










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Enology and Viticulture
On-line version ISSN 2224-7904
Print version ISSN 0253-939X
S. Afr. J. Enol. Vitic. vol.35 n.1 Stellenbosch 2014
Effect of berry weight and its components on the contents of sugars and anthocyanins of three varieties of Vitis vinifera L. under different water supply conditions
M. FerrerI, *; G. EcheverríaII; A. CarbonneauIII
IFacultad de Agronomia, Av. Garzon 780, Montevideo, Uruguay
IIFacultad de Agronomia, Av. Garzon 780, Montevideo, Uruguay
IIISup-Agro 2, Place Viala, Montpellier, France (carbonne@supagro.inra.fr)
ABSTRACT
The influence of berry size on wine quality is a matter of debate. With the aim to contribute to that debate, a study of the influence of the varieties Tannat, Merlot and Cabernet Sauvignon on berry weight, on berry components (flesh, seeds and skin) and on the varieties' oenological potential under different water supply conditions was carried out in Uruguay between 2001 and 2004. Replicated samples of 250 berries per vineyard were gathered at technological maturity at harvest. Berry weight and volume, weight ratio among each of the components and anthocyanin potential (ApH1 ) were measured and a basic analysis of the must was carried out. The vine water status was established through the measurement of pre-dawn leaf water potential. The tests showed that berry weight and sugar content were positively correlated and depended on variety, as was evident from the proportions of the berry components. Flesh weight was the most important determinant of berry weight, and flesh weight changed with the weight of the seeds. Larger values of these two components explained the heavier berries of the Tannat variety. On the other hand, Tannat berries had higher contents and concentrations of anthocyanins and sugars, and displayed the largest variation in these chemical compounds. Berry weight, weight and proportion of skin or their relationships did not determine anthocyanin contents or concentration. In response to the years' water conditions, different varieties showed different behaviours, explaining the variation in weight and in the composition of the berries.
Key words: Berry size, composition, components, water availability, varieties
INTRODUCTION
The influence of berry size on grape composition and hence on the quality of red wine is the subject of much debate. Authors have shown the relationship between berry weight and the contents of components determining must quality (Roby & Matthews, 2004; Santesteban & Royo, 2006, Dai et al., 2011). However, according to Roby et al., (2004), the berry content of all solutes increased approximately in proportion to the increase in berry size, and other researchers have reported that other factors besides the size of the berry, such as temperature, cultivation techniques, rainfall, etc., are involved in grape composition (Mori et al., 2005, 2007; Walker et al., 2005; Holt et al., 2008; Chaves et al., 2007). Walker et al. (2005) and Matthews & Nuzzo (2007)
concluded that the resultant winemaking traits of large or small berries depend more on the factor modifying the weight of the berry (variety, water deficit, etc.) than on the size itself. The way in which composition is formulated (content or concentration) also needs to be taken into account when the effects of berry size are being analysed (Matthews & Nuzzo, 2007). Variety is one of the major factors determining the difference in size and composition of the berry as a result of specific genetic characteristics associated with growth and the relative proportion of the components (flesh, seeds and skin) and their relationships (Matthews & Nuzzo, 2007; Constantini et al., 2008; Attia et al., 2010; Barbagallo et al., 2011; Dai et al., 2011; Nicolas et al., 2013). Genetics determines composition and has an influence on the ability of the variety to accumulate compounds, on the way in which photosynthetic products are distributed within the plant and on their influence on secondary metabolism (Edson et al., 1995; Gomez del Campo et al., 2005; Liu et al., 2007; Sadras et al., 2008; Dai et al., 2011). Among the components of the berry, flesh represents the largest proportion of weight and is the place where most sugars accumulate (Matthews & Nuzzo, 2007; Barbagallo et al., 2011). Weight and number of seeds have a direct impact on total berry weight due to the hormonal regulation exerted by the seeds on cell proliferation and expansion (Ristic & Iland, 2005; Walker et al, 2005; Friend et al, 2009; Gray & Combe, 2009; Attia et al, 2010). The skin is where the synthesis of anthocyanins takes place (Coombe, 1989). The relationships between the various components provide information on the oenological potential of the wine grape (Attia et al., 2010; Chaves et al., 2007; Barbagallo et al, 2011).
Environmental conditions and the response of the variety to different situations are also directly involved in determining the size and composition of the grapes. The relationship between vine water status, the size of the berry and grape composition has been described by numerous researchers (Mathews and Anderson, 1988; Williams & Matthews, 1990; Ojeda et al., 2001, Roby et al, 2004; Chaves et al, 2007; Ferrer et al., 2008; Girona et al., 2009). Berry size changes in response to water deficit, mostly when low water supply occurs during the first stages of development, as a result of an increase in abscisic acid (ABA), which limits cell division and expansion. Different components respond to stress conditions according to the variety, with the flesh being the most decisive one in reducing berry size (Ojeda et al., 2001; Roby & Matthews, 2004; Dai et al., 2011). Several authors have stated that the response to water conditions (excess or deficit) is not similar in different varieties and that water availability has a greater influence on berry composition than on size (Williams & Matthews, 1990; Mori et al., 2005; Santesteban & Royo, 2006; Castellarin et al., 2007; Ozden et al., 2010). Under stress conditions, an increasing supply of ABA generates responses in fruit composition. It is possible that ABA is synthesised under water stress, culminating in accelerated sugar and anthocyanin accumulation and malic acid breakdown (Castellarin et al., 2007). According to Roby et al. (2004), in addition to decreasing berry size, water deficits increase the amount of skin anthocyanin per berry and the concentration of skin anthocyanins. Their results show that there are effects of vine water status on fruit composition that arise independently of the resultant differences in fruit size. The effect of vine water status on the concentration of skin anthocyanin was greater than the effect of fruit size on this same variable. However, the increase in skin anthocyanin that accompanied water deficits appeared to result more from differential growth sensitivity of the inner mesocarp and exocarp than direct effects on phenolic biosynthesis. Moreover, stress conditions happening in the presence of calcium stimulate the activity of PAL (phenylalanine ammonia lyase), which acts as a primary enzyme or entrance input to the phenylpropanoid pathway, one of the most important paths in the synthesis of phenolic compounds.
In order to contribute to this discussion, a study on the influence of vine varieties (Tannat, Merlot and Cabernet Sauvignon) on berry size, berry components and their oenological potential under different water supply conditions was carried out in Uruguay between 2001 and 2004.
MATERIALS AND METHODS
The experiment was established in a seven-year-old commercial vineyard (cvs Cabernet Sauvignon, Merlot and Tannat) located in southern Uruguay (34°35'12.43" S; 56°15'2.26 W). The vines were planted at a spacing of 2.5 m between rows and 1.25 m between vines, trained to a VSP trellis system with rows oriented North-South, a canopy height of 1.85 m, and spur pruned leaving 18 buds per linear metre, with an average density of 3.85 to 4.20 leaf layers. The climate of the region is classified by the Multicriteria Climatic Classification System (Tonietto & Carbonneau, 2004; Ferrer, 2007) as temperate, temperate nights, moderately dry, with a sea influence. The maximum and minimum average temperatures in summer are 28.9°C and 16.4°C respectively.
