










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.49 n.3 Pretoria Jul. 2023
http://dx.doi.org/10.17159/wsa/2023.v49.i3.4023
RESEARCH PAPER
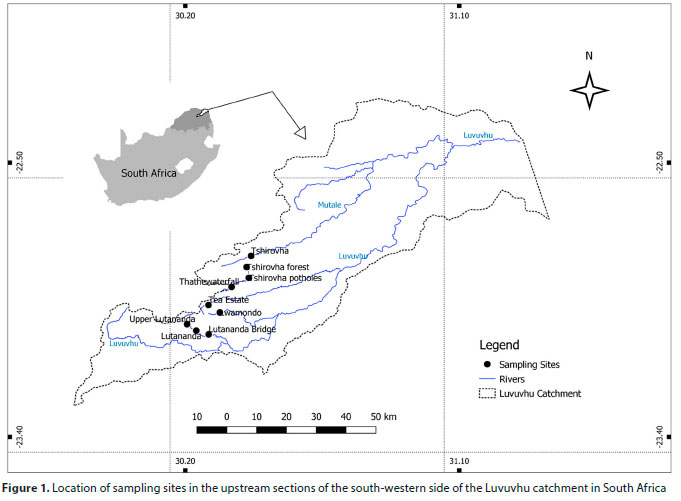
Relationships between reference site quality and baetid mayfly assemblages in mountainous streams of the Luvuvhu catchment, South Africa
Pfananani RamulifhoI; Nick Rivers-MooreII, III; Stefan FoordIV
IDepartment of Environmental Sciences, Florida Science Campus, University of South Africa, South Africa
IICentre for Water Resources Research, University of KwaZulu-Natal, Pietermaritzburg, South Africa
IIIFreshwater Research Centre, Cape Town, South Africa
IVChair in Biodiversity Value and Change in the Vhembe Biosphere Reserve, Department of Zoology, University of Venda, Thohoyandou, South Africa
ABSTRACT
With water quality deteriorating rapidly at a global scale, river sections suited to serve as reference sites are being increasingly lost. It thus becomes critical to develop rapid methods to confirm that previously monitored sites continue meet the requirements of reliable reference sites. In the absence of pristine sites, 9 near-natural sites, as defined by the Kleynhans (1996) classification, were used as reference sites for the Luvuvhu River catchment to compare the quality of physico-chemical factors against a biological metric. Baetid mayfly community structure at a site was chosen as an index of water quality, since this family is common in all types of freshwaters, highly diverse and adapted to unpolluted running water. Baetid larvae were sampled monthly from stones-in-current biotopes across 9 sites for over 1 year, between December 2016 and January 2018. A Spearman's correlation test was used to evaluate the relationship between physico-chemical factors and identify redundant variables. Water quality standards were measured against the national water quality guidelines for aquatic ecosystems. We used a generalized linear model to determine the effect of physico-chemical variables on baetid species, and canonical correspondence analysis to show the relationships between baetid species, sites, and physico-chemical variables. A total of 3 039 individuals belonging to 12 mayfly species were recorded. Our findings indicated that while the physico-chemical factors were highly variable, they were within favourable ranges to reflect reference site conditions. While water temperature was the most important driver of baetid community structure in general, as it negatively affected their abundances, a subset of species (Pseudoponnota sp., Pseudocloeon sp., Acanthiops varius and Demoulinia crassi) showed clear responses to changes in TDS and stream width. We conclude that specific baetid species show good potential as biological indicators of reference sites and chronic water temperature stress, making assessment of reference sites easier.
Keywords: bioindicator, ecological integrity, mayflies, reference condition, water temperature
INTRODUCTION
Despite the recognized importance of rivers in providing critical services to both humans and natural organisms, their water quality is deteriorating at an alarming rate due to human activities (Tampo et al., 2020). Worldwide, the quality ofwater in rivers is increasingly threatened (Dudgeon et al., 2006), most specifically those in developing countries, due to industrialization, urbanization processes, and constant changes in land uses (López-López and Sedeno-Díaz, 2015). The quality of river and stream water is very sensitive to anthropogenic influences (urban, industrial and agricultural activities, and increasing consumption of water resources), as well as natural processes like soil erosion and weathering of the earth's crustal material (Croijmans et al., 2020; Rashid and Romshoo, 2013; Hamid et al., 2020). In South Africa, extensive efforts of monitoring both the ecological and water quality conditions of rivers using nationally approved indices (e.g., River Eco-status Monitoring Programme formerly known as the River Health Programme, also the Rapid Habitat Assessment Methods and Models, etc.) is the responsibility of the Resource Quality Information Services Directorate of the national Department of Water and Sanitation (DWS). In the 60-year long records, time-series data show a growing deterioration of water quality that needs to be addressed more vigorously (Pitman, 2011). This is also reflected in the most recent national ecosystems and biodiversity status report, which indicated that the condition of natural river ecosystems has declined by 11% between 1999 and 2011 (Skowno et al., 2019). From the 222 stream ecosystems assessed in South Africa, 64% were found to be threatened and 43% among them were critically endangered. Similarly, in some developed countries, such as Australia, the United States of America, and some European countries, the monitoring of streams is a government obligation (López-López and Sedeno-Díaz, 2015; Couceiro et al., 2012). Sustained action needs to be taken worldwide to prevent further deterioration of rivers, failure of which might pose a health risk to aquatic life and people.
A key issue in the management and biomonitoring of aquatic systems is the establishment of reference conditions against which to assess change and ecological trends over time (McDowell at al., 2013). A practical definition of 'reference condition' is the chemical, physical and biological conditions that can be expected in streams and rivers with minimal or no anthropogenic influence (Soranno et al., 2011). Reference condition provides a baseline from which to compare changes in water quality parameters and biological composition. There is a range of methods used to estimate reference conditions, as mentioned in McDowell at al. (2013). However, in all the methods, the biological community of a stressed or disturbed ecosystem is compared with that of relatively undisturbed reference sites that have similar environmental conditions, when assessing the impact of disturbance in multiple sites (Kaboré et al., 2018). If the test-site community differs from the reference condition site, the conclusion can be drawn that the site is impacted (Reece and Reynoldson, 2001). Stream sections that are best suited to serve as reference condition are increasingly challenging to locate because of increasingly widespread anthropogenic impacts across catchments (Soranno et al., 2011).
