










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.35 n.1 Stellenbosch 2014
Comparison of proanthocyanidins with different polymerisation degrees among berry skins of 'Shiraz', 'Cabernet Sauvignon', and 'Marselan'
Q. LiI; Y.-X. LiuII; Q.-H. PanI; C.-Q. DuanI; Y. ShiI,
ICentre for Viticulture and Oenology, College of Food Science & Nutritional Engineering, China Agricultural University, Beijing 100083, China
IIBeijing Key Laboratory of Bioactive Substances and Functional Foods, College of Arts and Science, Beijing Union University, Beijing 100191, China
ABSTRACT
Proanthocyanidins in grape berries are synthesised mainly before véraison, and very little attention is paid to the evolution of proanthocyanidins (PAs) in grapes from véraison to harvest. The present study focused on the changes of flavan-3-ols with different degrees of polymerisation in grape skins and the difference in proanthocyandin composition of 'Shiraz', 'Cabernet Sauvignon' and 'Marselan' grapes (Vitis vinifera L.). The results show that the content of flavan-3-ols, the percentage of prodelphinidins (%P) and mean degree polymerisation (mDP) found in 'Cabernet Sauvignon' berry skins at post-véraison were higher than those in 'Shiraz' and 'Marselan' skins. Only monomelic, dimeric, trimeric and polymeric flavan-3-ols were detected in the three grape cultivars. Polymers with more than tenfold flavan-3-ol units accounted for a relatively high proportion in grape berry skins, and the content in the three cultivars declined continuously during ripening. Principal component analysis showed that proanthocyanidin content, composition and mDP at grape harvest stage depended strongly on grape cultivar. This study provides some useful information for understanding the accumulation of PAs during berry maturation and this information can be used to improve wine quality.
Key words: Shiraz, Cabernet Sauvignon, Marselan, berry skin, proanthocyanidins, post-véraison
INTRODUCTION
Proanthocyanidins (PAs), a group of flavan-3-ol polymers, are synthesised via the flavonoid pathway in many plants, such as grape (Flamini, 2003; Peyrot des Gachons & Kennedy, 2003), cacao (Nelson & Sharpless, 2003), apple and hops (Hammerstone et al., 1999). In grapes, PAs are present in the berry skin, seed, stem and pulp tissues, and constitute a complex mixture of monomers, oligomers and even polymers. PAs generally consist of (+)-catechin (C), (-)-epicatechin (EC), (-)-epigallocatechin (EGC) and (-)-epicatechin-3-O-gallate (ECG) linked by C(4)-C(6) or C(4)-C(8) inter-flavanoid bonds. During winemaking, PAs are extracted primarily from the skin and seed into wine and contribute greatly to astringent and bitter properties and the colour stability of the wine (Robichaud & Noble, 1990; Soares et al., 2013). The composition or degree of polymerisation of PAs has been reported to perform different functions on the sensory properties of bitterness and astringency. For example, astringency becomes more intense with mean degree polymerisation (mDP) increasing, EC provides a more astringent taste than C, and an increasing degree of galloylation will contribute to a more coarse perception (Peleg et al., 1999; Maury et al., 2001; Vidal et al., 2003), but the increase in the percentage of B-ring trihydroxylation seems to decrease astringency (Vidal et al., 2003). Therefore, the distribution in polymerisation degrees of PAs in mature grape berries will determine the sensory quality of wine products to a large extent.
Some analytical methods have been developed for the quantification of PAs, of which the most common is to use acid n-butanol (Porter et al., 1985) and vanillin (Price et al., 1978) to quickly determine the content of PAs in plants. With the development of chromatographic and mass spectrogram technologies, researchers analyse PAs mostly through acid-catalysed cleavage in the presence of excess phloroglucinol or benzyl mercaptan (Gupta & Haslam, 1978; Matthews et al., 1997), and it is measured using reverse-phase high performance liquid chromagraphy (HPLC) equipped with mass spectrogram (MS), such as ion trap MS and TOF-MS. This method can acquire the composition and total contents of flavan-3-ols in grapes and wines and provide information on mDP, rather than the contents of individual oligomeric or polymeric flavan-3-ols. In addition, a normal-phase HPLC technique has also been developed to separate and measure flavan-3-ols with different polymerisation degrees (DP) (Gu et al, 2003; Vidal et al, 2004; Hellström & Mattila, 2008; Liu et al, 2010). It is thought that this normal-phase HPLC method shows more precise quantification of oligomers (2 < DP < 10) in comparison with other methods.