The soil of the experimental site (Argiudol, USDA) is characterised by a typical profile: A (0 to 15 cm), Bt (15 to 85 cm) and C (> 85 cm), with the presence of a water table that is within reach of the roots. The average textural composition of the upper horizon is: 19.7% sand, 44.7% silt and 39.7% clay. The soil water-holding capacity is 141.6 mm per m soil depth.
The trials took place in adjacent plots over four years (2001 to 2004). In all cases the rootstock was SO4 and the experiment was laid out as a completely randomised design, with 30 plants of each variety.
Grape samples
Samples of 250 berries were taken at harvest, with two repetitions for each vineyard, employing the method proposed by Carbonneau et al. (1991). Three to five berries were sampled from clusters located on shoots in the middle of the vine. Berries were selected randomly from the upper and lower parts of a cluster. The harvest took place at "technological maturity" in all cases. For that purpose, the relationship between the sugar content (g/L), total acidity (g/L sulphuric acid) and pH of the grapes was determined once a week, starting at véraison, by taking samples of 250 berries with two sampling repetitions for each variety.
The decision to harvest was taken according to the variety and the characteristics of the year. Considering the maximal sugar contents (g/L), total acidity values (g H2SO4) were in the range 5.1 to 5.4 for Tannat, 4.1 to 4.4 for Cabernet Sauvignon and 3.6 to 4.3 for Merlot; and the pH values to obtain mature grapes were in ranges between 3.33 and 3.40 for Tannat, 3.44 and 3.66 for Cabernet Sauvignon and 3.40 and 3.67 for Merlot. Besides this, the evolution of the weight of the berries (g) was taken into account in order to prevent them becoming dehydrated.
Basic analysis of grapes and berry structure
Half of the berries of each sample were used to undertake a basic analysis and determine berry weight, the density of the musts and the relative proportions of skins, seeds and flesh. The weight of the berries was determined with an Ohaus Scout scale (Ohaus Corp., USA). The berries were then crushed manually in a mortar in order to remove the skins, seeds and flesh. The skins and seeds were rinsed under water to completely separate the pulp and dissolve the residual sugars, after which they were dried with filter paper and weighed. The weight of the flesh of each sample was calculated by using the difference between the berry weight and the skin and seed weights. The relative proportion of each part of the grape (flesh, skins and seeds) was calculated. Basic composition as realised in the juice obtained from the manual crushing of the berries and the crushing of the pulp with a juice extractor, Phillips HR2290 (Phillips, Netherlands) was determined by employing classical analyses (sugar content, total acidity and pH). Analyses were carried out according to the O.I.V. (2007), using a Atago N1 refractometer (Atago, Japan) and a Hanna HI8521 pH meter (Hanna Instruments, Italy). The must density was estimated from sugar concentrations determined by refractometry.
Phenolic potential of the grapes
Half of the berries of each sample were analysed according to Glories and Augustin (1995) to determine the total potential in anthocyanins (ApH1). After grinding the grapes with a blender, model solutions at pH 1.0 were added and the mixture was homogenised and macerated for four hours. The extracts were filtered and centrifuged for 3 min at 3 500 rpm, using a MSE Mistral 2000 centrifuge (Sanyo-Gallenkamp, Great Britain), before analysis. Anthocyanin content at pH 1.0 was measured according to Ribéreau-Gayon and Stonestreet (1965). The measurements were carried out by duplication with a Shimadzu UV-1240 Mini (Shimadzu, Japan) spectrophotometer, using glass (for the anthocyanin analyses) and quartz cells (for the analyses of absorbance at 280 nm) with 1 cm path length. The indexes were calculated, considering the respective dilution of the grape extracts, according to González-Neves et al. (2004). The total content of sugars and anthocyanins was calculated using the method proposed by Vila et al. (2010):
Total sugar (g/berry) = soluble solids (g/L) x berry weight (g)/(0.0046 x Brix + 0.9927)/1000.
Total anthocyanins (mg/berry) = anthocyanin pH1 (mg/L) x berry weight (g)/(0.0046 x Brix + 0,9927)/1000. Sugar expressed as g/berry, and g/g berry; anthocyanin as mg/berry and mg/g berry.
Determination of pre-dawn leaf water potential (ΨPD)
The pressure chamber technique (Scholander et al., 1965) was used (Soil moisture equipment mod. 3005-1412) to determine pre-dawn leaf water potential. These measurements were made before dawn at véraison, in 20 adult, healthy and fresh leaves per vineyard (10 on the eastern side of the canopy and 10 on the western side of the canopy).
Determination of dry matter per organ
The annual amount of dry matter of each variety was assessed at harvest from 15 representative vine shoots holding bunches. The material was placed in an oven at 60°C until the weight had stabilised. After removing the materials from the oven, each organ (leaves, bunches, shoots) was weighed using an Ohaus Scout ± 0.1 g (Ohaus Corporation, USA) scale.
Statistical analyses
Analysis of variance was performed on the surveyed variables, followed by the Tukey test (p <0.05), and correlations were established using the Pearson r method. The statistical package InfoStat Version 2011 was used.
RESULTS AND DISCUSSION
Berry weight and berry components
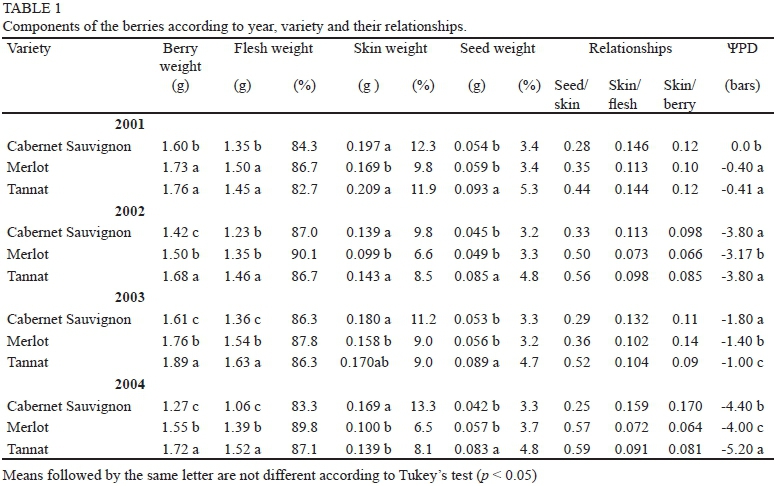
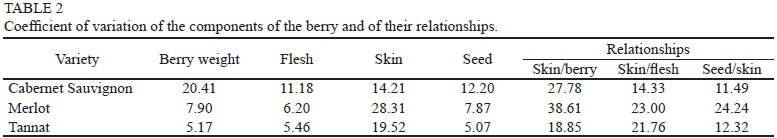
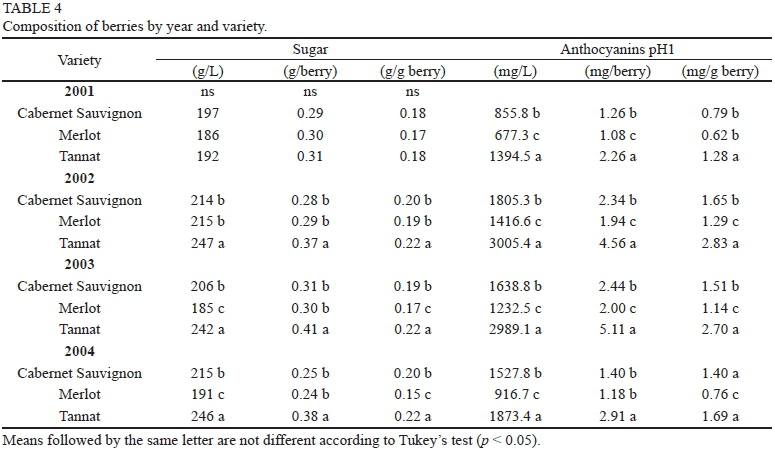
Berry weight showed significant differences depending on the variety and vintage, in accordance with Roby andMatthews, (2004), Attia et al. (2010) Chaves et al. (2007)and Dai et al. (2011). Tannat berries were heavier and the differences were significant in three years. The greatest weight variation was recorded for Cabernet Sauvignon (cv. 20.41, 0.29 g), followed by Merlot (cv. 7.90, 0.13 g) and the more stable Tannat (cv. 5.17, 0.09 g) (Tables 1 and 2).
