










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkWater SA
versión On-line ISSN 1816-7950
versión impresa ISSN 0378-4738
Water SA vol.44 no.1 Pretoria ene. 2018
http://dx.doi.org/10.4314/wsa.v44i1.08
http://dx.doi.org/10.4314/wsa.v44i1.08
A glimpse into the littoral nutrient dynamics of a lake system connected to the sea
S TaljaardI, II, *; L van NiekerkI, II; DA LemleyII
INatural Resources and the Environment, Council for Scientific and Industrial Research (CSIR), Stellenbosch, South Africa
IIDepartment of Botany, Nelson Mandela University, PO Box 77000, Port Elizabeth 6031, South Africa
ABSTRACT
In South Africa more than 80% of estuaries are small, dynamic and predominantly linear systems. Nutrient characteristics in these systems are mostly influenced by external catchment processes, except during extended periods of closure. However, a small percentage of the country's estuaries (< 3%) comprise larger estuarine lakes mostly evolved from drowned river valleys. The physical properties of these systems suggest relatively low flushing rates, and the potentially stronger influence of in-situ processes on nutrient characteristics. This study investigates dissolved inorganic nutrient dynamics in the littoral zones of these estuarine lake systems, and potential influencing factors, using the Wilderness Lake System as a case study. Comparison of inter-annual and seasonal trends in dissolved inorganic nutrients (NOx-N, NH4-N and PO4-P) in river inflow and in the three lakes confirmed that external catchment fluxes are most likely not the sole, or even the dominant drivers of average long-term or seasonal dissolved inorganic nutrient patterns in the lake littoral zones. Relatively low NOx:NH4 ratios (mostly less than 1) in the lakes (especially in Eilandvlei and Langvlei) indicated a stronger influence of in-situ processes, such as bacteriological remineralisation, similar to observations in other lake type systems with low flushing rates. Low NO3:NH4 ratios are known to stimulate cyanobacteria, having the metabolic ability to potentially produce microcystins. Therefore, incremental dissolved inorganic nutrient and organic matter enrichment may cause these systems to tip into eutrophic, possibly toxic, aquatic states that will be difficult, if not impossible, to reverse given their weak flushing mechanisms. The findings from this study highlight the importance of appropriate water resource management, both in the catchments and within the floodplains of estuarine lake systems.
Keywords: Wilderness, estuarine lake, littoral zone, dissolved inorganic nutrients, catchment fluxes, in-situ processes
INTRODUCTION
Nutrient characteristics in lakes and estuaries are influenced by numerous factors, both externally and internally. Key external factors derive from the catchment (e.g. surface and sub-surface flow) and the sea (e.g. tidal intrusion), where the influence is mainly determined by the proportional volume contribution from the sources, and mixing processes (Nixon et al., 1996). Key activities influencing dissolved inorganic nutrient input from catchments include wastewater discharges (point sources), as well as urban runoff and agricultural return flow (non-point sources) (Fraterrigo and Downing, 2008; Poe et al., 2003). Over-fertilization is often a key contributor from agricultural areas, especially nitrate (NO3-N) and phosphate (PO4-P) (Casalí et al., 2008; Pearce and Schumann, 2001). The rate of nutrient loading into receiving water bodies (e.g. lakes and estuaries) also depends on a catchment's transport capacity, i.e., primarily defined by the compositing physical effect of hydrology, geology and topography (García-Garizábal and Causapé, 2006; Fraterrigo and Downing, 2008). For example, nutrient characteristics in water bodies fed by high transport capacity catchments are most influenced by catchment (or external) processes (Fraterrigo and Downing, 2008), while nutrient characteristics in water bodies fed by low transport capacity catchments tend to be more strongly influenced by internal (or in-situ) sources. Irrigation practices also affect a watershed's transport capacity by reducing or changing the hydrological patterns (García-Garizábal and Causapé, 2006).
Important in-situ sources influencing dissolved inorganic nutrient characteristics in estuaries include biological processes (e.g. primary production) and bacteriological processes (e.g. remineralisation and denitrification) (Dagg et al., 2004; Eyre, 1994; Eyre and Balls, 1999; Roy et al., 2001), where flushing rate affects the proportional influence of such processes. For example, in rapidly flushed systems nutrient characteristics are primarily a function of the concentrations in, and proportional mixing of, inflow from the external sources (e.g. Balls, 1994; Eyre, 2000). However, in systems with low flushing rates, in-situ processes such as biological uptake and bacteriological remineralisation tend to play a more significant role (Church, 1986; Lillebø et al., 2005; Poe et al., 2003). Essington and Carpenter (2000) demonstrated this in their comparative study on the effect of hydrologic flushing on nutrient cycling, contrasting smaller streams (i.e. relatively high flushing rates) with lakes (i.e. relatively low flushing rates). Emergent (e.g. Phragmites australis) and submerged (e.g. Potamogeton) aquatic vegetation can also contribute significantly to in-situ processes by taking up dissolved inorganic nutrients, but also producing large stocks of organic matter fuelling remineralisation (e.g. Gessner, 2000; Howard-Williams and Allanson, 1981). This is especially relevant in the shallow, littoral zones of lake systems (e.g. Wilderness) because these zones often support dense beds of emergent and submerged aquatic vegetation (e.g. Howard-Williams, 1980). Also relevant in terms of in-situ sources are large bird populations, especially where they aggregate in large numbers (e.g. during communal roosting, over-wintering, flocking and colonial breeding), contributing to dissolved inorganic nutrient and organic matter loading (e.g. through excretion) (e.g. Hahn et al., 2007; Hahn et al., 2008). Important anthropogenic factors that can be viewed as in-situ sources are those situated in the floodplains of water bodies such as septic tank systems from low-lying developments (e.g. Chen, 1988; Fraterrigo and Downing, 2008).
The smaller, dynamic and more linear nature of over 80% of South Africa's estuaries, situated in temperate and subtropical southern hemisphere climates (Whitfield, 1992; Van Niekerk et al., 2013), causes them to have relatively high flushing rates (or weaker retention efficiency) compared with, for example, the larger estuarine systems of the temperate northern hemisphere (e.g. North America and Western Europe) (Eyre, 2000). As a result, the nutrient characteristics of these highly dynamic systems are mostly influenced by external catchment- and tidal-scale processes, rather than in-situ processes, except during periods of extended closure (Taljaard et al., 2009; Fraterrigo and Downing, 2008). However, a small percentage of South Africa's estuaries (< 3%) comprise larger estuarine lakes mostly evolving from drowned river valleys (Hill, 1975; Whitfield, 1992). These large basin-like systems are characterised by relatively low river inflow in comparison with their volumes, and include the Wilderness, Swartvlei, St Lucia and Kosi systems (Allanson and Howard-Williams, 1984; Whitfield, 1992; Allanson, 2001). They can also close off from the sea for varying periods, but even when they are connected to the sea, tidal exchange is limited due to narrow linking channels (Whitfield, 1992). Thus, the physical properties of these systems suggest relatively low flushing rates, and the potentially stronger influence of in-situ processes on nutrient characteristics (Church, 1986; Lillebø et al., 2005; Fraterrigo and Downing, 2008).
