










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.48 n.3 Pretoria Jul. 2022
http://dx.doi.org/10.17159/wsa/2022.v48.i3.3939
RESEARCH PAPER
Yeast supplementation alleviates the negative effects of greywater irrigation on lettuce and maize
LP TshapaI; G NaidooI; SershenII, III; KK NaidooIV
ISchool of Life Sciences, University of KwaZulu-Natal, Private Bag X54001, Westville, 4000, South Africa
IIDepartment for Biodiversity & Conservation Biology, University of the Western Cape, Private Bag X17, Bellville, 7535, South Africa
IIIInstitute of Natural Resources, PO Box 100396, Scottsville, 3209, South Africa
IVDepartment of Nature Conservation, Mangosuthu University of Technology, PO Box 12363, Jacobs, 4026, South Africa
ABSTRACT
Water scarcity has led to increased use of wastewater, particularly greywater, for crop irrigation. This study investigated whether the addition of yeast can alleviate the potential negative effects of greywater use on lettuce (Lactuca sativa L.) and maize (Zea mays L.). Seeds and seedlings were treated with 4 concentrations (0.005; 0.01; 0.015 and 0.020 g·mL−1) of yeast-treated tapwater (YTW) and greywater (YGW). Tapwater (TW) and greywater (GW) without yeast served as controls. In general, an increase in yeast concentration compromised seed germination in Petri dishes, but improved germination in soil. Tapwater was more effective than GW in promoting germination and growth in both species. Lower concentrations of yeast generally increased germination capacity in both species compared to the controls. Total biomass, number of leaves, chlorophyll content, leaf area, photosynthetic rate and maximum quantum yield of photosystem II (Fv/Fm) were significantly higher in yeast treatments in both species, compared with the controls. Biomass accumulation, total leaf area, chlorophyll content and photosynthesis were higher in YGW than controls and YTW. Differences in biomass allocation between treatments may be due to changes in soil moisture, pH and electrical conductivity of the soil caused by yeast supplementation. This study showed that plants treated with YGW performed better than those treated with YTW and without yeast. Yeast supplementation of greywater could increase water recycling and provide a cheap bio-fertilizer to home growers, whilst significantly improving yield in both species. This innovative approach may enhance water and food security of subsistence farmers in rural areas.
Keywords: bio-fertilizer, growth, germination, subsistence agriculture
INTRODUCTION
Rapid population growth has increased water requirements for domestic, industrial and agricultural uses. The demand for water has escalated to the point that countries need to invest heavily in alternative sources such as recycled water (Pinto and Maheshwari, 2015). Widespread water shortage is a threat to conservation and socio-economic development. It is predicted that by 2025, 3 billion people will be living in either water-stressed or water-limited regions (WHO, 2019). Therefore, to meet this demand, innovative water conservation techniques are required. In the face of climate change and rainfall variability, greywater reuse is important in minimizing water shortages and hunger and in poverty alleviation (Pinto and Maheshwari, 2015).
Greywater refers to untreated wastewater from the bath, shower, kitchen, washbasin, and laundry of households (Rodda et al., 2011). Greywater reuse is an alternative and effective source to reduce pressure on freshwater for food production and poverty alleviation in third-world countries (Radingoana et al., 2020). However, this resource is regarded as unclean and unfit for human use and crop irrigation. Health risk is the most important factor in household-level decisions about reuse. Poorly-treated wastewater results in the deposition of heavy metals, microbial pathogens and organic chemicals in soils and plants (Misra et al., 2010; Singh, 2021). Use of greywater is limited because of the lack of awareness of its potential in supplementing freshwater, especially in developing countries. Studies have shown that the views and perspectives of potential users of recycled wastewater generated in the household influence the adoption of technologies at the household level and beyond (Portman et al., 2022).
Adoption of water recycling measures is important, especially when considering urban development needs in the face of warming climates and declining water resources and when aiming to increase sustainability in urban planning and development (Portman et al., 2022). The World Health Organization highlights how countries such as the USA, Australia, China, and Japan have widely accepted greywater reuse. This practice, however, is limited in some African and Middle Eastern countries (WHO, 2019). Finding a safe and affordable treatment process is the key to sustainable reuse of greywater for domestic, agricultural, or industrial purposes. By having a proper treatment system in place, health concerns resulting from the presence of heavy metals and pathogens in greywater can be avoided (Pinto and Maheshwari, 2015). The use of greywater for irrigation may be an approach to conserving water, especially in drought-stricken areas (Nel and Jacobs, 2019). Greywater may also contain bacteria, fibre particles, and dead skin cells, which may contribute to soil fertility (Glick, 2012). Greywater decreased germination of Amaranthus dubius and reduced the rate of shoot emergence of Solanum nigrum relative to tapwater irrigation (Lubbe et al., 2016). Also, elevated salinity, pH, and the boron content of greywater adversely impact soil and stunt growth in carrot, lettuce, and red pepper (Finley et al., 2009).
The excessive use of inorganic fertilizers threatens the integrity of ecosystems and global biogeochemical cycles by causing pollution and loss of biota (Paszko et al., 2016). The application of bio-fertilizers could improve soil aggregation, structure, aeration, and infiltration (Dhanasekar and Dhandapani, 2012). Microbes within bio-fertilizers can provide mutually beneficial nutrients to plants (Bargaz et al., 2018). In addition, microbe-based bio-stimulants may enhance rhizophagy without having any negative effects on soil health and the environment (Anandaraj and Delapierre, 2010). Bio-fertilizers and bio-stimulants are also cheaper, more effective and a renewable source of nutrients for plants compared to inorganic fertilizers (Boraste et al., 2009; Al-Mefleh et al., 2021). A commonly used bio-stimulant is bread yeast (Saccharomyces cerevisiae) which improves growth and yield in many crops (Fahad et al., 2016).
