (e.g. the Pd. damnosus, Pd. parvulus, Pd. inopinatus, Lactobacillus casei and Lb. plantarum group) showed identical results on the DGGE gel. As this primer set is universal, faint bands of AAB and the Bacillus isolate were displayed, but not in the concentration range of LAB.
As ribosomal genes are present in several copies with different sequences (Rantsiou et al., 2004), all primer systems considered generated diverse amplicons, resulting in multiple bands on the gel. Thus, another primer set targeting the RNA polymerase beta subunit gene rpoB, which is only present as a single copy (Rantsiou et al., 2004), was also included in the tests. This primer set, originally developed by Renouf et al. (2006a), had already been used to study the effect of different oenological practices on LAB populations and their evolution during winemaking. Based on the rpoB1/rpoB1o/rpoB2GCprimer system, the bands of the reference strains were well separated within this study (Fig. 2). However, the optimally expected single band per strain appeared as a main band with weak "double bands" in its neighbourhood (Renouf et al., 2006a). Amongst others, these bands may be due to an enzymatic process involving the TAQ polymerase (Janse et al., 2004). Nevertheless, the main bands were clearly separated and visible. In the case of wine samples, the unambiguous detection of main bands could even be improved with a mixture of various species. Otherwise, reference strains of some LAB species already found in wine could not be detected beside the accompanying bacterial flora (e.g. Lc. lactis, Lb. brevis, Lb. casei, Lb. coryniformis, Lb. curvatus, Lb. delbrueckii, Lb. fermentum, Lb. fructivorans, Lb. hilgardii, Lb. lindneri, Lb. nageli, Lb. zeae and Pd. inopinatus), or generated only faint bands (e.g. Lb. paracasei and Pd. parvulus). Although Renouf et al. (2006a) could determine the wine-relevant species Lb. brevis and Lb. hilgardii by PCR-DGGE, we could not produce amplicons for the used reference strains of these two species in our study when applying this primer system. However, Lb. brevis could be identified in one of the subsequently tested samples.

As PCR-DGGE patterns obtained with the reference strains should allow the tentative identification of DNA fragments in each sample, wine-associated LAB species were identified by matching their band distances to those of the reference strains using the BioNumerics software. O. oeni, Lb. brevis, members of the Pediococcus genus, the Lb. buchneri, Lb. casei and Lb. plantarum group were detectable when investigating the samples using each selected primer system (Table 3). In addition to these LAB, weak bands corresponding to the species Le. mesenteroides were displayed by the WLAB set. This species, as well as the species detected by all primer sets, belong to the main LAB isolated from must and wines (Pozo-Bayón et al., 2009). Compared to the other primer systems used in our experiment, O. oeni was rarely detected by the 341fGC/518r primer set, whereas this species was frequently identified by the Lac and rpoB primer systems with specific primers for O. oeni. Furthermore, species of the Lb. buchneri group and the Pediococcus genus were rarely found by the WBAC primer set.
The detection of the diversity of species by various primer sets may be influenced by their differing affinity to different species (Bae et al., 2006). According to our observations, the primer pair used itself also affects the detection limit of PCR-DGGE. Thus, O. oeni could not or hardly be identified in sample 23 by the rpoB and 341fGC/518r primer sets respectively, whereas its presence was clearly detected by all other primer systems (Table 3). This detection limit even increases when competitive template DNAs are present (Andorra et al., 2008). Furthermore, Bae et al. (2006), as well as Renouf et al. (2006b), concluded independently that their applied primer sets were only able to reveal the predominant species.
When applying the primer sets described above to investigate 27 wine samples and enrichment cultures, the same species were mostly detected by the Lac1/Lac1o/Lac2GC and WLAB1/WLAB2GC primer sets, followed by the rpoB1/rpoB1o/rpoB2GCprimer systems. The biggest diversity of LAB species was also verified by these primer sets. The WBAC1/WBAC2GCand 341fGC/518r primer systems showed the poorest compliance.