Although grape seeds contain abundant PAs, the compounds from grape berry skins are considered to be extracted more easily from plant tissues and into wines in the process of winemaking (Amrani & Mercierz, 1994). Moreover, the relative lack of ECG together with the presence of EGC has been speculated to confer a softening mouthfeel to wines (Vidal et al., 2004), which means that PAs from berry skins play an important role in improving the quality of red wines. The biosynthesis of PAs occurs mainly in the early stages of berry development and ends around véraison. The content of PAs decreases from véraison to ripening, possibly because they bind to proteins on the internal surface of the tonoplast and to polysaccharides on the cell wall (De Gaulejac et al, 1997). Cell wall extension during berry ripening increases the PA-binding capacity on cell walls, and the structure and degree of polymerisation of PAs also influence their affinity for binding materials (Hanlin et al, 2010; Bindon & Kennedy, 2011). Therefore, characterising the composition and content of PAs in grape skins after véraison will make it possible to better foresee their sensory contribution to wine.
The French term "terroir" indicates an interactive ecosystem, in a given place, including climate, soil and the vine (Cohen et al., 2012). The effect of "terroir" on the composition and content of flavan-3-ols or proanthocynadins has been studied, and the results have indicated that climatic factors, soil type and water status all could affect, to different extents, the biosynthesis and accumulation of flavan-3-ols (Bucchetti et al, 2011; Šeruga et al, 2011; Cohen et al, 2012; Koyama et al, 2012). The climate in the wine-growing regions of eastern China is characterised by relatively high temperatures and humidity in summer, which is different in many respects from the climate of other countries in the world. In this climate it still is necessary to determine what characters grape skin PAs possess. In the present study, the profiling of flavan-3-ols with different DP was followed during berry maturation and compared between three grape cultivars ('Shiraz', 'Cabernet Sauvignon' and 'Marselan') , with the objective of discovering the variation in flavan-3-ol profiling in mature grape berries and their potential impact on wine quality.
MATERIALS AND METHODS
Grape berry samples
In 2008, grape berries were collected from the end of véraison to commercial harvest from a commercial vineyard in Huailai County (40°N, 115°E), Hebei Province, China. The 'Cabernet Sauvignon', 'Shiraz' and 'Marselan' berries began colouring on August the 3rd, 5th and 7th respectively, with colouration completed about one week later. We sampled according to the method described by He et al. (2010) to represent a vineyard population. Three 100-berry samples were selected from at least seven clusters at a similar position on 30 whole vine selections. In the present study, the berry seeds were separated from the rest of the grape berries and were immediately frozen in liquid nitrogen, then ground to a fine powder, freeze-dried, and stored at -50°C until analysed.
Reagents and chemicals
The standards of (+)-catechin, (-)-epicatechin, (-)-epigal-locatechin, (-)-epicatechin-3-O-gallate and procyanidin B1 were all purchased from Sigma Chemical Co. (St. Louis, MO, USA). HPLC-grade methanol, methylene chloride, ace-tonitrile and acetic acid were obtained from Fisher Company (Fairlawn, NJ, USA); ascorbic acid and all the other chemicals were purchased from Sigma-Aldrich Co.; and deionised water (< 18ΜΩ resistibility) from a Milli-Q Element water purification system (Millipore, Bedford, MA, USA).
Extraction and purification of grape berry skin flavan-3-ols
The extraction and purification procedures were performed as described by Liu et al. (2010), with some modification. Briefly, the sample powder (0.5 g) was extracted with mixed solvent (10 mL, acetone/water, 70:30 v/v) containing ascorbic acid (1 g/L) by shaking, followed by sonication for 15 min. The tube was then incubated at 35°C for 30 min, and centrifuged at 10 000 χ g for 15 min. The precipitate was extracted four times, as discussed previously, and the supernatants were combined. Acetone was removed by a rotary evaporation at 30°C and then lyophilised to a dry powder. The crude proanthocyanidin was dissolved in 3 mL of 30% (v/v) aqueous methanol and then purified using a Sephadex LH-20 column (6 χ 1.5 cm).