Many studies make ecological inferences based on the degree of water quality as reflected by the presence or absence of aquatic organisms (Aazami et al., 2015; Varnosfaderany et al., 2010; Venkatesharaju et al., 2010; Beyene et al., 2009; Sharma and Rawat, 2009). There are several good reasons why macroinvertebrates are useful as indicators of the reference conditions of rivers. These reasons include their persistence across seasons, their species diversity, and ubiquitous occurrence in almost all types of the world's freshwater ecosystems (Buss and Salles, 2007). Amongst the macroinvertebrate taxa found in the tropics and the southern hemisphere, baetidae are more endemic and show more important adaptation traits to local afrotropical conditions than others (Barber-James et al., 2008; Gattolliat and Nieto, 2009). Several studies have demonstrated that baetid community structure reflects the environmental state of rivers effectively (Kubendran et al., 2017; Buss and Salles, 2007; Bauernfeind and Moog, 2000). Mayflies are characterized by narrow habitat tolerance and only occur in very clean freshwater, which makes them good bioindicators for very good water quality (Alhejoj et al., 2023; Buss and Salles, 2007; Kubendran et al., 2017).
The interactions between environmental factors and baetid abundances is crucial since this nexus has potential to enhance ecosystem services that baetid species provide. Available evidence shows that they provide many essential services that maintain and enhance ecosystem function, such as energy flow dynamics (Boyero et al., 2011; Jacobus et al., 2019). Some baetid species are good manipulators of organic matter like periphyton and sediment (Buss and Salles, 2007; Baptista et al., 2006). Baetids process large amounts of organic matter, allochthonous carbon and nutrients from riparian vegetation and soil materials (Moulton et al., 2004), which are used by organisms at higher trophic levels (Wallace and Webster, 1996; Boyero et al., 2011). According to Wallace and Webster (1996), most baetid species are generally primary prey for invertebrate predators and they also contribute in various ways to energy flow and nutrient cycling. Some filter feeders of mayflies (including most baetid species) contribute to water purification and are part of arguably the most important of these predator-prey relationships - as the diet of fish - which is also a driver of the domestic food and local economy (Jacobus et al., 2019).
To our knowledge, no exclusive studies have been undertaken on the relationship between reference conditions based on physico-chemical parameters and baetid community composition. The similarities of baetid composition between reference condition sites have not been explored and it is unknown if the physico-chemical composition differs across these mountainous rivers. Furthermore, despite the widespread occurrence of this mayfly family, it is still unclear if this family is influenced by physico-chemical parameters at these sites. We ask the question of whether the value of sites to still act as reference sites can be quantified using baetid species. In this survey, we used correlation models to assess the degree of similarity between physico-chemical characteristics and baetid species composition at reference sites. Our objective was to quantify the relative role of physico-chemical factors in structuring baetid species assemblages, using the Luvuvhu River catchment as a case study.
METHODOLOGY
Study area
The study was conducted in the south-eastern streams of the Soutpansberg Mountains, Limpopo Province, South Africa. Nine sampling sites were selected along four streams (Dzindi, Mutshundudi, Lutanandwa and Tshirovha), all of which are major streams of the Luvuvhu River catchment. These sites are located in the uppermost 5 km stream segment within their respective streams, and they hold both an instream and riparian zone habitat integrity of 60% and 90%, respectively (Kleynhans, 1996). These streams have continuous flow of water throughout the year during both the dry and rainy seasons. All sites showed high similarities in their physical characteristics and biotopes and were in the foothill zone (Rowntree and Wadeson, 1999), with stream orders of 1 and at elevations of 622-1 022 m amsl. (Fig. 1). The catchment experiences wet summers from October to April with peak rainfall in January and February. The mean annual precipitation is 608 mm, while the mean annual air temperatures are 17°C in mountainous areas and 24°C near the Kruger National Park (Singo et al., 2012). The width of the active channel of the sampled sites ranged from 3.45 m at Thathe waterfall to 11.42 m at Tshirovha. These sites are near-natural, with intact vegetation cover and very little to no human impact. Based on the habitat integrity assessment of Kleynhans (1996), these sites have limited indigenous vegetation removal, little exotic vegetation encroachment and water abstraction. Sites were chosen to represent a pristine gradient of physico-chemical and environmental conditions and macroinvertebrate community assemblages.
Biological and physico-chemical sampling
All biological samples were taken from the 'stones-in-current' hydraulic biotope because sub-imagos (nymphs) of many baetid species inhabit this riffle section of streams and river (Bauernfeind and Moog, 2000). These sites were all well aerated (Fig. 2) and provide a home to a variety of macroinvertebrate organisms (Ramulifho et al., 2020). Each site was sampled on a monthly basis from December of 2016 to January of 2018. All sites were sampled within the same single week at daylight during each sampling month to allow for consistency in weather and flow conditions across sites. Six stones containing organisms were sampled at each site using a standard SASS net. All contents from a net were emptied into a sample bottle and sorted in the laboratory, and baetid larvae were then identified. Most of the material was identified to species level, while some early instar larvae were only identified to the genus or morpho-species level using taxonomic keys (De Moor et al., 2003). Specimens were preserved in 70% ethanol and are housed at the reference collection section of the SARCHi offices, University of Venda.
At each site, one measurement of four physico-chemical parameters (water temperature, conductivity, total dissolved solids (TDS), pH) was taken using a portable pH/EC/TDS/ temperature multi-meter. Other variables (environmental) whose single reading was measured include elevation, habitat area (stone size), flow depth, stream width, and flow velocity rate using a Flow Globe FP101 reader.
Data analysis
The baetid composition was analysed using species richness and abundance metrics. Baetid species abundance data taken from 6 sampling stones at each site was pooled to then represent a single monthly abundance sample at each site. Biological data were tested for normality using the normal quantile-quantile (Q-Q) test which showed that the data were normally distributed.
Physico-chemical data was standardized using log10 (x+1)to achieve the assumed conditions of normality and homoscedasticity (Buss and Salles, 2007), while no species in the biological data was down-weighted. Standardization of physico-chemical data was necessary to reduce the influence of large differences and double zeros, to normalize and render data homoscedastic (Clarke and Gorley, 2006).
