










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.48 n.2 Pretoria Apr. 2022
http://dx.doi.org/10.17159/wsa/2022.v48.i2.3918
REVIEW
Rising environmental temperatures and polluted surface waters: the prelude to the rise of mycoses in South Africa
Heidi SteffenI; Caylin BoschI; Gideon WolfaardtI, II; Alfred BothaI
IDepartment of Microbiology, Stellenbosch University, Private Bag X1, Matieland 7602, South Africa
IIDepartment of Chemistry and Biology, Ryerson University, Toronto, ON, Canada
ABSTRACT
South Africa's rivers are frequently used by communities lacking proper sanitation infrastructure for domestic purposes; however, these surface waters may pose a health risk to immunocompromised individuals due to the presence of opportunistic pathogenic fungi in the polluted water. Although only a few studies have focused on the presence of clinically relevant fungal species in South African rivers, many known opportunistic pathogenic species were found to be predominant in these waters. Furthermore, strong evidence exists that increased numbers of clinically relevant species may be observed in future due to fungi acquiring thermotolerance in response to the global increase in temperature. Thermotolerance is a major factor contributing to pathogenesis in fungi, due to the generally low tolerance of most fungi toward mammalian body temperatures. It is therefore contended that combinatorial effects of water pollution and rising environmental temperatures could lead to an increase in the incidence of mycoses in South Africa. This is especially concerning since a relatively large population of immunocompromised individuals, represented mostly by HIV-infected people, resides in the country.
Keywords: river water, pollution, global warming, mycoses, thermotolerance
INTRODUCTION
Despite the global increase in the incidence of fungal infections, which affect 1 billion individuals and cause nearly 1.6 million deaths annually (Brown et al., 2012; Bongomin et al., 2017; Almeida et al., 2019), only a few fungal taxa have been identified as causative agents of mycoses (Robert and Casadevall, 2009; Almeida et al., 2019). This phenomenon was ascribed to the relatively low maximum thermal tolerances of most fungi, which are lower than mammalian body temperatures (Robert and Casadevall, 2009). Nevertheless, fungi that can grow at mammalian body temperatures cause infections that may vary from being superficial, to subcutaneous to systemic (Dupont et al., 2000; Garber, 2001; Bicanic and Harrison, 2014; Almeida et al., 2019). Incidents of such mycoses are scrupulously reported in developed countries; however, the opposite is true for developing countries, where the statistics on fungal infections are likely underestimated to a great degree. Furthermore, the largest populations of individuals suffering from HIV/AIDS reside in developing countries (Vearey, 2011; UNAIDS, 2017; Schwartz and Denning, 2019), which could potentially increase the incidence of fungal diseases in these regions, since such immunocompromised individuals are at greater risk of acquiring fungal infections (Dupont et al., 1994; Schwartz and Denning, 2019). This is especially relevant to South Africa, with its disproportionately large population of HIV-infected people (UNAIDS, 2021; Schwartz and Denning, 2019; Weimann and Oni, 2019).
There are various mycoses known to be associated with immunocompromised individuals (Guarner, 2017). Candidiasis, caused by ascomycetous yeasts belonging to the genus Candida, is the most common opportunistic fungal infection among HIV-infected patients (Garber, 2001; Moran et al. 2012; Guarner, 2017). Of the estimated 3.2 million fungal infections reported annually in South Africa, candidiasis represents nearly 2 million (Schwartz and Denning, 2019). Other important mycoses include invasive aspergillosis, cryptococcal meningitis, histoplasmosis, as well as Pneumocystis- and dermatophyte infections (Guarner, 2017). The causative agents of these infections are regularly isolated from natural and anthropogenic environments (Garber, 2001).
Although mycoses may be obtained from nature, hospital-acquired fungal infections are most common and represent the majority of candidiasis and other fungal disease cases reported in literature (Garber, 2001; Suleyman and Alangaden, 2016; Moazeni et al., 2018). These clinical cases often include fungal infections as a result of contaminated catheters, needles, and decreased immunity during organ transplants (Suleyman and Alangaden, 2016). Additionally, infections caused by members of the genus Aspergillus are acquired due to the exposure of airborne propagules associated with, among others, construction sites and ventilation systems of hospitals (Cortez et al., 2008; Benedict et al., 2017). Aspergillosis may also develop due to close interaction with certain contaminated foodstuffs and vegetation (Walsh and Dixon, 1989). Other habitats, such as decaying trees and pigeon guano, harbour notorious members of Cryptococcus, which can infect humans in proximity via inhalation (Restrepo et al., 2000; Garber, 2001). Additionally, representatives of pathogenic fungal species were recovered from polluted river systems (Luplertlop et al., 2016; Postma, 2016; Assress et al., 2019). Although pathogenic fungi are plentiful in nature, studies commonly focus on nosocomial mycoses (Jarvis, 1995; Guinea, 2014). The prevalence and interactions of opportunistic fungal pathogens within the natural environment, such as rivers, are rarely studied, despite indications that fungal virulence results from adaptations that have evolved for protection against adverse conditions in nature (Steenbergen et al., 2001; Bosch et al., 2020). These conditions include rising environmental temperatures and other anomalies as a result of climate change, and have been linked with increases in the prevalence of fungal diseases in mammals, including humans (Garcia-Solache and Casadevall, 2010; Araújo et al., 2017).
Research on the ecology of opportunistic fungi in environments conducive to their proliferation, such as river systems, is therefore expedient. Focus should be given to developing countries such as South Africa, with its large population of immunocompromised individuals (UNAIDS, 2021) and inadequate infrastructure accompanied by rapid urbanisation (Schwartz and Denning, 2019). With the ultimate goal of obtaining an indication of the potential effects of climate change and anthropogenic pollution on the ecology of opportunistic pathogenic fungi in South African rivers, the aim of this literature review was to survey existing knowledge on the occurrence of these fungi in the river systems of the region. Additionally, this review reflects on the potential effect of increased environmental temperatures, in combination with pollution, on the biology of opportunistic pathogenic fungi occurring in these rivers.
POLLUTED SURFACE WATERS
Rivers may become polluted as a result of various anthropogenic activities: agricultural practices, industrial processes, expanding urban communities with poor sanitation infrastructure, as well as faulty wastewater treatment operations (Vearey, 2011; Wang et al., 2012; Glinska-Lewczuk et al., 2016; Liyanage and Yamada, 2017; Cullis et al., 2019). These may largely contribute to the fungal load in water due to the nutrient-rich characteristics of most pollutants. For several decades, the quality of water, whether it be used for drinking, ablutions, or irrigation purposes, was evaluated by determining the microbiological load thereof (Havelaar et al., 1986). Bacterial indicators have been, and are being, extensively used to determine water quality, especially in relation to the level of faecal pollution (Havelaar et al., 1986; Haack et al., 2009). Only relatively recently were studies conducted linking fungal numbers to pollution levels in river systems (De Almeida, 2005; Brandão et al., 2010; Medeiros et al., 2012; Stone et al., 2013). Previously the only indicators of fungal contaminants used were taste, odour and the number of culturable moulds in the water (Doggett, 2000). While very few studies were aimed at investigating the link between fungal numbers and pollution, several authors have studied fungal diversity in a wide range of surface water sources - including tap water, rivers, lakes and estuaries (Kwasnieskwa, 1988; Hageskal et al., 2009; Pereira et al., 2009; Magwaza et al., 2017; Assress et al., 2019). Some of the fungi identified during these studies include known clinically relevant yeasts and filamentous fungi.
CLINICALLY RELEVANT FUNGI
Pathogenic and opportunistic pathogenic fungi are ubiquitous in the environment (Restrepo et al., 2000; Gostincar et al., 2011; Babic et al., 2017). While the occurrence of some of these fungi has been studied extensively in atmospheric (Newbound et al., 2009) and terrestrial environments including animals, plants and soils (Restrepo et al., 2000; Gostincar et al., 2011), relatively little knowledge exists on the ecology of clinically relevant fungi in aquatic environments, with only a few studies focusing on polluted water. Regardless of the lack of available information, some findings have allowed researchers to hypothesise that fungi, particularly yeasts, have the potential to be used as indicators of pollution in rivers and other water bodies (De Almeida, 2005; Brandão et al., 2010; Medeiros et al., 2012; Stone et al., 2013; Postma, 2016). However, it should also be made a priority to determine the pathogenic fungal taxa occurring in polluted rivers, since individuals utilizing these water sources are at risk of acquiring mycoses (Weimann and Oni, 2019).
Filamentous fungi
The most common water-borne filamentous fungi are members of the genera Penicillium, Trichoderma, Acremonium, Cladosporium, Aspergillus, Fusarium and Mucor (Babic et al., 2017). Representatives of the latter three genera are considered opportunistic, causing invasive fungal infections in immune-deficient individuals. Members of the remaining genera cause allergies, as well as subcutaneous and superficial infections, especially among those suffering from immunodeficiency. Despite the low health risk, compared to invasive infections, superficial infections remain concerning due to the associated negative impact on quality of life (Weimann and Oni, 2019).
To date, no substantial evidence exists on the presence of pathogenic filamentous fungi in the river systems of South Africa. This, however, does not negate the danger associated with the presence of these fungi in rivers, but rather encourages researchers to investigate neglected environments (Schwartz and Denning, 2019). Investigating such understudied environments is of particular importance considering current evidence of disease-causing filamentous fungi detected in different European water sources - especially tap water (Novak Babic et al., 2018). It is well known that surface waters are sources for drinking and tap water (UN Water, 2006; WHO, 2017). These water sources are treated before distribution, but the treatment processes are not feasible for rivers, lakes and other surface waters that are used as primary water sources by some communities (Colvin et al., 2016; Edokpayi et al., 2018). Recent studies, however, have revealed that fungi can survive conventional treatment processes and persist in tap water, as was observed in some hospitals (Arroyo et al., 2020; Caggiano et al., 2020). Nevertheless, the fungal composition of treated surface water could be significantly different from untreated surface water and therefore in this review only fungi present in rivers and other natural water sources were evaluated.
Globally, two major fungal groups were detected in the few river systems that were studied: Mucoromycota and Ascomycota (Arvanitidou et al., 2005; Pietryczuk et al., 2018; Machido et al., 2015). Mucoralean fungi were found to be the only representatives of the phylum Mucoromycota that occurred within rivers in Greece. These included members of Mucor, Absidia and Rhizopus (Arvanitidou et al., 2005). Species within these genera cause mucormycosis - an invasive fungal infection which can manifest in the lungs, cutaneous and subcutaneous dermis, nose cavities, brain, intestines and bloodstream (Spellberg and Maertens, 2019). Recently, cases of mucormycosis have increasingly been reported among patients suffering from Covid-19 (Garg et al., 2021; Werthman-Ehrenreich et al., 2021). Members of Absidia have also caused infections in individuals suffering from burn wounds (Christiaens et al., 2005). These infections are invasive, invading organs and the bloodstream, with detrimental health outcomes (Harrison and Brouwer, 2009).
It must be noted that although immunosuppressed individuals are especially susceptible to invasive fungal infections, rare cases of rhinocerebral mucormycosis, as well as other mycoses, have been observed in immunocompetent patients (Hussain et al., 1995; Leyngold et al., 2014). In contrast, non-invasive superficial fungal infections caused by a variety of different fungi are very common among patients with immune deficiencies (Huang et al., 2004), and noticeable superficial infections on epidermal regions are considered a presenting feature of individuals diagnosed with AIDS (Lohoue Petmy et al., 2004; Benedict et al., 2017).
Some fungi that are known to occur in rivers are capable of causing both invasive and superficial infections (Arvanitidou et al., 2005). These include members of the genera Aspergillus and Penicillium belonging to the orders Eurotiales; Alternaria and Curvularia of the order Pleosporales; as well as Fusarium and Verticillium belonging to Hypocreales. Representatives ofthe genera Aspergillus and Penicillium spp. were among the most frequently isolated filamentous fungi found during a survey of rivers in Greece. Although many beneficial strains representing Penicillium exist that are used in the food and pharmaceutical industries (Sousa et al., 2001; Elander, 2003), it is known that some members of this genus can cause keratitis, pneumonia, endocarditis, necrotising esophagitis, endophthalmitis, urinary tract infections and peritonises (Hu et al., 2013). Aspergillus fumigatus and Aspergillus niger that were found in Polish rivers (Pietryczuk et al., 2018) are both associated with aspergillosis, which is one of the leading causes of death among AIDS individuals (GAFFI, 2017). Recently, cases of Covid-19 associated pulmonary chronic aspergillosis have also been reported (Arastehfar et al., 2020).
Verticillium, another genus harbouring waterborne fungi (Schiavano et al., 2014; Novak Babic et al., 2018), is represented by species associated with peritonises, as well as subcutaneous infections. The latter ensues when a microorganism breaches the epidermal barrier and migrates into the innermost layer of skin where blood vessels and nerves are located (Kujath and Kujath, 2010). While it is known that some bacteria produce proteins that enable infiltration of blood vessels, little information is available for fungi presenting the same mechanisms (Lipke, 2018). Nonetheless, in the probable event that some pathogenic fungi harbour these proteins, the proximity between the microorganisms and blood vessels in subcutaneous tissue could lead to systemic infections where the infectious agents are disseminated to other parts of the body. The subcutaneous tissue is also found directly on top of the musculoskeletal tissue, an area that some representatives of Fusarium are known to infect (Koehler et al., 2016). Both Fusarium and Verticillium spp. have been associated with superficial and invasive infections (Kujath and Kujath, 2010; Koehler et al., 2016), while other members of the Hypocreales from rivers, such as species of Acremonium and Cylindrocarpon, are more commonly known to cause superficial infections, such as mycetoma (Welsh et al., 2007; Arvanitidou et al., 2005; Pietryczuk et al., 2018).
Of the pleosporalean fungi that were found in Greek rivers, members of Curvularia and Pyrenochaeta were rarely encountered, whereas representatives of Alternaria were frequently isolated (Arvanitidou et al., 2005). Curvularia and Pyrenochaeta harbour species associated with cutaneous infections and invasive fungemia. Alternaria species, however, are involved in other less invasive infections such as rhinosinusitis, onychomycosis and oculomycosis (Pastor and Guarro, 2008). Additionally, in a recent study done by Pietryczuk et al. (2018), Alternaria alternata was identified in 4 out of 5 rivers surveyed in Poland.
Onygenalean fungi, which represent members that cause tineas, were also detected in natural surface waters (Arvanitidou et al., 2005; Machido et al., 2015; Novak Babic et al. 2018). Two genera, Emmonsia and Chrysosporium, were frequently encountered in Greek rivers (Arvanitidou et al., 2005) and represent fungi typically associated with disseminated- or superficial infections, respectively (Kenyon et al., 2013; Mijiti et al., 2017). Other members of the Onygenales which were isolated from river systems in Europe and Africa were representatives of Microsporum and Trichophyton (Arvanitidou et al., 2005; Machido et al., 2015), genera that include keratolytic fungi and are frequently associated with tineas (Schwinn et al., 1995; Brito-Santos et al., 2017). For example, Microsporum canis, which has been isolated from surface waters, as well as Trichophyton tonsurans and Trichophyton violaceum that have been isolated from rivers (Machido et al., 2015; Pietryczuk et al., 2018; Novak Babic et al., 2018), are all known to cause tinea capitis (Sombatmaithai et al., 2015; Pasquetti et al., 2017; Morales et al., 2019).
Other ascomycetous filamentous fungi occurring in surface waters that have mostly been associated with cutaneous infections are members of the order Chaetothyriales (Novak Babic et al., 2018; Pietryczuk et al., 2018). A chaetothyrialean fungus, Exophalia dermatitidis, associated with both cutaneous infections, as well as with respiratory infections in cystic fibrosis patients, was isolated from Polish rivers (Pietryczuk et al., 2018). Also isolated from these rivers were microascalean fungi belonging to the genus Scopulariopsis. Representatives of this genus are known to be the causative agents for different mycoses, ranging from superficial, to pulmonary and systemic infections (Iwen et al., 2012; SandovalDenis et al., 2013). It is important to mention that Microascales also harbours emerging pathogens such as members of the genus Scedosporium, which are commonly associated with wastewater sources (Skiada et al., 2017). Considering that wastewater is a major component of the pollution in rivers, further investigation into the diversity of Scedosporium spp. and other opportunistic filamentous fungi in natural surface waters is imminent.
Yeasts
The majority of fungal infections are caused by yeasts (Miceli et al., 2011; Bongomin et al., 2017; Lamoth et al., 2018; Ocansey et al., 2019; Schwartz and Denning, 2019). These opportunistic unicellular fungi usually belong to genera such as Candida and Rhodotorula, which were among the prevalent yeasts occurring in polluted surface waters (Slävikovä and Vadkertiovä, 1997; De Almeida et al., 2005; Gadanho et al., 2006; Coelho et al., 2010; Medeiros et al., 2012; Pietryczuk et al., 2014; Monapathi et al., 2020). The invasive infections caused by these yeasts often have devastating effects on human organs, including the liver, heart, lungs, and brain. These infections, however, are rare among immunocompetent individuals and occur mostly among immunocompromised patients, such as those suffering from HIV/AIDS (Low and Rotstein, 2011).