Studies conducted on lake systems elsewhere in the world have indicated high variability in the dominant dissolved inorganic nitrogen form in these types of systems, largely depending on the dominant source of nutrient input. For example, NO3-N was found to be the common form of dissolved inorganic nitrogen entering lakes from adjacent (external) drainage basins. On entering the lake, the NO3-N is taken up by autotrophs and bacteria, and transformed into organic matter. Thus, when flushing rates decrease NH4-N (i.e. mostly generated through in-situ processes such as remineralisation of organic matter) tended to accumulate in respect to NO3-N (Quiros, 2003). Experiments conducted in the littoral zones of Canadian lake systems showed bacterial remineralisation of organic matter deposited on the surface sediment was the major in-situ source of dissolved inorganic N (NH4-N) (Levine and Schindler, 1992). However, in these lakes this process was not a major in-situ source for PO4-P, possibly indicating stronger retention of P in the sediment through, for example, adsorption, immobilization by sediment microbes, or incorporation into minerals. Thus, remineralisation provided a disproportionally higher source of in-situ dissolved inorganic N (NH4-N) compared with dissolved inorganic P (PO4-P). Studies conducted in reservoirs (also representative of water bodies with low flushing rates), in turn, suggested that increases in NH4-N concentrations (causing a decrease in NO3:NH4 ratios), stimulate cyanobacteria in these systems, having the metabolic ability to produce microcystins (Harris et al., 2016).
Detailed studies on the nutrient dynamics in estuarine lake systems in South Africa are limited. Earlier studies conducted on the phosphorus (P) dynamics in Swartvlei found that exchange of soluble P between Potamogeton beds in the littoral zone and the open waters was low (Howard-Williams and Allanson, 1981). This slow exchange was considered an important physical factor contributing to the 'closed' cycling of P in the Potamogeton beds (Howard-Williams and Allanson, 1981). A study on the Wilderness Lake systems investigated the effect of a drought and subsequent flood event on selected physico-chemical properties (e.g. salinity, dissolved oxygen, nitrate and phosphate) (Allanson and Whitfield, 1984). Most recently, Russell (1999; 2013) studied the spatio-temporal variability in salinity, temperature, pH, dissolved oxygen and turbidity in this system, but did not include nutrient dynamics.
The research presented in this paper, therefore aims to build on earlier research on the nutrient dynamics of estuarine lake systems, specifically focusing on the littoral zones of the three individual lakes (namely Eilandvlei, Langvlei and Rondevlei) that form part of the Wilderness Estuarine Lake System in South Africa. Available long-term data sets are used to characterise inter-annual and seasonal patterns in dissolved inorganic nutrient (N and P) concentrations, and to compare patterns across the three lakes. Finally, the major influencing factors on dissolved inorganic nutrient patterns - external catchment sources versus internal or in-situ processes - are explored, drawing on international learning from similar lake-type systems.
METHODS
Study area
Situated along South Africa's south coast is a series of estuarine lake systems of Pleistocene origin, connected to the sea via sinuous estuaries (Allanson and Howard-Williams, 1984). One of these is the Wilderness Lake System (33°50′ to 34°30′S; 22°33′ to 22°50′E), comprising three interconnected lakes (namely Eilandvlei, Langvlei and Rondevlei) that are joined to an estuary (the Touw Estuary) via a long, narrow channel (the Serpentine Channel) (Fig. 1).
Mean annual precipitation in the region ranges between 900 and 1 000 mm with no identifiable seasonal variation (Russell and Kraaij, 2008). Freshwater inflow to the lakes originate from the Duiwe Catchment, a small catchment (mean annual runoff [MAR] ~ 6 × 106 m3) flowing into Eilandvlei, and the Langvleispruit, an even smaller catchment (MAR ~1.5 × 106 m3) draining into Langvlei (CSIR, 1981). According to Robarts and Allanson (1977), runoff from these catchments was naturally oligotrophic (low nutrient concentrations), flowing predominantly over Table Mountain Sandstone. Large parts of the Duiwe and Langvleispruit catchments support intensive agriculture (Robarts and Allanson 1977).
The Touw Catchment (MAR ~22.6 × 106 m3), draining into the Touw Estuary, comprises mostly indigenous vegetation in its upper reaches with limited agricultural, forestry and urban development in its lower reaches (Robarts and Allanson 1977). The estuary is small and shallow, and only temporarily connected to the sea (approximately 30% of the time). Naturally, the berm breached between +3.0 and +3.5 m amsl, but since the 1800s the system has been breached at much lower levels to protect low-lying developments (Fijen, 1995). As a result, the average duration of mouth opening is shorter compared with natural, and scouring potential of sediment from the system has also been reduced (Fijen, 1995).
The hydrodynamics of the Wilderness Estuarine Lake System are complex and have been markedly modified because of premature breaching practices (Fijen, 1995). Surface flows through the Serpentine Channel (connecting the three lakes with the estuary) are dependent on numerous interrelated factors, including freshwater inflow from the Touw Catchment and tidal intrusion when the estuary mouth is open to the sea. Water levels in Langvlei and Rondevlei are much less responsive to exchange across the Serpentine Channel than Eilandvlei, because the channels between the three lakes are more constricted (i.e. elevated and narrow) compared with the Serpentine Channel (Fijen, 1995). Water exchange across the Serpentine Channel occurs mostly during the closed period when connectivity is improved as a result of higher water levels (Fijen, 1995). Because the estuary mouth is currently being breached at lower water levels, the water exchange across the Serpentine Channel, and further upstream across the channels between the lakes, has also been reduced. In addition, the water transported to the lakes via the Serpentine Channel is less saline because of a decrease in the periods that the mouth remains open to the sea. Thus, most of the existing salt content in the lakes is attributed to historical connectivity when breaching levels were higher (Fijen, 1995). A reverse gradient in salt content now exists where the lakes furthest removed from the estuary, namely Langvlei and Rondevlei, generally having higher salinity compared to Eilandvlei, the lake closest to the estuary (Allanson and Whitfield, 1983; Russell, 2009; Russell, 2013). Further, direct freshwater inflow into Eilandvlei (via the Duiwe catchment) and Langvlei (via the Langvleispruit), and the absence of surface runoff into Rondevlei, also contribute to this reverse salinity gradient (Fijen, 1995).