To avoid multi-colinearity between physicochemical variables, we calculated the non-parametric rank-based Spearman correlation between these variables. The departure from reference condition of physicochemical variables was made by comparison with the corresponding standards prescribed for aquatic ecosystems in South Africa (DWAF, 1996). Water temperature should not vary (standard deviate) from the mean temperature for that specific site by > 2°C, while the mean TDS should not vary (standard deviate) by > 15% (DWAF, 1996). Most freshwaters in South Africa are relatively well buffered and more or less neutral, with pH ranges between 6 and 8, and pH should not vary (standard deviate) from the mean values for a specific site by > 0.5 (DWAF, 1996). Where no specific reference condition criteria were prescribed by national water quality guidelines (e.g. EC concentrations), peer-reviewed reports and articles published from areas of similar geographic or climatic region to this study with such specification were used.
We used generalized linear mixed model (GLMM) with negative binomial regression from the 'MASS' package and 'glmer.nb' function (Nakagawa and Schielzeth, 2013; Jamil and Ter Braak, 2013) to evaluate the relative importance of each physico-chemical variable on the abundance of baetidae species from all nine sites combined. GLMM is an extension to the generalized linear model (GLM) in which the linear predictor contains random effects in addition to the usual fixed effects (Venables and Ripley, 2002). Since no collinearity existed between physico-chemical variables, we ran one full model of GLMM with all physico-chemical variables. During the analyses, sites were used as a random factor to account for temporal pseudo-replication, while all physico-chemical variables were included as fixed variables (Li et al., 2018). The goodness-of-fit of the models was assessed using the relations between the residuals (the differences between observations and predictions by the retained model) and physico-chemical variables. We also determined the correspondence of physico-chemical variables, sites and baetid species during the sampling period, using the forward addition of correspondence variables technique in canonical correspondence analysis (CCA) (Ter Braak and Verdonschot, 1995). The statistical significance of each variable selected in CCA was judged using a Monte-Carlo permutation test (Klonowska-Olejnik and Skalski, 2014). All the statistical analyses were performed in R (R Core Team, 2022).
RESULTS
Sites condition and correlation between physico-chemical variables
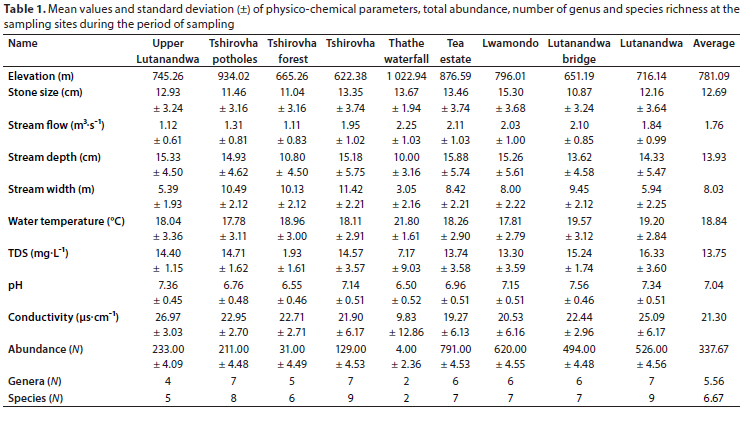
The sampling sites (Table 1) had a mean elevation of 781.09 m, with the highest at 1 022.94 m and the lowest at 622.38 m. Conductivity ranged from 1.9 to 42 μs cm-1, averaging at 21.30 μs·cm-1. Water temperatures ranged from 13.7 to 26.3°C, with a mean of 18.84°C. TDS ranged from 2.96 to 31.9 mg-L-1 with 6 of the 9 sites fluctuating by over 15% of a mean of 13.57 mg-L-1. The mean pH value of these sites ranged between 6.50 and 7.56, with some (55% of sites) having a change (standard deviation) of > 0.5 over time. Stream flow velocities ranged from 0.2 to 6.4 m-s-1, with a mean of 1.76 m-s-1. Mean stream flow depth was 13.93 cm and was highly variable, ranging between 2 and 48.5 cm. Stream width varied between 2.1 and 20 m, with a mean of 8.03 m. Stones had a mean size of 12.69 cm and ranged from 4.66 to 46 cm.
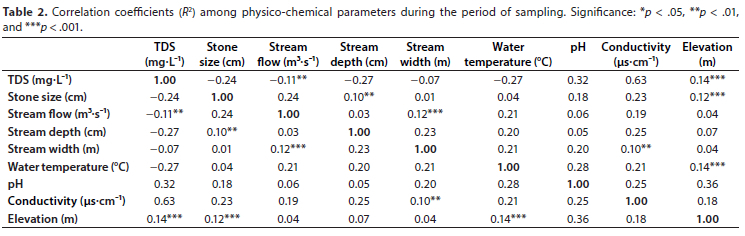
No correlation between physico-chemical variables was > 0.7 (Table 2). More than 85% of these variables were positively correlated. As expected, highest positive correlation was found between conductivity and TDS (R2 = 0.63), since these two water quality parameters are related and are used to describe salinity levels in water. There were highly significant correlations (p < 0.001) between elevation and three variables (TDS, stone size and water temperature; R2 < 0.15). Similarly, stream flow and stream width were significantly correlated with R2 < 0.15. Some significant correlations (p < 0.01) that were also observed included those between stream flow and TDS, stream depth and stone size, and between conductivity and stream width. The lowest negative correlation had a coefficient of -0.27 and was between TDS and stream depth, and between TDS and water temperature.
Abundance of baetids and effect of physico-chemical factors
A total of 3 039 individuals of baetidae belonging to 9 genera and 12 species were recorded in this study. The highest number of individuals caught was 28 specimens at Lwamondo (during low-flow period), while the average catch across the sites was 6 specimens. The highest number of baetid species was recorded at mid-Lutanandwa and the Tshirovha confluence, with 9 species (Table 1). The tea estate site had the highest number of individuals (791), and together with Lutanandwa bridge and Lwamondo sites had the third highest diversity, with 7 species after Tshirovha potholes (Table 1). The lowest number of species was recorded at Thathe waterfall, with only 2 species. The most abundant species in the streams were Baetis Harissoni and Dabulamanzia media, and these were also the most widespread species, occurring at 8 of the 9 sites. Centroptiloides bifasciata and Demoulinia crassi were each limited to 1 site, with low numbers of individuals at Lwamondo and Upper Lutanandwa (3 and 4, respectively).