Compared to filamentous fungi, more literature is available on the prevalence and presence of unicellular fungi in surface waters (Buzzini et al., 2017). This can be attributed to the non-spreading nature of yeast colonies on microbiological media, enabling researchers to more readily enumerate yeasts and link their numbers to environmental factors (Slävikovä and Vadkertiovä, 1997; De Almeida, 2005; Gadanho et al., 2006; Coelho et al., 2010; Medeiros et al., 2012; Stone et al., 2012; Van Wyk et al., 2012). Additionally, more yeast-related surveys could have been conducted in water sources than what were carried out for filamentous fungi, due to the disproportional disease burden associated with yeasts and the implicated higher health risk (Miceli et al., 2011; Bongomin et al., 2017; Lamoth et al., 2018; Ocansey et al., 2019; Schwartz and Denning, 2019).
A number of surveys on yeasts occurring in rivers was conducted across the globe (Table 1), yet there is still a paucity of published information available on yeasts associated with river systems in South Africa (Table 2). Limited data originating from only two provinces have been published thus far: Western Cape and North-Western Province (Stone et al., 2012; Van Wyk et al., 2013; Monapathi et al., 2017; Monapathi et al., 2020).
The most dominant of the opportunistic pathogenic yeasts identified in rivers of South Africa (Table 2), as well as other countries (Table 1), were found to be ascomycetous yeasts, especially members of the genus Candida (De Almeida, 2005; Coelho et al., 2010; Medeiros et al., 2012; Van Wyk et al., 2012; Monapathi et al., 2017). Whether the predominance of these clinically relevant ascomycetes is associated with certain environmental parameters is still unclear. Considering the positive correlation observed between pollution indicators and yeast numbers, as well as the prevalence of ascomycetes in some studies, research on the link between yeasts and physicochemical parameters is imminent (Slavikova and Vadkertiova, 1997; Gadanho et al., 2006; Coelho et al., 2010; Medeiros et al., 2012; Stone et al., 2012; Van Wyk et al., 2012). Thus far, when researchers analysed the pollution levels of river water, results varied: some authors reported higher than normal concentrations of nitrogen and phosphate (Slavikova and Vadkertiova, 1997; Medeiros et al., 2012), others reported that yeast numbers correlate with temperature and pH (Gadanho et al., 2006; Van Wyk et al., 2012) or that yeast prevalence is related to faecal coliform numbers (Coelho et al., 2010; Stone et al., 2012).
The South African rivers that were investigated were considered to be diffusely polluted, contaminated with exceedingly high levels of dissolved solids, nitrates and phosphates (Van Wyk et al., 2012; Monapathi et al., 2017), as well as faecal matter (Stone et al., 2012). The latter is of considerable concern since members of clinically relevant species including Pichia kudriavzevii (syn. Candida krusei), Candida tropicalis, Candia parapsilosis, Candida rugosa, Papiliotrema laurentii (syn. Cryptococcus laurentii), Cyberlindnera jadinii (syn. Candida utilis), Meyerozyma guilliermondii (syn. Candida guilliermondii), Rhodotorula glutinis and Rhodotorula mucilaginosa (syn. Rhodotorula rubra) have previously been associated with sewage and sewage-polluted water (Cooke et al., 1960). In South Africa, many municipalities have neglected the maintenance of sewage treatment infrastructure, causing the release ofraw and partially treated sewage into river systems (Herbig, 2019). The heavy load of excrement and other pollutants in sewage is well known to increase the quantity of organic carbon, including chemicals and other substrates that sustain growth of potentially harmful microorganisms (Liu et al., 2015). In tandem, anthropogenic waste depository from individuals residing near riverbanks extends the list of pollutants entering surface waters. Among the anthropogenic pollution entering river systems are chemicals such as antibiotics and even antifungals (Chitescu et al., 2015). While the above-mentioned aberrations with regard to physicochemical parameters are known to indicate pollution in river systems, no attempt was made thus far to correlate these parameters with the numbers of pathogenic yeast species in these polluted waters.
To enumerate and isolate clinically relevant fungi, including yeasts, from surface waters, an incubation temperature of 37°C was employed by some authors, (Buck and Bubucis, 1978; De Almeida, 2005; Yamaguchi et al., 2007; Coelho et al., 2010). Others incubated their isolation plates at room temperature to determine overall yeast numbers in surface waters. Interestingly, Coelho et al. (2010) identified yeasts incubated at both room temperature and 37°C, and found that the numbers of the predominant species cultivated at the latter temperature correlated with E. coli numbers and therefore with the level of faecal contamination. Yeast numbers occurring on isolation plates that were incubated at room temperature, however, showed no significant correlation with faecal coliform numbers.
The positive correlation between faecal coliform numbers and yeast counts obtained after incubation at 37°C, further suggests that these unicellular fungi are allochthonous - originating from other sources and subsequently introduced to the river water (Weimann and Oni, 2019; Arvanitidou et al., 2002). A potential source of these yeasts can be anthropogenic pollution (Dynowska, 1997; Coelho et al., 2010; Herbig and Meissner, 2019; Weimann and Oni, 2019). While the inhabitants of rural communities may be one of the sources of river pollution, they also tend to be at risk, especially individuals predisposed to infection because of immune deficiencies (Vearey, 2011).
It is disconcerting that some of the most clinically relevant species of Candida were isolated from South African rivers (Table 2): C. albicans, C. tropicalis, C. glabrata and C. parapsilosis (Miceli et al., 2011). The same representatives were isolated from surface waters in other countries such as Portugal, Greece, Poland and Brazil (Arvanitidou et al., 2005; De Almeida, 2005; Coelho et al., 2010; Medeiros et al., 2012, Pietryczuk et al., 2018). The occurrence of renowned clinically relevant yeasts in different geographical regions of the world emphasises the need for more ecological surveys for the presence of these unicellular fungi in ecosystems. To understand why these yeasts persist and grow within ecosystems, such surveys should be accompanied by analyses of anthropological activity including assessment of pollution indicators and measurement of physicochemical parameters. The latter includes parameters such as the nutrient levels, pH, as well as the temperature of surface waters.
RISING ENVIRONMENTAL TEMPERATURES
Life on Earth is sustained by the natural greenhouse gases found in the atmosphere; however, industrialisation and other anthropogenic activities involving fossil fuel combustion have impinged on the natural environment and largely contributed to climate change (IPCC, 2018; IPCC, 2021). Since the industrial revolution that started in 1760, the increased emissions of greenhouse- and other deleterious gases have contributed to a rise in global temperature. In addition, the exponential increase in the population caused agriculture to expand in order to meet the high food demand. Livestock farming, for instance, is now recognised as another contributor to global warming due to deforestation (land to farm) as well as an increase in released methane (Ilea, 2009). This has led to the total increase of an estimated 1°C in global temperatures and a further increase of up to 2°C has been hypothesised for future decades (IPCC, 2018; IPCC, 2021). In accordance with global predictions, South Africa has seen a 2°C increase since the industrial revolution (1760) and it is projected that a 0.12-0.5°C/ decade rise will be observed in future (USAID, 2015).
This world-wide rise in temperatures has been collectively referred to as global warming (Lineman et al., 2015). Disastrous consequences of this anthropogenically influenced phenomenon include the loss of ice in Antarctica and Greenland, accompanied by a rise in sea levels (Hansen et al., 2016; IPCC, 2021). Thus far, global warming has had detrimental effects on ecosystems and biomes across the globe. Increased temperatures were found to heighten the threat of species extinction within many habitats, such as montane forests and other terrestrial environments (Malcolm et al., 2006). Recent studies have shown that climate change also increases the temperature of freshwater aquatic environments (Morrison et al., 2002; Van Vliet et al., 2011; Chen et al., 2016; Nusslé et al., 2015; Pohle et al., 2019; Kedra, 2020; Liu et al., 2020). The rising river water temperatures in some of these studies were found to be congruent with the rise in air temperatures, suggesting a direct link between thermally altered waters and global warming (Van Vliet et al., 2011; Chen et al., 2016; Pohle et al., 2018; Kedra, 2020; Liu et al., 2020). As such, excess heat because of anthropogenic activities - also referred to as thermal pollution - may therefore be one of the main causes for increased water temperatures (Nordell et al., 2003; Verones et al., 2010; Liu et al., 2020).
The effects on the microorganisms present in thermally altered waters have not been fully described, despite their crucial roles in biogeochemical cycling and other processes essential for a fully functional and diverse ecosystem (Sigee, 2005). Fungi, in particular, are largely understudied in aquatic ecosystems, with little to no knowledge available on how thermal pollution in water affects these microbes (Grossart et al., 2019). However, potential links between the emergence of pathogenic fungi and rising ambient temperatures have been discussed by some mycologists (Garcia-Solache and Casadevall, 2010; Casadevall et al., 2020), and positive correlations have been observed between the presence of fungal pathogens in river systems and water pollution levels (De Almeida, 2005; Brandão et al., 2010; Medeiros et al., 2012; Stone et al., 2012; Postma, 2016). Thus, a holistic approach is needed to investigate the combinatorial effects of both water pollution and global warming on the emergence of fungal pathogens.
GLOBAL WARMING AND EMERGING MYCOSES
Fungi are well known infectious agents of various organisms, including plants and mammals as well as amphibians (Garber, 2001). Batrachochytrium dendrobatidis is a good example of a notorious causative agent of chytridiomycosis among many frog species. Since the emergence of this fungus, researchers have contended that the decline in frog species due to this pathogen is linked to the global rise in temperature (Longcore et al., 1999; Wake and Vredenburg, 2008; Fisher et al., 2009). Although the correlation analysis linking global warming to the decline of amphibian species has been queried by some (Rohr et al., 2008), many other researchers have emphasised the role of rising temperatures in species extinction (Pounds et al., 2006; Alford et al., 2007; D'Amen and Bombi, 2009). These studies employed statistical models to determine correlations between rising temperatures and declining species numbers (Pounds et al., 2006; Alford et al., 2007; Rohr et al., 2008; D'Amen and Bombi, 2009), but failed to consider other biological factors. Recently, however, researchers have begun to study the specific relationship between host and pathogen during climate changes (Cohen et al., 2017; Neely et al., 2020). The thermal mismatch theory was formulated, which states that as environmental conditions deviate from the optimal conditions required for host survival, susceptibility towards infectious agents will increase (Chen et al., 2011; Cohen et al., 2017). Although this theory is limited to cold-adapted hosts, it has provided opportunities to discover important traits of the associated pathogens. For example, it was found that microbial symbionts, specifically pathogens, have a broader thermal breadth than their hosts (Neely et al., 2020), due to the higher mass-specific metabolic rates of smaller organisms, thus allowing for a more rapid adaptation to environmental change (Cohen et al., 2017). Understanding these and similar microbial characteristics provides opportunities to determine how environmental fungi may adapt to climate change and evolve to survive the conditions of a mammalian host. It must be noted, however, that little research has been conducted on intraspecies differences in thermal breadth, between pathogenic fungi recovered from natural environments and that of laboratory or clinical strains. Such studies may provide better insight into how environmental stressors like elevated temperatures could facilitate adaptation.
Garcia-Solache and Casadevall (2010) presented an important hypothesis averring that mammalian fungal infections will increase as a result of global warming. This hypothesis requires an understanding of how pathogens adapt to specifically overcome the restrictions associated with the mammalian body. In humans, fungal infections have mainly occurred in individuals with immune deficiencies, while relatively few cases were recorded among immunocompetent persons (Badiee and Zare, 2017). This is largely owing to the complex and effective immune systems unique to jawed vertebrates, which provide extensive protection against fungal pathogens (Shoham and Levitz, 2005). In addition, an equally effective preventative mechanism is the thermal restriction zone. Endothermic animals can regulate body temperature in response to infection during which basal temperatures increase in a process known as fever. Microorganisms such as unicellular- and filamentous fungi have a relatively low tolerance to high temperatures and therefore deteriorate during fever conditions (Robert and Casadevall, 2009). The high thermal susceptibility, in addition to the intolerance of the already high body temperature, effectively prevents fungal pathogens from frequently causing invasive infections. Thus, with the increase in ambient temperatures, and the potential ability of fungi to adapt to increased environmental temperatures, global warming threatens to expedite the emergence of mycoses by introducing more thermally tolerant fungi.
Two major restrictions prevent an exponential increase of mycoses cases in humans: a complex, highly adaptable immune system (Blanco and Garcia, 2008) and elevated body temperatures. The law of parsimony (Sober, 1981; McMeekin et al., 2008) would dictate that investigating the microorganism's ability to adapt to the thermal restrictions in mammals could provide a better indication of potential virulence than investigating its ability to evade host immunity. Therefore, this review will continue by discussing available fundamental knowledge on fungal thermotolerance.
THERMALLY TOLERANT FUNGI
For microorganisms to be considered thermotolerant, they should be able to withstand temperatures that exceed the population's optimal growth temperature (Robert et al., 2015). The upper thermal limit can range from 35°C to 62°C depending on the type of microorganism (Tansey and Brock, 1972; Maheshwari et al., 2000). Many bacterial species have been identified as thermally tolerant and some groups have also been recognised as thermophilic. Most fungi, however, have failed to attain this characteristic and remain susceptible to higher temperatures (Robert and Casadevall, 2009). Exceptions to this include some industrially important yeasts that tolerate high temperatures, which is essential for the success of some processes. Understanding the mechanisms of thermotolerance in these yeasts is crucial for industries to optimise bread and wine fermentation as well as biofuel production (Parapouli et al., 2020). These mechanisms have thus been scrupulously investigated due to their economic value. Conversely, studies investigating thermotolerant fungi associated with infection and disease are lacking.
Robert et al. (2015) investigated thermotolerance among a wide range of yeast species (CBS Culture Collection, Netherlands) and highlighted that many were able to grow at temperatures above 35°C. Although these thermotolerant species represented a polyphyletic group of fungi, it was noted that thermotolerance was significantly more common among the ascomycetous yeasts than among the basidiomycetes. However, the maximum temperature of growth (Tmax) for basidiomycetous yeast species has increased over the past decades. This suggests that basidiomycetes could be increasingly implicated in future novel fungal infections. Regardless, increasing Tmax trends were observed for all yeasts investigated over the past few decades, potentially due to adaptation to the global change in temperature (Robert et al., 2015). Environmental stressors, such as temperature, are common agents in the evolution of microorganisms (Baquero, 2009) and it was contended that acquired thermotolerance among fungi could increase the incidence of infections (Araújo et al., 2017). In knowing the original environmental habitats of potentially pathogenic fungi and gaining a fundamental comprehension of their thermotolerant nature, researchers could in future determine the risk of these microorganisms becoming emerging pathogens (Araújo et al., 2017; Jackson et al., 2019). Well-known examples of such thermotolerant fungi that are contended to have emerged from the environment as pathogens are Candida auris, as well as the pathogenic cryptococci and aspergilli.
Candida auris
Candida auris is a multidrug-resistant pathogen, which is also known for its nosocomial transmission (Rhodes and Fisher, 2019). The simultaneous rise of infections caused by four genetically discrete clades of C. auris, in distinct geographical regions, has raised questions regarding the cause of the emergence of this pathogen (Kean et al., 2020). It was proposed that C. auris could be the model organism explaining emergence of novel mycoses as a result of global warming (Casadevall et al., 2020). Candida auris sp. nov. was first isolated from a hospitalised patient's ear (Satoh et al., 2009), an organ that represents cooler environments compared to the rest of the human bo dy. It was therefore contended that the yeast could have gradually gained temperature tolerance while inhabiting the ear, possibly enabling invasive infection and subsequently causing many outbreaks of C. auris-related candidiasis (Casadevall et al., 2020; Jackson et al., 2019; Kean et al., 2020). Alarmingly, the frequency of these outbreaks and severity of C. auris infections has highlighted this yeast's epidemic potential (Meis and Chowdhary, 2019). Recently, cases of C. auris infections have increasingly been reported among patients suffering from Covid-19 (Rodriguez, et al., 2020; Almeida et al., 2021; Villanueva-Lozano et al., 2021).
With little knowledge available on the natural origin of C. auris, determining the cause of this yeast's emergence remains cumbersome. Studies have suggested that C. auris is of environmental origin and emerged as a pathogen only after being introduced to healthcare systems by patients carrying the yeast (Casadevall et al., 2020; Kean et al., 2020). Some hypothesise that, as observed for certain pathogenic cryptococcal species (Moschetti et al., 2017), C. auris may have initially inhabited birds (body temperature of 42°C) where it consequently attained its thermotolerance. The migration of these animals to areas where they are in close contact with humans could also explain the emergence of C. auris at distinct geographical regions (Moschetti et al., 2017; Kean et al., 2020). Another proposed habitat for this yeast is wetland systems (Casadevall et al., 2019) that are known to have anaerobic zones, in which C. auris would be able to exist. This species also tolerates elevated salt concentrations, which may explain why C. auris was recently isolated from a salt marsh (Arora et al., 2021). Except for one isolate, most of the environmental isolates were found to be antifungal-resistant and grew well at 37°C and 42°C. Subsequent phylogenetic analyses revealed the existence of single nucleotide polymorphism differences between the environmental isolates and clinical isolates belonging to the same clade. The authors suggested that, once introduced into anthropogenic habitats, C. auris is capable of rapid adaptation to obtain both antifungal resistance and thermotolerance. They concurred, however, that more research is required to obtain a better understanding of the genetic diversity of this species in natural habitats. To date this species has not been detected in any other aquatic environment; however, frequent isolations of Candida spp. from aquatic environments (Table 1) as well as the recent findings of Arora et al. (2021) indicate that C. auris will potentially be found in more of these environments.