The littoral zones of the three lakes are relatively narrow (between +2 and 0 m MSL), bordering deeper pelagic zones. Maximum bathymetry of the pelagic zones in both Eilandvlei and Langvlei is about −4 m amsl, but less in Langvlei at about −2 m amsl (CSIR, 1982). Although surface water areas vary over time, the average areas for Eilandvlei, Langvlei and Rondevlei are estimated at 140, 200 and 100 ha, respectively (L van Niekerk, CSIR, personal observation). These littoral zones support dense beds of submerged macrophytes generally up to a water depth of 3.0 m (Allanson and Whitfield, 1983). The littoral zones of all three lakes are fringed by a narrow margin of emergent aquatic macrophytes including Phragmites australis, Typha capensis and Schoenoplectus scirpoideus (Russell, 2003, Russell et al., 2014). Large numbers of water birds, especially herbivorous and graminivorous Anatidae species, together with the herbivorous Red-knobbed Coot, are associated with this dense aquatic vegetation (Russell et al., 2009; Russell et al., 2014).
Data sources and analysis
Long-term water chemistry data were obtained from South Africa's national department responsible for water (currently, the Department of Water and Sanitation; www.dwaf.gov.za/iwqs/wms/data/WMS_pri_txt.asp). The Department runs a long-term monitoring programme on the country's rivers, dams and lakes, and for the purposes of this study available data on the electrical conductivity (EC) and dissolved inorganic nutrient data in the Duiwe River and three lakes were extracted. The most downstream station on the Duiwe River [K3H11] was used as it is most representative of inflow to the lakes. No long-term data were available for inflow from the Langvleispruit into Langvlei, or for EC and dissolved inorganic nutrient fluxes across the Serpentine Channel into Eilandvlei. Stations in the three lakes were all located in the littoral zones (Eilandvlei [K3R5], Langvlei [K3R4] and Rondevlei [K3R3]) (Fig. 1). The spatial uniformity previously observed in biogeochemical parameters in the lakes (e.g. Russell, 2013) warranted the use of the single location measurements for the lakes. However, Russell (2013) did occasionally observe spatial variations in some biogeochemical conditions (e.g. dissolved oxygen) between the shallower littoral zones and the deeper pelagic zones in the lakes, where oxygen in the littoral zone was comparatively higher due to its association with submerged macrophyte beds. Submerged aquatic plants are known to also influence nutrient dynamics in estuaries (Taljaard et al., 2009). Therefore, while the dissolved inorganic nutrient data collected from these stations in the littoral zones can be considered representative of those zones, such data may not necessarily represent conditions in the deeper, pelagic zones where submerged macrophyte beds are usually absent, mainly due to water depth.
Data on total dissolved inorganic oxidised nitrogen (NOx-N) concentrations (reported as NO2- + NO3- in µg·L−1 N, detection limit 25 µg·L−1 N ), total dissolved ammonia (NH4-N) concentrations (reported as NH4+ in µg·L−1 N, detection limit 25 µg·L−1 N) and dissolved inorganic phosphate (PO4-P) concentrations (reported as PO43- µg·L−1 P, detection limit 5 µg·L−1 P) were extracted for all stations over the period 1985-2015 (for the Duiwe River data was only available from the period 1998-2015). Sampling frequency was mostly on a monthly basis, but where multiple samples were collected in a given month, an average value was calculated for the month to provide time-series data at a monthly resolution. To compare long-term inter-annual trends, average annual concentrations over the study period were calculated for each location, while seasonal trends were compared using median monthly concentrations. Analyses of dissolved inorganic nutrient samples were conducted by the Department's analytical laboratories using the photometric methods as described in the methods manual of the Resource Quality Services department (DWA, 2009).
No reliable long-term flow data for the Duiwe and Langvleispruit rivers, or volume fluxes across the Serpentine Channel, were available for this study. Long-term precipitation from a station near Rondevlei (Station K3E3, coordinates: 22.72033 E, −33.99177 S), therefore, was considered as a proxy for river runoff (Fig. 1).
Data were analysed using Dell Statistica Version 13 (Dell Inc., 2016). The data were tested for normality using the Shapiro-Wilks test. Relationships between monthly median values for the various parameters were analysed using either the parametric correlation coefficient or the non-parametric Spearman's rank correlation depending on the normality of the data. Lastly, long-term inter-annual trends were statistically analysed for each system using linear regressions. All analyses were done at α = 0.05.
RESULTS AND DISCUSSION
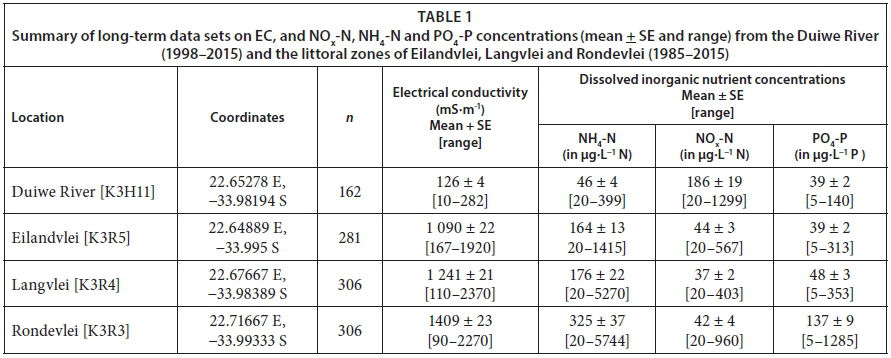
Table 1 provides a summary of the long-term data sets on EC, NOx-N, NH4-N and PO4-P (mean ± SE; range) from the Duiwe River (draining into Eilandvlei), as well as the littoral zones of Eilandvlei, Langvlei and Rondevlei.
Comparing the data from the Duiwe River with that of the three lakes, average EC levels in the river were an order of magnitude lower compared with levels in the lakes (Table 1). The higher EC levels in the lakes are indicative of connectivity to the sea via the Serpentine Channel (Russell 2009; 2013). While average NH4-N concentrations in the Duiwe River were lower compared with concentrations in the lakes, average NOx-N concentrations showed the contrary, i.e., concentrations in the river were higher compared with the three lakes. Average PO4-P concentrations in the river were comparable with those in Eilandvlei and Langvlei, but concentrations in Rondevlei were higher compared with those in the river and in the other two lakes.
Across-lake comparisons show that average EC levels in the littoral zones of the three lakes increased incrementally moving from Eilandvlei to Langvlei and into Rondevlei, echoing the reverse salinity gradient explained in previous studies (e.g. Allanson and Whitfield, 1983; Russell, 2009; Russell, 2013). Eilandvlei and Rondevlei displayed similar average NH4-N levels (164 and 176 µg·L−1 N, respectively) but in Rondevlei concentrations were double (324 µg·L−1 N). This pattern was also evident in average PO4-P concentrations, where levels in Eilandvlei and Rondevlei were similar (39 and 48 µg·L−1 N, respectively), with concentrations in Rondevlei being much higher (137 µg·L−1 N). A similar pattern emerged in average NOx-N concentrations, but was not as profound as for NH4-N and PO4-P.