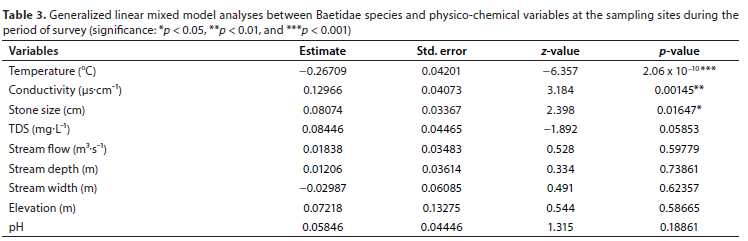
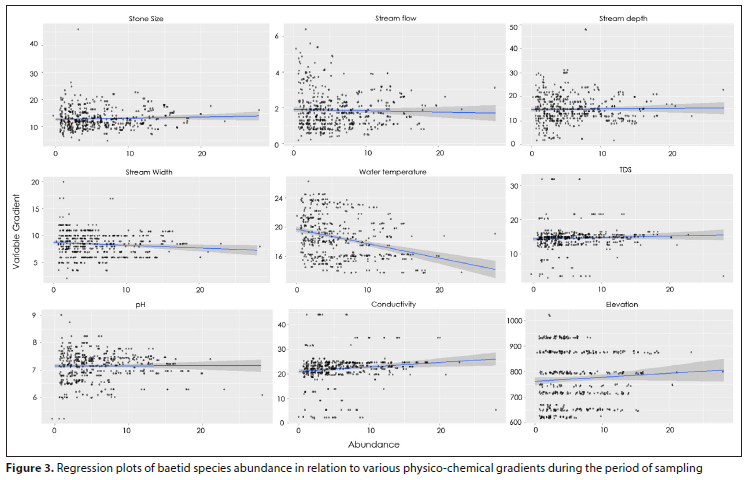
The results suggest that, overall, the most significant drivers of baetid species abundance were water temperature (GLMM: estimate = -0.26, p < 0.001), followed by conductivity (GLMM: estimate = 0.12, p < 0.01) and stone size (GLMM: estimate = 0.08, p < 0.05) (Table 3). An increase in water temperature negatively affected the abundance of baetid species, as opposed to an increase of both conductivity and stone size, which had a positive population effect (Fig. 3). Non-significant drivers of baetid abundance included stream flow, stream width, stream depth, TDS, pH, and elevation. Amongst all these drivers, only stream width increased with a decrease in baetid species abundance (Fig. 3).
Site preference of baetid species
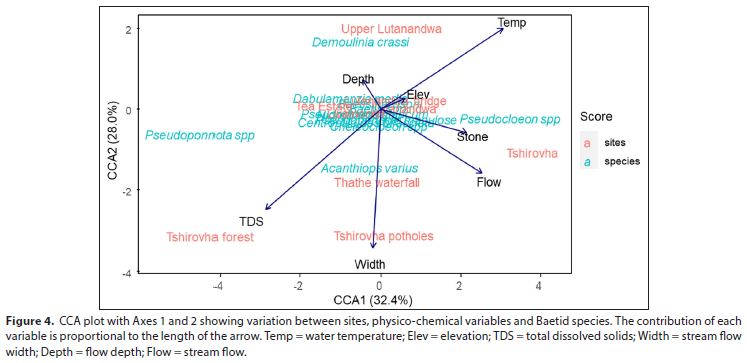
The percentage ofvariance explained by Axes 1-4 of the canonical correspondence analysis amounted to 76.88%, with the 1st and 2nd axes explaining 32.4% and 28%, while only a small variation of 9.56% and 6.92% of the total variance was explained by the 3rd and 4th axes, respectively. According to the canonical correspondence analysis, there are two distinct patterns of baetid species preferences for sites. The first correspondence component (CA1) represented a gradient where most physico-chemical variables (temperature, elevation, stone size, stream flow) load strongly positively on this component, while TDS and stream flow depth load negatively. This positive trend of physico-chemical variables is associated with the Tshirovha site, and Pseudocloeon sp. CA1 is also associated with a decrease in TDS which is closely linked to the decline in abundance of Pseudoponnota sp. at Tshirovha forest site. CA2 showed a prominent negative loading of physico-chemical variables such as TDS, stream width, stream flow and stone size, associated with Tshirovha, Thathe waterfall, Tshirovha potholes, and Tshirovha forest. Further observation showed that Acanthiops varius is negatively affected, while Demoulinia crassi abundance increases with an increase in temperature, elevation and stream depth, and is more closely related to the Upper Lutanandwa site. The majority of baetid species (8 of 12 species or 66.66%) showed no clear response to changes in physico-chemical parameters in the study area (Figure 4). These species are clustered between the nine sampling sites.
DISCUSSION
Physico-chemical variables and baetid community composition
Strong positive correlations between physico-chemical variables which were highly significant (e.g., elevation to TDS, stone size and water temperature) were observed in this study at numerous sites (Table 2). This was expected from sampling sites which are seemingly influenced by both closely related sources (the Soutpansberg mountains) and land uses, as shown in Abowei (2010). This low variation within physico-chemical variables was characteristic of all sampling sites. The absence of industrial activities at these sites is evident by pH levels for sampled sites which were all within the recommended South African aquatic system pH range of 6-8 (DWAF, 1996), and also as observed by Monyai et al. (2016). Acidic effluents from industrial activities are known to cause low pH levels in rivers (e.g., mine drainage, paper, tanning and leather industries). The visual evidence from stream water showed no black or brown (tea-coloured) water or any filamentous algae (Fig. 2), which is usually caused by changing pH levels. The water temperature standard for sustaining aquatic life is 20-30°C (Weldemariam, 2013). This study was dominated by sites with relatively lower temperature range within the accepted thresholds (Table 1). This could be due to forest cover at the sites, which reduces light incidence keeping the stream water temperature at low values (Siegloch et al., 2014; Klonowska-Olejnik and Skalski, 2014), and the effect of altitude The concentration of TDS and EC at the nine sites was well within the WHO standard for inland surface water of 1 000 mg-L-1 and 300 μs·cm-1 (WHO, 2011). This was expected at all these sites due to the absence of practices such as enrichment by soaps and detergents from people washing or bathing in streams, which would result in high levels of TDS and EC, placing stress on aquatic species (Monyai et al., 2016). This might also mean the absence of land-use practices such as overgrazing, non-contour ploughing, removal of riparian vegetation and forestry operations adjacent to these sites. These practices accelerate erosion or result in increased loads of suspended solids in rivers (Monyai et al., 2016; Adu and Oyeniyi, 2019). Most ofthe environmental variables recorded in this study were within levels prescribed by DWAF and WHO and should be able to support aquatic life (DWAF, 1996; WHO, 2011; Weldemariam, 2013).