The varieties presented differences in weight and proportion of components, which were dependent on the environmental conditions of the year. Considering the four years, the highest flesh proportion was registered for Merlot (86.8%). Cabernet Sauvignon was the variety with the highest skin percentage (11.5%), and the largest seed weight proportion was found in Tannat (4.9%). The registered values are within the ranges cited by Roby & Matthews (2004), Matthews & Nuzzo (2007) and Attia et al. (2010) (Table 1).
Flesh weight
Flesh weight was the main determinant of berry weight and its variation, and the established correlation was r = 0.97 (p = 0.006), in accordance with Barbagallo et al. (2011). Flesh weight was significantly different in the three varieties (p < 0.05), being heavier for Tannat in three of the four years analysed. The largest variation in weight was recorded for Cabernet Sauvignon (cv. 11.18, 0.14 g), followed by Merlot (cv. 6.20, 0.09 g) and, finally, Tannat (cv. 5.46, 0.08 g). This accounted for most of the variation in berry weight within varieties.
Seed weight
Seed weight was correlated with total berry weight (r = 0.94, p < 0.0001), in accordance with Barbagallo et al. (2011), and with flesh weight (r = 0.70, p = 0.06). Average seed weight for the Tannat variety was higher than for the other varieties (p < 0.05), whereas no significant differences were found between the seeds of Cabernet Sauvignon and Merlot. This correlation between the weight of berries and seeds is explained by growth regulators supplied by the latter. Growth regulators promote cell multiplication and enlargement (Ristic & Iland, 2005; Walker et al., 2005; Friend et al., 2009; Gray & Combe, 2009; Attia et al., 2010). Constantini et al. (2008) indicated genetic differences among varieties related to QTL on LG18, associating berry size with seed weight. These results could explain the variation in total berry weight of each variety and the heavier weight of the Tannat berries. The largest variation in seed weight was found in Cabernet Sauvignon (cv. 12.20, 0.06 g), followed by Merlot (cv. 7.87, 0.05 g) and Tannat (cv. 5.07, 0.04 g).
Skin weight
Skin weight did not show a correlation with berry weight, contradicting assertions by Mathews & Nuzzo (2007), Attia et al. (2010) and Barbagallo et al. (2011) for other varieties. The skin of Cabernet Sauvignon berries was found to be heavier than the skin of Merlot and significantly heavier than the skin of Tannat berries in one of the four years of the trial.
Berry weight differences depending on variety, particularly on the components flesh and skin, can be explained on the basis of research done by Nicolas et al. (2013). These authors identified a gene (VvCEB1) associated with berry growth as a consequence of the cell enlargement that occurs in development stages 1 and 3. The expression of this gene is specific to the species and the variety of vine and is expressed in the fruit in the mesocarp and exocarp. The largest variation in the skin weight was recorded in Merlot (cv. 28.31, 0.04 g), followed by Tannat (cv. 19.42, 0.03 g) and finally Cabernet Sauvignon (cv. 14.24, 0.02 g). From an oenological point of view, this result is of interest to the extent that the skin is where the synthesis and accumulation of anthocyanins take place (Coombe, 1989).
The analysis of the relationship of the skin with other components provides information on the oenological potential of the wine grape (Attia et al., 2010; Barbagalloet al., 2011).
The ratio skin weight/berry weight was significant (r = 0.66, p = 0.10), which would indicate that smaller berries have a relatively larger skin, as is the case in the Cabernet Sauvignon. This is a variety in which this relationship was higher in three of the four years of the trial (cv. = 27.78). The maximum variation in the ratio was in Merlot (cv. = 38.61) and the lowest in Tannat (cv. = 18.85).
Relationship between seed weight and skin weight The relationship between seed weight and skin weight was lower in Cabernet Sauvignon than in the other two cultivars (p < 0.05), which did not show any differences. The greatest variation was recorded in Merlot (cv. = 24.24), followed by Tannat (cv. = 12.32) and Cabernet Sauvignon (cv. = 11.49). Chaves et al. (2007) argue that the skin/flesh relationship is higher in smaller berries and that this would explain their higher content of anthocyanins. In our study, the correlation between berry weight and this relationship, although negative, was not significant (r = -0.34, p = 0.18). Accordingly, the statements made by these authors cannot be supported by our data. The greatest variation in this relationship was recorded in Merlot (cv. = 23.00), followed by Tannat (cv. = 21.76) and Cabernet Sauvignon (cv. = 14.33) (Tables 1 and 2).