Building on this general overview of the historical data set, the more detailed analysis of the inter-annual and seasonal patterns in and across the three individual lakes follows. Major influencing factors either derived from the external catchment sources or in-situ processes (e.g. remineralisation) are postulated drawing on other studies.
Inter-annual variability
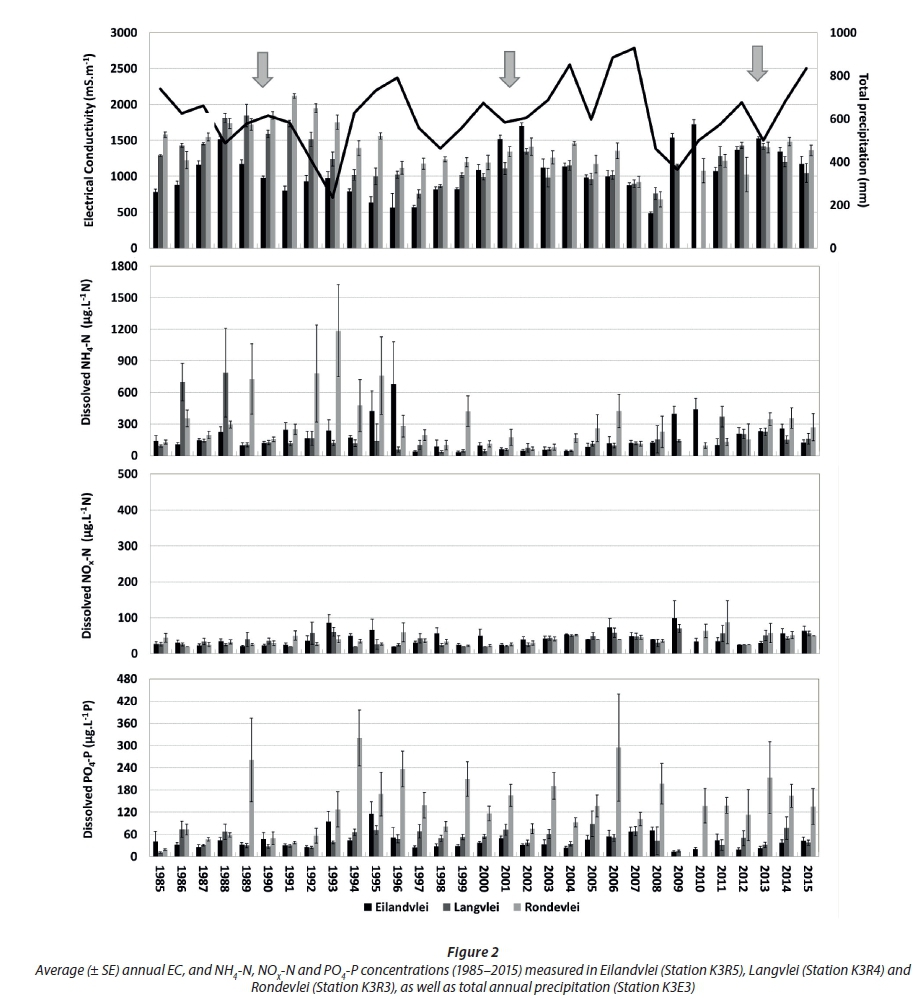
Average inter-annual NOx-N concentrations remained relatively low across the littoral zones in all three lakes during the period 1985 to 2015, mostly near detection limit (25 µg·L−1 N) (Fig. 2).
Inter-annual variations in both NH4-N and PO4-P concentrations varied greatly in, and across, the three lakes. Average annual concentrations of these did not show any consistent trends over the study period (P > 0.05). Although, during the period 1997 to 2004, NH4-N concentrations in all three lakes were more stable and lower, often near detection limit (25 µg·L−1 N). Interestingly, average annual electrical conductivity (EC) levels in the littoral zones of all three lakes revealed inter-annual oscillations, peaking approximately every 10 years (as indicated by the arrows in Fig. 2), but did not match any of the dissolved inorganic nutrient distribution patterns. The relative similarity in inter-annual oscillation was strongest between the adjacent lakes Rondevlei and Langvlei (n = 29; r = 0.72; P < 0.05) and, to a lesser extent, between adjacent lakes Langvlei and Eilandvlei (n = 30; r = 0.43; P < 0.05), but did not match between the two end lakes (i.e. Eilandvlei and Rondevlei ~ P > 0.05). These inter-annual oscillations could not be explained by total annual precipitation (Fig. 3), which suggested a much more complex interaction between various influencing factors such as catchment runoff, precipitation, evaporation and exchange across the Serpentine Channel. Large-scale atmospheric processes such as the El Niño and the Southern Oscillation tend to influence the meteorological conditions in South Africa in reoccurring cycles of 9-13, 13-18 and 18-39 years (Reason and Rouault, 2002). This frequency tends to coincide with the 10-year cycle apparent in the inter-annual oscillation in EC, but no conclusive correlations can be assumed without further, more detailed analyses.
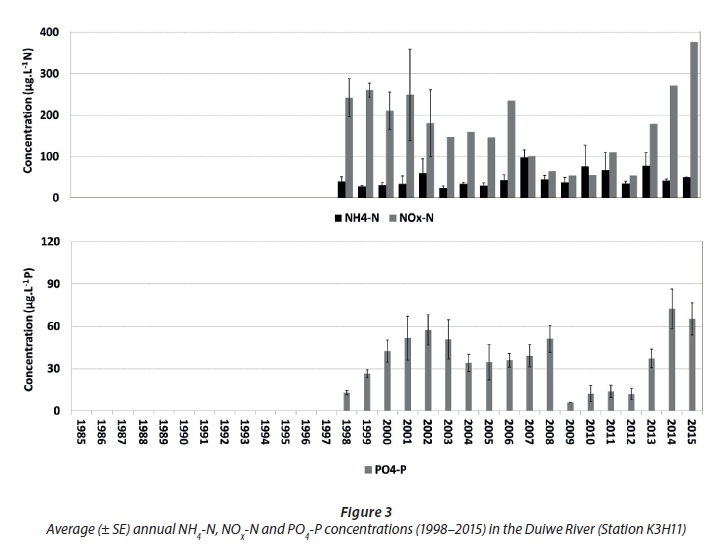
To establish the potential influence from external catchment sources, dissolved inorganic nutrient data from the Duiwe River were analysed (Fig. 3). The Duiwe River was naturally oligotrophic (Robarts and Allanson, 1977), so extensive agriculture in the catchment (e.g. over-fertilization) is considered the most likely source of elevated PO4-P, and especially NOx-N in river inflow (e.g. García-Garizábal and Causapé, 2006). Inter-annual patterns in PO4-P displayed a positive correlation with NOx-N (n = 18; r = 0.57; P < 0.05), suggesting some proportionality in the contribution from anthropogenic sources in the catchment.