It is evident from this study that not all species have the same response to environmental parameters (Table 3 and Fig. 4). Water temperature explained the most significant amount of variation in relative abundance, as has been reported in other studies (Ramulifho et al., 2020; Buss and Salles, 2007; Jacobus et al., 2019; Adu and Oyeniyi, 2019; Bauernfeind and Moog, 2000). Water temperature in these sites is driven by riparian vegetation. Riparian vegetation is vital for maintaining and ensuring suitable water temperature and the amount of light available as it also forms a buffer area for the stream. A similar study of undisturbed sites by Klonowska-Olejnik and Skalski (2014) found that the intactness of riparian vegetation is one of the most important factors structuring communities. Another similar study of upper catchment sites by Svitok (2006) concluded that mayfly abundance was most strongly related to elevation, which also relates to the climatic variable of air temperature. The findings of this signal important concerns regarding potential species movement and survival in the face of climate change predictions.
Eight species of baetid did not show any conclusive response to physico-chemical variables and sites. Only a few species were associated with specific sampling sites (Fig. 4). Pseudoponnota sp., Pseudocloeon sp., and A. varius showed a considerable degree of preference for TDS, temperature and stream width linked to Tshirovha potholes, Tshirovha forest, and Thathe waterfall, which are sites found in close proximity each other and on the same stream. Ubiquitous species such as Baetis harissoni and Dabulamanzia media showed no preference for measured conditions in streams. These species are generalist in their nature in the Luvuvhu catchment as they tolerate a range of conditions (Ramulifho et al., 2020). Studies globally have largely used different species of baetids as valid biological indicators of water quality because they are highly sensitive to substrate changes (Kubendran et al., 2017; Buss and Salles, 2007; Bauernfeind and Moog, 2000). In this study, it is evident that baetids showed varied tolerance levels to pollution, but generally are considered intolerant organisms and require water of good quality to survive, as also shown by Alhejoj et al. (2014).
Benefits of baetids in confirming reference site quality
Results from this study further enhance the use of baetid species as a low-cost indicator for aquatic reference sites that allow quick, widespread, long-term, routine monitoring and direct comparison of sites, time periods and studies (Butana et al., 2010). The use of natural variation of baetid species in reference conditions also helps to avoid the setting of quantitative limits or targets of physico-chemical factors (of reference sites) that are either too restrictive or impossible to meet in the face of changing land use and rapid industrialization (McDowell et al., 2013). Thus, even in areas where there is a deficiency of physico-chemical and environmental data, by using baetid species there is still a possibility that reference conditions may well be established. This biological approach to reference site selection enables the measurement of a natural continuum of the substantial benefits of baetid species ecosystem processes, such as nutrient cycling, algal distribution, retention and distribution of organic matter, and predator-prey interactions (Jacobus et al., 2019; Sartori and Brittain, 2015; Wallace and Webster, 1996).
The usefulness of this research lies in its contribution towards closing an existing gap on a biological index of baetids species in reference sites in South Africa. This research establishes preliminary baseline biological characteristics of the potential reference sites in mountain rivers, as opposed to widely used selection criteria like chemical and physical (i.e., abiotic) factors as surrogates (Agboola et al., 2020). The biological indices are widely recommended and a valuable tool in monitoring macroinvertebrate response, reference conditions and anthropogenic disturbances in rivers in many regions including Europe (Lewin et al., 2013) and west Africa (Kaboré et al., 2018). If this is adopted for local streams, accurate estimation of reference conditions based on biological indices will provide information on anthropogenic impacts and stress for sites in upstream catchments and potential areas for restoration of reference conditions (McDowell et al., 2013).
CONCLUSIONS
Our findings indicated that the physico-chemical factors at the selected sites are highly variable but are still in a favourable range for reference site conditions. Direct effects of measured physico-chemical factors on the entire baetid community were evident largely for Pseudoponnota sp., Pseudocloeon sp., Demoulinia crassi and Acanthiops varius. Since the presence or absence of certain mayflies was strongly influenced by water temperature, TDS, and stream width (as observed from the models), this study confirmed that these species are a powerful tool as descriptors of reference sites. These results are of relevance for protection of these species and reference sites in catchments in South Africa.
ACKNOWLEDGMENTS
This study was supported financially and logistically by the National Research Foundation and the Department of Science and Technology through the South African Research Chairs Initiative (SARChI) Chair on Biodiversity Value and Change in the Vhembe Biosphere Reserve, hosted and supported by the University of Venda. We thank the Department of Zoology, University of Venda, for providing laboratory and field work facilities.
DATA AVAILABILITY STATEMENT
Data, models or codes that support the findings of this study are available from the corresponding author upon request.
AUTHOR CONTRIBUTIONS
Pfananani Ramulifho conceived the study, wrote the initial draft of the manuscript and performed data analyses. Nick Rivers-Moore and Stefan Foord edited and commented on the manuscript. All authors contributed to discussions that shaped the manuscript.
CONFLICT OF INTEREST
The authors declare no conflict of interest.