Several new technologies, with yeast as the core, have been widely applied in water treatment and shown great potential and broad prospects in recent years (Wang et al., 2018). At present, yeasts have been applied in many kinds of industrial wastewater treatment, domestic sewage purification and other fields (Yang and Zheng, 2014). The efficiency of yeast in the treatment of wastewater has attracted wide attention. Treatment of wastewater with yeast produces lipids, glycolipids and enzymes (Yang et al., 2013). Therefore, it is widely used in the treatment of wastewater containing high concentrations of organics, heavy metals and domestic sewage. Yeast has been shown to convert most of the organic matter into non-toxic and nutritious single-cell protein (Yang and Zheng, 2014). Application of yeast as a biostimulant increased the growth of hyacinth beans compared to those without bio-stimulants (Velhal et al., 2014). Yeast also enhanced nitrogen and phosphorus uptake in tomato and sugarcane (Lonhienne, 2014). Yeast promotes cell division and extension, seed and tuber germination and improves root development. Yeast also increases ion uptake, vegetative growth, and improves pigment formation and photosynthesis (Hashem et al., 2008). The stimulatory effects of yeast are due to enhanced levels of cytokinins which increase cell division and cell enlargement (Jensen et al., 2004; Tiwari et al., 2020).
Yeast has been shown to be effective in the treatment of various wastewater types (Uysal et al., 2014) and enhances the germination and growth of many species (Abu-Dieyeh et al., 2017). There is little or no information on the use of yeast in alleviating the negative effects of polluted water on crop growth. In this study, we pretreated greywater with yeast and determined its effects on germination, growth and gas exchange in lettuce (Lactuca sativa L.) and maize (Zea mays L.). We tested the hypothesis that the addition of yeast to greywater would enhance growth of lettuce and maize. This study will contribute toward the wider acceptance of domestic greywater reuse for subsistence and agricultural irrigation. Furthermore, in water-stressed countries like South Africa, improved subsistence and commercial crop productivity are based on innovative approaches to increasing water availability for irrigation and this may be largely dependent on the reuse of wastewater for irrigation.
MATERIALS AND METHODS
Plant material
Instant baker's yeast, (Anchor Yeast Pty Ltd., South Africa) containing Saccharomyces cerevisiae was used in the study. Lettuce and maize seeds were purchased from Grovida Pty Ltd. (Durban, South Africa). Yeast was activated in warm water (27°C; 0.10 g.L−1), stirred, and then placed in an incubator at 30°C for 30 min.
Greywater synthesis
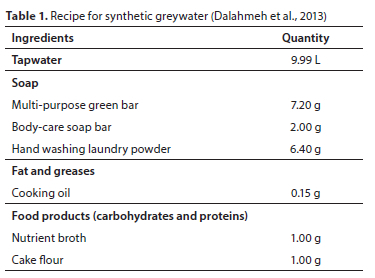
Synthetic greywater was prepared with ingredients representing the various components found in typical household greywater, such as soap, oil, grease, carbohydrates, and proteins (Table 1) (Lubbe et al., 2016). The greywater used conformed to chemical measures (chemical oxygen demand, biological demand, total nitrogen and total phosphorus) associated with mixed domestic greywater (Dalahmeh et al., 2013). Tapwater used for the preparation of all irrigation solutions was de-chlorinated by bubbling air with an aquarium jet filter pump (MAGI-700, RESUN, Shenzhen, China) and left to stand for 24 h before use.
Seed germination
Five millilitre (5 mL) solutions of each control (TW = tapwater and GW = greywater) and 4 concentrations of yeast-treated tapwater (YTW) and greywater (YGW) (0.005; 0.01; 0.015 and 0.02 g.mL−1 were applied to 3 replicates of 25 seeds of each species. The seeds were placed between Whatman No. 4 filter paper (90 mm diameter) within Petri dishes and incubated in a growth room for 14 days (24°C, 16 h day, 8 h night). Each Petri dish received a total of 5 mL of irrigation solution every 2nd day for 2 weeks for maize and 7 days for lettuce. Germination was monitored daily for radical emergence greater than 1 mm in lettuce and 2 mm in maize. The time (in days) taken to achieve 50% of final germination capacity was used to compare germination rates across treatments (Perumal et al., 2014). At the end of the 14 days, shoot and root lengths were measured and total germination capacity was calculated.
Seedling growth
Seeds were sown in pots (1 cm depth) and maintained in a greenhouse for 62 days (n = 20). Climatic conditions in the glasshouse were 25°C (day), 18°C (night), average maximal photosynthetic photon flux density (PPFD 800 µmol.m−2.s−1) and relative humidity 75%.
Each plastic pot (2 L for lettuce and 4 L for maize) was filled with the same volume (3 kg for lettuce and 5 kg for maize) of topsoil (Ukulinga Research Farm, University of KwaZulu-Natal) and watered to field capacity with irrigation solution (80 mL for lettuce and 100 mL for maize). Shoot emergence was recorded weekly.
Morphometric measurements
After 62 days, plants were harvested and roots were carefully washed with water to remove soil. The roots, shoots, and leaves of each plant were separated and total leaf area measured using a leaf area meter (Model CI-202, CID Inc., Germany). Shoot and root length of individual seedlings were measured. Plants material was then dried in an oven at 55°C for 7 days and dry weight determined. Specific leaf area (SLA), leaf area ratio (LAR), root:shoot ratio, and biomass allocation percentages were calculated from the biometric measurements.