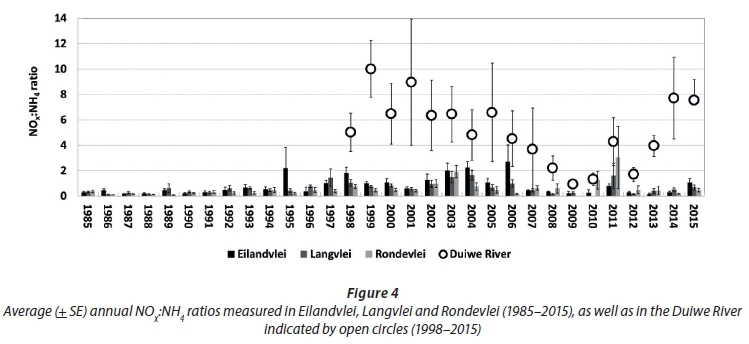
Comparing the average annual dissolved inorganic nutrient patterns of the three lakes (Fig. 2) with that of the Duiwe River (Fig. 3), there are no clear inter-annual relationships, not even in Eilandvlei (into which the Duiwe River drains). This suggests that external catchment fluxes may not be the dominant, direct drivers of average annual dissolved inorganic nutrient patterns in the lake littoral zones. This is expected, considering the relatively small catchment sizes of the rivers in comparison to the large size of the lakes into which they drain. However, an earlier study did show that a large flood event introduced a significant pulse of dissolved inorganic nutrients (i.e. NOx-N and PO4-P) into the lakes (Allanson and Whitfield, 1984), although the observed effect on lake nutrient levels was short-lived, possibly as a result of rapid uptake by aquatic plants (Quiros, 2003). The temporal scale of the long-term data sets (e.g. monthly data average over a year) therefore, may not be refined enough to record these types of pulsed events, and thus may under-represent the influence of the catchment (i.e. external sources) on nutrient processes in the lakes. Secondary processes, for example, where seasonal catchment fluxes are taken up by the fringing, aquatic vegetation which then enters the organic matter pool in the littoral zone to fuel remineralisation, may also be relevant. The non-correlation between river and lake dissolved inorganic nutrient patterns are reflected in their different average annual NOx:NH4 ratios (Fig. 4). For the Duiwe River, ratios were mostly above 1 (i.e. dissolved inorganic N is dominated by NOx-N) while those in the lakes were mostly below 1 (i.e. dissolved inorganic N is dominated by NH4-N). The ratios in the Duiwe River were primarily determined by variation in NO3-N concentrations (Fig. 3), peaking between the late 1990s to early 2000s (8-10), dropping to the lowest ratio in 2009 (~1), but then increasing again towards 2015 (7.5). However, ratios in the lakes were mostly influenced by variation in NH4-N concentrations, where the drop in concentrations between 1997 to 2004 (Fig. 2) resulted in highest NOx:NH4 ratios (> 1) in the lakes.
Although not evident in average annual patterns, external catchment fluxes may well affect dissolved inorganic nutrient patterns in the lakes, for example, during flood events or through secondary mechanisms, as such fuelling the growth of fringing vegetation which then enters the littoral zone's organic matter pool (in turn fuelling in-situ remineralisation). Thus, the dissolved inorganic nutrient dynamics of these lakes are more complex, involving both external catchment sources (Fraterrigo and Downing, 2008; Poe et al., 2003; Casalí et al., 2008), but also in-situ processes (e.g. primary production, decomposition, remineralisation and excretion from large bird populations) (Dagg et al., 2004; Eyre, 1994; Eyre and Balls, 1999; Lemley et al., 2014; Roy et al., 2001), possible dissolved inorganic nutrient fluxes across the Serpentine Channel (e.g. influenced by connectivity to the sea), as well as potential anthropogenic sources within the floodplain (e.g. sewage seepage from low-lying development). The relatively low NOx:NH4 ratios observed in the lakes support the stronger influence of in-situ processes in the littoral zones, where ratios below 1 indicate NH4-N accumulation (mostly generated through in-situ bacteriological processes) in respect to NO3-N (i.e. mostly introduced from external sources), as was observed in other lake type systems with low flushing rates (Quiros, 2003).
Seasonal trends
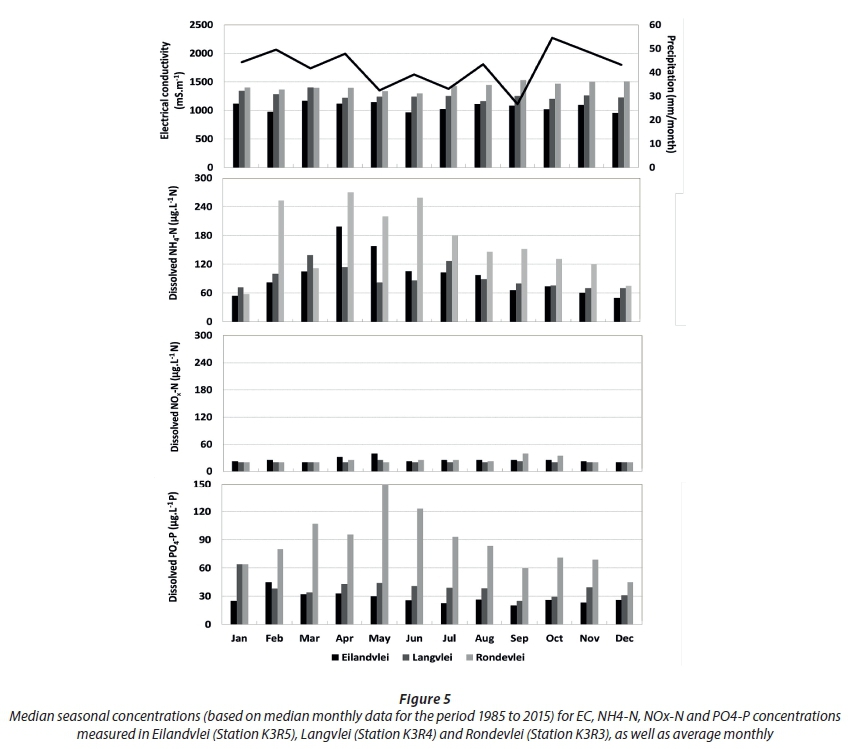
Across all lakes, seasonal NOx-N concentrations in the littoral zones also remained low across all seasons, mostly near detection limit (25 µg·L−1 N) (Fig. 5).