Analysis of flavan-3-ols by normal-phase HPLC-MS
The extracts obtained above were filtered through 0.22 μm filters. According to Liu et al. (2010), the content of flavan-3-ols with a different polymerisation degree was determined by normal-phase HPLC using an Agilent 1200 series LC/MSD Trap VL mass spectrometer (Agilent Technologies, Palo Alto, CA, USA) equipped with an electrospray ionisation (ESI) interface, using an Agilent Zorbax RX-SIL (5 μm, 2.1 χ 150 mm) column protected with a Zorbax RX-SIL (5 μm, 4.6 χ 12.5 mm) guard column. Three mobile phases were used, as follows: A, methylene chloride, B, methanol, and C, acetic acid and water (1:1 v/v). The injection volume was 2 μL and the column temperature was set at 30°C. Elution was carried out at a constant flow rate of 0.2 mL/min: 0 to 20 min, 14.0 to 25.0% B linear; 20 to 40 min, 25.0 to 33.2% B linear; 40 to 45 min, 33.2 to 86.0% B linear; 45 to 55 min, 86.0% B isocratic; 55 to 60 min, 86.0 to 14.0% B linear; followed by 10 min of re-equilibration of the column before the next run. A constant 4.0% C was kept throughout the gradient.
Analysis of flavan-3-ols by reverse-phase HPLC-MS
The method of acid-catalysed cleavage of the proanthocyanidins in the presence of excess phloroglucinol and the analysis method of flavan-3-ols were as carried out by Kennedy and Jones (2001). Reverse-phase HPLC was run on an Agilent 1200 Series HPLC-DAD-ESI-MS/MS, and MS analysis was performed in the negative ionisation mode. The flavan-3-ol components were subjected to a reverse-phase chromato- graphic separation at 25°C using a Zorbax SB-C18 column (250x4.6 mm, 5 μm). A binary gradient elution was carried out at a constant flow of 1 mL/min. The mobile phases consisted of the following: A, 0.2% v/v aqueous acetic acid; and B, acetonitrile: mobile phase A (4:1). The elution conditions were modified for the solvent B gradient as follows: 0 min, 10%; 20 min, 10%; 30 min, 15%; 40 min, 20%; 50 min, 33%; 55 min, 40%; 58 min, 100%; 63 min, 100%; 64 min, 10%. Injection volumes were 25 μL and the DAD detection wavelength was 280 nm. The column was then re-equilibrated with 10% B for 5 min before the next injection.
Statistical analysis
Means and standard deviations were obtained from at least three repetitions. One-way ANOVA and Tukey's range test were used to evaluate the differences between grape cultivars, and principal component analysis was used to achieve a better description and discrimination of the composition of proanthocyanidins among the grape cultivars. Statistical analysis was performed by SPSS (SPSS Inc., Chicago, IL) for Windows, version 20.0.
RESULTS AND DISCUSSION
Comparison of flavan-3-ol units among three grape cultivars based on reverse-phase HPLC analysis
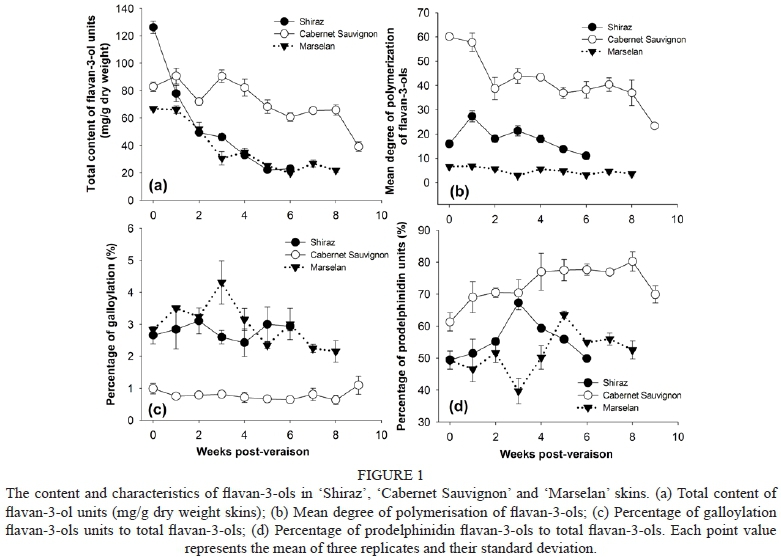
The content and composition of polymeric flavan-3-ols was determined in the berry skins of 'Shiraz', 'Cabernet Sauvignon' and 'Marselan' from véraison to commercial harvest by reverse-phase HPLC-MS after acid-catalysed cleavage in the presence of excess phloroglucinol. The total content of flavan-3-ol units (Fig. 1a) and their mDP (Fig. 1b) in the three cultivars all decreased gradually along with berry maturity, which is consistent with what has been reported previously (Downey et al., 2003; Gagné et al, 2006). Amongst the three cultivars, 'Cabernet Sauvignon' grape skins contained the highest level of flavan-3-ol units and mDP (60 to 23). At the beginning of véraison, the content of flavan-3-ols was relatively high in 'Shiraz' skins, but rapidly declined with berry maturity and reached a similar level to that of 'Marselan'. The mDP of flavan-3-ols in 'Shiraz' skins ranged from 13 to 27, which was in between the values of the other two cultivars studied. An increase in mDP from 21 to 36.6 has been observed in 'Cabernet Sauvignon' skins by Bordiga et al. (2011), and in 'Shiraz' skins the increase ranged from 25 to 40, as observed by Downey et al. (2003). The present analyses showed that there were lower levels of mDP when compared with the previous research mentioned described above, suggesting that this phenomenon might be related to differences in planting environment (such as ecological climate and vineyard microclimate), as flavan-3-ol biosynthesis is affected by different terroirs, cultural practices and vintages (Gagné et al., 2006; Fujita et al., 2007; Cohen et al, 2012; Koyama et al, 2012).