Physiological measurements
A hand-held chlorophyll absorbance meter (Minolta SPAD-502, Minolta Camera Co. Ltd.) was used to measure chlorophyll content on the 3rd leaf from the apex of each plant. Measurements were taken 7 days before harvest. The maximum quantum efficiency of photosystem II (PSII) (Fv/Fm) was measured using an integrated gas exchange and chlorophyll fluorescence system (Li-6400XT, LI-COR, Lincoln, USA) (n = 20). Plants were dark-adapted for 40 min to permit electrons to drain from the photosystems (Miradi and Ismail, 2007) for Fv/Fm measurements. One measurement per plant was taken on the lamina, midway between the base and the tip of the 3rd leaf from the apex (Naidoo, 2009). Gas exchange, stomatal conductance and transpiration were measured (n = 20) between 11:00 and 14:00. Plants were randomly selected and measurements were taken on the 3rd leaf from the apex under a light intensity of 1 000 µmol.m−2.s−1 and a CO2 concentration of 400 ppm.
Soil characteristics
Soil moisture content was measured weekly using a soil moisture meter (HH2 moisture meter; Delta T Devices, London, United Kingdom). At the final harvest, 3 pots were assessed for soil pH [Checker HI 98103; South Africa]) and electrical conductivity (CM 100-2; Reid & Associates, South Africa).
Data analyses
All data were subjected to analysis of variance (ANOVA SPSS, Version 22). Means were separated using Tukey's test. Pearson's correlation coefficient was used to test for relationships between selected parameters. Percentage data were arcsine or log-transformed before parametric analyses. Non-parametric data were determined by the Kolmogorov-Smirnov test. Where data remained non-parametric, inter-treatment differences were tested by the Kruskal-Wallis test. Differences were considered significant at P < 0.05.
RESULTS
Seed germination
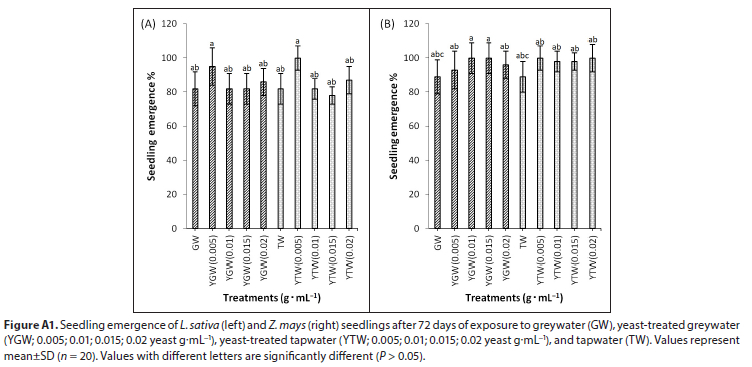
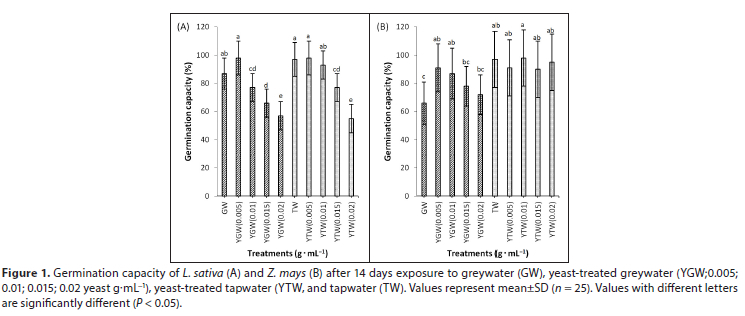
Seed germination in L. sativa was lower at yeast concentrations >0.015 g.mL−1 compared to the control (Fig. 1A). Generally, an increase in yeast concentration decreased germination in L. sativa (R2 = 0.412) but had no effect on seedling emergence in soil (Fig. A1, Appendix). In Z. mays, GW reduced germination capacity significantly (P < 0.05) relative to TW. The addition of yeast had no effect on germination in GW or TW (Fig. 1B).
Seedling growth
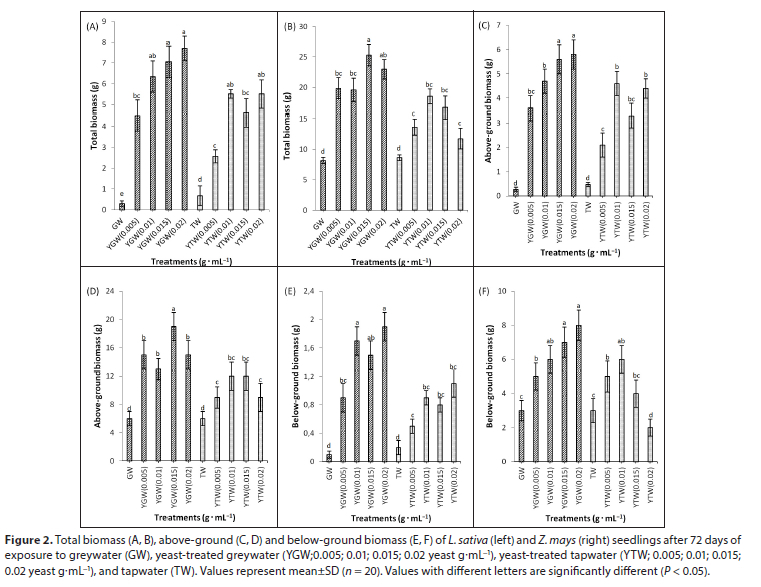
Total biomass in the yeast treatments was significantly higher than that of the controls in both species (Figs 2A, 2B). In L. sativa, the total biomass of the GW control was lower than that in TW, while in Z. mays there were no differences between treatments. Generally, an increase in yeast concentration increased biomass in both species (Table 2). Yeast-treated greywater was positively correlated with total biomass in both species (R2 = 0.84 for L. sativa and R2 = 0.71 for Z. mays); thus, an increase in yeast concentration caused an increase in total biomass (P < 0.005). Above-ground biomass was significantly greater (P < 0.05) in yeast treatments compared to controls in both species (Figs 2C, 2D). There was a positive correlation between above-ground biomass and yeast greywater in L. sativa; hence, an increase in yeast concentration caused an increase in above-ground biomass (R2 = YGW: 0.869 and YTW:0.787). Differences between YGW and YTW were not significant in both species (Figs 2C, 2D). Below and above-ground biomass did not differ between GW and TW in both species (Fig. 2).