Change in berry size - influence of water availability
Berry weight
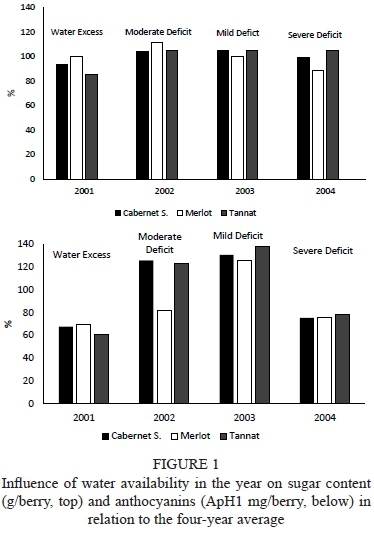
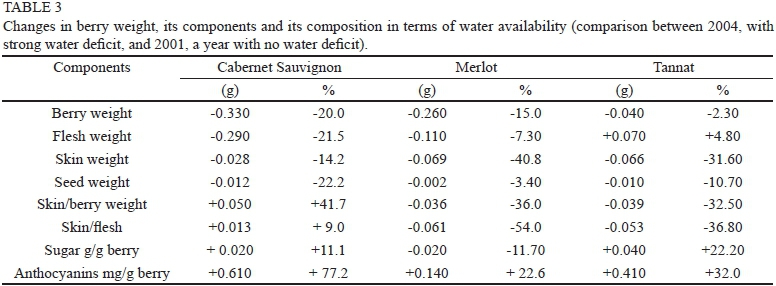
The four-year trial showed variable water conditions. Measured as pre-dawn leaf water potential (WD), those conditions influenced berry weight (r = - 0.63, p = 0.10), and that influence depended on the variety (Fig. 1), in agreement with the assertions of several authors (Ojeda et al., 2001; Roby & Matthews, 2004; Roby et al., 2004; Chaves et al., 2007; Ferrer, 2007; Matthews & Nuzzo, 2007; Ferrer et al., 2008; Girona et al., 2009; Attia et al., 2010). The impact on weight of water deficits during the early stage of fruit development is due to changes in water flow originatingmainly in the xylem. An increase in ABA in response to the deficit limits division and cell expansion, mainly in the fruit flesh. This differential response to water deficit has a genetic component, linked to the expression of several proteins identified in the first stage of fruit development. The genetic component determines differences in water-use efficiency, as happens in varieties defined as isohydric - drought sensitive, or anisohydric - resistant to water stress. In our study, in agreement with Chalmers (2007), Cabernet Sauvignon exhibited isohydric behaviour and, in contrast with what is generally considered by others authors, the correlation between Ψ and berry weight was r = -0.95 (p < 0.001); in Merlot, which displayed intermediate behaviour, the correlation was r = -0.92, (p < 0.001) (Chaves et al., 2007), and in Tannat, with anisohydric behaviour, the correlation was r = -0.69 (p = 0.12) (Pereyra & Ferrer, 2013; unpublished data). According to Friend et al. (2009), the increase in berry volume is the result of incorporating water and sugars in the mesocarp, and this process is limited by the expansion of skin cells. Varieties with more efficient water use (anisohydric), like Tannat, would have advantages in increasing the size of their berries over less efficient varieties, like Cabernet Sauvignon. Comparing the year 2004, a dry vintage, with 2001, which was the wettest vintage, it was observed that, under water stress conditions, Cabernet Sauvignon decreased berry weight by 0.33 g, Merlot by 0.26 g and Tannat by 0.04 g. These results show the influence of water availability and variety on variations in berry weight. This statement is also supported by the analysis of water conditions in 2002. Cabernet Sauvignon and Tannat recorded an equal value for ΨPD (-3.80 bar), while the berry weight was significantly lower in Cabernet Sauvignon and the weight reduction in this variety, compared to a year without water restriction (2001), was 0.18 g (11.2%). The reduction in Tannat was 0.08 g. (4.5%). Merlot, showing significantly higher values of ΨPD (-3.17 bars), presented smaller berries and the weight decrease was 0.23 g (13.4%) compared to 2001 (Table 3), in accordance with Ojeda et al., (2001), Ferrer (2007) and Ferrer et al. (2008). This comparison leads to the conclusion that the weight of berries in Tannat is less sensitive to water-limited conditions, in agreement with Attia et al. (2010). This finding is attributed to anisohydric behaviour. The iso- or anisohydric behaviour of the tested varieties can be supported by comparing vegetative growth -estimated through pruning weight - in the wettest year, 2001, with the values shown in the dry year, 2004. The pruning weight in Cabernet Sauvignon was 1.21 kg/plant in 2001 and 0.58 kg/plant in 2004. In Tannat, the same variable amounted to 0.88 kg/plant in 2001 and to 0.80 kg/plant in 2004; and in Merlot it was 0.93 kg/plant in 2001 and 0.86 kg/plant in 2004. Cabernet Sauvignon was the variety most affected by hydric deficit.
Flesh weight
Flesh weight was limited by water stress conditions, with differences depending on variety in accordance with Roby and Matthews (2004) and Chaves et al. (2007). In Merlot and Cabernet Sauvignon, the correlation between the weight of the flesh and ΨPD was -0.90 (p < 0.001) and -0.91 (p < 0.001) respectively. In Tannat the correlation was not significant. Comparing the wettest or wet with dry vintages (2001 vs. 2004), Cabernet Sauvignon lost 0.29 g flesh weight (-21.5%), Merlot lost 0.11 g (-7.3%) and Tannat increased by 0.07 g (+4.8%). The conclusions reported by Roby and Matthews (2004) and Chaves et al. (2007) could not be generalised for all varieties in the sense that, under stress conditions, berry weight loss is due almost exclusively to flesh weight loss. Tannat's behaviour was contrary to expectations, reinforcing the claim of the low sensitivity of berry weight alteration in this variety under water stress conditions.
Seed weight
Seed weight decreased under drought stress, depending on the variety and in accordance with Attia et al. (2010). In Cabernet Sauvignon and Tannat the correlation between seed weight and ΨPD was r -0.99 (p < 0.0001) and -0.95 (p < 0.001), respectively, while in the case of Merlot the correlation was not significant Comparing years with and without water availability (2001 vs. 2004), the seed weight decreased in all three varieties in 2004, with larger weight losses in Cabernet Sauvignon (0.012 g). In this variety, seed was the berry component that suffered the highest proportional weight loss (-22.2%). This differs from the results reported by Roby and Matthews (2004).
Skin weight
Skin weight showed a correlation of r = -0.69 (p = 0.12) with water stress. The analysis in terms of variety showed a correlation in Tannat of r = -0.88 (p = 0.01), in Cabernet Sauvignon of r = -0.78 (p = 0.05), with Merlot being the most sensitive variety, with r = -0.98 (p < 0.001). A comparison of years with different water availability (2001 and 2004) shows that, under conditions of stress (2004), weight loss was 0.028 g in Cabernet Sauvignon, 0.069 g in Merlot and 0.066 g. in Tannat. In the last two varieties, the skin was the component that lost proportionally more weight (-40.8% and -31.6% respectively) (Table 3). These results do not support those reported by Roby and Matthews (2004).
By analysing the change in the weight of the different components of the berry (flesh, seeds and skin) it was established that the three components responded to water deficit conditions and to varietal differences, in accordance with Roby and Matthews (2004) and Attia et al. (2010).
Ratio between skin weight and berry weight
No significant correlation was established between skin weight, berry weight and predawn water potential. The varieties showed different behaviour in response to water stress. The ratio increased in Cabernet Sauvignon (41.7%) due to a loss of berry weight. In Tannat, the decrease (-32.5%) due to weight loss by the skin was the largest relative weight loss among the components of Tannat berries. In Merlot, the relationship also diminished as a result of weight losses in the flesh and skin (-36%), with the last component being the one that lost the highest percentage (Table 3).
Comparing the dry vintage of 2004 with the wettest vintage of 2001, a large variation occurred in the ratio skin weight/berry weight in Tannat, caused by a large weight loss in the skin (-40.8%), while the weight of the berry remained almost unchanged under hydric deficit. This test result shows the different responses of the varieties to the hydric conditions of the year, and the different responses of the components of the berries to these changing climatic conditions (Table 1). Bucchetti et al. (2011) reported that water deficit decreased berry weight in Merlot, but did not inhibit skin growth (fresh weight), and the changes in berry weight did not result in a significant increase in the skin/berry weight. Roby et al. (2004) reported that flesh growth was inhibited more than skin growth in Cabernet Sauvignon, resulting in a greater skin/berry weight ratio. The growth responses may depend on the variety, and on the specifics of timing and severity of water deficits. On the other hand, Ortega-Regules et al. (2008) reported that Merlot showed higher quantities of cell wall material in the skins compared to Cabernet Sauvignon. In the pulp, Merlot berries showed the lowest quantity of cell wall material. These differences in morphology of the cell wall structure (skin and flesh) may partly explain the differences in sensitivity to water deficit of the varieties and the tissues.