NH4-N concentrations showed a tendency to peak over the period May to July (late-autumn into winter), especially in Rondevlei. In Rondevlei, this pattern was also evident for PO4-P, but not in the other two lakes where PO4-P concentrations remained uniform throughout the year. In fact, the seasonal patterns in Rondevlei between NH4-N and PO4-P showed a positive correlation (n = 12; r = −0.61; P < 0.05), indicating that the in-situ source in Rondevlei contributed relatively proportionally to NH4-N and PO4-P concentrations in the littoral zone, in contrast to the sources in Eilandvlei and Langvlei (where seasonal increases in NH4-N were disproportional to PO4-P which remained relatively uniform). The disproportionality between NH4-N and PO4-P in Eilandvlei and Langvlei reflect similar patterns observed in the littoral zones of Canadian lake systems, where bacterial remineralisation of organic matter (deposited on the surface sediment) was the major in-situ source of dissolved inorganic NH4-N, but not for PO4-P because the latter was more effectively retained at the sediment surface by other processes (Levine and Schindler, 1992). In the case of Rondevlei there is no significant relationship between seasonal precipitation (Fig. 5) and NH4-N and PO4-P that would suggest external fluxes from adjacent catchments or rainfall as the source. An alternative explanation is that the sampling location in Rondevlei may be influenced by different in-situ sources, such as a sewage disposal system of a low-lying development or a large bird roosting area, both known to occur here. These sources are known to increase N and P in water bodies (e.g. Chen, 1988; Hahn et al., 2007; Hahn et al., 2008), releasing dissolved inorganic nutrients directly into the water column which could explain the relative proportionality of PO4-P and NH4-N at this location. However, this hypothesis needs to be confirmed through a more thorough sampling programme.
Seasonality in dissolved inorganic nutrient patterns of the Duiwe River was also analysed (Fig. 6). Seasonal NH4-N concentrations in the river remained relatively low with no apparent seasonal patterns. Although both NOx-N and PO4-P concentrations increased during autumn (March/April), only NOx-N continued to increase into winter, peaking in late winter (August). This is most likely indicative of the different ways in which N and P is mobilised from or retained in catchment soils (e.g. Levine and Schindler, 1992). Again, the non-correlation between seasonal dissolved inorganic nutrient concentrations in the Duiwe River (Fig. 6) and precipitation (Fig. 5) must be interpreted with caution as precipitation may not be a suitable proxy for river flow rates in this particular catchment as many other factors could influence transport capacity (e.g. García Garizábal and Causapé, 2006; Fraterrigo and Downing, 2008).

Reflecting on the seasonal patterns in dissolved inorganic nutrients of the three lakes (Fig. 5) and that of the Duiwe River (Fig. 6), no clear, comparable seasonal trends are evident. Seasonal variation in NOx:NH4 ratios of the Duiwe River and three lakes also reflects this disconnect (Fig. 7). NOx:NH4 ratios in the Duiwe River were highest during autumn/winter, matching the period of elevated NOx-N concentrations in river inflow (Fig. 6). In the lakes, the lowest NOx:NH4 ratios were evident during this period (winter) because NH4 concentrations peaked (Fig. 5), especially in Rondevlei where ratios decreased to ~0.1.

Seasonal results support observed patterns in the inter-annual analyses in that external catchment fluxes are most likely not the sole, or even the dominant, driver of seasonal dissolved inorganic nutrient patterns in the littoral zones of the lakes, with findings rather suggesting in-situ processes (e.g. remineralisation) as most significant, although influences from catchment fluxes during flood events, or through secondary mechanisms (e.g. fuelling growth of fringing aquatic vegetation which enters the littoral zone's organic matter pool, in turn fuelling remineralisation), cannot be ignored.
CONCLUSION
This study expands research on dissolved inorganic nutrient dynamics in the littoral zones of estuarine lake systems, and potential influencing factors, using the Wilderness Lake System as a case study. NH4-N concentrations in (external) catchment inflow (e.g. Duiwe River) remained relatively low. Inter-annual patterns of PO4-P in river inflow displayed a positive relationship with NOx-N concentrations, suggesting some proportionality in the contribution from sources in the catchment. The relatively high NOx:NH4 ratios confirmed the dominant influence of NO3 enrichment from agricultural activities (e.g. over-fertilization) (Fraterrigo and Downing, 2008; Casalí et al., 2008). In all three lakes NO3-N concentrations remained low, while inter-annual variations in both NH4-N and PO4 varied greatly in and across the lakes, with no clear inter-annual trends. Relatively low NOx:NH4 ratios (mostly less than 1) in the lakes (especially in Eilandvlei and Langvlei) indicated a stronger influence of in-situ bacteriological processes, similar to observations in other lake type systems with low flushing rates (Quiros, 2003).
The positive correlation in Rondevlei between NH4-N and PO4-P was indicative of a relatively proportional input from source/s in this lake compared with input from source/s in Eilandvlei and Langvlei (where seasonal increases in NH4-N were disproportional to PO4-P). The disproportionality between NH4-N and PO4-P in Eilandvlei and Langvlei reflect similar patterns observed in the littoral zones of Canadian lake systems, where in-situ bacterial remineralisation of organic matter was found to be a major source of dissolved inorganic NH4, but not for PO4-P because the latter was more effectively retained at the sediment surface by other processes (Levine and Schindler, 1992). The situation in Rondevlei, however, speaks to the contribution of potentially different in-situ sources such as a sewage disposal system of the low-lying development or large bird roosting areas.
Comparison of inter-annual and seasonal trends in dissolved inorganic nutrients (NOx-N, NH4-N and PO4-P) in river inflow and the three lakes indicates that external catchment fluxes are most likely not the sole, or even the dominant, driver of average long-term or seasonal dissolved inorganic nutrient patterns in the lake littoral zones. This can be expected in a way, considering the relatively small catchment sizes of the rivers in comparison to the large size of the lakes into which they drain. However, the influence of catchment fluxes during flood events (Allanson and Whitfield, 1984), or through secondary mechanisms where catchment-derived dissolved inorganic nutrients are taken up by the fringing, aquatic vegetation and then enter the organic pool in the littoral zone to fuel remineralisation (Gessner, 2000; Howard-Williams and Allanson, 1981), can still be significant.