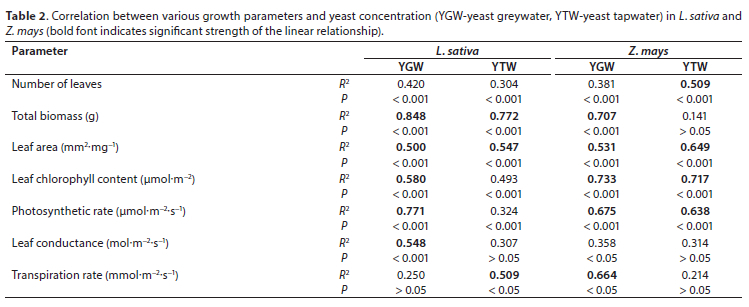
Yeast treatments generally increased below-ground biomass of both species relative to the controls (Figs 2E, 2F). Below-ground biomass was positively correlated to yeast-treated greywater in Z. mays (R2 = 702). In both species, YGW significantly increased (P < 0.05) below-ground biomass relative to the control (Figs 2E, 2F). In general, yeast-treated greywater increased below-ground biomass compared to yeast-treated tapwater in both species. Yeast increased plant height significantly (P < 0.05) in both species relative to the control (Figs 3A, 3B). Plant height of L. sativa in the yeast treatments was higher than those in the control and tapwater treatments (Figs 3A, 3B). In general, plant height of L. sativa increased with added yeast for YTW and YGW (Fig. 3A). Generally, there were no differences in the total number of leaves and plant height between GW and TW in both species (Figs 3 and 4).
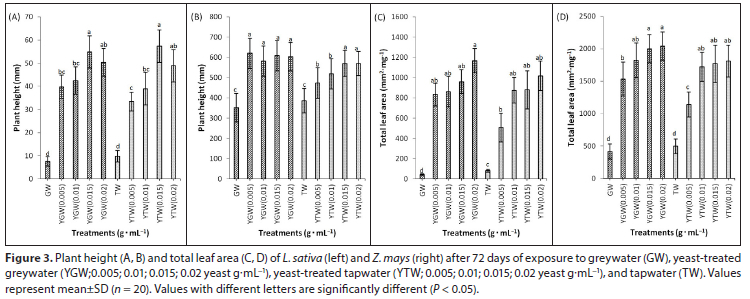
Total leaf area increased significantly (P < 0.05) with yeast additions compared to the control in both species (Figs 3C, 3D). Leaf area was positively correlated with yeast concentration in both species (Table 2). Yeast-treated GW and TW (0.02 g.mL−1) increased total leaf area in both species (Figs 3C, 3D). Yeast supplementation increased the total number of leaves in both species compared to the control (Figs 4A, 4B). The total number of leaves was positively correlated with yeast concentration in Z. mays (Table 2). Yeast-treated greywater did not affect root length in both species in the greywater treatments (Figs. 4C, 4D). Tapwater significantly increased root length of Z. mays compared to the yeast treatments (P < 0.05). In Z. mays, high yeast concentrations reduced root elongation in the tapwater treatments (Fig. 4D). Root length in tapwater treatments was significantly higher (P < 0.05) than those in greywater treatments (Figs 4C, 4D). The root length of both species was significantly greater (P < 0.05) in tapwater compared to greywater (Figs 4C, 4D). In L. sativa, tapwater significantly increased root elongation (P < 0.05) compared to the treatments with added yeast (Fig. 4C).
Physiology
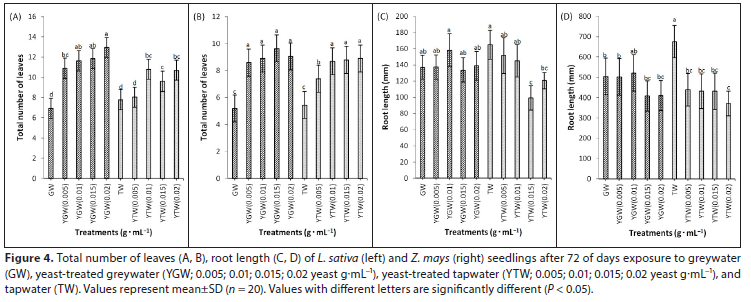
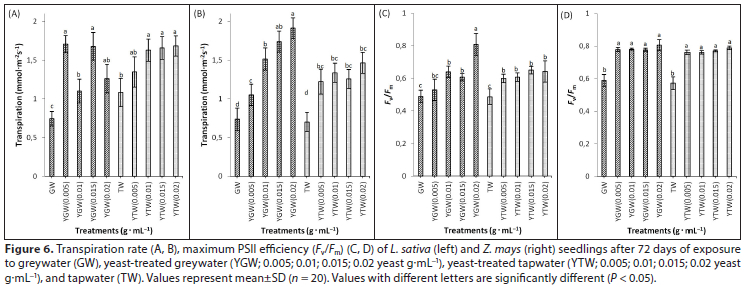
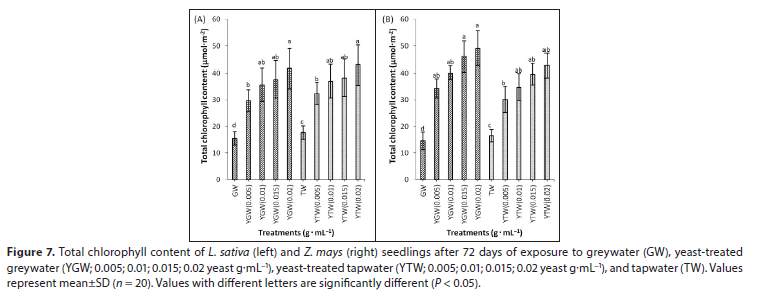
Yeast supplementation increased photosynthesis, stomatal conductance, transpiration, chlorophyll fluorescence and total chlorophyll content in both species, compared to the controls (Figs 5, 6 and 7). Photosynthetic rate and chlorophyll content were positively correlated with yeast concentration (Table 2). The highest concentration of yeast (0.02 g.mL−1) resulted in significantly higher (P < 0.05) photosynthesis and total chlorophyll content than other treatments in both species (Figs 5 and 7). Greywater significantly reduced (P < 0.05) total chlorophyll content compared to tapwater in both species (Fig. 7). Yeast supplementation increased Fv/Fm significantly (P < 0.05) compared to the respective controls in both species (Figs 6C and 6D). The highest concentration of yeast greywater (0.02 g.mL−1) increased Fv/Fm significantly compared to other treatments in L. sativa (Fig. 6C).