Seed weight versus skin weight
No significant correlation was established between seed weight, skin weight and water potential. However, when analysing the varieties separately the correlation was r = -0.99 (p = 0.004) in Merlot; r = -0.68 (p = 0.11) in Tannat, and not significant in the case of Cabernet Sauvignon. Under stress, the ratio rose for the varieties Tannat (+ 34.1%) and Merlot (+ 62.8%), in agreement with the findings of Roby and Matthews (2004) and Attia et al. (2010), while the ratio for Cabernet Sauvignon decreased by -10.7%.
Relationship skin weight and flesh weight Regarding the relationship between skin and flesh weight, and depending on the variety, the following correlations were established with water potential: in Merlot it was r = 0.97 (p = 0.02), in Tannat it was r = 0.76, (p = 0.09) and it was not significant in the case of Cabernet Sauvignon. The relationship skin weight/flesh weight increased in response to stress conditions in Cabernet Sauvignon (+ 8.9%), while it decreased in the varieties Tannat and Merlot (-36.8% and -54.0% respectively). This is consistent with the findings of Chaves et al. (2007) concerning these varieties. In the two relations involving the skin, Cabernet Sauvignon showed the opposite behaviour to the other varieties. This can be explained mainly by the skin component, which increases its proportion, while the flesh and seeds show a larger decrease under conditions of strong water stress. In agreement with the results of Chalmer (2007) and Peryera & Ferrer (unpublished), different behaviour under water stress conditions by different varieties can explain variations in the weight loss of the berries, and this behaviour could be the consequence of their different isohydric and anisohydric behaviour. Cabernet Sauvignon lost more berry weight (-20%) as a result of weight loss in the three components (skin, seeds and flesh). Merlot berries lost weight (-15%) as a consequence of decreases in the flesh and skin, while the weight loss by Tannat berries was the lowest (-2.3%), and was the result of the decrease in seed weight and skin weight. The varieties that lost flesh weight are those that showed greater variability in berry weight, given the relative importance of this component. Moreover, the greater weight of the seeds of Tannat and its more efficient water use under stress may explain its larger berries (Table 3 and Fig. 1).
Composition of berries and their components
Depending on the variety and the conditions in the year there were significant differences in the content and concentration of sugars and the potential and concentration of anthocyanins.
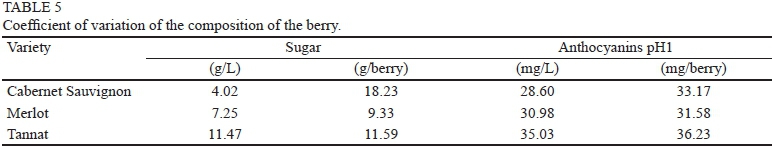
Total sugar content (g/berry) depends on berry weight (r = 0.79, p = 0.11), but independent of berry weight when sugars are expressed in concentration (g/L), in accordance with the statements of Roby et al. (2004) and Matthew and Nuzzo (2007). Tannat, with larger berries, recorded sugar contents (g/L, g/berry or g/g berry) that were higher in three of the four years analysed, in accordance with González-Neves et al. (2012) (Table 4 and Fig. 2). Dry matter production is an indicator of photosynthetic activity. Its distribution among plant organs is an indicator of the source/sink ratio, which has been shown to be dependent on the variety. A significant negative correlation was established between the proportion of dry matter contained in the berries and in the vegetative organs (leaves -0.73, p < 0.001; branch -0.77, p <0.001). This demonstrates existing competition between sinks, in accordance with Edson et al. (1995). Tannat showed a significantly higher proportion of dry matter in the berries (62.2%) compared to the other varieties (Cabernet Sauvignon 49% and Merlot 33.8%). Dry matter was significantly higher to the shoots in Cabernet Sauvignon (36.0%), while in Merlot it was the leaves that contained the highest proportion of dry matter (36.5%). The different allocation of carbohydrates produced during photosynthesis by the different varieties showed that the berry was the most important dry matter sink in the case of Tannat. This result explains the higher sugar content in Tannat berries. Some authors hypothesise this to be due to genetic causes (Edson et al., 1995; Gomez del Campo et al., 2005; Liu et al., 2007; Sadras et al., 2008). The most important variation in sugar concentration between vintages was found in Tannat (cv 11.47, 26.6 g/L), but in Cabernet Sauvignon when expressed in g/berry (cv = 18.23, 0.05 g), while Merlot had the lowest coefficient of variation expressed in g/berry (Table 5). According to Matthews and Nuzzo (2007) and Barbagallo et al. (2011), the flesh is the place where sugars accumulate and it represents the largest proportion of berry weight. Measuring in grams of sugar per berry, a strong and significant positive correlation could be established with flesh weight (r = 0.81, p = 0.07), seed weight (r = 0.79, p = 0.10) and with the relationship seed weight/ skin weight (r = 0.59, p = 0.05). The last two correlations are due to the positive correlation with flesh weight. These results explain the higher content of sugars in the berries of Tannat. The correlation with skin weight and with ratio skin weight/flesh weight was not significant.
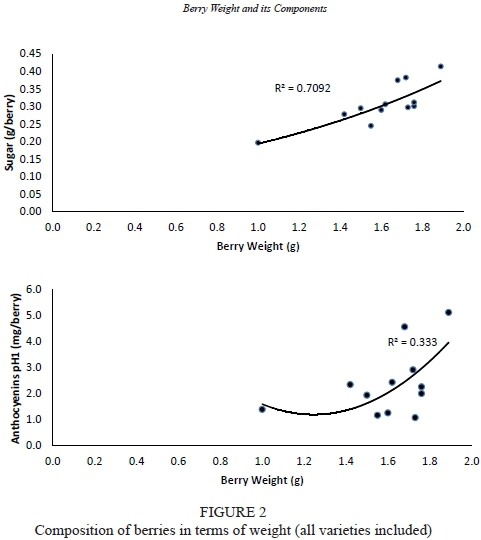
Total anthocyanin content or concentration was independent of berry weight and higher in Tannat berries in all four years of the study. The extreme values between years in Tannat were double to the ones recorded in Cabernet Sauvignon. According to Barbagallo et al. (2011), anthocyanin biosynthesis is not affected by the size of the berry, but by the conditions of the year. This compound showed the greatest variation (Table 4 and Fig. 2). The analysis of the coefficient of variation, expressed in mg/L or mg/berry, is the highest for Tannat. But the differences in coefficient of variation in relation to the other two varieties were lower than the ones recorded for sugars (Table 5). The established correlation between the sugar content per berry and anthocyanin content per berry was r = 0.83 (p = 0.14). These results agree with Hunter et al. (1991), who established that sugar levels in berry skins seemed to be associated with anthocyanin concentration. The skin is the place where anthocyanins are synthesised and accumulate (Coombe, 1989). The weight of the skin in relation to the total anthocyanins or their concentration was dependent on the variety, and was significant and negative for Cabernet Sauvignon (-0.82, p = 0.04) and Tannat (-0.52, p = 0.10), but not significant for Merlot. Comparing the two varieties, skin weight was significantly higher in Cabernet Sauvignon in 2004, while the difference with Tannat regarding anthocyanins was not significant. These results would indicate that skin weight was not a determining factor and that other causes (genetic, environmental, etc.) could be responsible for determining the anthocyanin potential of the berries. This is in accordance with the findings of Mori et al. (2005, 2007), Barbagallo et al. (2011) and Dai et al. (2011). The relationships suggested by some authors (Matthews & Nuzzo, 2007; Attia et al., 2010) to measure the potential anthocyanins, involving the skin (skin weight/berry weight, seed weight/skin weight or skin weight/flesh weight), were not significant.