Cryptococcus spp.
Of the wide diversity of Cryptococcus spp. identified so far, infection is mainly caused by representatives of the Cryptococcus gattii / Cryptococcus neoformans species complex (Hagen et al., 2015; May et al., 2016). Their ability to withstand the mammalian core body temperature and to evade the immune response are key characteristics that facilitate systemic infections in humans and other mammals (Perfect, 2006) and it is known that cryptococcosis is one of the leading causes of mortality among immunocompromised individuals (UNAIDS, 2021). This infection is acquired after exposure to pathogenic cryptococci present in natural environments (May et al., 2016). Common natural habitats for these microorganisms are decaying plant material, such as trees, as well as the bodies and excreta of avian species (Ellis and Pfeiffer, 1990; Ellis and Pfeiffer, 1992; Lazera et al., 1996; Nielsen et al., 2007). The latter suggests that birds can serve as vectors for these cryptococci and subsequently distribute the pathogen to urban areas where interaction with humans might occur. These yeasts are introduced to the mammalian body through inhalation of aerosolised propagules (Perfect, 2006).
Recently, transcriptional regulation and signalling pathways were uncovered in pathogenic cryptococci that increase the virulence of these yeasts; many of these pathways are also up-regulated at higher temperatures (Juvvadi et al., 2014; Chatterjee and Tatu, 2017; Oliveira et al., 2020; Toplis et al., 2020; Bosch et al., 2021). Typical virulence factors observed in these yeasts include capsule enlargement and melanin production, which enable successful evasion of host immunity. Some of these virulence factors were also observed in non-pathogenic species (Petter et al., 2001; Watkins et al., 2017). For example, the basidiomycetous yeast, Saitozyma podzolica (syn. Cryptococcus podzolicus), produces virulence factors but is unable to cause disease in mammals due to its sensitivity towards higher temperatures (Petter et al., 2001). With the increased Tmaxobserved among basidiomycetes (Robert et al., 2015), it is plausible that more non-pathogenic species of the basidiomycetous yeasts will cause fungal diseases in future in the event of acquired thermotolerance. Similar phenomena may also be observed for other fungal species; however, without establishing the level of thermotolerance of these potential pathogens, one cannot determine the risk of such a fungus becoming an emergent pathogen.
Aspergillus spp.
For most fungi, the thermal limit for survival is low and rarely breaches the basal temperatures of mammalian species (Robert and Casadevall, 2009). An exception, however, is the upper limit of some species of Aspergillus, a genus of filamentous fungi of which some representatives cause respiratory diseases among both immunocompromised and immunocompetent individuals (Kousha et al., 2011; Badiee and Zare, 2017). Members of this genus, such as A. fumigatus, can survive up to 70°C and actively grow at 37°C (Albrecht et al., 2010). The ability to grow at such high temperatures would suggest that this organism might be considered thermophilic; however, A. fumigatus and other species within this genus are capable of rapidly growing at mesophilic temperatures, allowing them to be predominant in natural environments (Paulussen et al., 2017). This characteristic distinguishes Aspergillus from thermophilic groups and delineates aspergilli as thermotolerant, with a broad thermal range. The ability to grow at mammalian body temperature enables the above-mentioned aspergilli, such as A. fumigatus, to infect humans even during fever conditions (Bhabhra and Askew, 2005). Aspergillosis is known for manifesting in the respiratory system, similar to what is seen for cryptococcosis. The spores of Aspergillus spp. are ubiquitous in nature, found in soil, air (both in natural environments and in buildings) as well as in natural water sources (O'Gorman and Fuller, 2008; Paulussen et al., 2017). Human contact with these fungi is therefore inevitable and poses a great risk to individuals predisposed to fungal infection because of defective immune systems. Additionally, antifungal resistance among Aspergillus spp. continues to complicate the treatment of aspergillosis. This resistance has also been observed in environmental isolates that have not yet been implicated in clinical situations (Hoda et al., 2019). Such intrinsic resistance towards antifungals adds further pathogenic potential to these aspergilli and poses a great threat to immunocompromised individuals who encounter these thermotolerant fungi in the environment.
MOLECULAR ASPECTS OF THERMOTOLERANCE
Temperature-related stress largely affects the physiology of an organism (Buckley and Huey, 2016). Alterations in a cell responding to elevated temperatures can be studied by investigating proteomic and genomic functionality under stressful conditions. In doing so, researchers identified stress response pathways that overlap among distinct fungal groups (Tereshina, 2005; Fuchs and Mylonakis, 2009; Leach et al., 2012; Juvvadi et al., 2014). The most common response pathway that is associated with thermotolerance is the heat shock response (HSR; Tereshina, 2005), which responds to sudden high increases in external temperature. It is a complex adaptation mechanism that focuses on downregulating housekeeping genes and upregulating cytoprotective genes in order to prevent or restore any damage due to the imposing heat shock (Verghese et al., 2012). The products and cofactors of these regulatory processes are proteins or chaperones known as heat shock proteins (HSPs). Their functions, in addition to gene regulation, include the denaturing, folding, refolding and transport of cytosolic proteins affected by the external stress (Tereshina, 2005). Additionally, HSPs assist in the transcriptional regulation of the cell wall integrity (CWI) and environmental stress response (ESR) pathways (Fuchs and Mylonakis, 2009; Verghese et al., 2012), both of which play a role in thermotolerance. The ESR involves many other signalling transductions responding to external changes in the microorganism's environment such as oxidative-, osmotic-, and pH stress (Verghese et al., 2012). The HSR, for instance, represents a subset within the ESR, since all of the genes involved in this pathway are included in the ESR regulon. The CWI pathway, on the other hand, harbours many other functions separate to environmental stress, despite having overlapping pathways with both the ESR and HSR (Dhar et al., 2013). Nevertheless, multiple regulatory proteins and genes in the CWI pathway have been identified and linked to thermotolerance. Within all these pathways and stress responses, a plethora of proteins and transcriptional factors all cooperate and cross-communicate to provide the micro organism with the proper machinery to adapt to thermal stress (Fuchs and Mylonakis, 2009; Verghese et al., 2013; Dhar et al., 2013).
Long-term thermotolerance and microevolution
Yeasts and other fungi adapt to environmental stress using several mechanisms which depend on the degree of stimulation as well as the duration thereof (Causton et al., 2001; Berry and Gasch, 2008; Dhar et al., 2013; Pereira et al., 2018). As for cells undergoing heat shock, the harsh stimulus is perceived, and various signals are sent to the rest of the cell to initiate repair as well as prevent further anticipated damage caused by the heat shock (Tereshina, 2005). During this process, housekeeping genes are paused while the cell rapidly responds to the external threat via HSR upregulation. However, during prolonged exposure to stressful conditions other signalling transductions occur simultaneously which aid in long-term adaptation, including regulatory factors of the CWI pathway and ESR (Causton et al., 2000; Berry and Gasch, 2008; Fuchs et al., 2009; Chen et al., 2012; Dunayevich et al., 2018; Sanz et al., 2018). Although most studies focused on industrially important yeasts, some researchers have investigated the molecular signalling constituents of thermotolerance in clinically relevant fungi (Argüelles, 1997; Juvvadi et al., 2003; Chang et al., 2004; Nichols et al., 2007; Chen et al., 2012; Chen et al., 2013; Chow et al., 2017; Yang et al., 2017; Brandão et al., 2018; Jung et al., 2018; So et al., 2018; Bloom et al., 2019). While the regulatory mechanisms of thermotolerance may vary among fungi, the underlying response pathways are similar (Alonso-Monge et al., 2009; Brown et al., 2020).
Generally, signalling pathways are induced by stimuli received through proteins located on the surface of a cell (Levin, 2005). These proteins or sensors can differ among species and rarely have singular roles. In fungi, precursors that form part of the CWI pathway mostly initiate signalling linked to thermotolerance (Verna et al., 1997; Zu et al., 2001; Verghese et al., 2012; Huang et al., 2018). The messages that these precursors receive are sent to other proteins that are part of various pathways, including the mitogen-activated protein kinase (MAPK)-, RAS-cAMP- and calcineurin signalling pathways (Fuchs et al., 2009; Parts et al., 2011; Verghese et al., 2012; Juvvadi et al., 2014). These pathways are also involved in many other cellular processes and are therefore not exclusively responsible for thermal stress adaptation (Causton et al., 2001; Berry and Gasch, 2008; Fuchs et al., 2009; Chen et al., 2012; Dunayevich et al., 2018; Sanz et al., 2018). Particular genes and proteins that form part of these pathways were linked to thermotolerance. However, the genome and transcriptome of yeasts were investigated only at severe heat shock conditions which might not provide an adequate representation of adaptation to gradual temperature increase (Fuchs et al., 2009; Parts et al., 2011; Verghese et al., 2012; Juvvadi et al., 2014). To gain a better perspective of fungi adapting to prolonged temperature increases, the genetic changes that occur over generations should be investigated.
Huang et al. (2018) carried out experimental evolution on a laboratory strain of S. cerevisiae by initiating adaptation through a stepwise increase in temperature, after which the genome was sequenced to investigate acquired thermotolerance. Considering that global temperatures increase gradually (IPCC, 2018), the approach of Huang and co-workers might provide an adequate representation of microevolution in fungi occurring in natural environments with temperature increases. The goal of their study was to experimentally evolve a yeast to obtain a high temperature growth phenotype (Htg+), as well as determining major contributing factors by investigating the mutations that occurred during the experimental evolution (Huang et al., 2018). The authors found mutations such as single nucleotide variants (SNVs), insertions and deletions (INDELs) and segmental duplications/deletions that could be associated with the Htg+ phenotype. Many of these mutations were nonsynonymous which altered the amino acid sequence of some proteins. Moreover, the mutations were multiple and sometimes parallel, increasing the significance of the mutation and thus the probability of association with the Htg+ phenotype. As a result of their study, genetic mutations were identified that can contribute to long-term fungal thermotolerance. In addition, a combination of non-essential gene mutations was examined and it was determined that a genetically modified thermotolerant strain can be attained (Huang et al., 2018). Although this might seem like a breakthrough for industrial purposes (Amore and Faraco, 2012), it becomes increasingly concerning when considering the implications that global warming might have on fungal microevolution and consequently the emergence of novel fungal pathogens.
The dirty river, the fungus and the heat
So far, studies aimed at investigating microevolutionary changes in clinically relevant fungi have mainly focused on virulence and physiological reactions within the host (Magditch et al., 2012; Wartenberg et al., 2014; Ene et al., 2018). However, the combined contribution of pollution and global warming on fungal evolution has not been studied. Apart from the regulatory pathways or physiological reactions to environmental changes, little is known regarding fungal adaptation on a genetic level (Causton et al., 2001; Dhar et al., 2013). Moreover, studies often fail to consider the microevolutionary effects that result from interactions with other biotic and abiotic factors, e.g., other organisms, including plant life, and environmental conditions. The latter, for example, might include anthropogenic pollutants and climate change. Studies investigating pollution-associated microevolution are lacking despite the valuable insight that can be gained from understanding the effects of pollution on fungal growth. This is especially concerning since the pollutants may include antifungals released into the rivers (Chitescu et al., 2015). Thus, during the process of acquiring thermotolerance with the aid of global warming, fungi in polluted aquatic environments could potentially gain resistance towards antifungals, due to the presence of these compounds in surface waters. Future emerging pathogens could therefore be resistant to treatment before even causing infections. Pollution, such as organic pollution, also promotes the growth of fungi by serving as a rich source of nutrients (Wen et al., 2017), which might allow some species to outcompete more fastidious microorganisms and consequently disturb the natural mycobiome (Ortiz-Vera et al., 2018). Certain pollutants may also initiate virulence phenotypes in fungi, e.g., as seen for persistent organic pollutants (POPs) such as pentachlorophenol (Martins et al., 2018). Increasing anthropogenic activities resulting in polluted river water could therefore potentially cause a rise in fungal numbers, particularly pathogenic species. Considering the wide range of factors that may impact on fungi in natural environments, it is prudent to employ not only genomics but also more traditional techniques, such as biochemical, ecological, genetic, morphological, and ontological approaches (Naranjo-Ortiz and Gabaldon, 2019), to delineate the complex synergetic effect of climate change and increased pollution levels on fungal biology.
CONCLUSIONS
With the ever-increasing urbanisation to accommodate the equally growing population, increased environmental temperatures and pollution of rivers will have detrimental outcomes on human health (Fig. 1). South Africa is a developing country experiencing rapid urbanisation in informal communities with municipalities struggling to keep up and failing to implement proper sanitation infrastructure, causing communities to be dependent on natural water sources for their livelihoods (Colvin et al., 2016). Although little research has been conducted for South Africa, there is growing evidence that a wide diversity of fungi, including opportunistic species that can tolerate the mammalian body temperature, occur in South African rivers. The extent of the risk posed by waterborne fungi to community health therefore needs to be determined, especially in view of the fact that opportunistic yeasts, such as Candida spp., that infect individuals suffering from HIV/AIDS appear to be common in polluted rivers. Furthermore, rising river water temperatures will most probably induce increased thermotolerance amongst waterborne fungi. Worryingly, the fungal metabolism can adapt to these increases and current evidence indicates that more fungi will become thermotolerant as global temperatures rise. This, together with the fact that sewage-polluted rivers contain elevated concentrations of fungi, points to an imminent increase in the incidence of mycoses in the not-so-distant future. Important to note is that non-clinical isolates implicated in mycoses might represent antifungal-resistant fungi due to the various chemicals derived from agricultural-, industrial- and anthropogenic waste that end up in surface waters. However, few studies have been conducted to elucidate the resistance acquired from antifungal exposure in water sources. Nonetheless, the combinatorial effects of pollution (organic or inorganic) and global warming could be drivers in fungal microevolution and potentially lead to a rise in emergent pathogenic fungi, for which there is a limited range of anti-fungal drugs available. Efforts to better record the occurrence and delineate the ecology and antifungal resistance of clinically relevant fungi in South African river systems should therefore be a priority.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge funding from The Water Research Commission, Pretoria, South Africa (WRC Project No. 2019/2020-00137). The National Research Foundation (NRF) is thanked for the personal funding of Heidi Steffen. Further thanks are given to fellow researchers in the Botha Laboratory (Department of Microbiology, Stellenbosch University) for general advice and insights given during the write-up of this review article.
AUTHOR CONTRIBUTIONS
Heidi Steffen contributed to write-up and conceptualisation. Alfred Botha, Gideon Wolfaardt and Caylin Bosch contributed to conceptualisation and revision.