To understand differences in proanthocyandin aracteristics among grape cultivars, we also computed the percentages of galloylation derivatives (%G) and prodelphinidin (EGC) units (%P) (Fig. 1c and d, respectively). Vidal et al. (2004) showed that an increase in astringency correlated positively with the %G, which also would produce a potential impact on the intensity of coarseness. Moreover, a higher proportion of EGC smoothed the astringent perception of the wine (Vidal et al., 2004). The present study revealed that prodelphinidin (EGC) units in these three cultivars accounted for close on or more than 50%, which was much higher than the percentages of galloylation derivatives. In particular, the proportion of EGC units to total flavan-3-ols in 'Cabernet Sauvignon' reached 61% to 80%, and the percentage of galloylation units was only approximately 1%, which meant that proanthocyanidins in 'Cabernet Sauvignon' skins might provide more soft astringency to wine than that in 'Shiraz' and 'Marselan' skins.
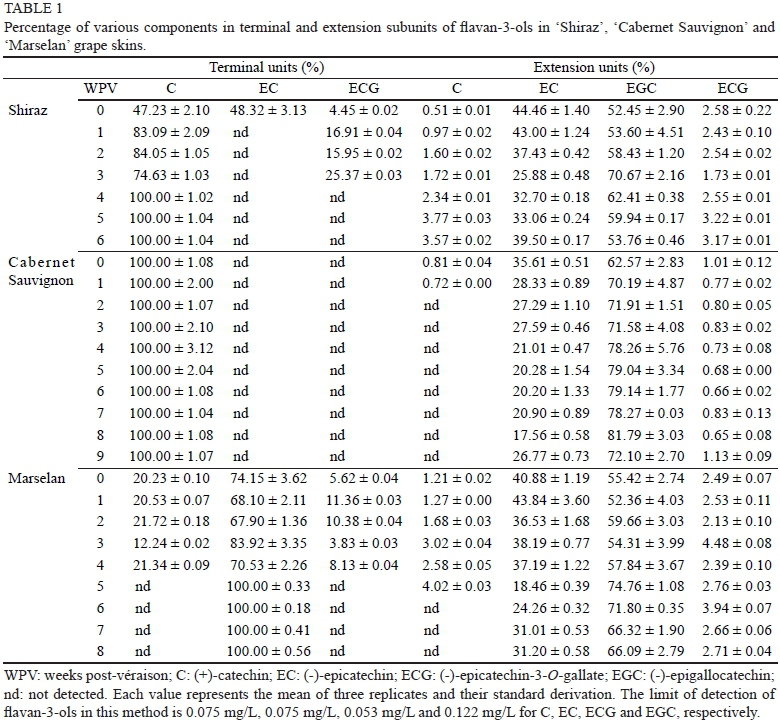
Table 1 showed the evolution of the extractable PAs fraction isolated by phloroglucinolysis from the berry skins of three grape cultivars during mature. Epigallocatechin-phloroglucinol (EGC-P), (-)-epicatechin-phloroglucinol (EC-P), (+)-catechin-phloroglucinol (C-P) and epicatechin-3- O-gallate-phloroglucinol (ECG-P) were identified as extension proanthocyanidin subunits, whereas (+)-catechin (C), (-)-epicatechin (EC) and (-)-epicatechin-3-О-gallate (ECG) were identified as terminal subunits in the berry skins. The differences in composition and content of terminal and extension subunits amongst grape cultivars reflected the variation in biosynthetic metabolism of flavan-3-ols to some extent.