However, it was possible to establish a significant positive relationship between anthocyanin potential and seed weight of r = 0.69 (p = 0.01). The results would indicate that, in order to explain the contents of anthocyanins from these relationships, the relative weight of the skin had less value than the relative weight of the seeds or flesh. The variety Tannat, which recorded the highest values for anthocyanins, has a larger seed weight/skin weight ratio and heavier seeds.
Variation in the composition of the berry - influence of water availability
Composition of sugars
The dynamics of the accumulation of sugars and water in the berry are closely related and may differ, among other factors, due to variety (Ristic & Iland 2005). The water content in the berries was reported as inversely proportional to the content of sugars by Liu et al. (2007) and confirmed by Sadras et al. (2008). These patterns in water dynamics may differ depending on the variety. The above statements are confirmed by the results of measurements of the percentage of water, estimated as the difference between the fresh weight and dry weight of berries of different varieties. Tannat berries contain 78.93% water, Cabernet Sauvignon 80.92% and Merlot 82.27%. This would agree with the sugar content of the berries from the varieties tested (Table 4).
Correlations were established between sugar content per berry and (Ψ), depending on variety, and were in accordance with Matthews et al. (1990). In Cabernet Sauvignon, that correlation was -0.84 (p = 0.02) and in Tannat it was -0.60 (p = 0.12), whereas it was not significant for Merlot, at -0.19 (p = 0.38). This response corresponds to the dependence on variety of the stress level and anisohydric or isohydric behaviour, as already mentioned. This is in agreement with Castellarin et al. (2007), who reported that the accumulation of sugars in Merlot does not respond to water deficit, but that it does so in Cabernet Sauvignon. The increase in sugar content and its concentration in response to water deficit depended on variety, both in the amount of the increase and in the level of stress. According to Wheeler et al. (2009), this response would be associated with ABA. Under water deficit conditions, abscicic acid promotes increased membrane elasticity and turgor of the cells, thus favouring the entry of sugars into the fruit, sugar movement from the leaves and also sucrose transportation within the plant. Under moderate stress conditions, sugar increases were recorded in all three varieties: in Cabernet Sauvignon it was 11.1%; in Tannat it was 22.2% and in Merlot it was 11.8%. Under strong stress conditions, Cabernet Sauvignon and Tannat kept increasing their sugar contents, while in Merlot the content decreased by 11.7%. The sugar increases recorded in Cabernet Sauvignon, regardless of the level of stress (moderate or severe), are probably explained by an equilibrium in the source/sink relationship regarding vegetative growth being reached, as was shown in our study through the lower pruning weight under limited hydric conditions. The proportion of dry matter directed to the shoots and leaves was 59.6% in 2001 and 45% in 2002, with the carbohydrates produced during photosynthesis being directed to the berries
In Merlot, which has been shown to first serve the leaves with dry matter, a more negative Ψ value could affect photosynthesis, and therefore the production of sugars. The proportion of dry matter directed to the leaves was 35% in 2001 and 27% in 2002.
The sugar content changed according to water availability in the different years. From Fig. 1 it is clear that the content of sugars, related to the average variety/year, showed a lower percentage in years with excess water (2001) for all three varieties. In years with mild or moderate water deficit (2002 and 2003), the differences depend on variety (Table 4, Fig. 1). The lowest sugar values were recorded under conditions of excess water (year 2001). The variety affected the most was Tannat. As a result of the dilution, sugars decreased 14.3% on average across the four years for this variety. Cabernet Sauvignon suffered a 5.2% decrease and Merlot was unaffected. The dilution effect probably was less pronounced in this variety because it was harvested earlier, so it did not suffer the consequence of the rains of the next 20 days. Tannat and Cabernet Sauvignon were affected by this rainy weather. In the period between ripening and harvest in 2001, 425 mm of rain fell on the Merlot variety (harvested on 25 February) and 537 mm on the Cabernet Sauvignon and Tannat varieties (harvested on 15 March).
Ozden et al. (2010) report similar values regarding decreases in total anthocyanins and soluble solids in irrigated vines when irrigation exceeded 436 mm in the period from June to August (HN, Northern Hemisphere).
Anthocyanin composition
Total anthocyanin changed under the influence of the hydrological conditions of the year, and this change was higher than for sugar. Fig. 1 shows that the contents of anthocyanins in the three varieties, based on the average variety/year and measured as percentage, was lower in years with excess water than in years with a strong water deficit (2001 and 2004 respectively). The highest values corresponded to the years with mild or moderate water deficit (2002 and 2003), with differences within the same year depending on the variety (Table 4, Fig. 1). If we compare water conditions in two years with widely different precipitation (2001 and 2004), the results show that the anthocyanin content increased in absolute terms in the three varieties in response to high stress conditions. The magnitude of this increase was different depending on the variety. According to these results, excess water conditions are more negative for anthocyanin contents than strong deficit conditions. Several research studies relate this varietal response to regulation due to genes responsible for the biosynthesis of flavonoids (Castellarin et al., 2007), or the appearance and intensity of a peak of ABA that would be caused by the expression of genes related to the last steps of the biosynthesis of anthocyanins (Wheeler et al., 2009). Cabernet Sauvignon was the variety that recorded the highest increase in anthocyanin concentration (+77.2%), followed by Tannat (+32%) and Merlot (+22.6%). As discussed before, the weight of the skin does not explain this response. Under moderate deficit conditions (year 2002), increased anthocyanin was recorded in all three varieties. Cabernet Sauvignon showed the largest increase (+135.7%), followed by Tannat (+121.1%) and Merlot (+108.1%). Under this moderate deficit situation, the proportion of both berry weight and berry skin decreased as a result of water deficit conditions. According to Roby and Matthews (2004) and Attia et al. (2010), this result would show that the intensity of the deficit plays a key role and that the magnitude of the response is determined by variety. Excess water reduced the concentration of anthocyanins, a reduction that was more significant than the reduction in sugars. This was probably due also not only to rain, but to the effect of increasingly cloudy days with reduced luminosity. The reduction in anthocyanins in 2001, compared to the average of the four years, was 41% in Cabernet Sauvignon, 39.9% in Merlot and 34.7% in Tannat (Fig. 1). Koundouras et al. (2009) also reported that water deficits increased the skin solute concentration without significantly affecting the skin/ berry weight. These relationships indicate that an increase in the relative skin/berry weight is not necessary to realise increased concentrations (mg/g berry FW) of anthocyanins and tannins in harvested fruit as a consequence of water deficits.
Another factor widely reported in the literature is the effect of temperature on the balance of synthesis/degradation of anthocyanins (Tonietto & Carbonneau, 2004; Mori et al., 2005, 2007; Hunter & Bonnardot, 2011). The range of optimal night temperatures during ripening for this balance ranged from 15°C to 16°C. This temperature was 18.8°C in 2001, 16°C in 2002, 15.6°C in 2003 and 14.2°C in 2004. These temperature differences would explain the low content of anthocyanins in the berries in 2001 and 2004.