ORCIDS
Alfred Botha: https://orcid.org/0000-0003-0988-647X
REFERENCES
ALBRECHT D, GUTHKE R, BRAKHAGE AA and KNIEMEYER O (2010) Integrative analysis of the heat shock response in Aspergillus fumigatus. BMC Genomics. 11 (1). https://doi.org/10.1186/1471-2164-11-32 [ Links ]
ALFORD RA, BRADFIELD KS and RICHARDS SJ (2007) Ecology: Global warming and amphibian losses. Nature. 447 (7144) 3-6. https://doi.org/10.1038/nature05940 [ Links ]
ALMEIDA F, RODRIGUES ML and COELHO C (2019) The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 10 (FEB) 1-5. https://doi.org/10.3389/fmicb.2019.00214 [ Links ]
ALMEIDA JND, FRANCISCO EC, HAGEN F, BRANDÃO IB, PEREIRA FM, PRESTA DIAS PH, DE MIRANDA COSTA MM, DE SOUZA JORDÃO RT, DE GROOT T and COLOMBO AL (2021) Emergence of Candida auris in Brazil in a COVID-19 intensive care unit. J. Fungi. 7 (3) 220. https://doi.org/10.3390/jof7030220 [ Links ]
ALONSO-MONGE R, ROMÁN E, ARANA DM, PLA J and NOMBELA C (2009) Fungi sensing environmental stress. Clin. Microbiol. Infect. 15 (SUPPL. 1) 17-19. https://doi.Org/10.1111/j.1469-0691.2008.02690.x [ Links ]
AMORE A and FARACO V (2012) Potential of fungi as category I Consolidated BioProcessing organisms for cellulosic ethanol production. Renew. Sustain. Energ. Rev. 16 (5) 3286-3301. https://doi.org/10.1016/j.rser.2012.02.050 [ Links ]
ARASTEHFAR A, CARVALHO A, VAN DE VEERDONK FL, JENKS JD, KOEHLER P, KRAUSE R, CORNELY OA, PERLIN D, LASS-FLORL C and HOENIGL M (2020) COVID-19 associated pulmonary aspergillosis (CAPA)-from immunology to treatment. J. Fungi. 6 (2) 91. https://doi.org/10.3390/jof6020091 [ Links ]
ARAÚJO GRDS, DE SOUZA W and FRASES S (2017) The hidden pathogenic potential of environmental fungi. Future Microbiol. 12 (16) 1533-1540. https://doi.org/10.2217/fmb-2017-0124 [ Links ]
ARGUELLES JC (1997) Thermotolerance and trehalose accumulation induced by heat shock in yeast cells of Candida albicans. FEMS Microbiol. Lett. 146 (1) 65-71. https://doi.org/10.1016/S0378-1097(96)00455-7 [ Links ]
ARORA P, SINGH P, WANG Y, YADAV A, PAWAR K, SINGH A, PADMAVATI G, XU J and CHOWDHARY A. (2021) Environmental isolation of Candida auris from the coastal wetlands of Andaman Islands, India. Mbio 12e03181-20. https://doi.org/10.1128/mBio.03181-20 [ Links ]
ARROYO MG, FERREIRA AM, FROTA OP, BRIZZOTTI-MAZUCHI NS, PERESI JTM, RIGOTTI MA, MACEDO CE, SOUSA AFLD, ANDRADE DD and ALMEIDA MTGD (2020) Broad diversity of fungi in hospital water. Sci. World J. 2020. https://doi.org/10.1155/2020/9358542 [ Links ]
ARVANITIDOU M, KANELLOU K and VAGIONA DG (2005) Diversity of Salmonella spp. and fungi in northern Greek rivers and their correlation to fecal pollution indicators. Environ. Res. 99 (2) 278-284. https://doi.org/10.1016/j.envres.2005.01.002 [ Links ]
ARVANITIDOU M, KANELLOU K, KATSOUYANNOPOULOS V and TSAKRIS A (2002) Occurrence and densities of fungi from northern Greek coastal bathing waters and their relation with faecal pollution indicators. Water Res. 36 (20) 5127-5131. https://doi.org/10.1016/S0043-1354(02)00235-X [ Links ]
ASSRESS HA, SELVARAJAN R, NYONI H, NTUSHELO K, MAMBA BB and MSAGATI TAM (2019) Diversity, co-occurrence and implications of fungal communities in wastewater treatment plants. Sci. Rep. 9 (1) 1-15. https://doi.org/10.1038/s41598-019-50624-z [ Links ]
BABIC MN, GUNDE-CIMERMAN N, VARGHA M, TISCHNER Z, MAGYAR D, VERÍSSIMO C, SABINO R, VIEGAS C, MEYER W and BRANDÃO J (2017) Fungal contaminants in drinking water regulation? A tale of ecology, exposure, purification and clinical relevance. Int. J. Environ. Res. Public Health. 14 (6). https://doi.org/10.3390/ijerph14060636 [ Links ]
BADIEE P and ZARE M (2017) Consideration of invasive fungal infections in immunocompetent hosts Archiv. Clin. Infect. Dis. 12 (4) 66111. https://doi.org/10.5812/archcid.66111. [ Links ]
BAQUERO F (2009) Environmental stress and evolvability in microbial systems. Clin. Microbiol. Infect. 15 (SUPPL. 1) 5-10. https://doi.org/10.1111/j.1469-0691.2008.02677.x [ Links ]
BARON E, ANAISSIE E, DUMPHY F, MCCREDIE K and FAINSTEIN V (1988) Hansenula anomala fungemia. Clin. Infect. Dis. 10 (6) 1182-1186. https://doi.org/10.1093/clinids/10.6.1182 [ Links ]
BELFORD I (2013) Investigating the removal of Candida and other potential pathogens from wastewater via an experimental rhizofiltration system. Doctoral dissertation, Stellenbosch University. https://scholar.sun.ac.za/handle/10019.1/79806?show=full [ Links ]
BENEDICT K, RICHARDSON M, VALLABHANENI S, JACKSON BR and CHILLER T (2017) Emerging issues, challenges, and changing epidemiology of fungal disease outbreaks. Lancet Infect. Dis. 17 (12) e403-e411. https://doi.org/10.1016/S1473-3099(17)30443-7 [ Links ]
BERECZKI L, BARTHA N, KOCSUBÉ S, SÓKI J, LENGYEL G, TÁLOSI G, MÁDER K, DEÁK J and DÓCZI I (2012) Fungaemia caused by Candida pulcherrima. Med. Mycol. 50 (5) 522-524. https://doi.org/10.3109/13693786.2011.644590 [ Links ]
BERGMAN AG and KAUFFMAN CA (1984) Dermatitis due to Sporobolomyces infection. Archiv. Dermatol. 120 (8) 1059-1060. https://doi.org/10.1001/archderm.1984.01650440089026 [ Links ]
BERRY DB and GASCH AP (2008) Stress-activated genomic expression changes serve a preparative role for impending stress in yeast. Mol. Biol. Cell. 19 (11) 4580-4587. https://doi.org/10.1091/mbc.e07-07-0680 [ Links ]
BEYDA ND, CHUANG SH, JAHANGIR ALAM M, SHAH DN, NG TM, MCCASKEY L and GAREY KW (2013) Treatment of Candida famata bloodstream infections: Case series and review of the literature. J. Antimicrob. Chemotherapy. 68 (2) 438-443. https://doi.org/10.1093/jac/dks388 [ Links ]
BHABHRA R and ASKEW DS (2005) Thermotolerance and virulence of Aspergillus fumigatus: Role of the fungal nucleolus. Med. Mycol. 43 (SUPPL.1) 87-93. https://doi.org/10.1080/13693780400029486 [ Links ]
BICANIC TA and HARRISON TS (2014) Systemic fungal infections. Medicine. 42 (1) 26-30. https://doi.org/10.1016/j.mpmed.2013.10.006 [ Links ]
BLANCO JL and GARCIA ME (2008) Immune response to fungal infections. Vet. Immunol. Immunopathol. 125 (1-2) 47-70. https://doi.org/10.1016/j.vetimm.2008.04.020 [ Links ]
BLOOM ALM, JIN RM, LEIPHEIMER J, BARD JE, YERGEAU D, WOHLFERT EA and PANEPINTO JC (2019) Thermotolerance in the pathogen Cryptococcus neoformans is linked to antigen masking via mRNA decay-dependent reprogramming. Nat. Commun. 10 (1) 1-13. https://doi.org/10.1038/s41467-019-12907-x [ Links ]
BOLIGNANO G and CRISEO G (2003) Disseminated nosocomial fungal infection by Aureobasidium pullulans var. melanigenum: A case report. J. Clin. Microbiol. 41 (9) 4483-4485. https://doi.org/10.1128/JCM.41.9.4483-4485.2003 [ Links ]
BONGOMIN F, GAGO S, OLADELE R and DENNING D (2017) Global and multi-national prevalence of fungal diseases-estimate precision. J. Fungi 3 (4) 57. https://doi.org/10.3390/jof3040057 [ Links ]
BOSCH C, BHANA Z, TOPLIS B, VOLSCHENK H and BOTHA A (2021) Transcriptomic response of Cryptococcus neoformans to ecologically relevant nitrogen concentrations. FEMS Yeast Res. https://doi.org/10.1093/femsyr/foab028 [ Links ]
BOSCH C, TOPLIS B, VREULINK JM, VOLSCHENK H and BOTHA A (2020) Nitrogen concentration affects amphotericin B and fluconazole tolerance of pathogenic cryptococci. FEMS Yeast Res. 20 (2) p.foaa010. https://doi.org/10.1093/femsyr/foaa010 [ Links ]
BOYD AS, WHELESS L, BRADY BG and ELLIS D (2017) Cutaneous Yarrowia lipolytica infection in an immunocompetent woman. JAAD Case Rep. 3 (3) 219-221. https://doi.org/10.1016/j.jdcr.2017.02.010 [ Links ]
BRANDÃO F, ESHER SK, OST KS, PIANALTO K, NICHOLS CB, FERNANDES L, BOCCA AL, POÇAS-FONSECA MJ and ALSPAUGH JA (2018) HDAC genes play distinct and redundant roles in Cryptococcus neoformans virulence. Sci. Rep. 8 (1) 1-17. https://doi.org/10.1038/s41598-018-21965-y [ Links ]
BRANDÃO LR, MEDEIROS AO, DUARTE MC, BARBOSA AC and ROSA CA (2010) Diversity and antifungal susceptibility of yeasts isolated by multiple-tube fermentation from three freshwater lakes in Brazil. J. Water Health. 8 (2) 279-289. https://doi.org/10.2166/wh.2009.170 [ Links ]
BRANDÃO LR, MEDEIROS AO, DUARTE MC, BARBOSA AC and ROSA CA (2010) Diversity and antifungal susceptibility of yeasts isolated by multiple-tube fermentation from three freshwater lakes in Brazil. J. Water Health. 8 (2) 279-289. https://doi.org/10.2166/wh.2009.170 [ Links ]
BRITO-SANTOS F, FIGUEIREDO-CARVALHO MHG, COELHO RA, SALES A and ALMEIDA-PAES R (2017) Tinea capitis by Microsporum audouinii: case reports and review of published global literature 2000-2016. Mycopathologia. 182 (11) 1053-1060. https://doi.org/10.1007/s11046-017-0181-1 [ Links ]
BROSS JE, MANNING P, KACIAN D and TALBOT GH (1986) Pseudomeningitis caused by Sporobolomyces salmonicolor. Am. J. Infect. Control 14 (5) 220-223. https://doi.org/10.1016/0196-6553(86)90121-5 [ Links ]
BROWN AJP, LARCOMBE DE and PRADHAN A (2020) Thoughts on the evolution of Core Environmental Responses in yeasts. Fungal Biol. 124 (5) 475 -481. https://doi.org/10.1016/j.funbio.2020.01.003 [ Links ]
BROWN GD, DENNING DW, GOW NA, LEVITZ SM, NETEA MG and WHITE TC (2012) Hidden killers: human fungal infections. Sci. Translational Med. 4 (165) 165-165. https://doi.org/10.1126/scitranslmed.3004404 [ Links ]
BUCK JD and BUBUCIS PM (1978) Membrane filter procedure for enumeration of Candida albicans in natural waters. Appl. Environ. Microbiol. 35 (2) 237-242. https://doi.org/10.1128/aem.35.2.237-242.1978 [ Links ]
BUCKLEY LB and HUEY RB (2016) How extreme temperatures impact organisms and the evolution of their thermal tolerance. Integr. Compar. Biol. 56 (1) 98-109. https://doi.org/10.1093/icb/icw004 [ Links ]
BUZZINI P, LACHANCE MA and YURKOV A (2017) Yeasts in natural ecosystems: Diversity. Yeasts Nat. Ecosyst. Div. (March 2018) 1-499. https://doi.org/10.1007/978-3-319-62683-3 [ Links ]
CAGGIANO G, DIELLA G, TRIGGIANO F, BARTOLOMEO N, APOLLONIO F, CAMPANALE C, LOPUZZO M and MONTAGNA MT (2020) Occurrence of fungi in the potable water of hospitals: A public health threat. Pathogens. 9 (10) 783. https://dx.doi.org/10.3390%2Fpathogens9100783 [ Links ]
CARREGA G, RICCIO G, SANTORIELLO L, PASQUALINI M and PELLICCI R (1997) Candida famata fungemia in a surgical patient successfully treated with fluconazole. Eur. J. Clin. Microbiol. Infect. Dis. 16 (9) 698. https://doi.org/10.1007/bf01708564 [ Links ]
CASADEVALL A (2020) Climate change brings the specter of new infectious diseases. J. Clin. Invest. 130 (2) 553-555. https://doi.org/10.1172/JCI135003 [ Links ]
CASADEVALL A, KONTOYIANNIS DP and ROBERT V (2019) On the emergence of Candida auris: climate change, azoles, swamps, and birds. MBio. 10 (4) e01397-19. https://doi.org/10.1128/mBio.01397-19 [ Links ]
CAUSTON HC, REN B, SANG SEOK KOH, HARBISON CT, KANIN E, JENNINGS EG, TONG IHN LEE, TRUE HL, LANDER ES and YOUNG RA (2001) Remodeling of yeast genome expression in response to environmental changes. Mol. Biol. Cell. 12 (2) 323-337. https://doi.org/10.1091/mbc.12.2.323 [ Links ]
CHAKRABARTI A, SINGH K, NARANG A, SINGHI S, BATRA, R., RAO, K.L.N., RAY, P., GOPALAN, S., DAS, S., GUPTA, V. and GUPTA, A.K., 2001. Outbreak of Pichia anomala infection in the pediatric service of a tertiary-care center in Northern India. J. Clin. Microbiol. 39 (5) 1702-1706. https://doi.org/10.1128/jcm.39.5.1702-1706.2001 [ Links ]
CHANG YC, TSAI HF, KAROS M and KWON-CHUNG KJ (2004) THTA, a thermotolerance gene of Aspergillus fumigatus. Fungal Genet. Biol. 41 (9) 888-896. https://doi.org/10.1016Zj.fgb.2004.06.004 [ Links ]
CHATTERJEE S and TATU U (2017) Heat shock protein 90 localizes to the surface and augments virulence factors of Cryptococcus neoformans. PLoS Neglected Trop. Dis. 11 (8) 1-20. https://doi.org/10.1371/journal.pntd.0005836 [ Links ]
CHEN D, HU M, GUO Y and DAHLGREN RA (2016) Changes in river water temperature between 1980 and 2012 in Yongan watershed, eastern China: Magnitude, drivers and models. J. Hydrol. 533 191199. https://doi.org/10.1016/j.jhydrol.2015.12.005 [ Links ]
CHEN IC, HILL JK, OHLEMÜLLER R, ROY DB and THOMAS CD (2011) Rapid range shifts of species associated with high levels of climate warming. Science. 333 (6045) 1024-1026. https://doi.org/10.1126/science.1206432 [ Links ]
CHEN Y, KONIECZKA JH, SPRINGER DJ, BOWEN SE, ZHANG J, SILAO FGS, BUNGAY AAC, BIGOL UG and NICOLAS MG (2012) Convergent evolution of calcineurin pathway roles in thermotolerance and virulence in Candida glabrata. G3 Genes Genomes Genet. 2 (June) 675-691. https://doi.org/10.1534/g3.112.002279 [ Links ]
CHEN YL, LEHMAN VN, LEWIT Y, AVERETTE AF and HEITMAN J (2013) Calcineurin governs thermotolerance and virulence of Cryptococcusgattii. G3: Genes Genomes Genet. 3 (3) 527-539. https://doi.org/10.1534/g3.112.004242 [ Links ]
CHITESCU CL, KAKLAMANOS G, NICOLAU AI and STOLKER AAML (2015) High sensitive multiresidue analysis of pharmaceuticals and antifungals in surface water using U-HPLC-Q-Exactive Orbitrap HRMS. Application to the Danube river basin on the Romanian territory. Sci. Total Environ. 532 501-511. https://doi.org/10.1016/j.scitotenv.2015.06.010 [ Links ]
CHOW EWL, CLANCEY SA, BILLMYRE RB, AVERETTE AF, GRANEK JA, MIECZKOWSKI P, CARDENAS ME and HEITMAN J (2017) Elucidation of the calcineurin-Crz1 stress response transcriptional network in the human fungal pathogen Cryptococcus neoformans. PLoS Genet. 13 (4) 1-29. https://doi.org/10.1371/journal.pgen.1006667 [ Links ]
CHRISTIAENS G, HAYETTE MP, JACQUEMIN D, MELIN P, MUTSERS J and DE MOL P (2005) An outbreak of Absidia corymbifera infection associated with bandage contamination in a burns unit. J. Hosp. Infect. 61 (1) 88. https://doi.org/10.1016/j.jhin.2004.12.011 [ Links ]