Terminal subunit composition
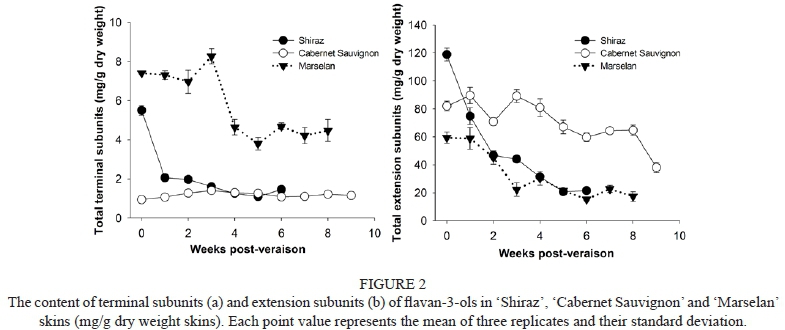
In 'Cabernet Sauvignon' berry skins, catechin was only one component examined as terminal subunit from véraison to harvest, while in the 'Shiraz' berry skin, epicatechin, catechin and a low percentage of epicatechin-gallate were observed as terminal subunits at véraison, but epicatechin and epicatechin-gallate disappeared one after the other at post-véraison (Table 1). Some literature has reported that the terminal subunit in the berry skins of 'Shiraz' and 'Cabernet Sauvignon' mainly comprises catechin (Downey et al., 2003; Busse-Valverde et al., 2010; Hanlin et al., 2010). Few studies have examined the PAs of 'Marselan' grapes. Here we found that epicatechin took up a large proportion (67% to 100%), while catechin (11% to 19%) and epicatechin-3-O-gallate (3% to 10%) could be examined only during the first four weeks post-véraison (Table 1). The contents of terminal subunits in 'Cabernet Sauvignon' berry skins were almost unchanged throughout berry ripening. In contrast, the contents of terminal subunits in 'Shiraz' berry skin were higher than in 'Cabernet Sauvignon' at véraison, then declined considerably from véraison to harvest. The terminal subunit in 'Marselan' berry skins was the highest detected in the three cultivars (Fig. 2a).
Extension subunit composition
The contents of extension subunits in the three grape berry skins declined gradually during ripening (Fig. 2b). For 'Shiraz', 'Cabernet Sauvignon' and 'Marselan' berry skins, epigallocatechin had the highest proportion of extension subunit, and its ratio to total content of extension subunits along with berry maturity fluctuated, ranging between 52.45% and 70.67% in 'Shiraz', between 62.57% and 81.79% in 'Cabernet Sauvignon', and between 52.36% and 74.76% in 'Marselan'. In addition, epicatechin was another major extension subunit for the three cultivars. The proportion of epicatechin first decreased and then increased slightly. The overall trend from véraison to harvest changed downwards. The catechin and epicatechin-3-O-gallate extension subunits accounted for less than 5%, and were undetectable after one week of post-véraison in 'Cabernet Sauvignon' and five weeks of post-véraison in 'Marselan'. These results were consistent with earlier reports, in which epigallocatechin and epicatechin were considered to be the most abundant extension subunits, and catechin and epicatechin-3-O-gallate were present in much lower proportions in 'Shiraz' and 'Cabernet Sauvignon' berry skins (Hanlin et al., 2010). The above results indicate that the composition of PAs from the skins of these three cultivars are different in proportion. It therefore is entirely possible that they would provide wines with different mouthfeel characters.
Comparison of flavan-3-ols with different polymerisation degrees in three grape cultivars based on normal-phase HPLC analysis
Using normal-phase HPLC, the separation of PAs was based on their degree of polymerisation (DP). Flavan-3-ols consist of monomers (DP = 1), oligomers (2 ≤ DP ≤ 10) and polymers (DP > 10) with different degrees of polymerisation (DP) (Gu et al., 2003). In the three grape cultivars, monomers decreased as the berries ripened, while the dimers and trimers fluctuated throughout maturity. The mass spectra signals of other oligomers (4 ≤ DP ≤ 10) could be tested, but no chromatogram information corresponding to these oligomers was found in HPLC, and the mass spectra signals weakened with maturity (data not shown). Polymers (DP > 10) were the main form of flavan-3-ols in the grape berry skins.