DISCUSSION
Germination
As yeast concentration increased, germination and root elongation of L. sativa decreased in Petri dishes. When grown in soil, however, germination and root elongation of L. sativa did not decline with an increase in yeast concentration (Fig. A1, Appendix). Inhibition of germination may be due to phytotoxicity caused by low pH due to yeast fermentation in Petri dishes (Lin and Xin, 2007). High yeast concentrations (> 0.01 g.mL−1) may create an anaerobic environment when grown in Petri dishes. Greywater reduced germination capacity significantly compared to tapwater, probably due to the presence of excessive salts, total suspended solids, and nutrients such as nitrogen, ammonia, and phosphate (Corwin et al., 2005). Low yeast concentrations (0.0015 g.mL−1) produced maximum germination compared to the controls, probably due to the presence of hydrolase enzymes and growth-promoting rhizobacteria, compared to the controls (Delshadi et al., 2017). Low concentrations of yeast (0.5 %) also improved seed germination, yield and fruit quality parameters of olive trees (Ahmed and Ragab, 2002). In contrast, concentrations of yeast above 0.01 g.mL−1 in greywater reduced growth and caused abnormal seedlings in L. sativa.
High concentrations of yeast (0.02 g.mL−1) reduced root and shoot length of L. sativa in Petri dishes probably due to the acidic pH caused by yeast fermentation (Ge et al., 2016). Lettuce seeds were more sensitive to irrigation using greywater than maize. This could be attributed to genotypic variation between these two species (Pinto et al., 2010).
In both species, tapwater improved germination, root length, vigour, and biomass than greywater. Greywater with high yeast (> 0.01 g.mL−1) reduced root and shoot elongation, vigour, and biomass of L. sativa (data not shown), probably due to the greater electrical conductivity of the soil (Hasanuzzaman et al., 2013). In soil, yeast concentration of 0.02 g.mL−1 in both greywater and tapwater did not have any detrimental effect in both species (Fig. A1, Appendix).
Seedling growth
The highest concentration of yeast-treated greywater (0.02 g.mL−1) improved the growth and biomass of both species, probably due to the buffering capacity of the soil. Shoot emergence was > 75% in L. sativa and > 85% in Z. mays (data not shown), which indicated soil buffering of greywater and yeast. The lowest concentration of yeast-treated tapwater (0.0015 g.mL−1) stimulated seedling growth in L. sativa. A low concentration of active bread yeast (0.5%) improved seed germination, yield, and fruit quality of olive trees (Ahmed and Ragab, 2002). Bread yeast nourishes plants when applied to soil because of the production of growth regulators such as gibberellins and auxins (Sarhan et al., 2011) and useful enzymes (Dinkha and Khazragji, 1990). In the absence of yeast, there was a low percentage of shoot emergence in Z. mays due to the absence of bio-fertilizer. Others showed that yeast can synthesize various phytohormones, enzymes, and solubilise minerals (Panhwar et al., 2012) while indirectly inhibiting phytopathogens (Hao et al., 2011).
In general, total number of leaves and total leaf area were higher in yeast treatments compared to the controls in both species. Yeast treatment of greywater and tapwater improved growth and yield in both species. Enhanced growth was probably due to the presence of growth-promoting rhizobacteria (PGPR) that solubilize nutrients for growth (Lonhienne et al., 2014). Bread yeast was shown to increase leaf number, leaf area, leaf mass, and photosynthesis in cucumber (Shahata et al., 2012). Yeast treatment of grey- and tapwater improved root growth and biomass by solubilizing nutrients (Lonhienne et al., 2014), including nitrogen and phosphorus (Costa et al., 2014). The increase in crop yield, number of leaves, vigour, shoot height, and chlorophyll content with yeast treatments was due to stimulation of growth hormones like auxins, gibberellins, and cytokinin (Jensen, 2004). Greywater treated with yeast improved shoot elongation, number of leaves, vigour, and biomass of both species relative to yeast-treated tapwater. This was probably due to higher nitrates and phosphorus in the soil (Lonhienne et al., 2014). The increase in total leaf area in both species was probably due to stimulation of growth by auxins produced by yeast (Gollan and Wright, 2006). Yeast promotes biologically fixed nitrogen, phytohormones, volatiles, defense compounds, and enzymes (Kuklinsky et al., 2004). Greywater reduced leaf area, vigour, and total biomass significantly in both species, probably due to its effect on soil pH, salinity, and the presence of toxic elements (Lubbe et al., 2016). Many researchers reported that bread yeast enhances yield in different crop plants (Costa et al., 2014, Lonhienne et al., 2014).