Understanding these more subtle influences of external catchment fluxes (e.g., through flood events or a secondary mechanism of fuelling growth of fringing vegetation) is critical, especially in aquatic ecosystems characterised by weak flushing potential, such as lakes. As a result of weak flushing these systems tend to accumulate organic matter. Over time a situation may arise where dissolved inorganic nutrients and organic matter that incrementally enter the lakes (e.g. during flood events, or through continuous die-back of vegetation), often aggravated by inappropriate management of catchment activities (e.g. over-fertilisation or wastewater disposal), exceed the system's assimilative ability. The consequence, especially in weakly flushed systems, is an increase in-situ remineralisation rates, causing elevated NH4-N concentrations and a marked decrease in NOx:NH4 ratios. Low NO3:NH4 ratios are known to stimulate cyanobacteria, having the metabolic ability to potentially produce microcystins (e.g. Harris et al., 2016). Already the Wilderness Estuarine Lake System has experienced sporadic nuisance algal blooms (Russell, 2013), at times accompanied by mortality in fish (e.g. Russell, 1994). Preliminary investigations on Lake Sibaya and Kosi Bay - large lake systems along South Africa's east coast - also showed signs of relatively low NOx:NH4 ratios similar to that observed in the Wilderness (Taljaard, CSIR, unpublished data). Therefore, further studies on these estuarine lake systems with low flushing potential are required to better understand and quantify the subtle and complex processes influencing their nutrient dynamics. This is necessary to inform water resource management, both in the catchments and within the floodplains, so that incremental enrichment ultimately does not exceed critical limits causing them to tip into eutrophic, possibly toxic, aquatic systems in the future that will be difficult, if not impossible, to reverse.
ACKNOWLEDGEMENTS
The authors thank the Department of Water and Sanitation for access to their long-term water quality datasets and Dr Ian Russell (South African National Parks) for his support and sharing of valuable data, information and knowledge on the Wilderness Lake System. Also thanks to Prof Janine Adams (Nelson Mandela University) for her support throughout this study. This project was financially supported by the Council for Scientific and Industrial Research (CSIR), South Africa.
REFERENCES
ALLANSON BR and WHITFIELD AK (1983) The Touw River Floodplain. Part I: Ecological structure in relation to planning and management. A report prepared by the Institute for Freshwater Studies, Rhodes University, in collaboration with the Cooperative Scientific Programmes, Inland Water Ecosystems, of the Council for Scientific and Industrial Research. South African National Scientific Programmes Report No. 79. CSIR, Pretoria. [ Links ]
CASALÍ J, GASTESI R, ÁLVAREZ-MOZOS J, DE SANTISTEBAN LM, DEL VALLE DE LERSUNDI J, GIMÉNEZ R, LARRAÑAGA A, GOÑI M, AGIRRE U, CAMPO MA, LÓPEZ JJ and DONÉZAR M (2008) Runoff, erosion, and water quality of agricultural watersheds in central Navarre (Spain). Agric. Water Manage. 95 1111-1128. https://doi.org/10.1016/j.agwat.2008.06.013 [ Links ]
CHEN M (1988) Pollution of ground water by nutrients and fecal coliforms from lakeshore septic tank systems. Water Air Soil Pollut. 37 407-417. https://doi.org/10.1007/BF00192950 [ Links ]
CORBELLA S and STRETCH DD (2014) Directional wave spectra on the east coast of South Africa. J. S. Afr. Inst. Civ. Eng. 56 (3) 53-64. [ Links ]
CSIR (1981) Evaluation of prototype data and the application of a numerical model to the Wilderness Lakes and Touw River flood plain. CSIR Report No. C/SEA 8113. CSIR, Stellenbosch, South Africa. [ Links ]
CSIR (1982) Wilderness Report No. 2. Evaluation of prototype data and the application of a numerical model to conditions when the estuary mouth was opened. CSIR Report No. C/SEA 8255. CSIR, Stellenbosch, South Africa. [ Links ]
DAGG M, BENNER R, LOHRENZ S and LAWRENCE D (2004) Transformation of dissolved and particulate materials on continental shelves influenced by large rivers: plume processes. Cont. Shelf Res. 24 833-858. https://doi.org/10.1016/j.csr.2004.02.003 [ Links ]
DWA (Department of Water Affairs, South Africa) (2009) Resource Quality Services methods manual: Inorganic chemistry laboratory. Method 5001.5, 5001.7, and 5001.11. Dated 21 December 2009. DWA, Pretoria. [ Links ]
DWS (Department of Water and Sanitation, South Africa) (2014) Reserve determination studies for surface water, groundwater, estuaries and wetlands in the Gouritz Water Management Area: Estuaries RDM Report - Rapid Assessment, Volume 2 (Wilderness System). Prepared by the Council for Scientific and Industrial Research (CSIR) for Scherman Colloty and Associates cc. Report no. RDM/WMA16/04/CON/0713, Volume 2. DWS, Pretoria. [ Links ]
EYRE B (1994) Nutrient biogeochemistry in the tropical Moresby River Estuary system, North Queensland, Australia. Estuar. Coast. Shelf Sci. 39 15-31. https://doi.org/10.1006/ecss.1994.1046 [ Links ]
EYRE B and BALLS P (1999) A comparative study of nutrient behaviour along the salinity gradient of tropical and temperate estuaries. Estuaries 22 (2A) 313-326. https://doi.org/10.2307/1352987 [ Links ]
FRATERRIGO JM and DOWNING JA (2008) The influence of land use on lake nutrients varies with watershed transport capacity. Ecosystems 11 1021-1034. https://doi.org/10.1007/s10021-008-9176-6 [ Links ]
FIJEN APM (1995) Wilderness Lakes catchment, Touw and Duiwe Rivers, water management strategy, Vol. 2: Water resources. Department of Water Affairs and Forestry, Report No. WQ. K100/00/0395. DWAF, Pretoria. 57 pp. [ Links ]
FINDLAY S, GROFFMAN P and DYE S (2003) Effects of Phragmites australis removal on marsh nutrient cycling. Wetlands Ecol. Manage. 11 157-165. https://doi.org/10.1023/A:1024255827418 [ Links ]
GARCÍA-GARIZÁBAL I and CAUSAPÉ J (2010) Influence of irrigation water management on the quantity and quality of irrigation return flows. J. Hydrol. 385 36-43. https://doi.org/10.1016/j.jhydrol.2010.02.002 [ Links ]
GESSNER MO (2000) Breakdown and nutrient dynamics of submerged Phragmites shoots in the littoral zone of a temperate hardwater lake. Aquat. Bot. 66 9-20. https://doi.org/10.1016/S0304-3770(99)00022-4 [ Links ]