As shown in Table 2, the levels of monomers in the three cultivars were relatively low at véraison and decreased slightly to trace level before harvest. Meanwhile, dimers and trimers in 'Shiraz' and 'Marselan' increased in the first two weeks post-véraison, and then maintained a high level, with some fluctuations in the period following maturity. 'Cabernet Sauvignon' berry skins showed a notable increase in the content of oligomeric flavan-3-ols from véraison to the fourth week post-véraison, followed by a decline at harvest. The contents of polymeric flavan-3-ols (DP > 10) in the three cultivars declined continuously during ripening. Regarding the content of various flavan-3-ols at commercial harvest, we found that the skins of 'Shiraz' contained more oligomers than the other two grape cultivars, and the content of polymers in the skins of 'Cabernet Sauvignon' was higher than that in 'Shiraz' and 'Marselan'. 'Marselan' had the lowest level of oligomers and polymers.
To better understand the evolution of the polymerisation of flavan-3-ols during berry maturation, we evaluated the relative proportion of flavan-3-ols of different polymerisation to all the detected flavan-3-ols. At post-véraison, the proportions of monomers in 'Shiraz' and 'Cabernet Sauvignon' were virtually constant, but declined sharply in 'Marselan' (Fig. 3a). In 'Shiraz' and 'Marselan', a similar tendency of change was observed in the oligomers and polymers. For these two cultivars, the proportion of dimers (Fig. 3b) and trimers (Fig. 3c) increased gradually, and the proportion of polymers (Fig. 3d) decreased from véraison to harvest by 34% and 20%, respectively.
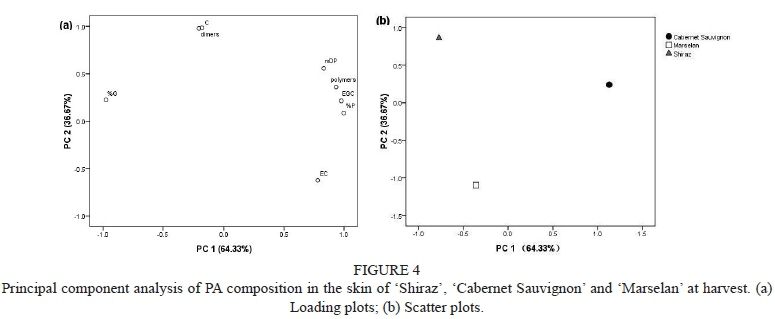
Principal component analysis (PCA)
To obtain an intuitive understanding of the varietal difference in PA composition, we conducted PCA using a number of variables, including the content of EGC, C, EC units, dimers, polymers and mDP, %P and %G at harvest, which were significantly different among the cultivars according to ANOVA (p < 0.05). As shown in Fig. 4a, the first and second principal components together explained 100% (PC1 64.28% and PC2 35.72%) of the total variance. PC1 was negatively correlated with the levels of C, dimers and %G, whereas PC2 was negatively related to EC. A projection of the cases of the first two components showed that the three cultivars could be differentiated readily (Fig. 4b). 'Cabernet Sauvignon' was found on the first factorial plane, which had a strong positive correlation with EGC, mDP, %P and polymers. 'Shiraz' was located on the second factorial plane, which showed a strongly positive correlation with C, dimers and %G, while 'Marselan' did not correspond well to the variables we chose here. The results further showed a great difference in PA composition among the three cultivars at harvest. 'Cabernet Sauvignon' skin was richer in EGC subunits and polymers, and the values of %P and mDP were higher when compared to the other two cultivars, while 'Shiraz' contained more C subunits and dimers, and the proportion of galloylation derivatives was higher than in the 'Cabernet Sauvignon' and 'Marselan' skins.
CONCLUSIONS
By integrating the data above, we found that the pattern of change in the proanthocyanidins in the studied cultivars was siilar, but there also were some differences in PA contents and composition. As all the samples were collected from similar vineyards with similar soil types and cultural practices, these differences observed between the three cultivars should be due mainly to the intrinsic characteristics of the selected cultivars. Among them, 'Cabernet Sauvignon' skin was characterised by the highest levels of flavan-3-ol units, polymers, percentage of prodelphinidins (%P) and mDP; 'Shiraz' skin presented the highest levels of catechins, dimers and percentage of galloylation (%G), and the lowest level of flavan-3-ols; while 'Marselan' skins exhibited the lowest mDP and the highest proportions of monomers and oligomers (dimers and trimers) to total flavan-3-ols. Several studies have indicated that perceived astringency and bitterness are significantly correlated with flavan-3-ol levels and composition. As a result, we speculated that 'Cabernet Sauvignon' could provide more of a soft, astringent perception to wine than 'Shiraz', and that the latter could provide more astringency and bitterness to wine when compared to 'Marselan'.