COELHO MA, ALMEIDA JMF, MARTINS IM, DA SILVA AJ and SAMPAIO JP (2010) The dynamics of the yeast community of the Tagus river estuary: testing the hypothesis of the multiple origins of estuarine yeasts. Anton. Leeuw. 98 (3) 331-342. https://doi.org/10.1007/s10482-010-9445-1 [ Links ]
COHEN JM, CIVITELLO DJ, VENESKY MD, MCMAHON TA and ROHR JR (2017) Thermal mismatches explain how climate change and infectious disease drove widespread amphibian extinctions. bioRxiv 164814. https://doi.org/10.1101/164814 [ Links ]
COLES M, COX K and CHAO A (2020) Candida haemulonii: An emerging opportunistic pathogen in the United States? IDCases. 21 e00900. https://doi.org/10.1016/j.idcr.2020.e00900 [ Links ]
COLOMBO AL, PADOVAN ACB and CHAVES GM (2011) Current knowledge of Trichosporon spp. and trichosporonosis. Clin. Microbiol. Rev. 24 (4) 682-700. https://doi.org/10.1128/CMR.00003-11 [ Links ]
COLVIN C, MURUVEN D, LINDLEY D, GORDEN H and SCHACHTSCHENEIDER K (2016) Water : Facts and futures rethinking South Africa's water future. WWF-SA, Cape Town. 1-100. www.wwf.org.za [ Links ]
COOKE WB, PHAFF HJ, MILLER MW, SHIFRINE M and KNAPP EP (1960) Yeasts in polluted water and sewage. Mycologia. 52 (2) 210230. https://doi.org/10.1080/00275514.1960.12024896 [ Links ]
CORTEZ KJ, ROILIDES E, QUIROZ-TELLES F, MELETIADIS J, ANTACHOPOULOS C, KNUDSEN T, BUCHANAN W, MILANOVICH J, SUTTON DA, FOTHERGILL A, and co-authors (2008) Infections caused by Scedosporium spp. Clin. Microbiol. Rev. 21 (1) 157-197. https://doi.org/10.1128/CMR.00039-07 [ Links ]
CULLIS JDS, HORN A, ROSSOUW N, FISHER-JEFFES L, KUNNEKE MM and HOFFMAN W (2019) Urbanisation, climate change and its impact on water quality and economic risks in a water scarce and rapidly urbanising catchment: Case study of the Berg River Catchment. H2Open J. 2 (1) 146-167. https://doi.org/10.2166/H2OJ.2019.027 [ Links ]
D'AMEN M and BOMBI P (2009) Global warming and biodiversity: Evidence of climate-linked amphibian declines in Italy. Biol. Conserv. 142 (12) 3060-3067. https://doi.org/10.1016Zj.biocon.2009.08.004 [ Links ]
DATTA N, ARENDRUP MC and SAUNTE JP (2015) First report of Candida palmioleophila endogenous endophthalmitis. Acta Ophthalmol. 93 (6) e517-e518. https://doi.org/10.1111/aos.12662 [ Links ]
DE ALMEIDA JMGCF (2005) Yeast community survey in the Tagus estuary. FEMS Microbiol. Ecol. 53 (2) 295-303. https://doi.org/10.1016/j.femsec.2005.01.006 [ Links ]
DHAR R, SÄGESSER R, WEIKERT C and WAGNER A (2013) Yeast adapts to a changing stressful environment by evolving cross-protection and anticipatory gene regulation. Mol. Biol. Evol. 30 (3) 573-588. https://doi.org/10.1093/molbev/mss253 [ Links ]
DOGGETT, MS (2000) Characterization of fungal biofilms within a municipal water distribution system. Appl. Environ. Microbiol. 66 (3) 1249-1251. https://doi.org/10.1128%2Faem.66.3.1249-1251.2000 [ Links ]
DUNAYEVICH P, BALTANÄS R, CLEMENTE JA, COUTO A, SAPOCHNIK D, VASEN G and COLMAN-LERNER A (2018) Heat-stress triggers MAPK crosstalk to turn on the hyperosmotic response pathway. Sci. Rep. 8 (1) 1-15. https://doi.org/10.1038/s41598-018-33203-6 [ Links ]
DUPONT B, BROWN HHC, WESTERMANN K, MARTINS MD, REX JH, LORTHOLARY O and KAUFFMANN CA (2000) Mycoses in AIDS. Med. Mycol. 38 (i) 259-267. https://doi.org/10.1080/mmy.38.s1.259.267 [ Links ]
DYNOWSKA M (1997) Yeast-like fungi possessing bio-indicator properties isolated from the Lyna river. Acta Mycol. 32 (2) 279-286. https://doi.org/10.5586/am.1997.024 [ Links ]
EDOKPAYI JN, ROGAWSKI ET, KAHLER DM, HILL CL, REYNOLDS C, NYATHI E, SMITH JA, ODIYO JO, SAMIE A, BESSONG P and DILLINGHAM R (2018) Challenges to sustainable safe drinking water: a case study of water quality and use across seasons in rural communities in Limpopo province, South Africa. Water. 10 (2) 159. https://doi.org/10.3390/w10020159 [ Links ]
EISENBERG ES, ALPERT BE, WEISS RA, MITTMAN N and SOEIRO R (1983) Rhodotorula rubra peritonitis in patients undergoing continuous ambulatory peritoneal dialysis. Am. J. Med. 75 (2) 349352. https://doi.org/10.1016/0002-9343(83)91216-0 [ Links ]
EL HELOU G and PALAVECINO E (2017) Candida pararugosa: First Reported Bloodstream Infection in an Adult. Cureus. 9 (5). https://doi.org/10.7759/cureus.1283 [ Links ]
ELANDER RP (2003) Industrial production of β-lactam antibiotics. Appl. Microbiol. Biotechnol. 61 (5) 385-392. https://doi.org/10.1007/s00253-003-1274-y [ Links ]
ELLIS DH and PFEIFFER TJ (1990) Natural habitat of Cryptococcus neoformans var. gattii. J. Clin. Microbiol. 28 (7) 1642-1644. https://doi.org/10.1128/jcm.28.7.1642-1644.1990 [ Links ]
ELLIS DH and PFEIFFER TJ (1992) The ecology of Cryptococcus neoformans. Eur. J. Epidemiol. 8 (3) 321-325. https://doi.org/10.1007/bf00158562 [ Links ]
ENE IV, FARRER RA, HIRAKAWA MP, AGWAMBA K, CUOMO CA and BENNETT RJ (2018) Global analysis of mutations driving microevolution of a heterozygous diploid fungal pathogen. Proc. Natl Acad. Sci. 115 (37) E8688-E8697. https://doi.org/10.1073/pnas.1806002115 [ Links ]
FAUSTO A, RODRIGUES ML and COELHO C (2019) The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 10 (FEB) 1-5. https://doi.org/10.3389/fmicb.2019.00214 [ Links ]
FISHER MC, GARNER TW and WALKER SF (2009) Global emergence of Batrachochytrium dendrobatidis and amphibian chytridiomycosis in space, time, and host. Annu. Rev. Microbiol. 63 291-310. https://doi.org/10.1146/annurev.micro.091208.073435 [ Links ]
FUCHS BB and MYLONAKIS E (2009) Our paths might cross: The role of the fungal cell wall integrity pathway in stress response and cross talk with other stress response pathways. Eukaryotic Cell. 8 (11) 1616-1625. https://doi.org/10.1128/EC.00193-09 [ Links ]
GADANHO M, LIBKIND D and SAMPAIO JP (2006) Yeast diversity in the extreme acidic environments of the Iberian pyrite belt. Microb. Ecol. 52 (3) 552-563. https://doi.org/10.1007/s00248-006-9027-y [ Links ]
GAFFI (2017) Fungal disease frequency. https://www.gaffi.org/why/fungal-disease-frequency/ (Accessed 28 January 2022). [ Links ]
GARBER G (2001) An overview of fungal infections. Drugs. 61 (SUPPL. 1) 1-12. https://doi.org/10.2165/00003495-200161001-00001 [ Links ]
GARCIA-SOLACHE MA and CASADEVALL A (2010) Global warming will bring new fungal diseases for mammals. mBio. 1 (1) 1-3. https://doi.org/10.1128/mBio.00061-10 [ Links ]
GARG D, MUTHU V, SEHGAL IS, RAMACHANDRAN R, KAUR H, BHALLA A, PURI GD, CHAKRABARTI A and AGARWAL R (2021) Coronavirus disease (Covid-19) associated mucormycosis (CAM): case report and systematic review of literature. Mycopathologia. 186 (2) 289-298. https://doi.org/10.3410/f.739505445.793585416 [ Links ]
GHAJARI A, LOTFALI E, NOROUZI M and ARAB-MAZAR Z (2018) First report of Vulvovaginitis due to Cryptococcus magnus in Iran. Curr. Med. Mycol. 4 (1) 30. https://dx.doi.org/10.18502%2Fcmm.4.1.32 [ Links ]
GLINSKA-LEWCZUK K, GOLAS I, KOC J, GOTKOWSKA-PLACHTA A, HARNISZ M and ROCHWERGER A (2016) The impact of urban areas on the water quality gradient along a lowland river. Environ. Monit. Assess. 188 (11) 1-15. https://doi.org/10.1007/s10661-016-5638-z [ Links ]
GOLD JW, POSTON W, MERTELSMANN R, LANGE M, KIEHN T, EDWARDS F, BERNARD E, CHRISTIANSEN K and ARMSTRONG D (1981) Systemic infection with Trichosporon cutaneum in a patient with acute leukemia: report of a case. Cancer. 48 (10) 2163-2167. https://doi.org/10.1002/1097-0142(19811115)48:10<2163::AID-CNCR2820481008>3.0.CO;2-T [ Links ]
GOLDANI LZ, CRAVEN DE and SUGAR AM (1995) Central venous catheter infection with Rhodotorula minuta in a patient with AIDS taking suppressive doses of fluconazole. Journal of Medical and Veterinary Mycology. 33 (4) 267-270. https://doi.org/10.1080/02681219580000531 [ Links ]
GOSTINCAR C, GRUBE M and GUNDE-CIMERMAN N (2011) Evolution of Fungal Pathogens in Domestic Environments? Fungal Biol. 115 (10) 1008-1018. https://doi.org/10.1016/j.funbio.2011.03.004 [ Links ]
GRLER NC, TOSUN I and AYDIN F (2017) The identification of Meyerozyma guilliermondii from blood cultures and surveillance samples in a university hospital in Northeast Turkey: A ten-year survey. J. Mycol. MUdicale. 27 (4) 506-513. https://doi.org/10.1016/j.mycmed.2017.07.007 [ Links ]
GROSSART HP, VAN DEN WYNGAERT S, KAGAMI M, WURZBACHER C, CUNLIFFE M and ROJAS-JIMENEZ K (2019) Fungi in aquatic ecosystems. Nat. Rev. Microbiol. 17 (6) 339-354. https://doi.org/10.1038/s41579-019-0175-8 [ Links ]
GUARNER J (2017) Human immunodeficiency virus and fungal infections. Seminars Diagn. Pathol. 34 (4) 325-331. https://doi.org/10.1053/j.semdp.2017.04.007 [ Links ]
GUINEA J (2014) Global trends in the distribution of Candida species causing candidemia. Clinical Microbiol. Infection. 8 5-10. https://doi.org/10.1111/1469-0691.12539 [ Links ]
GUITARD J, ANGOULVANT A, LETSCHER-BRU V, L'OLLIVIER C, CORNET M, DALLE F, GRENOUILLET F, LACROIX C, VEKHOFF A, MAURY E, and co-authors (2013) Invasive infections due to Candida norvegensis and Candida inconspicua: Report of 12 cases and review of the literature. Medical Mycology. 51 (8) 795-799. https://doi.org/10.3109/13693786.2013.807444 [ Links ]
GYAURGIEVA OH, BOGOMOLOVA TS and GORSHKOVA GI (1996) Meningitis caused by Rhodotorula rubra in an HIV-infected patient. J. Med. Vet. Mycol. 34 (5) 357-359. https://doi.org/10.1080/02681219680000611 [ Links ]
HA MV, CHOY MS, MCCOY D, FERNANDEZ N and SUH JS (2018) Candida catenulata candidemia and possible endocarditis in a cirrhotic patient successfully de-escalated to oral fluconazole. J. Clin. Pharm. Therapeutics. 43 (6) 910-913. https://doi.org/10.1111/jcpt.12728 [ Links ]
HAACK SK, DURIS JW, FOGARTY LR, KOLPIN DW, FOCAZIO MJ, FURLONG ET and MEYER MT (2009) Comparing wastewater chemicals, indicator bacteria concentrations, and bacterial pathogen genes as faecal pollution indicators. J. Environ. Qual. 38 (1) 248-258. https://doi.org/10.2134/jeq2008.0173 [ Links ]
HAGEN F, KHAYHAN K, THEELEN B, KOLECKA A, POLACHECK I, SIONOV E, FALK R, PARNMEN S, LUMBSCH HT and BOEKHOUT T (2015) Recognition of seven species in the Cryptococcus gattii/ Cryptococcus neoformans species complex. Fungal Genet. Biol. 78 16-48. https://doi.org/10.1016/j.fgb.2015.02.009 [ Links ]
HAGESKAL G, LIMA N and SKAAR I (2009) The study of fungi in drinking water. Mycol. Res. 113 (2) 165-172. https://doi.org/10.1016/j.mycres.2008.10.002 [ Links ]
HANSEN J, SATO M, HEARTY P, RUEDY R, KELLEY M, MASSON-DELMOTTE V, RUSSELL G, TSELIOUDIS G, CAO J, RIGNOT E and co-authors (2016) Ice melt, sea level rise and superstorms: evidence from paleoclimate data, climate modelling, and modern observations that 2°C global warming could be dangerous. Atmos. Chem. Phys. 16 3761-3812. https://doi.org/10.5194/acp-16-3761-2016,2016. [ Links ]
HARRISON TS and BROUWER AE (2009) Systemic fungal infections. Medicine. 37 (12) 660-664. https://doi.org/10.1016/j.mpmed.2009.09.016 [ Links ]
HAVELAAR AH, FURUSE K and HOGEBOOM WM (1986). Bacteriophages and indicator bacteria in human and animal faeces. J. Appl. Bacteriol. 60 (3) 255-262. https://doi.org/10.1111/j.1365-2672.1986.tb01081.x [ Links ]
HERBIG FJW (2019) Talking dirty - effluent and sewage irreverence in South Africa: A conservation crime perspective. Cogent Soc. Sci. 5 (1). https://doi.org/10.1080/23311886.2019.1701359 [ Links ]
HOANG JK and BURRUSS J (2007) Localized cutaneous Cryptococcus albidus infection in a 14-year-old boy on etanercept therapy. Pediatric Dermatol. 24 (3) 285-288. https://doi.org/10.1111/j.1525-1470.2007.00404.x [ Links ]
HODA S, AGARWAL H, AHLUWALIA SK and VIJAYARAGHAVAN MVP (2019) Antifungal Resistance Analysis of Environmental Isolates of Aspergillus in North India. J. Pure Appl. Microbiol. 13 (1) 385-392. https://doi.org/10.22207/jpam.13.L42 [ Links ]
HSUEH PR, TENG LJ, HO SW and LUH KT (2003) Catheter-related sepsis due to Rhodotorula glutinis. J. Clin. Microbiol. 41 (2) 857-859. https://doi.org/10.1128/JCM.41.2.857-859.2003 [ Links ]
HU Y, ZHANG J, LI X, YANG Y, ZHANG Y, MA J and XI L (2013) Penicillium marneffei infection: an emerging disease in mainland China. Mycopathologia. 175 (1) 57-67. https://doi.org/10.1007/s11046-012-9577-0 [ Links ]
HUANG CJ, LU MY, CHANG YW and LI WH (2018) Experimental evolution of yeast for high-temperature tolerance. Mol. Biol. Evol. 35 (8) 1823-1839. https://doi.org/10.1093/molbev/msy077 [ Links ]
HUANG DB, OSTROSKY-ZEICHNER L, WU JJ, PANG KR and TYRING SK (2004) Therapy of common superficial fungal infections. Dermatol. Therapy. 17 517-522. https://doi.org/10.1111/j.1396-0296.2004.04055.x [ Links ]
HUNTER-ELLUL L, SCHEPP ED, LEA A and WILKERSON MG (2014) A rare case of Cryptococcus luteolus-related tenosynovitis. Infection. 42 (4) 771-774. https://doi.org/10.1007/s15010-014-0593-5 [ Links ]
HUSSAIN S, SALAHUDDIN N, AHMAD I, SALAHUDDIN I and JOOMA R (1995) Rhinocerebral invasive mycosis: occurence in immunocompetent individuals. Eur. J. Radiol. 20 (2) 151-155. https://doi.org/10.1016/0720-048x(95)00644-6 [ Links ]
ILEA RC (2009) Intensive livestock farming: Global trends, increased environmental concerns, and ethical solutions. J. Agric. Environ. Ethics. 22 (2) 153-167. https://doi.org/10.1007/s10806-008-9136-3 [ Links ]
IPCC (2018) Summary for Policymakers. Global Warming of 1.5°C. In: Masson-Delmotte V, Zhai P, Portner H-O, Roberts D, Skea J, Shukla PR, Pirani A, Moufouma-Okia W, Pean C, Pidcock R, Connors S, Matthews JBR, Chen Y, Zhou X, Gomis MI, Lonnoy E, Maycock T, Tignor M, and Waterfield T (eds.) Global Warming of 1.5°C. World Meteorological Organization, Geneva. 32 pp. https://www.ipcc.ch/sr15/ [ Links ]