ORCIDS
Pfananani Ramulifho: https://orcid.org/0000-0002-1589-7899
Nick Rivers-Moore: https://orcid.org/0000-0002-6546-4215
Stefan Foord: https://orcid.org/0000-0002-9195-2562
REFERENCES
AAZAMI J, ESMAILI-SARI A, ABDOLI A, SOHRABI H and VAN DEN BRINK PJ (2015) Monitoring and assessment of water health quality in the Tajan River, Iran, using physicochemical, fish and macroinvertebrates indices. J. Environ. Health Sci. Eng. 13 29. https://doi.org/10.1186/s40201-015-0186-y [ Links ]
ABOWEI JFN (2010) Salinity, dissolved oxygen, pH and surface water temperature conditions in Nkoro River, Niger Delta, Nigeria. Adv. J. Food Sci. Technol. 2 (1) 36-40. [ Links ]
ADU BW and OYENIYI EA (2019) Water quality parameters and aquatic insect diversity in Aahoo stream, southwestern Nigeria. J. Basic Appl. Zool. 80 15 https://doi.org/10.1186/s41936-019-0085-3 [ Links ]
AGBOOLA OA, DOWNS CT and O'BRIEN G (2020) A multivariate approach to the selection and validation of reference conditions in KwaZulu-Natal Rivers, South Africa. Front. Environ. Sci. 8 584923. https://doi.org/10.3389/fenvs.2020.584923 [ Links ]
ALHEJOJ I, ELIAS S and KLAUS B (2014) Mayflies (Order Ephemeroptera): an effective indicator of water bodies conditions in Jordan. Int. J. Sci. Res. Environ. Sci. 2 (10) 361-370. https://doi.org/10.12983/ijsres-2014-p0361-0370 [ Links ]
ALHEJOJ I, HIASAT TH, SALAMEH E, HAMAD AA and AL KUISI M (2023) Use of the aquatic mayfly (Insecta: Ephemer-optera) as environmental bio-indicator in Jordan. Int. J. Design Nat. Ecodyn. 8 (1) 133-139. https://doi.org/10.18280/ijdne.180115 [ Links ]
BAPTISTA DF, BUSS DF, DIAS LG, NESSIMIAN JL, DA SILVA ER, NETO AD, DE CARVALHO SN, DE OLIVEIRA MA, ANDRADE LR (2006) Functional feeding groups of Brazilian Ephemeroptera nymphs: Ultrastructure of mouthparts. Int. J. Limnol. 42 (2) 87-96. https://doi.org/10.1051/limn/2006013 [ Links ]
BAUERNFEIND E and MOOG O (2000) Mayflies (Insecta: Ephemer-optera) and the assessment of ecological integrity: a methodological approach, (pp. 71-83). In: Jungwirth M, Muhar S and Schmutz S (eds) Assessing the Ecological Integrity of Running Waters. Springer, The Netherlands. https://doi.org/10.1007/978-94-011-4164-2_6 [ Links ]
BEYENE A, ADDIS T, KIFLE D, LEGESSE W, KLOOS H and TRIEST L (2009) Comparative study of diatoms and macroinver-tebrates as indicators of severe water pollution: case study of the Kebena and Akaki rivers in Addis Ababa, Ethiopia. Ecological Indicators 9 381-392. https://doi.org/10.1016/j.ecolind.2008.05.001 [ Links ]
BOYERO L, PEARSON RG, DUDGEON D, GRAQA MA, GESSNER MO, ALBARINO RJ and CALLISTO M (2011) Global distribution of a key trophic guild contrasts with common latitudinal diversity patterns. Ecology 92 (9) 1839-1848. https://doi.org/10.1890/10-2244.1 [ Links ]
BULUTA S, FINAU I, BRODIE G and HODGE S (2010) A preliminary study into the potential of mayflies (Ephemeroptera: Baetidae and Caenidae) as bio-indicators of stream health in Fiji. S. Pacific J. Nat. Appl. Sci. 28 (1) 82-84. https://doi.org/10.1071/SP10011 [ Links ]
BUSS DF and SALLES FF (2007) Using Baetidae species as biological indicators of environmental degradation in a Brazilian river basin. Environ. Monit. Assess. 130 365-372. https://doi.org/10.1007/s10661-006-9403-6 [ Links ]
CHRISTIDIS F, CONNOLLY NM, NOLEN JA, PEARSON RG, CLAIR R, CAIRNS A and DAVIS L (2017) Stream macroinverte-brate assemblage uniformity and drivers in a tropical bioregion. Freshwater Biol. 62 (3) 544-558. https://doi.org/10.1111/fwb.12884 [ Links ]
CLARKE KR and GORLEY RN (2006) Primer v.6: User Manual/Tutorial. PRIMER-E. [ Links ]
COUCEIRO SR, HAMADA N, FORSBERG BR, PIMENTEL TP and LUZ SL (2012) A macroinvertebrate multimetric index to evaluate the biological condition of streams in the Central Amazon region of Brazil. Ecol. Indic. 18 118-125. https://doi.org/10.1016/j.ecolind.2011.11.001 [ Links ]
CROIJMANS L, DE JONG JF and PRINS HH (2020) Oxygen is a better predictor of macroinvertebrate richness than temperature -a systematic review. Environ. Res. Lett. 21;16(2):023002. https://doi.org/10.1088/1748-9326/ab9b42 [ Links ]
DE MOOR IJ, DAY JA and DE MOOR FC (2003) Guides to the freshwater invertebrates of Southern Africa. Vol. 7: Insecta I (Ephemeroptera, Odonata and Plecoptera). WRC Report No. TT 207/03. Water Research Commission, Pretoria. [ Links ]
DUDGEON D, ARTHINGTON AH, GESSNER MO, KAWABATA ZI, KNOWLER DJ, LÉVÊQUE C, NAIMAN RJ, PRIEUR-RICHARD AH, SOTO D, STIASSNY ML, SULLIVAN CA (2006) Freshwater biodiversity: importance, threats, status and conservation challenges. Biol. Rev. 81 163-182. https://doi.org/10.1017/S1464793105006950 [ Links ]
DWAF (Department of Water Affairs and Forestry, South Africa) (1996) South African Water Quality Guidelines. Volume 7: Aquatic Ecosystems. Department of Water Affairs and Forestry, Pretoria. [ Links ]
EDOKPAYI JN, ODIYO JO, POPOOLA OE and MSAGATI TAM (2016) Assessment of trace metals contamination of surface water and sediment: a case study of Mvudi River, South Africa. Sustainability 8 135. https://doi.org/10.3390/su8020135 [ Links ]
GATTOLLIAT JL and NIETO C (2009) The family Baetidae (Insecta: Ephemeroptera): synthesis and future challenges. Aquat. Insects 31 (sup1) 41-62. https://doi.org/10.1080/01650420902812214 [ Links ]