LITERATURE CITED
Amrani, J. & Mercierz, M., 1994. Localisation des tanins dans la pellicule de baie de raisin. Vitis 33, 133-138. [ Links ]
Bindon, K.A. & Kennedy, J.A., 2011. Ripening-induced changes in grape skin proanthocyanidins modify their interaction with cell walls. J. Agric. Food Chem. 59, 2696-2707. [ Links ]
Bordiga, M., Travaglia, F., Locatelli, M., Coïsson, J.D. & Arlorio, M., 2011. Characterisation of polymeric skin and seed proanthocyanidins during ripening in six Vitis vinifera L. cv. Food Chem. 127, 180-187. [ Links ]
Bucchetti, B., Matthews, M.A., Falginella, L., Peterlunger, E. & Castellarin, S.D., 2011. Effect of water deficit on Merlot grape tannins and anthocyanins across four seasons. Sci. Hortic. Amsterdam 128, 297-305. [ Links ]
Busse-Valverde, N., Gómez-Plaza, E., López-Roca, J.M., Gil-Munoz, R., Fernández-Fernández, J.I. & Bautista-Ortín, A.B., 2010. Effect of different enological practices on skin and seed proanthocyanidins in three varietal wines. J. Agric. Food Chem. 58, 11333-11339. [ Links ]
Cohen, S.D., Tarara, J.M., Gambetta, G.A., Matthews, M.A. & Kennedy, J.A., 2012. Impact of diurnal temperature variation on grape berry development, proanthocyanidin accumulation, and the expression of flavonoid pathway genes. J. Exp. Bot. 63, 2655-2665. [ Links ]
De Gaulejac, N.S.C., Augustin, M., Vivas, N. & Glories, Y., 1997. A biochemical approach to the evolution of procyanidins in grape seeds during the ripening of red grapes (Vitis vinifera L. cv. Merlot Noir). J. Wine Res. 8, 159-167. [ Links ]
Downey, M.O., Harvey, J.S. & Robinson, S.P., 2003. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine R. 9, 15-27. [ Links ]
Flamini, R., 2003. Mass spectrometry in grape and wine chemistry. Part I: Polyphenols. Mass Spectrom. Rev. 22, 218-250. [ Links ]
Fujita, A., Soma, N., Goto-Yamamoto, N., Mizuno, A., Kiso, K. & Katsumi, H., 2007. Effect of shading on proanthocyanidin biosynthesis in the grape berry. J. Jpn. Soc. Hortic. Sci. 76, 112-119. [ Links ]
Gagné, S., Saucier, C. & Gény, L., 2006. Composition and cellular localization of tannins in Cabernet Sauvignon skins during growth. J. Agric. Food Chem. 54, 9465-9471. [ Links ]
Gu, L., Kelm, M.A., Hammerstone, J.F., Beecher, G., Holden, J., Haytowitz, D. & Prior, R.L., 2003. Screening of foods containing proanthocyanidins and their structural characterization using LC-MS/MS and thiolytic degradation. J. Agric. Food Chem. 51, 7513-7521. [ Links ]
Gupta, R.K. & Haslam, E., 1978. Plant proanthocyanidins. Part 5. Sorghum polyphenols. Journal of the Chemical Society, Perkin Transactions 1, 892896. [ Links ]
Hammerstone, J.F., Lazarus, S.A., Mitchell, A.E., Rucker, R., & Schmitz, H.H., 1999. Identification of procyanidins in cocoa (Theobroma cacao) and chocolate using high-performance liquid chromatography/mass spectrometry. J. Agric. Food Chem. 47, 490-496. [ Links ]
Hanlin, R., Hrmova, M., Harbertson, J. & Downey, M.O., 2010. Review: Condensed tannin and grape cell wall interactions and their impact on tannin extractability into wine. Aus. J. Grape Wine Res. 16, 173-188. [ Links ]
He, J.J., Liu, Y.X., Pan, Q.H., Cui, X.Y., & Duan, C.Q., 2010. Different anthocyanin profiles of the skin and the pulp of Yan7 (Muscat Hamburg x Alicante Bouschet) grape berries. Molecules 15, 1141-1153. [ Links ]
Hellström, J.K. & Mattila, P.H., 2008. HPLC determination of extractable and unextractable proanthocyanidins in plant materials. J. Agric. Food Chem. 56, 7617-7624. [ Links ]