IPCC (2021) Summary for Policymakers. In: Masson-Delmotte V, Zhai P, Pirani A, Connors SL , Pean C, Berger S, Caud N, Chen Y, Goldfarb L, Gomis MI, Huang M, Leitzell K, Lonnoy E, Matthews JBR, Maycock TK, Waterfield T, Yelekci O, Yu R and Zhou B (eds.). Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press. https://www.ipcc.ch/report/ar6/wg1/ (Accessed 18 March 2022). [ Links ]
IWEN PC, SCHUTTE SD, FLORESCU DF, NOEL-HURST RK and SIGLER L (2012) Invasive Scopulariopsis brevicaulis infection in an immunocompromised patient and review of prior cases caused by Scopulariopsis and Microascus species. Med. Mycol. 50 (6) 561-569. https://doi.org/10.3109/13693786.2012.675629 [ Links ]
JACKSON BR, CHOW N, FORSBERG K, LITVINTSEVA AP, LOCKHART SR, WELSH R, VALLABHANENI S and CHILLER T (2019) On the origins of a species: What might explain the rise of Candida auris? J. Fungi. 5 (3). https://doi.org/10.3390/jof5030058JARVIS [ Links ]
JARVIS WR (1995) Epidemiology of nosocomial fungal infections, with emphasis on Candida species. Clin. Infect. Dis. 20 (6) 1526-1530. https://doi.org/10.1093/clinids/20.6.1526 [ Links ]
JOSHI A, SINGH R, SHAH MS, UMESH S and KHATTRY N (2010) Subcutaneous mycosis and fungemia by Aureobasidium pullulans: A rare pathogenic fungus in a post allogeneic BM transplant patient. Bone Marrow Transplant. 45 (1) 203-204. https://doi.org/10.1038/bmt.2009.111 [ Links ]
JUNEJA D, BORAH AK, NASA P, SINGH O, JAVERI Y and DANG R (2012) Candida sake candidaemia in non-neutropenic critically ill patients: A case series. Critical Care Resusc. 14 (1) 187-191. [ Links ]
JUNG J, MOON YS, YOO JA, LIM JH, JEONG J and JUN JB (2018) Investigation of a nosocomial outbreak of fungemia caused by Candida pelliculosa (Pichia anomala) in a Korean tertiary care center. J. Microbiol. Immunol. Infect. 51 (6) 794-801. https://doi.org/10.1016/j.jmii.2017.05.005 [ Links ]
JUNG KW, LEE KT, AVERETTE AF, HOY MJ, EVERITT J, HEITMAN J and BAHN YS (2018) Evolutionarily conserved and divergent roles of Unfolded Protein Response (UPR) in the pathogenic Cryptococcus species complex. Sci. Rep. 8 (1) 1-15. https://doi.org/10.1038/s41598-018-26405-5 [ Links ]
JUVVADI PR, KUROKI Y, ARIOKA M, NAKAJIMA H and KITAMOTO K (2003) Functional analysis of the calcineurin-encoding gene cnaA from Aspergillus oryzae: Evidence for its putative role in stress adaptation. Arch. Microbiol. 179 (6) 416-422. https://doi.org/10.1007/s00203-003-0546-3 [ Links ]
JUVVADI PR, LAMOTH F and STEINBACH WJ (2014) Calcineurin as a multifunctional regulator: Unraveling novel functions in fungal stress responses, hyphal growth, drug resistance, and pathogenesis. Fungal Biol. Rev. 28 (2) 56-69. https://doi.org/10.1016/j.fbr.2014.02.004 [ Links ]
KACZMARSKI EB, LIU YIN JA, TOOTH JA, LOVE EM and DELAMORE IW (1986) Systemic infection with Aureobasidium pullulans in a leukaemic patient. J. Infect. 13 (3) 289-291. https://doi.org/10.1016/S0163-4453(86)91388-5 [ Links ]
KASSAMALI H, ANAISSIE E, RO J, ROLSTON K, KANTARJIAN H, FAINSTEIN V and BODEY GP (1987) Disseminated Geotrichum candidum infection. J. Clin. Microbiol. 25 (9) 1782-1783. https://doi.org/10.1128/jcm.25.9.1782-1783.1987 [ Links ]
KEAN R, BROWN J, GULMEZ D, WARE A and RAMAGE G (2020) Candida auris: A decade of understanding of an enigmatic pathogenic yeast. J. Fungi. 6 (1) 30. https://doi.org/10.3390/jof6010030 [ Links ]
KEDRA M (2020) Regional response to global warming: Water temperature trends in semi-natural mountain river systems. Water. 12 (1). https://doi.org/10.3390/w12010283 [ Links ]
KENYON C, BONORCHIS K, CORCORAN C, MEINTJES G, LOCKETZ M, LEHLOENYA R, VISMER HF, NAICKER P, PROZESKY H, VAN WYK M and co-authors (2013) A dimorphic fungus causing disseminated infection in South Africa. New England J. Med. 369 (15) 1416-1424. https://doi.org/10.1056/nejmoa1215460 [ Links ]
KOEHLER P, TACKE D and CORNELY OA (2016) Bone and joint infections by Mucorales, Scedosporium, Fusarium and even rarer fungi. Crit. Rev. Microbiol. 42 (1) 158-171. https://doi.org/10.3109/1040841X.2014.910749 [ Links ]
KORDOSSIS T, AVLAMI A, VELEGRAKI A, STEFANOU I, GEORGAKOPOULOS G, PAPALAMBROU C and LEGAKIS NJ (1998) First report of Cryptococcus laurentii meningitis and a fatal case of Cryptococcus albidus cryptococcaemia in AIDS patients. Med. Mycol. 36 (5) 335-339. https://doi.org/10.1080/02681219880000521 [ Links ]
KOUSHA M, TADI R and SOUBANI AO (2011) Pulmonary aspergillosis: a clinical review. Eur. Respir. Rev. 20 (121) 156-174. https://doi.org/10.1183/09059180.00001011 [ Links ]
KUJATH P and KUJATH C (2010) Complicated skin, skin structure and soft tissue infections are we threatened by multi-resistant pathogens? Eur. J. Med. Res. 15 (12) 544-553. https://doi.org/10.1186/2047-783x-15-12-544 [ Links ]
KWASNIEWSKA, K (1988) Horizontal distribution and density of yeasts and filamentous fungi in Lake St. Clair water. J. Great Lakes Res. 14 (4) 438-443. https://doi.org/10.1016/S0380-1330(88)71577-4 [ Links ]
LAMOTH F, LOCKHART SR, BERKOW EL and CALANDRA T (2018) Changes in the epidemiological landscape of invasive candidiasis. J. Antimicrob. Chemotherapy. 73 i4-i13. https://doi.org/10.1093/jac/dkx444 [ Links ]
LAZERA MS, PIRES FDA, CAMILLO-COURA L, NISHIKAWA MM, BEZERRA CCF, TRILLES L and WANKE B (1996) Natural habitat of Cryptococcus neoformans var. neoformans in decaying wood forming hollows in living trees. J. Med. Vet. Mycol. 34 (2) 127-131. https://doi.org/10.1080/02681219680000191 [ Links ]
LEACH MD, KLIPP E, COWEN LE and BROWN AJP (2012) Fungal Hsp90: A biological transistor that tunes cellular outputs to thermal inputs. Nat. Rev. Microbiol. 10 (10) 693-704. https://doi.org/10.1038/nrmicro2875 [ Links ]
LEAW SN, CHANG HC, BARTON R, BOUCHARA JP and CHANG TC (2007) Identification of medically important Candida and non- Candida yeast species by an oligonucleotide array. J. Clin. Microbiol. 45 (7) 2220-2229. https://doi.org/10.1128/jcm.00543-07 [ Links ]
LEVENSON D, PFALLER MA, SMITH MA, HOLLIS R, GERARDEN T, TUCCI CB and ISENBERG HD (1991) Candida zeylanoides: Another opportunistic yeast. J. Clin. Microbiol. 29 (8) 1689-1692. https://doi.org/10.1128/jcm.29.8.1689-1692.1991 [ Links ]
LEYNGOLD I, OLIVI A, ISHII M, BLITZ A, BURGER P, SUBRAMANIAN PS and GALLIA G (2014) Acute chiasmal abscess resulting from perineural extension of invasive sino-orbital aspergillosis in an immunocompetent patient. World Neurosurg. 81 (1) 203-e1. https://doi.org/10.1016/j.wneu.2013.08.018 [ Links ]
LINEMAN M, DO Y, KIM JY and JOO GJ (2015) Talking about climate change and global warming. PloS ONE. 10 (9) p.e0138996. https://doi.org/10.1371/journal.pone.0138996 [ Links ]
LIPKE PN (2018) What we do not know about fungal cell adhesion molecules. J. Fungi. 4 (2). https://doi.org/10.3390/jof4020059 [ Links ]
LIU S, XIE Z, LIU B, WANG Y, GAO J, ZENG Y, XIE J, XIE Z, JIA B, QIN P, and co-authors (2020) Global river water warming due to climate change and anthropogenic heat emission. Glob. Planetary Change. 193 (July) 103289. https://doi.org/10.1016/j.gloplacha.2020.103289 [ Links ]
LIU X, WANG J, LIU T, KONG W, HE X, JIN Y and ZHANG B (2015) Effects of assimilable organic carbon and free chlorine on bacterial growth in drinking water. PLoS ONE. 10 (6) 1-11. https://doi.org/10.1371/journal.pone.0128825 [ Links ]
LIYANAGE CP and YAMADA K (2017) Impact of population growth on the water quality of natural water bodies. Sustainability. 9 (8). https://doi.org/10.3390/su9081405 [ Links ]
LO RE V, FISHMAN NO and NACHAMKIN I (2003) Recurrent catheter-related Rhodotorula rubra infection. Clin. Microbiol. Infect. 9 (8) 897-900. https://doi.org/10.1046/j.1469-0691.2003.00641.x [ Links ]
LOCKHART SR, MESSER SA, PFALLER MA and DIEKEMA DJ (2009) Identification and susceptibility profile of Candida fermentati from a worldwide collection of Candida guilliermondii clinical isolates. J. Clin. Microbiol. 47 (1) 242-244. https://doi.org/10.1128/JCM.01889-08 [ Links ]
LOHOUÉ PETMY J, LANDO AJ, KAPTUE L, TCHINDA V and FOLEFACK M (2004) Superficial mycoses and HIV infection in Yaounde. J. Eur. Acad. Dermatol. Venereol. 18 (3) 301-304. https://doi.org/10.1111/j.1468-3083.2004.00924.x [ Links ]
LONGCORE JE, PESSIER AP and NICHOLS DK (1999) Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia. 91 (2) 219-227. https://doi.org/10.1080/00275514.1999.12061011 [ Links ]
LOW CY and ROTSTEIN C (2011) Emerging fungal infections in immunocompromised patients. F1000 Med. Rep. 3 (1) 1-8. https://doi.org/10.3410/M3-14 [ Links ]
LUPLERTLOP N, PUMEESAT P, MUANGKAEW W, WONGSUK T and ALASTRUEY-IZQUIERDO A (2016) Environmental screening for the Scedosporium apiospermum species complex in public parks in Bangkok, Thailand. PLoS ONE. 11 (7) 1-10. https://doi.org/10.1371/journal.pone.0159869 [ Links ]
MACHIDO D, YAKUBU S and EZEONUEGBU B (2015) Composition of fungal flora in raw refinery effluent, effluent retention pond and a treated effluent recipient river. J. Appl. Sci. Environ. Manage. 18 (4) 592. https://doi.org/10.4314/jasem.v18i4.5 [ Links ]
MADHAVAN T, EISSES J and QUINN EL (1976) Infections due to Trichosporon cutaneum, an uncommon systemic pathogen. Henry Ford Hospital Med. J. 24 (1) 27-30. https://scholarlycommons.henryford.com/hfhmedjournal/vol24/iss1/5 [ Links ]
MAGDITCH DA, LIU TB, XUE C and IDNURM A (2012) DNA mutations mediate microevolution between host-adapted forms of the pathogenic fungus Cryptococcus neoformans. PLoS Pathogens. 8 (10). https://doi.org/10.1371/journal.ppat.1002936 [ Links ]
MAGWAZA NM, NXUMALO EN, MAMBA BB and MSAGATI TAM (2017) The occurrence and diversity of waterborne fungi in African aquatic systems: Their impact on water quality and human health. Int. J. Environ. Res. Public Health. 14 (5). https://doi.org/10.3390/ijerph14050546 [ Links ]
MAHESHWARI R, BHARADWAJ G and BHAT MK (2000) Thermophilic fungi: their physiology and enzymes. Microbiol. Mol. Biol. Rev. 64 (3) 461-488. https://doi.org/10.1128/mmbr.64.3.461-488.2000 [ Links ]
MALCOLM JR, LIU C, NEILSON RP, HANSEN L and HANNAH LEE (2006) Global warming and extinctions of endemic species from biodiversity hotspots. Conserv. Biol. 20 (2) 538-548. https://doi.org/10.1111/j.1523-1739.2006.00364.x [ Links ]
MARIER R, ZAKHIREH B, DOWNS J, WYNNE B, HAMMOND GL and ANDRIOLE VT (1978) Trichosporon cutaneum endocarditis. Scan. J. Infect. Dis. 10 (3) 255-256. https://doi.org/10.3109/inf.1978.10.issue-3.19 [ Links ]
MARTINS C, VARELA A, LECLERCQ CC, NÚÑEZ O, VÈTROVSKY T, RENAUT J, BALDRIAN P and SILVA PEREIRA C (2018) Specialisation events of fungal metacommunities exposed to a persistent organic pollutant are suggestive of augmented pathogenic potential. Microbiome. 6 (1) 208. https://doi.org/10.1186/s40168-018-0589-y [ Links ]
MAY RC, STONE NRH, WIESNER DL, BICANIC T and NIELSEN K (2016) Cryptococcus: From environmental saprophyte to global pathogen. Nat. Rev. Microbiol. 14 (2) 106-117. https://doi.org/10.1038/nrmicro.2015.6 [ Links ]
MCMEEKIN T, BOWMAN J, MCQUESTIN O, MELLEFONT L, ROSS T and TAMPLIN M (2008) The future of predictive microbiology: Strategic research, innovative applications and great expectations. Int. J. Food Microbiol. 128 (1) 2-9. https://doi.org/10.1016/j.ijfoodmicro.2008.06.026 [ Links ]
MEDEIROS AO, MISSAGIA BS, BRANDÃO LR, CALLISTO M, BARBOSA FAR and ROSA CA (2012) Water quality and diversity of yeasts from tropical lakes and rivers from the Rio Doce basin in Southeastern Brazil. Braz. J. Microbiol. 43 (4) 1582-1594. https://doi.org/10.1590/S1517-83822012000400043 [ Links ]
MEIS JF and CHOWDHARY A (2020) Candida auris-"ten years after." J. Fungi 6 (1) 10-11. https://doi.org/10.3390/jof6010002 [ Links ]
MERKUR AB and HODGE WG (2002) Rhodotorula rubra endophthalmitis in an HIV positive patient. Brit. J. Ophthalmol. 86 (12) 1444-1445. http://dx.doi.org/10.1136/bjo.86.12.1444 [ Links ]
MICELI MH, DÍAZ JA and LEE SA (2011) Emerging opportunistic yeast infections. Lancet Infect. Dis. 11 (2) 142-151. https://doi.org/10.1016/S1473-3099(10)70218-8 [ Links ]
MIJITI J, PAN B, DE HOOG S, HORIE Y, MATSUZAWA T, YILIFAN Y, LIU Y, ABLIZ P, PAN W, DENG D, and co-authors (2017) Severe chromoblastomycosis-like cutaneous infection caused by Chrysosporium keratinophilum. Front. Microbiol. 8 (JAN) 20122017. https://doi.org/10.3389/fmicb.2017.00083 [ Links ]
MOAZENI M, ASGARI S and NABILI M (2018) Nosocomial fungal infections: Epidemiology, diagnosis, treatment and prevention. J. Mazandaran Univ. Med. Sci. 28 (160) 182-212. http://jmums.mazums.ac.ir/article-1-11237-en.html [ Links ]
MOLINA-LEYVA A, RUIZ-CARRASCOSA JC, LEYVA-GARCIA A and HUSEIN-ELAHMED H (2013) Cutaneous Cryptococcus laurentii infection in an immunocompetent child. Int. J. Infect. Dis. 17 (12) e1232-e1233. https://doi.org/10.1016/j.ijid.2013.04.017 [ Links ]
MONAPATHI M, BEZUIDENHOUT C and RHODE O (2020) Physicochemical parameters and culturable yeast diversity in surface water: A consequence of pollution. Water SA. 46 (4) 593-601. https://doi.org/10.17159/wsa/2020.v46.i4.9072 [ Links ]
MONAPATHI ME, BEZUIDENHOUT CC and RHODE OHJ (2017) Water quality and antifungal susceptibility of opportunistic yeast pathogens from rivers. Water Sci. Technol. 75 (6) 1319-1331. https://doi.org/10.2166/wst.2016.580 [ Links ]
MORALES EG, IORIZZO M, LUCCHINI GM and MAINETTI C (2019) Trichophyton violaceum: an emerging pathogen in Southern Switzerland. Dermatology. 235 (5) 434-439. https://doi.org/10.1159/000501271 [ Links ]