Based on the sequence analysis performed for selected bands (Table 4), it turned out that the similarity of the sequences of LAB bands generated by primers targeting the 16S rRNA gene (WLAB, WBAC, Lac and 341fGC/518r systems) with those available in the database was > 97%, whereas all tested rpoB sequences corresponded to database sequences with a similarity of > 98%. Furthermore, sequences generated by the rpoB primer set were more discriminative for the identification of related LAB species than those produced by 16S rRNA gene primer systems, which is in accordance with the literature (Renouf et al., 2006a; Lv et al., 2012).
CONCLUSIONS
Due to poor compliance with the other primer sets, the WBAC1/WBAC2GCand 341fGC/518r primer systems are not suitable to investigate the diversity of LAB involved in winemaking. In addition, multiple bands were frequently produced for the reference strains tested, complicating the allocation of a particular band to a defined species. Compared to these primer systems, the Lac set with the modified primer Lac1o exhibited slightly better performance, although multiple bands also were obtained. Due to the proper separation of different species on the gel, as well as their verification by a distinctive sequence analysis, the rpoB1/rpoB1o/rpoB2GCprimer system seems to be a promising tool for monitoring the evolution of wine LAB. However, improvements should be made, as the detection limit of this set seems to be higher than that of the other primer sets. Owing to its pronounced sensitivity and its capability of discriminating to species level, the WLAB1/WLAB2GCprimer set turned out to be advantageous for LAB detection purposes in wine.
LITERATURE CITED
Andorra, I., Landi, S., Mas, A., Guillamón, J.M. & Esteve-Zarzoso, B., 2008. Effect of oenological practices on microbial populations using culture-independent techniques. Food Microbiol. 25, 849-856. [ Links ]
Bae, S., Fleet, G.H. & Heard, G.M., 2006. Lactic acid bacteria associated with wine grapes from several Australian vineyards. J. Appl. Microbiol. 100, 712-727. [ Links ]
Bousbouras, G.E. & Kunkee, R.E., 1971. Effect of pH on malo-lactic fermentation in wine. Am. J. Enol. Vitic. 22, 121-126. [ Links ]
Cho, G.-S., KrauB, S., Huch, M., Du Toit, M. & Franz C.M.A.P., 2011. Development of a quantitative PCR for detection of Lactobacillus plantarum starters during wine malolactic fermentation. J. Microbiol. Biotechnol. 21, 1280-1286. [ Links ]
Cocolin, L., Manzano, M., Cantoni, C. & Comi, G., 2001. Denaturing gradient gel electrophoresis analysis of the 16S rRNA gene V1 region to monitor dynamic changes in the bacterial population during fermentation of Italian sausages. Appl. Environ. Microbiol. 67, 5113-5121. [ Links ]
Davis, C.R., Wibowo, T.H.L. & Fleet, G.H., 1986. Growth and metabolism of lactic acid bacteria during and after malolactic fermentation of wines at different pH. Appl. Environ. Microbiol. 51, 539-545. [ Links ]
Endo, A. & Okada, S., 2005. Monitoring the lactic acid bacteria diversity during shochu fermentation by PCR-denaturing gradient gel electrophoresis. J. Biosci. Bioeng. 99, 216-221. [ Links ]
Ercolini, D., 2004. PCR-DGGE fingerprinting: Novel strategies for detection of microbes in food. J. Microbiol. Methods. 56, 297-314. [ Links ]
Janse, I., Bok, J. & Zwart, G., 2004. A simple remedy against artifactual double bands in denaturing gradient gel electrophoresis. J. Microbiol. Methods. 57, 279-281. [ Links ]
Knoll, C., Fritsch, S., Schnell, S., Grossmann, M., Krieger-Weber, S., Du Toit, M. & Rauhut, D., 2012. Impact of different malolactic fermentation inoculation scenarios on Riesling wine aroma. World J. Microbiol. Biotechnol. 28, 1143-1153. [ Links ]