Depending on the variety, the composition of the grape was affected more than berry growth by water availability. The accumulation of sugars in the grapes of Cabernet Sauvignon and Merlot was influenced less by water deficits than by grape weight, in accordance with Williams and Matthews (1990), while Tannat shows an opposite response. Sugar accumulation (expressed as a percentage) was ten times higher than the change in the weight of the berry. This result agrees with the results reported by Gomez del Campo et al. (2005). These researchers showed that, under stress conditions, the size of the fruit, the accumulation of dry matter and sugar content are affected, but total amounts are dependent on the variety. (Table 3, Fig. 1).
Regarding anthocyanins, the effect of water availability was significantly higher than berry size on their contents, and this parameter was not affected by variety. According to Roby et al. (2004), increased anthocyanin content under water deficit directly stimulates their biosynthesis (Table 3, Fig. 1).
CONCLUSIONS
Berry weight was dependent on the variety and was closely related to the proportions between its components (flesh, seeds and skin). Flesh weight was the most important determinant of berry weight, which in turn was dependent on the weight of the seeds. These two components explain why Tannat berries were heavier. The varieties on trial showed a different response under different weather-dependent hydrological conditions. This different behaviour could explain the variations in and weight loss of berries under conditions of stress. Cabernet Sauvignon berries were the most sensitive (-20%) as a result of the decline in the three components. Merlot berry weight losses (-15%) were the consequence of decreases in flesh weight and skin weight, while Tannat berries were affected the least (-2.3%) as a result of weight loss in the seeds and in the skin. The berries of the varieties in which flesh weight diminished were those that showed greater variation in weight, given the large proportion of this component in the berries. The sugar contents were dependent on berry weight. Anthocyanin content did not depend on the weight of the berry, weight, proportion of the skin on the berry, or their relationships, but was closely associated with the water availability in each year. Tannat berries showed the highest value of anthocyanins and sugars, and also were the variety that showed the greatest variation in these compounds. In Tannat, the berries were the other preferred storage sink for the products of photosynthesis and the larger size of the flesh explains the higher sugar content. Water surpluses and water deficits had a larger influence on berry composition than on berry weight, and these environmental factors had different effects on different varieties. On balance, the present results indicate that the source(s) of variation in berry size are more important in determining composition than size per se.
LITERATURE CITED
Attia, F., Garcia, F., Ben Mariem, F., Nuzzo, V., Dedieu, F., Garcia, M. & Lamaze, T., 2010. Water stress in Tannat and Duras grapevine cultivars (Vitis vinifera l.): Leaf photosynthesis and grape phenolic maturity. J. Int. Sci. Vigne Vin, 81-93. [ Links ]
Barbagallo, M., Guidoni, S. & Hunter, J.J., 2011. Berry size and qualitative characteristics of Vitis vinifera L. cv. Syrah. S. Afr. J. Enol. Vitic. 32(1), 129-136. [ Links ]
Bucchetti, B., Matthews, M.A., Falginella, L., Peterlunger, E. & Castellarin, S.D., 2011. Effect of water deficit on Merlot grape tannins and anthocyanins across four seasons. Scientia Horticulturae 128, 297-305. [ Links ]
Carbonneau, A., Moueix, A., Leclair, N. & Renoux, J., 1991a. Proposition d'une mèthode de prélèvement de raisin à partir de l'analyse de l'hétérogénité de maturation sur un cep. Bull. OIV 727/728, 679-690. [ Links ]
Castellarin, S., Matthews M., Di Gaspero G. & Gambetta, G., 2007. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 227, 101-112. [ Links ]
Chalmers, Y.M., 2007. Influence of sustained deficit irrigation on physiology and phenolic compounds in winegrapes and wine. PhD thesis, Adelaide University, Australia. http://digital.library.adelaide.edu.au/dspace/bitstream/2440/50101/1/02whole.pdf. [ Links ]
Chaves, M., Santos, T.P., Souza, C.R., Ortuño, M.F., Rodrigues, M.L., Lopes, C.M., Maroco, J.P. & Pereira, J.S., 2007. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol. 150, 237-252. [ Links ]
Coombe, B.J., 1989. The grape berry as a sink. Acta Hort. 239, 149-158. [ Links ]
Costantini, L., Battilana, J., Lama, F., Fanizza, G. & Grando, M., 2008. Berry and phenology - related traits in grapevine (Vitis vinifera L.). From quantitative trait loci to underlying genes. Plant Biology. Available www.biomedcentral.com/1471-2229/8/38 [ Links ]
Dai, Z., Ollat, N., Gomés, E., Decroocq, S., Tandonnet, J.P., Bordenave, L., Pieri, P., Hilbert, G., Kappel, Ch., Van Leeuwen, C., Vivin, P. & Delrot, S., 2011. Ecophysiological, genetic, and molecular causes of variation in grape berry weight and composition: A review. Am. J. Enol. Vitic. 62(4), 413-425. [ Links ]
Edson, C.E., Howell, G.S. & Flore, J.A., 1995. Influence of crop load on photosynthesis and dry matter partitioning of Seyval grapevines. III. Seasonal changes in dry matter partitioning, vine morphology, yield, and fruit composition. Am. J. Enol. Vitic. 46(4), 478-485. [ Links ]
Ferrer, M., 2007. Etude du climat des régions viticoles de l'Uruguay des variations climatiques et de l'interaction apportée par le microclimat et l'écophysiologie des systèmes de conduite Espalier et Lyre sur Merlot. Thése Doctorat, Ecole Nationale Supérieure Agronomique - Université de Montpellier II, France. [ Links ]
Ferrer, M., González-Neves, G., Priore, E., Montana, A. & Carbonneau, A., 2008. Los componentes del rendimiento, la expresión vegetativa y los indicadores fisiológicos de Vitis vinifera L. cv. Merlot en función de la arquitectura de la planta. Agrociencia XII(1), 6-19. [ Links ]
Friend, A.P., Trought, M.C.T. & Creasy, G.L., 2009. The influence of seed weight on the development and growth of berries and live green ovaries in Vitis vinifera L. cvs Pinot Noir and Cabernet Sauvignon. Aus. J. Grape Wine Res 15, 166-174. [ Links ]
Girona J., Marsal, J., Mata, M., Del Campo, J. & Basile, B., 2009. Phenological sensitivity of berry growth and composition of Tempranillo grapevines (Vitis vinifera L.) to water stress. Aus. J. Grape Wine Res 15, 268-277. [ Links ]
Glories, Y. & Augustin, M., 1995. Maturité phénolique du raisin, conséquences technologiques: application aux millésimes 1991 et 1992. Proceedings of the Journée Technique. CIVB, Bordeaux - France (Conseil interprofessionnel du vin de Bordeaux) pp 56-61. [ Links ]