MORAN GP, COLEMAN DC and SULLIVAN DJ (2012) Candida albicans versus Candida dubliniensis: why is C. albicans more pathogenic? Int. J. Microbiol. 2012. https://doi.org/10.1155/2012/205921 [ Links ]
MORRISON J, QUICK MC and FOREMAN MGG (2018) Climate change in the Fraser River Watershed: Flow and temperature climate change in the Fraser River watershed: flow and temperature projections. 1694 (June 2002) 230-244. https://doi.org/10.1016/S0022-1694(02)00065-3 [ Links ]
MOSCHETTI G, ALFONZO A, FRANCESCA N (2017) Yeasts in Birds. In: Buzzini P, Lachance MA and Yurkov A (eds) Yeasts in Natural Ecosystems: Diversity. Springer, Cham. https://doi.org/10.1007/978-3-319-62683-3_14 [ Links ]
MUÑOZ P, BOUZA E, CUENCA-ESTRELLA M, EIROS JM, PÉREZ MJ, SÁNCHEZ-SOMOLINOS M, RINCÓN C, HORTAL J and PELÁEZ T (2005) Saccharomyces cerevisiae fungemia: An emerging infectious disease. Clin. Infect. Dis. 40 (11) 1625-1634. https://doi.org/10.1086/429916 [ Links ]
NAGARATHNAMMA T, CHUNCHANUR SK, RUDRAMURTHY SM, VINEETHA KR, RAMAMURTHY K, JOSEPH J and AMBICA R (2017) Outbreak of Pichia kudriavzevii fungaemia in a neonatal intensive care unit. J. Med. Microbiol. 66 (12) 1759-1764. https://doi.org/10.1099/jmm.0.000645 [ Links ]
NARANJO-ORTIZ MA and GABALDÓN T.(2019) Fungal evolution: major ecological adaptations and evolutionary transitions. Biol. Rev. 94 (4) 1443-1476. https://doi.org/10.1111/brv.12510 [ Links ]
NEELY WJ, GREENSPAN SE, RIBEIRO LP, CARVALHO T, MARTINS RA, RODRIGUEZ D, ROHR JR, HADDAD CFB, TOLEDO LF and BECKER CG (2020) Synergistic effects of warming and disease linked to high mortality in cool-adapted terrestrial frogs. Biol. Conserv. 245 (October 2019) 108521. https://doi.org/10.1016/j.biocon.2020.108521 [ Links ]
NEWBOUND M, MCCARTHY MA and LEBEL T (2010) Fungi and the urban environment: A review. Landscape Urban Plann. 96 (3) 138-145. https://doi.org/10.1016/j.landurbplan.2010.04.005 [ Links ]
NICHOLS CB, PERFECT ZH and ALSPAUGH JA (2007) A Ras1-Cdc24 signal transduction pathway mediates thermotolerance in the fungal pathogen Cryptococcus neoformans. Mol. Microbiol. 63 (4) 1118-1130. https://doi.org/10.1111/j.1365-2958.2006.05566.x [ Links ]
NIELSEN K, DE OBALDIA AL and HEITMAN J (2007) Cryptococcus neoformans mates on pigeon guano: implications for the realized ecological niche and globalization. Eukaryotic Cell. 6 (6) 949-959. https://doi.org/10.1128/ec.00097-07 [ Links ]
NOBREGA DE ALMEIDA J, CAMPOS S, THOMAZ DY, THOMAZ L, DE ALMEIDA RKG, DEL NEGRO GMB, GIMENES VF, GRENFELL RC, MOTTA AL, ROSSI F and co-authors (2018) Candida blankii: An emergent opportunistic yeast with reduced susceptibility to antifungals correspondence. Emerging Microbes and Infections. 7 (1) 1-4. https://doi.org/10.1038/s41426-017-0015-8 [ Links ]
NORDELL B (2003) Thermal pollution causes global warming. Glob. Planet. Change. 38 (3-4) 305-312. https://doi.org/10.1016/S0921-8181(03)00113-9 [ Links ]
NOVAK BABIC M, ZALAR P, ZENKO B, DZEROSKI S and GUNDE-CIMERMAN N (2016) Yeasts and yeast-like fungi in tap water and groundwater, and their transmission to household appliances. Fungal Ecol. 20 30-39. https://doi.org/10.1016/j.funeco.2015.10.001 [ Links ]
NOVAK BABIC M, ZUPANCIC J, BRANDÃO J and GUNDE-CIMERMAN N (2018) Opportunistic water-borne human pathogenic filamentous fungi unreported from food. Microorganisms. 6 (3) 79. https://doi.org/10.3390/microorganisms6030079 [ Links ]
NUSSLÉ S, MATTHEWS KR and CARLSON SM (2015) Mediating water temperature increases due to livestock and global change in high elevation meadow streams of the golden trout wilderness. PLoS ONE. 10 (11) 1-22. https://doi.org/10.1371/journal.pone.0142426 [ Links ]
O'GORMAN CM and FULLER HT (2008) Prevalence of culturable airborne spores of selected allergenic and pathogenic fungi in outdoor air. Atmos. Environ. 42 (18) 4355-4368. https://doi.org/10.1016/j.atmosenv.2008.01.009 [ Links ]
OCANSEY BK, PESEWU GA, CODJOE FS, OSEI-DJARBENG S, FEGLO PK and DENNING DW (2019) Estimated burden of serious fungal infections in Ghana. J. Fungi. 5 (2) 38. https://doi.org/10.3390/jof5020038 [ Links ]
OLIVEIRA FFM, PAES HC, PECONICK LDF, FONSECA FL, MARINA CLF, BOCCA AL, HOMEM-DE-MELLO M, RODRIGUES ML, ALBUQUERQUE P, NICOLA AM, and co-authors (2020) Erg6 affects membrane composition and virulence of the human fungal pathogen Cryptococcus neoformans. Fungal Genet. Biol. 140 (March) 103368. https://doi.org/10.1016/j.fgb.2020.103368 [ Links ]
ORTIZ-VERA MP, OLCHANHESKI LR, DA SILVA EG, DE LIMA FR, MARTINEZ LR del PR, SATO MIZ, JAFFÉ R, ALVES R, ICHIWAKI S, PADILLA G, and co-authors (2018) Influence of water quality on diversity and composition of fungal communities in a tropical river. Sci. Rep. 8 (1) 1-9. https://doi.org/10.1038/s41598-018-33162-y [ Links ]
PAN W, LIAO W, HAGEN F, THEELEN B, SHI W, MEIS JF and BOEKHOUT T (2012) Meningitis caused by Filobasidium uniguttulatum: Case report and overview of the literature. Mycoses 55 (2) 105-109. https://doi.org/10.1111/j.1439-0507.2011.02054.x [ Links ]
PARAPOULI M, VASILEIADIS A, AFENDRA AS and HATZILOUKAS E (2020) Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 6 (1) 1-31. https://doi.org/10.3934/microbiol.2020001 [ Links ]
PARK JH, OH J, SANG H, SHRESTHA B, LEE H, KOO J, CHO S il, CHOI JS, LEE MH, KIM J, and co-authors (2019) Identification and antifungal susceptibility profiles of Cyberlindnera fabianii in Korea. Mycobiology 47 (4) 449-456. https://doi.org/10.1080/12298093.2019.1651592 [ Links ]
PARTS L, CUBILLOS FA, WARRINGER J, JAIN K, SALINAS F, BUMPSTEAD SJ, MOLIN M, ZIA A, SIMPSON JT, QUAIL MA, and co-authors (2011) Revealing the genetic structure of a trait by sequencing a population under selection. Genome Res. 21 (7) 11311138. https://doi.org/10.1101/gr.116731.110 [ Links ]
PASQUETTI M, MIN ARM, SCACCHETTI S, DOGLIERO A and PEANO A (2017) Infection by Microsporum canis in paediatric patients: A veterinary perspective. Vet. Sci. 4 (3) 2-7. https://doi.org/10.3390/vetsci4030046 [ Links ]
PASTOR FJ and GUARRO J (2008) Alternaria infections: laboratory diagnosis and relevant clinical features. Clin. Microbiol. Infect. 14 (8) 734-746. https://doi.org/10.1111/j.1469-0691.2008.02024.x [ Links ]
PAULUSSEN C, HALLSWORTH JE, ÁLVAREZ-PÉREZ S, NIERMAN WC, HAMILL PG, BLAIN D, REDIERS H and LIEVENS B (2017) Ecology of aspergillosis: insights into the pathogenic potency of Aspergillus fumigatus and some other Aspergillus species. Microb. Biotechnol. 10 (2) 296-322. https://doi.org/10.1111/1751-7915.12367 [ Links ]
PEREIRA T, VILAPRINYO E, BELLI G, HERRERO E, SALVADO B, SORRIBAS A, ALTÉS G and ALVES R (2018) Quantitative operating principles of yeast metabolism during adaptation to heat stress. Cell Reports. 22 (9) 2421-2430. https://doi.org/10.1016/j.celrep.2018.02.020 [ Links ]
PEREIRA VJ, BASÍLIO MC, FERNANDES D, DOMINGUES M, PAIVA JM, BENOLIEL MJ, CRESPO MT and SAN ROMÃO M (2009) Occurrence of filamentous fungi and yeasts in three different drinking water sources. Water Res. 43 (15) 3813-3819. https://doi.org/10.1016/j.watres.2009.05.024 [ Links ]
PERFECT JR (2006) Cryptococcus neoformans: The yeast that likes it hot. FEMS Yeast Res. 6 (4) 463-468. https://doi.org/10.1111/j.1567-1364.2006.00051.x [ Links ]
PETTER R, KANG BS, BOEKHOUT T, DAVIS BJ and KWON-CHUNG KJ (2001) A survey of heterobasidiomycetous yeasts for the presence of the genes homologous to virulence factors of Filobasidiella neoformans, CNLAC1 and CAP59. Microbiology. 147 (8) 2029-2036. https://doi.org/10.1099/00221287-147-8-2029 [ Links ]
PFALLER MA, DIEKEMA DJ, COLOMBO AL, KIBBLER C, NG KP, GIBBS DL, NEWELL VA, FINQUELIEVICH J, TIRABOSCHI N, ELLIS D and co-authors (2006) Candida rugosa, an emerging fungal pathogen with resistance to azoles: Geographic and temporal trends from the ARTEMIS DISK Antifungal Surveillance Program. J. Clin. Microbiol. 44 (10) 3578-3582. https://doi.org/10.1128/JCM.00863-06 [ Links ]
PIETRYCZUK A, CUDOWSKI A, HAUSCHILD T, SWISLOCKA M, WIECKO A and KARPOWICZ M (2018) Abundance and Species Diversity of Fungi in Rivers with Various Contaminations. Curr. Microbiol. 75 (5) 630-638. https://doi.org/10.1007/s00284-017-1427-3 [ Links ]
PINNA A, CARTA F, ZANETTI S, SANNA S and SECHI LA (2001) Endogenous Rhodotorula minuta and Candida albicans endophthalmitis in an injecting drug user. Brit. J. Ophthalmol. 85 (6) 754-754. http://dx.doi.org/10.1136/bjo.85.6.754-f [ Links ]
POHLE I, HELLIWELL R, AUBE C, GIBBS S, SPENCER M and SPEZIA L (2019) Citizen science evidence from the past century shows that Scottish rivers are warming. Sci. Total Environ. 659 53-65. https://doi.org/10.1016/j.scitotenv.2018.12.325 [ Links ]
POSTMA F (2016) Rhizofiltration of urban effluent: Microbial ecology and conceptual treatment mechanisms. Doctoral dissertation, Stellenbosch University. http://scholar.sun.ac.za/handle/10019.1/98274 [ Links ]
POTH T, SEIBOLD M, WERCKENTHIN C and HERMANNS W (2010) First report of a Cryptococcus magnus infection in a cat. Med. Mycol. 48 (7) 1000-1004. https://doi.org/10.3109/13693786.2010.489584 [ Links ]
POUNDS JA, BUSTAMANTE MR, COLOMA LA, CONSUEGRA JA, FOGDEN MPL, FOSTER PN, LA MARCA E, MASTERS KL, MERINO-VITERI A, PUSCHENDORF R and co-authors (2006) Widespread amphibian extinctions from epidemic disease driven by global warming. Nature. 439 (7073) 161-167. https://doi.org/10.1038/nature04246 [ Links ]
PRINSLOO B, WELDHAGEN GF and BLAINE RW (2003) Candida famata central nervous system infection. S. Afr. Med. J. 93 (8) 601-602. [ Links ]
QADRI SH, AL DAYEL F, STRAMPFER MJ and CUNHA BA (1988) Urinary tract infection caused by Hansenula anomala. Mycopathologia. 104 (2) 99-101. https://doi.org/10.1007/BF00436934 [ Links ]
RADOSAVLJEVIC M, KOENIG H, LETSCHER-BRU V, WALLER J, MALOISEL F, LIOURE B and HERBRECHT R (1999) Candida catenulata fungemia in a cancer patient. J. Clin. Microbiol. 37 (2) 475 -477. https://doi.org/10.1128/jcm.37.2.475-477.1999 [ Links ]
RESTREPO A, BAUMGARDNER DJ, BAGAGLI E, JR. CRC, MCGINNIS MR, LÁZERA MS, BARBOSA FH, BOSCO SMG, CAMARGO ZPF, COELHO KIR, and co-authors (2000) Clues to the presence of pathogenic fungi in certain environments. Med. Mycol. 38 (s1) 67-77. https://doi.org/10.1080/mmy.38.s1.67.77 [ Links ]
RHODES J and FISHER MC (2019) Global epidemiology of emerging Candida auris. Curr. Opin. Microbiol. 52 84-89. https://doi.org/10.1016/j.mib.2019.05.008 [ Links ]
RIZZITELLI G, GUANZIROLI E, MOSCHIN A, SANGALLI R and VERALDI S (2016) Onychomycosis caused by Trichosporon mucoides. Int. J. Infect. Dis. 42 61-63. https://doi.org/10.1016Zj.ijid.2015.11.011 [ Links ]
ROBERT V, CARDINALI G and CASADEVALL A (2015) Distribution and impact of yeast thermal tolerance permissive for mammalian infection. BMC Biol. 13 (1) 1-14. https://doi.org/10.1186/s12915-015-0127-3 [ Links ]
ROBERT VA and CASADEVALL A (2009) Vertebrate endothermy restricts most fungi as potential pathogens. J. Infect. Dis. 200 (10) 1623-1626. https://doi.org/10.1086/644642 [ Links ]
RODRIGUEZ JY, LE PAPE P, LOPEZ O, ESQUEA K, LABIOSA AL and ALVAREZ-MORENO C (2020) Candida auris: A latent threat to critically ill patients with coronavirus disease 2019. Clin. Infect. Dis. https://doi.org/10.1093/cid/ciaa1595 [ Links ]
ROHR JR, RAFFEL TR, ROMANSIC JM, MCCALLUM H and HUDSON PJ (2008) Evaluating the links between climate, disease spread, and amphibian declines. Proc. Natl Acad. Sci. USA. 105 (45) 17436-17441. https://doi.org/10.1073/pnas.0806368105 [ Links ]
RUAN SY, CHIEN JY, HOU YC and HSUEH PR (2010) Catheter-related fungemia caused by Candida intermedia. Int. J. Infect. Dis. 14 (2) e147-e149. https://doi.org/10.1016/j.ijid.2009.03.015 [ Links ]
SANDOVAL-DENIS M, SUTTON DA, FOTHERGILL AW, CANO-LIRA J, GENÉ J, DECOCK CA, DE HOOG GS and GUARRO J (2013) Scopulariopsis, a poorly known opportunistic fungus: Spectrum of species in clinical samples and in vitro responses to antifungal drugs. J. Clin. Microbiol. 51 (12) 3937-3943. https://doi.org/10.1128/JCM.01927-13 [ Links ]
SANZ AB, GARCÍA R, RODRÍGUEZ-PEÑA JM and ARROYO J (2018) The CWI pathway: Regulation of the transcriptional adaptive response to cell wall stress in yeast. J. Fungi. 4 (1) 1-12. https://doi.org/10.3390/jof4010001 [ Links ]
SATOH K, MAKIMURA K, HASUMI Y, NISHIYAMA Y, UCHIDA K and YAMAGUCHI H (2009) Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 53 (1) 41-44. https://doi.org/10.1111/j.1348-0421.2008.00083.x [ Links ]
SCHIAVANO GF, PARLANI L, SISTI M, SEBASTIANELLI G and BRANDI G (2014) Occurrence of fungi in dialysis water and dialysate from eight haemodialysis units in central Italy. J. Hosp. Infect. 86 (3) 194-200. https://doi.org/10.1016/j.jhin.2013.11.010 [ Links ]
SCHWARTZ I and DENNING D (2019) The estimated burden of fungal diseases in South Africa. J. Infect. Public Health 12 (1) 124. https://doi.org/10.1016/j.jiph.2018.10.064 [ Links ]
SCHWINN A, EBERT J and BROCKER EB (1995) Frequency of Trichophyton rubrum in tinea capitis. Mycoses. 38 (1-2) 1-7. https://doi.org/10.1111/j.1439-0507.1995.tb00001.x [ Links ]
SCOPPETTUOLO G, DONATO C, DE CAROLIS E, VELLA A, VACCARO L, LA GRECA A and FANTONI M (2014) Candida utilis catheter-related bloodstream infection. Med. Mycol. Case Rep. 6 70-72. https://doi.org/10.1016/j.mmcr.2014.10.003 [ Links ]
SCORZONI L, DE LUCAS MP, MESA-ARANGO AC, FUSCO-ALMEIDA AM, LOZANO E, CUENCA-ESTRELLA M, MENDES-GIANNINI MJ and ZARAGOZA O (2013) Antifungal efficacy during Candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE. 8 (3). https://doi.org/10.1371/journal.pone.0060047 [ Links ]