GUSTAFSON MP (2008) Effects of thermal regime on mayfly assemblages in mountain streams. Hydrobiologia 605 (1) 235-246. https://doi.org/10.1007/s10750-008-9357-5 [ Links ]
HAIDEKKER A and HERING D (2008) Relationship between benthic insects (Ephemeroptera, Plecoptera, Coleoptera, Trichoptera) and temperature in small and medium-sized streams in Germany: A multivariate study. Aquat. Ecol. 42 (3) 463-481. https://doi.org/10.1007/s10452-007-9097-z [ Links ]
HAMID A, BHAT SU and JEHANGIR A (2020) Local determinants influencing stream water quality. Appl. Water Sci. 10 (1) 1-16. https://doi.org/10.1007/s13201-019-1043-4 [ Links ]
HEATH RGM and CLASSEN M (1990) An overview of the pesticide and metal levels present in populations of the larger indigenous fish species of selected South African rivers. WRC Report No. 428/1/99. Water Research Commission, Pretoria. [ Links ]
JACOBUS LM, MACADAM CR and SARTORI M (2019) Mayflies (Ephemeroptera) and their contributions to ecosystem services. Insects 10 170. https://doi.org/10.3390/insects10060170 [ Links ]
JAMIL T and TER BRAAK CJF (2013) Generalized linear mixed models can detect unimodal species-environment relationships. PeerJ 1 e95. https://doi.org/10.7717/peerj.95 [ Links ]
KABORÉ I, MOOG O, OUÉDA A, SENDZIMIR J, OUÉDRAOGO R, GUENDA W and MELCHER AH (2018) Developing reference criteria for the ecological status of West African rivers. Environ. Monit. Assess. 190 2. https://doi.org/10.1007/s10661-017-6360-1 [ Links ]
KARR JR (1999) Defining and measuring river health. Freshwater Biol. 41 221-234. https://doi.org/10.1046/j.1365-2427.1999.00427.x [ Links ]
KLEYNHANS CJ (1996) A qualitative procedure for the assessment of the habitat integrity status of the Luvuvhu River (Limpopo system, South Africa). J. Aquat. Ecosyst. Health 5 41-54. https://doi.org/10.1007/BF00691728 [ Links ]
KLONOWSKA-OLEJNIK M and SKALSKI T (2014) The effect of environmental factors on the mayfly communities of headwater streams in the Pieniny mountains (west carpathians). Biologia 69 (4) 498-507. https://doi.org/10.2478/s11756-014-0334-3 [ Links ]
KUBENDRAN T, SELVAKUMAR C, SIDHU AK, NAIR A and KRISHNAN SM (2017) Baetidae (Ephemeroptera : Insecta) as biological indicators of environmental degradation in Tamiraparani and Vaigai river basins of Southern Western Ghats, India. Int. J. Curr. Microbiol. Appl. Sci. 6 (6) 558-572. https://doi.org/10.20546/ijcmas.2017.606.066 [ Links ]
KUSANGAYA S, WARBURTON ML, ARCHER VAN GARDEREN E and JEWITT GPW (2013) Impacts of climate change on water resources in southern Africa: A review. Phys. Chem. Earth 67 47-54. https://doi.org/10.1016/j.pce.2013.09.014 [ Links ]
LEWIN I, CZERNIAWSKA-KUSZA I, SZOSZKIEWICZ K, EWA-LAWNICZAK A and JUSIK S (2013) Biological indices applied to benthic macroinvertebrates at reference conditions of mountain streams in two ecoregions (Poland, the Slovak Republic). Hydrobiologia 709 183-200. https://doi.org/10.1007/s10750-013-1448-2 [ Links ]
LI L, ZHENG B and LIU L (2010) Biomonitoring and bioindicators used for river ecosystems: definitions, approaches and trends. Proced. Environ. Sci. 2 1510-1524. https://doi.org/10.1016/j.proenv.2010.10.164 [ Links ]
LI S, YANG W, WANG L, CHEN K, XU S and WANG B (2018) Influences of environmental factors on macroinvertebrate assemblages: differences between mountain and lowland ecoregions, Wei River, China. Environ. Monit. Assess. 190 152. https://doi.org/10.1007/s10661-018-6516-7 [ Links ]
LÓPEZ-LÓPEZ E and SEDENO-DÍAZ JE (2015) Biological indicators of water quality: the role of fish and macroinvertebrates as indicators of water quality. In: Armon R and Hänninen O (eds) Environmental Indicators. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-9499-2_37 [ Links ]
MANGADZE T, DALU T and FRONEMAN PW (2019) Biological monitoring in southern Africa: a review of the current status, challenges and future prospects. Sci. Total Environ. 648 1492-1499. https://doi.org/10.1016/j.scitotenv.2018.08.252 [ Links ]
MCDOWELL RW, SNELDER TH, COX N, BOOKER DJ and WILCOCK RJ (2013) Establishment of reference or baseline conditions of chemical indicators in New Zealand streams and rivers relative to present conditions. Mar. Freshwater Res. 64 387-400. https://doi.org/10.1071/MF12153 [ Links ]
MONYAI M, MAKHADO RA and NOVHE NO (2016) Water quality of the Luvuvhu River and its tributaries within the Thulamela Local Municipality, Limpopo Province, South Africa. Afr. J. Sci. Technol. Innovat. Dev. 8 (5-6) 439-445. https://doi.org/10.1080/20421338.2016.1219504 [ Links ]
MOULTON TP, SOUZA ML, SILVEIRA RML and KRSULOVIC FAM (2004) Effects of ephemeropterans and shrimps on periphyton and sediments in a coastal stream (Atlantic forest, Rio de Janeiro, Brazil). J. N. Am. Benthol. Soc. 23 868-881. https://doi.org/10.1899/0887-3593(2004)023<0868:EOEASO>2.0.CO;2 [ Links ]
NAKAGAWA S and SCHIELZETH H (2013) A general and simple method for obtaining R2 from generalized linear mixed effects models. Meth. Ecol. Evol. 42 133-142. https://doi.org/10.1111/j.2041-210x.2012.00261.x [ Links ]