Yeast treatments increased total chlorophyll content compared to controls significantly in both species. Leaf chlorophyll content usually increases with increasing leaf nitrogen (Marenco et al., 2009). The combination of yeast and greywater possibly mobilized nitrogen-fixing bacteria, enhancing nutrient availability (Gollan and Wright, 2006; Basak and Biswas, 2010; Taha et al., 2011). Yeast treatments increased CO2 uptake and Fv/Fm in both species compared to controls (Hashem et al., 2008). Similar results for Fv/Fm were reported in amaranth and lettuce with greywater irrigation (Qin et al., 2013). The maximum quantum yield is the maximum efficiency at which light is absorbed by light-harvesting antennae chlorophyll of PSII and converted to chemical energy (Baker and Rosenquist, 2004). Growth and physiology outcomes suggest that use of yeast-treated greywater for crop irrigation is effective, eco-friendly and economically viable (Table A1, Appendix).
CONCLUSION
Greywater alone impaired seed germination and seedling growth in both species compared to yeast supplementation. However, high concentrations of yeast impaired germination of both species, with effects being greater in Petri dishes than in soil. Yeast supplementation alleviated phytotoxicity effects of greywater in both species. Yeast treatments stimulated shoot length, number of leaves, leaf area, biomass, total chlorophyll content, and photosynthesis in both species relative to the controls. The use of yeast-treated greywater is an innovative and economically viable approach to the resuse of greywater water for irrigation and improving food security in resource-limited settings. Impoverished communities could use greywater supplemented with yeast as a cheap bio-fertilizer to increase yields. However, water quality requirements for greywater reuse vary considerably globally and the yeast concentrations applied here may need to be adjusted as needed. Further studies should focus on quality treatment processes for sustainable reuse of greywater for domestic, agricultural, or industrial purposes. Based on the scale and type of use, it may be unnecessary to treat greywater to the highest quality before use in subsistence farming. However, the composition of greywater varies with different sources and additional studies are needed to assess the benefits of yeast treatment of greywater for different crops.
ACKNOWLEDGEMENTS
The financial support of Mangosuthu University of Technology (grant number NSCI: 01/2003 to KK Naidoo) and the National Research Foundation (Grant No. 93560 to G Naidoo) are gratefully acknowledged. The University of KwaZulu-Natal provided technical support.
REFERENCES
ABU-DIEYEH MH, DIAB M and AL-GHOUTI MA (2017) Ecological and agriculture impacts of bakery yeast wastewater use on weed communities and crops in an arid environment. Environ. Sci. Pollut. Res. 24 14957-14969. https://doi.org/10.1007/s11356-017-9115-1 [ Links ]
AHMED FF and RAGAB MM (2002) A new trial to stimulate growth and nutritional status of Picual olive transplants. In: The First International Conference on Olive Cultivation, Protection and Processing. 25-27. [ Links ]
AL-MEFLEH NK, OTHMAN YA, TADROS MJ, AL-ASSAF A and TALOZI S (2021) An assessment of treated greywater reuse in irrigation on growth and protein content of Prosopis and Albizia. Horticulturae. 3 38. https://doi.org/10.3390/horticulturae7030038 [ Links ]
ANANDARAJ B and DELAPIERRE LRA (2010) Studies on influence of bio-inoculants (Pseudomonas fluorescens, Rhizobium sp., Bacillus megaterium) in green gram. J. Biosci. Technol. 1 95-99. [ Links ]
BAKER NR and ROSENQVIST E (2004) Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 55 1607-1621. https://doi.org/10.1093/jxb/erh196 [ Links ]
BARGAZ A, LYAMLOULI K, CHTOUKI M, ZEROUAL Y and DHIBU D (2018) Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front. Microbiol. 9 1606. https://doi.org/10.3389/fmicb.2018.01606 [ Links ]
BORASTE A, VAMSI KK, JHADAV A, KHAIRNAR Y, GUPTA N, TRIVEDI S and JOSHI B (2009) Bio-fertilizers: A novel tool for agriculture. Int. J. Microbiol. Res. 1 23-31. https://doi.org/10.9735/0975-5276.1.2.23-31 [ Links ]
BASAK BB and BISWAS DR (2010) Co-inoculation of potassium solubilizing and nitrogen fixing bacteria on solubilization of waste mica and their effect on growth promotion and nutrient acquisition by a forage crop. Biol. Fertil. Soils. 46 641-648. https://doi.org/10.1007/s00374-010-0456-x [ Links ]
CORWIN DL and LESCH SM (2005) Apparent soil electrical conductivity measurements in agriculture. Comput. Electron. Agric. 46 11-43. https://doi.org/10.1016/j.compag.2004.10.005 [ Links ]
COSTA R, GOMES, NCM, KROGERRECKLENFORT E, OPELT K, BERG G and SMALLA K (2014) Pseudomonas community structure and antagonistic potential in the rhizosphere: insights gained by combining phylogenetic and functional gene-based analyses. Environ. Microbiol. 9 2260-2273. https://doi.org/10.1111/j.1462-2920.2007.01340.x [ Links ]