Kennedy, J.A. & Jones, G.P., 2001. Analysis of proanthocyanidin cleavage products following acid-catalysis in the presence of excess phloroglucinol. J. Agric. Food Chem. 49, 1740-1746. [ Links ]
Koyama, K., Ikeda, H., Poudel, P.R. & Goto-Yamamoto, N., 2012. Light quality affects flavonoid biosynthesis in young berries of Cabernet Sauvignon grape. Phytochem. 78, 54-64. [ Links ]
Liu, Y.X., Pan, Q.H., Yan, G.L., He, J.J. & Duan, C.Q., 2010. Changes of flavan-3-ols with different degrees of polymerization in seeds of 'Shiraz', 'Cabernet Sauvignon' and 'Marselan' grapes after véraison. Molecules 15, 7763-7774. [ Links ]
Matthews, S., Mila, I., Scalbert, A., Pollet, B., Lapierre, C., Hervé du Penhoat, C.L., Rolando, C. & Donnelly, D.M., 1997. Method for estimation of proanthocyanidins based on their acid depolymerization in the presence of nucleophiles. J. Agric. Food Chem. 45, 1195-1201. [ Links ]
Maury, C., Sarni-Manchado, P., Lefebvre, S., Cheynier, V. & Moutounet, M., 2001. Influence of fining with different molecular weight gelatins on proanthocyanidin composition and perception of wines. Am. J. Enol. Vitic. 52, 140-145. [ Links ]
Nelson, B.C. & Sharpless, K.E., 2003. Quantification of the predominant monomeric catechins in baking chocolate standard reference material by LC/APCI-MS. J. Agric. Food Chem. 51, 531-537. [ Links ]
Peleg, H., Gacon, K., Schlich, P. & Noble, A.C., 1999. Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. J. Sci. Food Agr. 79, 1123-1128. [ Links ]
Peyrot des Gachons, C. & Kennedy, J.A., 2003. Direct method for determining seed and skin proanthocyanidin extraction into red wine. J. Agric. Food Chem. 51, 5877-5881. [ Links ]
Porter, L.J., Hrstich, L.N. & Chan, B.G., 1985. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochem. 25, 223-230. [ Links ]
Price, M.L., Van Scoyoc, S. & Butler, L.G., 1978. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 26, 1214-1218. [ Links ]
Robichaud, J.L. & Noble, A.C., 1990. Astringency and bitterness of selected phenolics in wine. J. Sci. Food Agric. 53, 343-353. [ Links ]
Šeruga, M., Novak, I. & Jakobek, L., 2011. Determination of polyphenols content and antioxidant activity of some red wines by differential pulse voltammetry, HPLC and spectrophotometric methods. Food Chem. 124, 1208-1216. [ Links ]
Soares, S., Kohl, S., Thalmann, S., Mateus, N., Meyerhof, W. & De Freitas, V., 2013. Different phenolic compounds activate distinct human bitter taste receptors. J. Agric. Food Chem. 61, 1525-1533. [ Links ]
Vidal, S., Francis, L., Guyot, S., Marnet, N., Kwiatkowski, M., Gawel, R., Cheynier, V.R. & Waters, E.J., 2003. The mouth-feel properties of grape and apple proanthocyanidins in a wine-like medium. J. Sci. Food Agric. 83, 564-573. [ Links ]
Vidal, S., Francis, L., Noble, A., Kwiatkowski, M., Cheynier, V. & Waters, E., 2004. Taste and mouth-feel properties of different types of tannin-like polyphenolic compounds and anthocyanins in wine. Anal. Chimica Acta 513, 57-65. [ Links ]
Submitted for publication: August 2013
Accepted for publication: September 2013
Aknowledgements: This work was supported by the National Natural Science Foundation of China (30871746) and the Earmarked Fund for China Agriculture Research System (Grant No. CARS-30)
* Corresponding author: e-mail address: e-mail address: shiy@cau.edu.cn [Tel.: +86-10-62737136; Fax: +86-10-62737136]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}