Gomez del Campo, M., Baeza, P., Ruiz, C. & Lissarrague, J.R., 2005. Effects of water stress on dry matter content and partitioning in four grapevine cultivars (Vitis vinifera L.). J. Int. Sci. Vigne Vin 39(1), 1-10. [ Links ]
González-Neves, G., Charamelo, D., Balado, J., Barreiro, L., Bochicchio, R., Gatto, G., Gil, G., Tessore, A., Carbonneau, A. & Moutounet, M., 2004. Phenolic potential of Tannat, Cabernet-Sauvignon and Merlot grapes and their correspondence with wine composition. Analytica Chimica Acta 513 (1), 191-196. [ Links ]
González-Neves, G., Ferrer, M. & Gil, G., 2012. Differentiation of Tannat, Cabernet Sauvignon and Merlot grapes from Uruguay according to their general composition and polyphenolic potential. Comunicata Scientiae 3(1), 41-49. [ Links ]
Gray, J. & Coombe, B.G., 2009. Variation in Shiraz berry size originates before fruitset but harvest is a point of resynchronisation for berry development after flowering. Aus. J. Grape Wine Res. 15, 156-165. [ Links ]
Holt, H.E., Francis, L., Field, J., Herderich, M.J. & Iland, P.G., 2008. Relationships between berry size, berry phenolic composition and wine quality scores for Cabernet Sauvignon (Vitis vinifera L.) from different pruning treatments and different vintages Aus. J. Grape Wine Res. 14, 191202. [ Links ]
Hunter, J.J. & Bonnardot, V., 2011. Suitability of some climatic parameters for grapevine cultivation in South Africa, with focus on key physiological processes. S. Afr. J. Enol. Vitic. 32(1), 137-154. [ Links ]
Hunter J.J., De Villiers, O.T. & Watts J.E., 1991. The effect of partial defoliation on quality characteristics of Vitis vinifera L. cv. Cabernet Sauvignon grapes. II. Skin color, skin sugar, and wine quality. Am. J. Enol. Vitic. 42(1), 13-18. [ Links ]
Koundouras, S., Hatzidimitriou, E., Karamolegkou, M., Dimopoulou, E., Kallithraka, S., Tsialtas, J.T., Zioziou, E., Nikolaou, N. & Kotseridis, Y., 2009. Irrigation and rootstock effects on the phenolic concentration and aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. J. Agric. Food Chem. 57, 7805-7813. [ Links ]
Liu, H.-F., Wu, B.-H., Fan, P.G., Xu, H.-Y. & Li, S.-H., 2007. Inheritance of sugars and acids in berries of grape (Vitis vinifera L.). Euphytica 153, 99-107. [ Links ]
Mathews, M.A. & Anderson, M.M., 1988. Fruit ripening in Vitis vinifera. Responses to seasonal water deficits. Am. J. Enol. Vitic. 39 (4), 313-320. [ Links ]
Matthews, M.A. & Nuzzo, V., 2007. Berry size and yield paradigms on grapes and wines quality. In: Nuzzo, V (ed). Proc. Intl. WS on Grapevine, Venosa, Italy, 31 October 2007. Acta Hort. 754. [ Links ]
Mori, K., Goto-Yamamoto, N., Kitayama, M. & Hashizume, K., 2007. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot. 58(8), 1935-1945. [ Links ]
Mori, K, Sugaya, S. & Gemma, H., 2005. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic. 105, 319-330. [ Links ]
Nicolas, P., Lecourieux, D., Gomès, E., Delrot, S. & Lecourieux, F., 2013. The grape berry-specific basic helix-loop-helix transcription factor VvCEB1 affects cell size. J. Exp. Bot. Available http://jxb.oxfordjournals.org/open_access.html [ Links ]
O.I.V., 2007. Récueil des méthodes internationales d'analyse des vins et des moûts. Office International de la Vigne et du Vin. Paris, 368 pp. [ Links ]
Ojeda, H., Deloire, A. & Carbonneau, A., 2001. Influence of water deficits on grape berry growth. Vitis 40(3), 141-145. [ Links ]
Ortega-Regules, A., Romero-Cascales, J., Ros Garcia, J., Bautista-Ortín, A., López-Roca, M., Fernández-Fernández, J. & Gómez-Plaza, E., 2008. Anthocyanins and tannins in four grape varieties (Vitis vinifera L.). Evolution of their content and extractability. J. Int. Sci. Vigne Vin 42 (3), 147-156. [ Links ]
Ozden, M., Vardin M., Simsek, H. & Karaaslan, M., 2010. Effects of rootstocks and irrigation levels on grape quality of Vitis vinifera L. cv. Shiraz. Afr. J. Biotechnol. 9(25), 3801-3807. [ Links ]
Ribéreau-Gayon, P. & Stonestreet, E., 1965. Le dosage des anthosyanes dans le vin rouge. Bulletin de la Société Chimique. 9, 2649-2652. [ Links ]
Ristic, R. & Iland, P., 2005. Relationships between seed and berry development of Vitis vinifera L. cv. Shiraz: Developmental changes in seed morphology and phenolic composition. Aus. J. Grape and Wine Res. 11, 43-58. [ Links ]
Roby, G. & Matthews, M., 2004. Relative proportions of seed, skin and flesh, in ripe berries from Cabernet Sauvignon grapevines grown in a vineyard either well irrigated or under water deficit. Aus. J. Grape & Wine Res. 10, 74-82. [ Links ]
Roby, G., Harbertson, J.F., Douglas, A.A. & Matthews, M.A., 2004. Berry size and vine water deficits as factors in wine grape composition: Anthocyanins and tannins. Aus. J. Grape Wine Res. 10, 100-107. [ Links ]
Sadras, V.O., Collins, M. & Soar, C.J., 2008. Modelling variety-dependent dynamics of soluble solids and water in berries of Vitis vinifera. Aus. J. Grape Wine Res. 14, 250-259. [ Links ]
Santesteban, L. & Royo, B., 2006. Water status, leaf area and fruit load influence on berry weight and sugar accumulation of cv. 'Tempranillo' under semiarid conditions Scientia Horticulturae 109, 60-65. [ Links ]
Tonietto, J. & Carbonneau, A., 2004. A multicriteria climatic classification system for grape-growing regions worldwide. Agric. Fort. Meto. 124, 8197. [ Links ]
Vila, H., Paladino, S., Nazrala, J. & Lucero, C., 2010. Manual de calidad de uva. Guia práctica para conocer y evaluar la calidad de la uva para vino. Ediciones INTA, Mendoza, Argentina. [ Links ]
Walker, R.R., Blackmore, D.H., Clingeleffer, P.R., Kerridge, G.H., Rühl, E.H. & Nicholas, P.R., 2005. Shiraz berry size in relation to seed number and implications for juice and wine composition. Aus. J. Grape and Wine Res. 11, 2-8. [ Links ]
Wheeler, S., Loveys, B., Ford, C. & Davies, C., 2009. The relationship between the expression of abscisic acid biosynthesis genes, accumulation of abscisic acid and the promotion of Vitis vinifera L. berry ripening by abscisic acid. Aus. J. Grape Wine Res. 15, 195-204. [ Links ]
Williams, L.E. & Matthews, M.A., 1990. Grapevine. In: Stewart, B.A. & Nielson, D.R. (eds). Irrigation of agricultural crops. Agronomy Monograph No. 30. ASA-CSSA-SSSA, Madison, WI, pp 1019-1059. [ Links ]
Submitted for publication: September 2013
Accepted for publication: December 2013
Aknowledgements: The authors gratefully acknowledge the technical assistance of the personnel of the Viticulture section, as well as financial support from the wine industry. Prof G. González-Neves and the Faculty of Agronomy are thanked for their support during the study periods
* Corresponding author: e-mail address: mferrer@fagro.edu.uy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}