DALAHMEH SS, PELL M, HYLANDER LD, LALANDER C, VINNERAS B and JONSSON H (2013) Effects of changing hydraulic and organic loading rates on pollutant reduction in bark, charcoal and sand filters treating greywater. J. Environ. Manage. 132 338-345.https://doi.org/10.1016/j.jenvman.2013.11.005 [ Links ]
DELSHADI S, EBRAHIMI M and SHIRMOHAMMADI E (2017) Influence of plant-growth-promoting bacteria on germination, growth and nutrients' uptake of Onobrychis sativa L. under drought stress. J. Plant Interactions 12 200-208. https://doi.org/10.1080/17429145.2017.1316527 [ Links ]
DHANASEKAR R and DHANDAPANI R (2012) Effect of bio-fertilizers on the growth of Helianthus annuus. Int. J. Plants, Anim. Environ. Sci. 2 143-147. [ Links ]
DINKHA RF and KHAZRAGJI AL (1990) Nutrition and Fungus Function Science. University of Salahaddin, Ministry of High Education, Iraq. 139-147. [ Links ]
FAHAD S, HUSSAIN S, SAUD S, HASSAN S, CHAUHAN BS, KHAN, F and ALHARBY H (2016) Responses of rapid viscoanalyzer profile and other rice grain qualities to exogenously applied plant growth regulators under high day and high night temperatures. PLoS One. 11 1-13. https://doi.org/10.1371/journal.pone.0159590 [ Links ]
FINLEY S, BARRINGTON S and LYEW D (2009) Reuse of domestic greywater for the irrigation of food crops. Water Air Soil Pollut. 199 235-245. https://doi.org/10.1007/s11270-008-9874-x [ Links ]
GE C, RADNEZHAD H, ABARI MF, SADEGHI M and KASHI G (2016) Effect of biofertilizers and plant growth promoting bacteria on the growth characteristics of the herb Asparagus officinalis. Appl. Ecol. Environ. Res. 14 547-558. https://doi.org/10.15666/aeer [ Links ]
GLICK BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica. 1 1-16. https://doi.org/10.6064/2012/963401 [ Links ]
GOLLAN JR and WRIGHT JT (2006) Limited grazing pressure by native herbivores on the invasive seaweed Caulerpa taxifolia in a temperate Australian estuary. Mar. Freshwater Res. 57 685-694. https://doi.org/10.1071/MF05253 [ Links ]
HAO Y, CHEN Z, WANG Y, BLAND D, BUCK J, BROWN-GUEDIRA G and JOHNSON J (2011) Characterization of a major QTL for adult plant resistance to stripe rust in US soft red winter wheat. Theor. Appl. Genet. 123 1401-1411. https://doi.org/10.1007/s00122-011-1675-8 [ Links ]
HASANUZZAMAN M, NAHAR K and FUJITA M (2013) Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. Ecophysiol. Responses Plants Salt Stress. 1 25-87. https://doi.org/10.1007/978-1-4614-4747-4_2 [ Links ]
HASHEM M, OMRAN YM and NASHWA SALLAM M (2008) Efficacy of yeasts in the management of root-knot nematode (Meloidogyne incognita) in flame seedless grape vines and the consequent on the productivity of the vines. Biocontrol Sci. Technol. 18 357-375. https://doi.org/10.1080/09583150801950568 [ Links ]
JENSEN S, JOHNSON AL, JOHNSTON LH and SEGAL M (2004) Temporal coupling of spindle disassembly and cytokinesis is disrupted by deletion of LTE1 in budding yeast. Cell Cycle. 3 815-820. https://doi.org/10.4161/cc.3.6.950 [ Links ]
KUKLINSKY SJ, ARAUJO WL, MENDES R, GERALDI IO, KLEINER PA and AZEVEDO JL (2004) Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ. Microbiol. 6 1244-1251. https://doi.org/10.1111/j.1462-2920.2004.00658.x [ Links ]
LIN D and XING B (2007) Phytotoxicity of nanoparticles: inhibition of seed germination and root growth. Environ. Pollut. 150 243-250. https://doi.org/10.1016/j.envpol.2007.01.016 [ Links ]
LONHIENNE T, MASON MG, RAGAN MA, HUGENHOLTZ P, SCHMIDT S and PAUNGFOO-LONHIENNE C (2014) Yeast as a bio-fertilizer alters plant growth and morphology. Crop Sci. 54 785-790. https://doi.org/10.2135/cropsci2013.07.0488 [ Links ]
LUBBE E, RODDA N and NAIDOO S (2016) Effects of greywater irrigation on germination, growth and photosynthetic characteristics in selected African leafy vegetables. Water SA. 42 203-212. https://doi.org/10.4314/wsa.v42i2.04 [ Links ]
MARENCO RA, ANTEZANA-VERA SA and NASCIMENTO HCS (2009) Relationship between specific leaf area, leaf thickness, leaf water content and SPAD-502 readings in six Amazonian tree species. Photosynthetica. 47 184-190. https://doi.org/10.1007/s11099-009-0031-6 [ Links ]
MIRADA F and ISMAIL AM (2007) Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 99 1161-1173. https://doi.org/10.1093/aob/mcm052 [ Links ]
MISRA RK, PATEL JH and BAXI VR (2010) Reuse potential of laundry greywater for irrigation based on growth, water and nutrient use of tomato. J. Hydrol. 386 95-102. https://doi.org/10.1016/j.jhydrol.2010.03.010 [ Links ]
NAIDOO G (2009) Differential effects of nitrogen and phosphorus enrichment on growth of dwarf Avicennia marina mangroves. Aquat. Bot. 90 184-190. https://doi.org/10.1016/j.aquabot.2008.10.001 [ Links ]
NEL N and JACOBS HE (2019) Investigation into untreated greywater re-use practices by suburban households under threat of intermittent water supply. Water Sanit. Hyg. Dev. 9 627-634. https://doi.org/10.2166/washdev.2019.055 [ Links ]