HAHN S, BAUER S and KLAASSEN M (2007) Estimating the contribution of carnivorous waterbirds to nutrient loading in freshwater habitats. Freshwater Biol. 52 2421-2433. https://doi.org/10.1111/j.1365-2427.2007.01838.x [ Links ]
HAHN S, BAUER S and KLAASSEN M (2008) Quantification of allochthonous nutrient input into freshwater bodies by herbivorous waterbirds. Freshwater Biol. 53 181-193. [ Links ]
HARRIS TD, SMITH, VH, GRAHAM JL, VAN DE WAAL DB, TEDESCO LP and CLERCIN N (2016) Combined effects of nitrogen to phosphorus and nitrate to ammonia ratios on cyanobacterial metabolite concentrations in eutrophic Midwestern USA reservoirs. Inland Waters 6 199-210. https://doi.org/10.5268/IW-6.2.938 [ Links ]
HENAO J and BAANANTE C (1999) Nutrient depletion in the agricultural soils of Africa. 2020 Brief No. 62. URL: www.ifpri.org/2020/briefs/number62.htm (Accessed 26 October 2016). [ Links ]
HOWARD-WILLIAMS C (1980) Aquatic macrophyte cummuniteis of the Wilderness Lakes: Community structure and associated environmental conditions. J. Limnol. Soc. S. Afr. 6 (2) 85-92. [ Links ]
HOWARD-WILLIAMS C and ALLANSON BR (1981) Phosphorus cycling in dense Potamageton pectinatus L. beds. Oecologia 49 56-66. https://doi.org/10.1007/BF00376898 [ Links ]
LEMLEY DA, SNOW GC and HUMAN LRD (2014) The decomposition of estuarine macrophytes under different temperature regimes. Water SA 40 (1) 117-124. https://doi.org/10.4314/wsa.v40i1.14 [ Links ]
LEVINE SN and SCHINDLER DW (1992) Modification of the N:P ratio in lakes by in situ processes. Limnol. Oceanogr. 37 (5) 917-935. https://doi.org/10.4319/lo.1992.37.5.0917 [ Links ]
NIJBURG JW and LAANBROEK HJ (1997) The fate of N-nitrate in healthy and declining Phragmites australis stands. Microb. Ecol. 34 254-262. https://doi.org/10.1007/s002489900055 [ Links ]
NIXON SW, AMMERMAN JW, ATKINSON LP, BEROUNSKY VM, BILLEN G, BOICOURT WC, BOYTON WR, CHURCH TM, DITORO DM, ELMGREN R, GARBER JH, GIBLIN AE, JAHNKE RA, OWENS NJP, PILSON MEQ and SEITZINGER SP (1996) The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean. Biogeochemistry 35 141-180. https://doi.org/10.1007/BF02179826 [ Links ]
PEARCE MW and SCHUMANN EH (2001) The impact of irrigation return flow on aspects of the water quality of the upper Gamtoos Estuary, South Africa. Water SA 27 (3) 367-372. https://doi.org/10.4314/wsa.v27i3.4981 [ Links ]
POE AC, PIEHLER MF, THOMPSON SP and PAERL HW (2003) Denitrification in a constructed wetland receiving agricultural runoff. Wetlands 23 (4) 817-826. https://doi.org/10.1672/0277-5212(2003)023[0817:DIACWR]2.0.CO;2 [ Links ]
REASON CJC and ROUAULT M (2002) ENSO-like decadal variability and South African precipitation, Geophys. Res. Lett. 29 (13) doi:10.1029/2002GL014663. https://doi.org/10.1029/2002GL014663 [ Links ]
ROBARTS RD and ALLANSON BR (1977) Meromixis in the lake-like upper reaches of a South African estuary. Arch. Hydrobiol. 80 531-540. [ Links ]
ROY PS, WILLIAMS RJ, JONES AR, YASSINI I, GIBBS PJ, COATES B, WEST RJ, SCANES PR, HUDSON JP and NICHOL S (2001) Structure and function of south-east Australian estuaries. Estuar. Coast. Shelf Sci. 53 351-384. https://doi.org/10.1006/ecss.2001.0796 [ Links ]
RUSSELL IA (1994) Mass mortality of marine and estuarine fish in Swartvlei and Wilderness lake systems, southern Cape. S. Afr. J. Aquat. Sci. 20 (1/2) 93-96. https://doi.org/10.1080/10183469.1994.9631353 [ Links ]
RUSSELL IA (1999) Changes in the water quality of the Wilderness and Swartvlei lake systems, South Africa. Koedoe 42 57-72. https://doi.org/10.4102/koedoe.v42i1.222 [ Links ]
RUSSELL IA (2003) Long-term changes in the distribution of emergent aquatic plants in a brackish South African estuarine-lake system. Afr. J. Aquat. Sci. 28 103-122. https://doi.org/10.2989/16085910309503776 [ Links ]
RUSSELL IA (2013) Spatio-temporal variability of surface water quality parameters in a South African estuarine lake system. Afr. J. Aquat. Sci. 38 (1) 53-66. https://doi.org/10.2989/16085914.2012.742006 [ Links ]
RUSSELL IA and KRAAIJ T (2008) Effects of cutting Phragmites australis along an inundation gradient, with implications for managing reed encroachment in a South African estuarine lake system. Wetlands Ecol. Manage. 16 383-393. https://doi.org/10.1007/s11273-007-9075-1 [ Links ]
RUSSELL IA, RANDALL RM and HANEKOM N (2014) Spatial and temporal patterns of waterbird assemblages in the Wilderness Lakes complex, South Africa. Waterbirds 37 (1) 1-18. https://doi.org/10.1675/063.037.0104 [ Links ]
RUSSELL IA, RANDALL RM, RANDALL BM and HANEKOM N (2009) Relationships between the biomass of waterfowl and submerged macrophytes in a South African estuarine lake system. Ostrich 80 (1) 35-41. https://doi.org/10.2989/OSTRICH.2009.80.1.5.763 [ Links ]
TALJAARD S, VAN NIEKERK L and JOUBERT W (2009) Extension of a qualitative model on nutrient cycling and transformation to include microtidal estuaries on wave-dominated coasts: Southern hemisphere perspective. Estuar. Coast. Shelf Sci. 85 407-421. https://doi.org/10.1016/j.ecss.2009.09.006 [ Links ]
TSAHAR E, MARTÍNEZ DEL RIO C, IZHAKI I and ARAD Z (2005) Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores. J. Exp. Biol. 208 1025-1034. https://doi.org/10.1242/jeb.01495 [ Links ]
VAN DER PUTTEN W (1997) Die-back of Phragmites australis in European wetlands: an overview of the European Research Programme on reed die-back and progression (1993-1994). Aquat. Bot. 59 (3/4) 263-275. https://doi.org/10.1016/S0304-3770(97)00060-0 [ Links ]
WEISSER PJ, WHITFIELD AK and HALL CM (1992) The recovery and dynamics of submerged aquatic macrophyte vegetation in the Wilderness lakes, Southern Cape. Bothalia 22 (2) 283-288. https://doi.org/10.4102/abc.v22i2.851 [ Links ]
WHITFIELD AK (1984) The effects of prolonged aquatic macrophyte senescence on the biology of the dominant fish species in a southern African coastal lake. Estuar. Coast. Shelf Sci. 18 315-329. https://doi.org/10.1016/0272-7714(84)90074-X [ Links ]
Received 14 December 2015
Accepted in revised form 5 December 2017
* To whom all correspondence should be addressed. +27218882494; fax: +2721882693; e-mail: staljaar@csir.co.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}