










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.42 n.4 Pretoria Oct. 2016
http://dx.doi.org/10.4314/wsa.v42i4.06
Testing household disinfectants for the inactivation of helminth eggs on surfaces and in spills during pit latrine emptying
D NaidooI; C ArcherI, *; B LoutonII; N RoddaI
ISchool of Life Sciences, University of KwaZulu-Natal (Westville Campus), Private Bag X54001, Durban 4000
IIPartners in Development, P.O. Box 11431, Dorpspruit, Pietermaritzburg, 3206
ABSTRACT
The lack of potable water, poor hygiene practices and inefficient sanitation systems in developing countries pose a risk to human health. Pit latrines aim to break the link between human faecal waste and human contact by serving as improved sanitation. The eThekwini Municipality has developed a programme that employs workers to empty pits every 3 to 5 years. This study aimed to find a cost-effective, easy way of disinfecting household surfaces and inactivating Ascaris eggs using household disinfectants, so that transmission of helminths is minimised when pits are emptied. Disinfectants tested in this study were based on sodium hypochlorite or carbolic acid. Experiments simulated inactivation of Ascaris eggs by wiping contaminated surfaces with disinfectants and by soaking small spills in disinfectants. Samples were processed using standard helminth egg enumeration methods for soil and sludge. Wiping of contaminated surfaces removed Ascaris eggs, facilitating egg transfer but not egg inactivation. Prolonged exposure to disinfectants, at concentrations of 50% and above, were required to inactivate eggs. Sodium hypochlorite-based disinfectants were the most successful for inactivation, whilst those based on carbolic acid were mostly ineffective. It is recommended that faecal sludge spills are soaked in a sodium hypochlorite-based disinfectant and contaminated surfaces be wiped with a sodium hypochlorite-based disinfectant-saturated cloth, which should then be soaked for 1 h in a similar disinfectant solution (50% dilution) to inactivate any eggs picked up on the cloth.
Keywords: Ascaris, carbolic acid, disinfectant, eggs, inactivation, pit latrine, sanitation, sodium hypochlorite
INTRODUCTION
The lack of improved water, sanitation and hygiene (WASH) infrastructure in developing countries poses significant health risks. An estimated 2.5 billion people worldwide do not have access to appropriate sanitation facilities, most living in sub-Saharan Africa, which has the poorest record for implementing WASH interventions, particularly sanitation (Heijnen et al., 2014). In 2009 it was estimated that 88% of worldwide diar-rhoeal deaths in children under 5 years was due to the lack of WASH facilities (UNICEF/WHO, 2009). While considerable progress has been made in the provision of improved sanitation in South Africa in recent decades, many informal settlements and rural areas still experience problems (Johannessen t al., 2014).
Diarrhoeal diseases are generally caused by bacteria and viruses, but can also be symptomatic of geohelminth infections (Brownell and Nelson, 2006). Ascariasis, the most widespread helminth infection in humans, is caused by the common roundworm, A. lumbricoides (Brownell and Nelson, 2006). It is prevalent in tropical and sub-tropical regions, infecting approximately one third of the world's population (Bethony et al., 2006). Although ascariasis is generally not lethal, it is a major contributor to impairment of physical and cognitive development, particularly in children < 5 years, in whom the sequelae may be life-long (Esrey et al., 1991). Symptoms associated with severe ascariasis include abdominal pain and distension due to intestinal blockage by the worms. Complications of ascariasis interfere with schooling through absenteeism and impaired development (Awasthi et al., 2003). Infected subjects may become immuno-compromised, increasing infection susceptibility, including multiple parasitic worm infestations (Bethony et al., 2006).
Ascaris lumbricoides is the most resilient of all helminths in terms of resistance of excreted fertilised eggs to inactivation. Since A. lumbricoides cannot be distinguished microscopically from A. suum (which infects pigs), only the genus will be specified here, unless referring specifically to the life cycle in humans. Ascaris eggs are resistant to desiccation, and are able to survive in both aerobic and anaerobic conditions (Pecson and Nelson, 2005). Consequently, Ascaris is used as an index organism for overall pathogen inactivation in faecal sludge.
Fertilised Ascaris lumbricoides eggs excreted by the human host are not immediately infective. They require a period of approximately 2-3 weeks after being passed out in faeces to develop to an infective stage larva. Infection is acquired by ingestion of these eggs containing 2nd stage larvae (Pecson and Nelson, 2005) which hatch in the small intestine. The released larvae penetrate the intestinal wall and migrate to the heart and lungs where they moult to the 3rd stage. They are then coughed up and swallowed, return to the small intestine, moult to the 4th stage, develop into adults, mate and produce eggs (Murrell et al., 1997; Schüle et al., 2014). Fertilised eggs can remain viable for up to 7 years (Pecson and Nelson, 2005) in the environment providing temperature (25°C) and humidity (> 55%) are optimal.