PANHWAR QA, OTHMAN R, RAHMAN ZA, MEON S and ISMAIL MR (2012) Isolation and characterization of phosphate-solubilizing bacteria from aerobic rice. Afr. J. Biotechnol. 11 2711-2719. https://doi.org/10.5897/AJB10.2218 [ Links ]
PASZKO T, MUSZYNSKI P, MATERSKA M, BOJANOWSKA M, KOSTECKA M and JACKOWSKA I (2016) Adsorption and degradation of phenoxyalkanoic acid herbicides in soils: A review. Environ. Toxicol. Chem. 35 271-286. https://doi.org/10.1002/etc.3212 [ Links ]
PERUMAL A, VARGHESE B, GOVENDER P, RAMDHANI S and BERJAK P (2014) Effects of elevated temperatures on germination and subsequent seedling vigour in recalcitrant Trichilia emetica seeds. S. Afr. J. Bot. 90 153-162. https://doi.org/10.1016/j.sajb.2013.11.005 [ Links ]
PINTO U and MAHESHWARI BL (2015) Sustainable graywater reuse for residential landscape irrigation-a critical review. Chin. J. Populat. Resour. Environ. 3 250-264. https://doi.org/10.1080/10042857.2015.1059790 [ Links ]
PINTO U, MAHESHWARI BL and GREWAL HS (2010) Effects of greywater irrigation on plant growth, water use and soil properties. Resour. Conserv. Recy. 54 429-435. https://doi.org/10.1016/j.resconrec.2009.09.007 [ Links ]
PORTMAN ME, VDOV O, SCHUETZE M, GILBOA Y and FRIEDLER E (2022) Public perceptions and perspectives on alternative sources of water for reuse generated at the household level. Water Reuse. 1 157-174. https://doi.org/10.2166/wrd.2022.002 [ Links ]
QIN L, GUO S, AI W, TANG Y, CHENG Q and CHEN G (2013) Effect of salt stress on growth and physiology in amaranth and lettuce: Implications for bioregenerative life support system. Adv. Space Res. 51 476-482. https://doi.org/10.1016/j.asr.2012.09.025 [ Links ]
RADINGOANA MP, DUBE T and MAZVIMAVI D (2020) Progress in greywater reuse for home gardening: Opportunities, perceptions and challenges. Phys. Chem. Earth A/B/C. 116 102853. https://doi.org/10.1016/j.pce.2020.102853 [ Links ]
RODDA N, SALUKAZANA L, JACKSON SAF and SMITH MT (2011) Use of domestic greywater for small-scale irrigation of food crops: Effects on plants and soil. Phys. Chem. Earth A/B/C. 36 1051-1062. https://doi.org/10.1016/j.pce.2011.08.002 [ Links ]
SARHAN TZ, MOHAMMAD GH and TELI JA (2011) Effects of humic acid and bread yeast on growth and yield of eggplant (Solanum melongena L.). J. Agric. Sci. Technol. 1 1091-1096. https://doi.org/10.26682/ajuod.2020.23.2.17 [ Links ]
SHEHATA SA, FAWZY ZF and EL-RAMADY HR (2012) Response of cucumber plants to foliar application of chitosan and yeast under greenhouse conditions. Aust. J. Basic Appl. Sci. 6 63-71. http://www.ajbasweb.com/ajbas/2012/April/63-71.pdf [ Links ]
SINGH A (2021) A review of wastewater irrigation: Environmental implications. Resour. Conserv. Recy. 168 105454. https://doi.org/10.1016/j.resconrec.2021.105454 [ Links ]
TAHA ZS, SMIRA TA and SANAA MSR (2011) Effect of bread yeast application and seaweed extract on cucumber (Cucumis sativus L.) plant growth, yield and fruit quality. Mesopotamia J. Agric. 39 26-34. https://doi.org/10.33899/magrj.2011.30359 [ Links ]
TIWARI P, BAJPAI M, SINGH LK, MISHRA S and YADAV AN (2020) Phytohormones producing fungal communities: metabolic engineering for abiotic stress tolerance in crops. In: Agriculturally Important Fungi for Sustainable Agriculture. Springer, Cham. 171-197. https://doi.org/10.1007/978-3-030-45971-0_8 [ Links ]
UYSAL A, DEMIR S, SAYILGAN E, ERASLAN F and KUCUKYUMUK Z (2014) Optimization of struvite fertilizer formation from baker's yeast wastewater: growth and nutrition of maize and tomato plants. Environ. Sci. Pollut. Res. 21 3264-3274. https://doi.org/10.1007/s11356-013-2285-6 [ Links ]
VELHAL C, SANT M, GODBOLE T, WAGHMODE S and KULKARNI C (2014) Effect of Rhizobium based bio-fertilizer combined with Saccharomyces cerevisiae on the growth of Hyacinth Bean. Int. J. Plant Soil Sci. 3 959-968. https://doi.org/10.9734/IJPSS/2014/9284 [ Links ]
WANG Y, QIU L and HU M (2018) Application of yeast in the wastewater treatment. In E3S Web of Conferences (Vol. 53, p. 04025). EDP Sciences. https://doi.org/10.1051/e3sconf/20185304025 [ Links ]
WHO (World Health Organization) (2019) Safer water, better health. World Health Organization, Geneva. 1-80. [ Links ]
YANG M and ZHENG S (2014) Pollutant removal-oriented yeast biomass production from high-organic-strength industrial wastewater: A review. Biomass Bioenerg. 64 356-362. https://doi.org/10.1016/j.biombioe.2014.03.020 [ Links ]
YANG Q, ZHANG H, LI X, WANG Z, XU Y, REN S and WANG H (2013) Extracellular enzyme production and phylogenetic distribution of yeasts in wastewater treatment systems. Bioresour. Technol. 129 264-273. https://doi.org/10.1016/j.biortech.2012.11.101 [ Links ]
 Correspondence:
Correspondence:
KK Naidoo
Email: kuben@mut.ac.za
Received: 30 August 2021
Accepted: 4 July 2022
APPENDIX



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}