OKONKWO JO and MOTHIBA M (2005) Physico-chemical characteristics and pollution levels of heavy metals in the rivers in Thohoyandou, South Africa. J. Hydrol. 308 122-127. https://doi.org/10.1016/j.jhydrol.2004.10.025 [ Links ]
PITMAN WV (2011) Overview of water resource assessment in South Africa: current state and future challenges. Water SA 37 (5) 659-664. https://doi.org/10.4314/wsa.v37i5.3 [ Links ]
RAMULIFHO PA (2020) Modelling flow and water temperature in the Luvuvhu catchment and their impact on macroinvertebrate assemblages. PhD thesis, University of Venda. [ Links ]
RAMULIFHO PA, FOORD SH and RIVERS-MOORE NA (2020) The role of hydro-environmental factors in Mayfly (Ephem-peroptera, Insecta) community structure: identifying threshold responses. Ecol. Evolut. 10 (14) 6919-6928. https://doi.org/10.1002/ece3.6333 [ Links ]
RASHID I and ROMSHOO SA (2013) Impact of anthropogenic activities on water quality of Lidder River in Kashmir Himalayas. Environ. Monit. Assess. 185 4705-4719. https://doi.org/10.1007/s10661-012-2898-0 [ Links ]
REECE PF, REYNOLDSON TB, RICHARDSON JS and ROSENBERG DM (2001) Implications of seasonal variation for biomonitoring with predictive models in the Fraser River catchment, British Columbia. Can. J. Fish. Aquat. Sci. 58 1411-1418. https://doi.org/10.1139/f01-076 [ Links ]
ROWNTREE KM and WADESON RA (1999) A hierarchical geomorphological model for the classification of selected South African rivers. WRC Report No. 497/1/99. Water Research Commission, Pretoria. [ Links ]
SARTORI M and BRITTAIN JE (2015) Order Ephemeroptera. In: Thorp J and Rodgers DC (eds) Thorp and Covich's Freshwater Invertebrates: Ecology and General Biology (4th edn.). Academic Press, New York. 873-891. https://doi.org/10.1016/B978-0-12-385026-3.00034-6 [ Links ]
SHARMA RC and RAWAT JS (2009) Monitoring of aquatic macroinvertebrates as bioindicators for assessing the health of wetlands: a case study in the Central Himalayas, India. Ecol. Indic. 9 118-128. https://doi.org/10.1016/j.ecolind.2008.02.004 [ Links ]
SIEGLOCH AE, SURIANO M, SPIES M. and FONSECA-GESSNER A (2014) Effect of land use on mayfly assemblages' structure in Neotropical headwater streams. An. Acad. Bras. Ciê. 86 (4) 1735-1747. https://doi.org/10.1590/0001-3765201420130516 [ Links ]
SINGO LR, KUNDU PM, ODIYO JO, MATHIVHA FI and NKUNA TR (2012) Flood frequency analysis of annual maximum stream flows for Luvuvhu River Catchment, Limpopo Province, South Africa. In: 16th SANCIAHS Hydrology Symposium, 1-3 October 2012, University of Pretoria. [ Links ]
SKOWNO AL, POOLE CJ, RAIMONDO DC, SINK KJ, VAN DEVENTER H, VAN NIEKERK L, HARRIS LR, SMITH-ADAO LB, TOLLEY KA, ZENGEYA TA, FODEN WB, MIDGLEY GF and DRIVER A (2019) National Biodiversity Assessment 2018: The status of South Africa's ecosystems and biodiversity. Synthesis Report. South African National Biodiversity Institute, an entity of the Department of Environment, Forestry and Fisheries, Pretoria. 214 pp. [ Links ]
SVITOK M (2006) Structure and spatial variability of mayfly (Ephemeroptera) communities in the upper Hron river basin. Biologia 61 (5) 547-554. https://doi.org/10.2478/s11756-006-0089-6 [ Links ]
TAMPO L, LAZAR IM, KABORÉ I, OUEDA A, AKPATAKU KV, DJANEYE-BOUNDJOU G, BAWA LM, LAZAR G and GUENDA WA (2020) A multimetric index for assessment of aquatic ecosystem health based on macroinvertebrates for the Zio river basin in Togo. Limnologica 83 125783. https://doi.org/10.1016/j.limno.2020.125783 [ Links ]
TER BRAAK CJF and VERDONSCHOT PFM (1995) Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 37 130-137. [ Links ]
THIRUPATHAIAH M, SAMATHA CH and SAMMAIAH C (2012) Analysis of water quality using physico-chemical parameters in lower manair reservoir of Karimnagar district, Andhra Pradesh Environ. Sci. 3 (1) 172-180. https://doi.org/10.6088/ijes.2012030131017 [ Links ]
VARNOSFADERANY NM, EBRAHIMI E, MIRGHAFFARY N and SAFYANIAN A (2010) Biological assessment of the Zayandeh Rud River, Iran, using benthic macroinvertebrates. Limnologica 40 226-232. https://doi.org/10.1016/j.limno.2009.10.002 [ Links ]
VENABLES WN, and RIPLEY BD (2002) Random and mixed effects. In: Modern Applied Statistics with S. Statistics and Computing. Springer, New York, NY. https://doi.org/10.1007/978-0-387-21706-2_10 [ Links ]
VENKATESHARAJU K, RAVIKUMAR P, SOMASHEKAR RK and PRAKASH KL (2010) Physico-chemical and bacteriological investigation on the river Cauvery of Kollegal Stretch in Karnataka. J. Sci. Eng. Technol. 6 (1) 50-59. https://doi.org/10.3126/kuset.v6i1.3310 [ Links ]
WALLACE JB and WEBSTER JR (1996) The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 41 115-139. https://doi.org/10.1146/annurev.en.41.010196.000555 [ Links ]
WELDEMARIAM MM (2013) Physico-chemical analysis of GudBahri river water of Wukro, Eastern Tigrai, Ethiopia. Int. J. Sci. Res. Publ. 3 (11) 1-4. [ Links ]
WHO (World Health Organisation) (2011) Guidelines for Drinking-Water Quality (4th edn). World Health Organisation Press, Geneva. [ Links ]
 Correspondence:
Correspondence:
Pfananani Ramulifho
Email:pfananani.ramulifho@gmail.com
Received: 18 September 2022
Accepted: 19 June 2023


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}