SETH-SMITH HMB, BÜCHLER AC, HINIC V, MEDINGER M, WIDMER AF and EGLI A (2020) Bloodstream infection with Candida kefyr/Kluyveromyces marxianus: case report and draft genome. Clin. Microbiol. Infect. 26 (4) 522-524. https://doi.org/10.1016/j.cmi.2019.11.014 [ Links ]
SFAKIANAKIS A, KRASAGAKIS K, STEFANIDOU M, MARAKI S, KOUTSOPOULOS A, KOFTERIDIS D, SAMONIS G and TOSCA A (2007) Invasive cutaneous infection with Geotrichum candidum - Sequential treatment with amphotericin B and voriconazole. Med. Mycol. 45 (1) 81-84. https://doi.org/10.1080/13693780600939948 [ Links ]
SHARMA V, SHANKAR J and KOTAMARTHI V (2006) Endogeneous endophthalmitis caused by Sporobolomyces salmonicolor. Eye. 20 (8) 945-946. https://doi.org/10.1038/sj.eye.6702051 [ Links ]
SHINDE R, MANTUR B, PATIL G, PARANDE M and PARANDE A (2008) Meningitis due to Rhodotorula glutinis in an HIV infected patient. Indian J. Med. Microbiol. 26 (4) 375-377. [ Links ]
SHOHAM S and LEVITZ SM (2005) The immune response to fungal infections. Brit. J. Haematol. 129 (5) 569-582. https://doi.org/10.1111/j.1365-2141.2005.05397.x [ Links ]
SIGEE D (2005) Freshwater Microbiology: Biodiversity and Dynamic Interactions of Microorganisms in the Aquatic Environment. John Wiley & Sons, Hoboken. [ Links ]
SILVA S, NEGRI M, HENRIQUES M, OLIVEIRA R, WILLIAMS DW and AZEREDO J (2012) Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 36 (2) 288-305. https://doi.org/10.1111/j.1574-6976.2011.00278.x [ Links ]
SKIADA A, PAVLEAS I and DROGARI-APIRANTHITOU M (2017) Rare fungal infectious agents: a lurking enemy. FlGGGResearch. 6 1917. https://doi.org/10.12688/f1000research.11124.1 [ Links ]
SLAVIKOVA E and VADKERTIOVA R (1997) Seasonal occurrence of yeasts and yeast-like organisms in the river Danube. Anton. Leeuw. Int. J. Gen. Mol. Microbiol. 72 (2) 77-80. https://doi.org/10.1023/A:1000287005253 [ Links ]
SO YS, JANG J, PARK G, XU J, OLSZEWSKI MA and BAHN YS (2018) Sho1 and msb2 play complementary but distinct roles in stress responses, sexual differentiation, and pathogenicity of Cryptococcus neoformans. Front. Endocrinol. 9 (December) 1-16. https://doi.org/10.3389/fmicb.2018.02958 [ Links ]
SOBER E (1981) The principle of parsimony. Brit. J. Philos. Sci. 32 (2) 145-156. https://doi.org/10.1093/bjps/32.2.145 [ Links ]
SOMBATMAITHAI A, PATTANAPRICHAKUL P, TUCHINDA P, SURAWAN T, MUANPRASART C, MATTHAPAN L and BUNYARATAVEJ S (2015) Tinea capitis caused by Trichophyton tonsurans presenting as an obscure patchy hair loss due to daily antifungal shampoo use. Dermatol. Practical Concept. 5 (2) 133-135. https://doi.org/10.5826/dpc.0502a27 [ Links ]
SOUSA MJ, ARDO Y and MCSWEENEY PLH (2001) Advances in the study of proteolysis during cheese ripening. Int. Dairy J. 11 (4-7) 327-345. https://doi.org/10.1016/S0958-6946(01)00062-0 [ Links ]
SPELLBERG B and MAERTENS J (2019) Mucormycosis. In: Principles and Practice of Transplant Infectious Diseases. 577-589. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-9034-4_34 [ Links ]
STEENBERGEN JN, SHUMAN HA and CASADEVALL A (2001) Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc. Natl Acad. Sci. 98 (26) 15245-15250. https://doi.org/10.1073/pnas.261418798 [ Links ]
STONE W, JONES B-L, WILSENACH J and BOTHA A (2012) External Ecological Niche for Candida albicans within Reducing, Oxygen-Limited Zones of Wetlands. Appl. Environ. Microbiol. 78 (7) 24432445. https://doi.org/10.1128/aem.06343-11 [ Links ]
SUGITA T, KAGAYA K, TAKASHIMA M, SUZUKI M, FUKAZAWA Y and NAKASE T (1999) A clinical isolate of Candida palmioleophila formerly identified as Torulopsis candida. Jap. J. Med. Mycol. 40 (1) 21-25. https://doi.org/10.3314/jjmm.40.21 [ Links ]
SULEYMAN G and ALANGADEN GJ (2016) Nosocomial fungal infections: epidemiology, infection control, and prevention. Infect. Dis. Clinics N. Am. 30 (4) 1023-1052. https://doi.org/10.1016/j.idc.2016.07.008 [ Links ]
TAJ-ALDEEN SJ, ABDULWAHAB A, KOLECKA A, DESHMUKH A, MEIS JF and BOEKHOUT T (2014) Uncommon opportunistic yeast bloodstream infections from Qatar. Med. Mycol. 52 (5) 549-553. https://doi.org/10.1093/mmycol/myu016 [ Links ]
TANG HJ, LAI CC and CHAO CM (2015) Central-line-associated bloodstream infection caused by Sporobolomyces salmonicolor. Infect. Control Hospital Epidemiol. 36 (9) 1111-1112. https://doi.org/10.1017/ice.2015.158 [ Links ]
TANSEY MR and BROCK TD (1972) The upper temperature limit for eukaryotic organisms. Proc. Natl Acad. Sci. 69 (9) 2426-2428. https://doi.org/10.1073/pnas.69.9.2426 [ Links ]
TERESHINA VM (2005) Thermotolerance in fungi: the role of heat shock proteins and trehalose. Microbiology. 74 (3) 247-257. https://doi.org/10.1007/s11021-005-0059-y [ Links ]
TOPLIS B, BOSCH C, SCHWARTZ IS, KENYON C, BOEKHOUT T, PERFECT JR and BOTHA A (2020) The virulence factor urease and its unexplored role in the metabolism of Cryptococcus neoformans. FEMS Yeast Res. 20 (4) p.foaa031. https://doi.org/10.1093/femsyr/foaa031 [ Links ]
TREGUIER P, DAVID M, GARGALA G, CAMUS V, STAMATOULLAS A, MENARD A-L, LENAIN P, CONTENTIN N, LEMASLE E, LANIC H, and co-authors (2018) Cyberlindnera jadinii (teleomorph Candida utilis) candidaemia in a patient with aplastic anaemia: a case report. JMM Case Reports. 5 (8) 1-3. https://doi.org/10.1099/jmmcr.0.005160 [ Links ]
TROWBRIDGE J, LUDMER LM, RIDDLE VD, LEVY CS and BARTH WF (1999) Candida lambica polyarthritis in a patient with chronic alcoholism. J. Rheumatol. 26 (8) 1846-1848. https://pubmed.ncbi.nlm.nih.gov/10451090/ [ Links ]
UN WATER (2006) UN World Water Development Report. URL: https://www.unwater.org/publications/water-shared-responsibility (Accessed 11 March 2021). [ Links ]
UNAIDS (2021) Global HIV & AIDS statistics - Fact sheet. https://www.unaids.org/en/resources/fact-sheet (Accessed 25 January 2022). [ Links ]
USAID (2015) Climate Change Information Fact Sheet SOUTHERN AFRICA. URL: https://www.usaid.gov/south-africa/environment (Accessed 18 March 2021). [ Links ]
VAN VLIET MTH, LUDWIG F, ZWOLSMAN JJG, WEEDON GP and KABAT P (2011) Global river temperatures and sensitivity to atmospheric warming and changes in river flow. Water Resour. Res. 47 (2). https://doi.org/10.1029/2010WR009198 [ Links ]
VAN WYK DAB, BEZUIDENHOUT CC and RHODE OHJ (2012) Diversity and characteristics of yeasts from water sources in the North West Province, South Africa. Water Sci. Technol. Water Suppl. 12 (4) 422-430. https://doi.org/10.2166/ws.2012.009 [ Links ]
VEAREY J (2011) Challenging urban health: towards an improved local government response to migration, informal settlements, and HIV in Johannesburg, South Africa. Glob. Health Action. 4. https://doi.org/10.3402/gha.v4i0.5898 [ Links ]
VERGHESE J, ABRAMS J, WANG Y and MORANO KA (2012) Biology of the heat shock response and protein chaperones: budding yeast (Saccharomyces cerevisiae) as a model system. Microbiol. Mol. Biol. Rev. 76 (2) 115-158. https://doi.org/10.1128/mmbr.05018-11 [ Links ]
VERGIDIS P, CLANCY CJ, SHIELDS RK, PARK SY, WILDFEUER BN, SIMMONS RL and NGUYEN MH (2016) Intra-abdominal candidiasis: The importance of early source control and antifungal treatment. PLoS ONE. 11 (4) 1-13. https://doi.org/10.1371/journal.pone.0153247 [ Links ]
VERNA J, LODDER A, LEE K, VAGTS A and BALLESTER R (1997) A family of genes required for maintenance of cell wall integrity and for the stress response in Saccharomyces cerevisiae. Proc. Natl Acad. Sci. 94 (25) 13804-13809. https://doi.org/10.1073/pnas.94.25.13804 [ Links ]
VERONES F, HANAFIAH MM, PFISTER S, HUIJBREGTS MAJ, PELLETIER GJ and KOEHLER A (2010) Characterization factors for thermal pollution in freshwater aquatic environments. Environ. Sci. Technol. 44 (24) 9364-9369. https://doi.org/10.1021/es102260c [ Links ]
VERVAEKE S, VANDAMME K, BOONE E, DE LAERE E, SWINNE D and SURMONT I (2008) A case of Candida lambica fungemia misidentified as Candida krusei in an intravenous drug abuser. Sabouraudia. 46 (8) 853-856. https://doi.org/10.1080/13693780802342552 [ Links ]
VILLANUEVA-LOZANO H, TREVIÑO-RANGEL RDJ, GONZÁLEZ GM, RAMÍREZ-ELIZONDO MT, LARA-MEDRANO R, ALEMAN-BOCANEGRA MC, GUAJARDO-LARA CE, GAONA-CHÁVEZ N, CASTILLEJA-LEAL F, TORRE-AMIONE G and MARTÍNEZ-RESÉNDEZ MF (2021) Outbreak of Candida auris infection in a COVID-19 hospital in Mexico. Clin. Microbiol. Infect. 27 (5) 813-816. https://doi.org/10.1016/j.cmi.2020.12.030 [ Links ]
WAGNER D, SANDER A, BERTZ H, FINKE J and KERN WV. (2005) Breakthrough invasive infection due to Debaryomyces hansenii (teleomorph Candida famata) and Scopulariopsis brevicaulis in a stem cell transplant patient receiving liposomal amphotericin B and caspofungin for suspected aspergillosis. Infection. 33 (5-6) 397-400. https://doi.org/10.1007/s15010-005-5082-4 [ Links ]
WAKE DB and VREDENBURG VT (2008) Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl Acad. Sci. 105 (Supplement 1) 11466-11473. https://doi.org/10.1073/pnas.0801921105 [ Links ]
WALSH TJ and DIXON DM (1989) Nosocomial aspergillosis: environmental microbiology, hospital epidemiology, diagnosis and treatment. Eur. J. Epidemiol. 5 (2) 131-142. https://doi.org/10.1007/bf00156818 [ Links ]
WANG J, LIU XD and LU J (2012) Urban river pollution control and remediation. Procedia Environ. Sci. 13 1856-1862. https://doi.org/10.1016/j.proenv.2012.01.179 [ Links ]
WARREN TA, MCTAGGART L, RICHARDSON SE and ZHANG SX (2010) Candida bracarensis bloodstream infection in an immunocompromised patient. J. Clin. Microbiol. 48 (12) 4677-4679. https://doi.org/10.1128/JCM.01447-10 [ Links ]
WARTENBERG A, LINDE J, MARTIN R, SCHREINER M, HORN F, JACOBSEN ID, JENULL S, WOLF T, KUCHLER K, GUTHKE R and KURZAI O (2014) Microevolution of Candida albicans in macrophages restores filamentation in a nonfilamentous mutant. PLoS Genet. 10 (12) 1004824. https://doi.org/10.1371/journal.pgen.1004824 [ Links ]
WATKINS R, KING J and JOHNSTON S (2017) Nutritional requirements and their importance for virulence of pathogenic Cryptococcus species. Microorganisms. 5 (4) 65. https://doi.org/10.3390/microorganisms5040065 [ Links ]
WEIMANN A and ONI T (2019) A systematised review of the health impact of urban informal settlements and implications for upgrading interventions in South Africa, a rapidly urbanising middle-income country. International Journal of Environmental Research and Public Health. 16 (19) 1-17. https://doi.org/10.3390/ijerph16193608 [ Links ]
WELSH O, VERA-CABRERA L and SALINAS-CARMONA MC (2007) Mycetoma. Clinics in Dermatology. 25 (2) 195-202. https://doi.org/10.1016/j.clindermatol.2006.05.011 [ Links ]
WEN Y, SCHOUPS G and GIESEN N van de (2017) Organic pollution of rivers : Combined threats of urbanization, livestock farming and global climate change. Nature Publishing Group (February) 1-9. https://doi.org/10.1038/srep43289 [ Links ]
WERTHMAN-EHRENREICH A (2021) Mucormycosis with orbital compartment syndrome in a patient with COVID-19. Am. J. Emerg. Med. 42 264-e5. https://doi.org/10.1016/j.ajem.2020.09.032 [ Links ]
WHITBY S, MADU EC and BRONZE MS (1996) Candida zeylanoides infective endocarditis complicating infection with the human immunodeficiency virus. Am. J. Med. Sci. 312 (3) 138-139. https://doi.org/10.1016/S0002-9629(15)41781-1 [ Links ]
WHO (2017) Water sanitation and hygeine (WASH). URL: https://www.who.int/health-topics/water-sanitation-and-hygiene-wash (Accessed 21 April 2021). [ Links ]
WIRTH F and GOLDANI LZ (2012) Epidemiology of Rhodotorula: An emerging pathogen. Interdisc. Perspect. Infect. Dis. 2012. https://doi.org/10.1155/2012/465717 [ Links ]
WONG B, KIEHN TE, EDWARDS F, BERNARD EM, MARCOVE RC, HARVEN EDE and ARMSTRONG D (1982) Bone infection caused by Debaryomyces hansenii in normal host: a case report. Notes. 16 (3) 545-548. https://doi.org/10.1128/jcm.16.3.545-548.1982 [ Links ]
YAMAGUCHI MU, RAMPAZZO RDCP, YAMADA-OGATTA SF, NAKAMURA CV, UEDA-NAKAMURA T and DIAS FILHO BP (2007) Yeasts and filamentous fungi in bottled mineral water and tap water from municipal supplies. Braz. Arch. Biol. Technol. 50 (1) 1-9. https://doi.org/10.1590/S1516-89132007000100001 [ Links ]
YANG DH, JUNG KW, BANG S, LEE JW, SONG MH, FLOYD-AVERETTE A, FESTA RA, IANIRI G, IDNURM A, THIELE DJ, and co-authors (2017) Rewiring of signaling networks modulating thermotolerance in the human pathogen Cryptococcus neoformans. Genetics. 205 (1) 201-219. https://doi.org/10.1534/genetics.116.190595 [ Links ]
YILMAZ-SEMERCI S, DEMIREL G and TASTEKIN A (2017) Wickerhamomyces anomalus blood stream infection in a term newborn with pneumonia. Turk. J. Pediatr. 59 (3) 349-351. https://doi.org/10.24953/turkjped.2017.03.021 [ Links ]
ZHOU J, CHEN M, CHEN H, PAN W and LIAO W (2014) Rhodotorula minuta as onychomycosis agent in a Chinese patient: First report and literature review. Mycoses. 57 (3) 191-195. https://doi.org/10.1111/myc.12143 [ Links ]
ZU T, VERNA J and BALLESTER R (2001) Mutations in WSC genes for putative stress receptors result in sensitivity to multiple stress conditions and impairment of Rlm1-dependent gene expression in Saccharomyces cerevisiae. Mol. Genet. Genomics. 266 (1) 142-155. https://doi.org/10.1007/s004380100537 [ Links ]
ZUZA-ALVES DL, SILVA-ROCHA WP and CHAVES GM (2017) An update on Candida tropicalis based on basic and clinical approaches. Front. Microbiol. 8 (OCT) 1-25. https://doi.org/10.3389/fmicb.2017.01927 [ Links ]
 Correspondence:
Correspondence:
Alfred Botha
Email: abo@sun.ac.za
Received: 18 July 2021
Accepted: 17 March 2022


{kind=link}
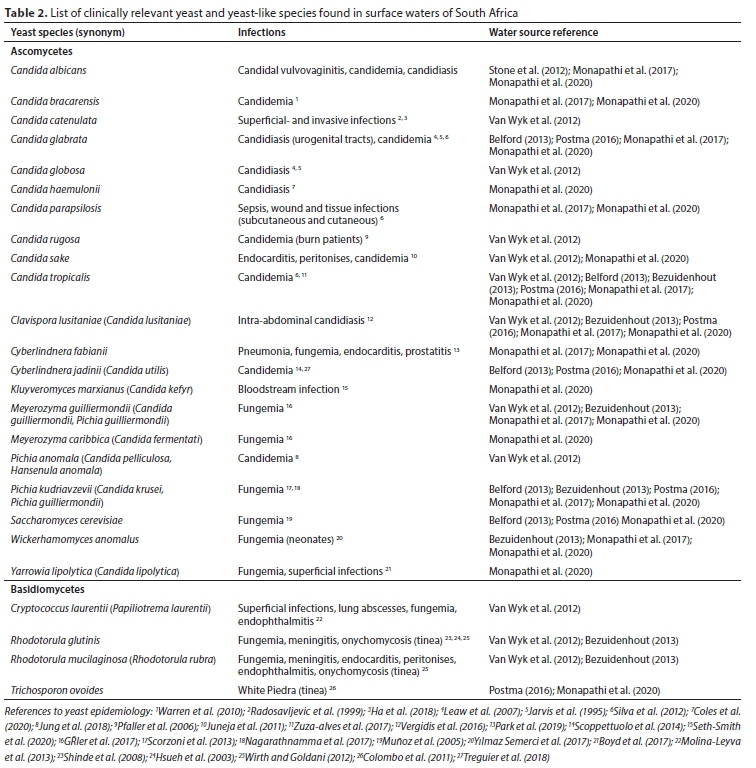{kind=link}
{kind=link}