Lerm, E., Engelbrecht L. & Du Toit, M., 2010. Malolactic fermentation: The ABC's of MLF. S. Afr. J. Enol. Vitic. 31, 186-212. [ Links ]
Lonvaud-Funel, A., 1999. Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie van Leeuwenhoek 76, 317-331. [ Links ]
Lopez, I., Ruiz-Larrea, F., Cocolin, L., Orr, E., Phister, T., Marshall, M., Vandergheynst, J. & Mills, D.A., 2003. Design and evaluation of PCR primers for analysis of bacterial populations in wine by denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 69, 6801-6807. [ Links ]
Lv, X.-C., Weng, X., Zhang, W., Rao, P.F. & Ni, L., 2012. Microbial diversity of traditional fermentation starters for Hong Qu glutinous rice wine as determined by PCR-mediated DGGE. Food Control 28, 426-434. [ Links ]
Mills, D.A., Rawsthorne, H., Parker, C., Tamir, D. & Makarova, K., 2005. Genomic analysis of Oenococcus oeni PSU-1 and its relevance to winemaking. FEMS Microbiol. Rev. 29, 465-475. [ Links ]
Moreno-Arribas, M.V. & Polo, M.C., 2005. Winemaking microbiology and biochemistry, current knowledge and future trends. Crit. Rev. Food Sci. and Nutr. 45, 265-286. [ Links ]
Muyzer, G., De Waal, E.C. & Uitterlinden, A.G., 1993. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 59, 695-700. [ Links ]
Pinzani, P., Bonciani, L., Pazzagli, M., Orlando, C., Guerrini, S. & Granchi, L., 2004. Rapid detection of Oenococcus oeni in wine by real-time quantitative PCR. Let. Appl. Microbiol. 38, 118-124. [ Links ]
Pozo-Bayón, M.A., Pardo, I., Ferrer, S. & Moreno-Arribas, M.V., 2009. Molecular approaches for the identification and characterisation of oenological lactic acid bacteria. Afr. J. Biotechnol. 8, 3995-4001. [ Links ]
Rantsiou, K., Comi, G. & Cocolin, L., 2004. The rpoB gene as a target for PCR-DGGE analysis to follow lactic acid bacterial population dynamics during food fermentations. Food Microbiol. 21, 481-487. [ Links ]
Renouf, V., Claisse, O. & Lonvaud-Funel, A., 2006a. RpoB gene: A target for identification of LAB cocci by PCR-DGGE and melting curves analysis in real time PCR. J. Microbiol. Methods 67, 162-170. [ Links ]
Renouf, V., Claisse, O. & Lonvaud-Funel, A., 2007. Inventory and monitoring of wine microbial consortia. Appl. Microbiol. Biotechnol. 75, 149-164. [ Links ]
Renouf, V., Claisse, O., Miot-Sertier, C. & Lonvaud-Funel, A., 2006b. Lactic acid bacteria evolution during winemaking: Use of rpoB gene as a target for PCR-DGGE analysis. Food Microbiol. 23, 136-145. [ Links ]
Renouf, V., Vayssieres, L.C., Claisse, O. & Lonvaud-Funel, A., 2009. Genetic and phenotypic evidence for two groups of Oenococcus oeni strains and their prevalence during winemaking. Appl. Microbiol. Biotechnol. 83, 85-97. [ Links ]
Spano, G., Lonvaud-Funel, A., Claisse, O. & Massa, S., 2007. In vivo PCR-DGGE analysis of Lactobacillus plantarum and Oenococcus oeni populations in red wine. Curr. Microbiol. 54, 9-13. [ Links ]
Walter, J., Hertel, C., Tannock, G.W., Lis, C.M., Munro, K. & Hammes, W.P., 2001. Detection of Lactobacillus, Pediococcus, Leuconostoc, and Weissella species in human feces by using group-specific PCR primers and denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 67, 2578-2585. [ Links ]
Wibowo, D., Fleet, G.H., Lee, T.H. & Eschenbruch, R.E., 1988. Factors affecting the induction of malolactic fermentation in red wines with Leuconostoc oenos. J. Appl. Bacteriol. 64, 421-428. [ Links ]
Submitted for publication: October 2013
Accepted for publication: May 2014
Acknowledgements: This study was funded by the Austrian Economic Chambers (WKO; Wirtschaftskammerpreis 2009)
* Corresponding author: sigrid.mayrhofer@boku.ac.at
Other authors: konrad.domig@boku.ac.at; wolfgang.kneifel@boku.ac.at

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}