The three-layered shell of the Ascaris egg is responsible for its resistance to inactivation (Rogers, 1956). The innermost layer is a lipoprotein (ascaraside) layer, which is desiccation-resistant. It surrounds the perivitelline space, which contains perivitelline fluid and the larva (Geng et al., 2002). This is followed by a thicker chitinous layer which provides the egg shell with rigidity and shape. The third is a vitelline layer comprised of glycoproteins and is a mammillated, uterine layer which develops in the uterus of the female worm (Rogers, 1956). Hypochlorite and certain environmental stressors may be used to remove the outermost layer of the shell, known as decortication (Murrell et al., 1997). The chitin and innermost lipopro-tein layer remain. A decorticated egg is still viable, but is more susceptible to inactivation (Rogers, 1956). Hatching of Ascaris eggs can be induced outside the host body if placed in appropriate solutions (Ransom and Foster, 1919). Ascaris eggs are hydrophobic and sticky (O'Lorcain and Holland, 2000; Capizzi and Schwartzbrod, 2001) and can adhere to a variety of household surfaces, fruits and vegetables, utensils and the skin as well as soil particles.
A functioning sanitation system, defined as one which protects people from contact with their excreted faeces, is required to break the cycle of A. lumbricoides infection from human host to the environment and then to another human host. Various forms of pit latrines are common in rural communities (Mara, 1984). A ventilated improved pit latrine is an example of the minimum standard of improved sanitation recognized by the South African Government (Mara et al., 2007; RSA, 1996). Once full, pits must be sealed or emptied. This may be done mechanically using a vacuum pump, but structural limitations, terrain type, dwelling density, and the presence of solid waste in pits, mean that presently manual emptying is the best option (Mara, 1984). Contamination of workers, households and the immediate environment with faecal sludge during this process defeats the purpose of improved sanitation; thus minimising this contact is essential.
The eThekwini Municipality (Durban, South Africa) has developed a pit-emptying programme which employs community members to empty pits. Workers are provided with personal protective equipment (PPE) such as gloves, boots, overalls and face masks (PID, 2013). However, a lack of health and hygiene awareness amongst pit-emptiers means that PPE use is inconsistent, thereby increasing infection risk in workers (Mara, 1984). Cross-contamination from workers to households may occur as a result of touching taps with contaminated gloves, walking through a property with contaminated boots, or failure to clean up spillage of sludge after pit emptying (PID, 2013).
Inactivation of helminth eggs has been studied under a range of conditions. At temperatures above 60°C Ascaris eggs may be inactivated in a few minutes, but can survive for more than a year at 40°C (Brownell and Nelson, 2006). Reported methods of inactivating Ascaris eggs include lime (Capizzi-Banas et al., 2004), gamma radiation (De Souza et al., 2011), solar disinfection (Heaselgrave and Kilvington, 2011), UV light (Liberti et al., 2002), urea addition (Vinnerås et al., 2003) and ozone (De Velasquez et al., 2013). These methods are expensive and unavailable to poor householders. Other treatments such as iodine, sodium hypochlorite, ethyl alcohol, potassium hydroxide, glutaraldehyde, potassium permanganate and phenol have been used on geohelminth eggs (Ayçiçek et al., 2001), but aside from sodium hypochlorite these treatments are also not available to householders.
Thus, although there is considerable literature on inactivation of Ascaris in a range of media, it is irrelevant to inactivation of Ascaris eggs on contaminated household surfaces and in small sludge spills in homes of poor communities. This study aimed to identify simple, cost-effective ways of inactivating Ascaris eggs using household disinfectants available at supermarkets and retailers. Household contamination scenarios were simulated with respect to the mode of contamination, type of surface contaminated, disinfectant product and mode of cleaning.
METHODS
Sample collection
Stool samples were collected from 46 pit-emptying workers and 50 control group members from the Eastern Cape. The pit emp-tiers worked for a franchise group and were considered to be at risk of infection, or already infected. The control group members worked for non-sanitation departments of the local municipality. All stool donors originated from East London metropolitan area, South Africa. Sample collection complied with all necessary ethical approval (Ethics Protocol Number: HSS/0653/013, Humanities and Social Sciences Research Ethics Committee, University of KwaZulu-Natal, Westville Campus). Donors gave informed, signed consent for study participation. Stool samples were collected and screened for Ascaris eggs by the Kato-Katz technique (WHO, 1994). Participants whose stool samples tested positive for helminth infections were then treated with anthel-minthics. Positive stool samples were retained for use in disinfection experiments.
Initial helminth egg extraction from positive samples
Positive stool samples from each donor group were washed and processed in order to extract helminth eggs for use in the experiments. Eggs from positive stool samples were used for experimentation in preference to eggs from pit latrine faecal sludge due to the probability that they would already have been exposed to detergents used by home owners to clean latrines.
Briefly, stool samples which were scored positive by the Kato-Katz method were processed according to Moodley et al. (2008), as modified by Pebsworth et al. (2012), in order to extract helminth eggs. Samples were suspended in water, then sieved through a 100 μm sieve placed over a 20 μπι sieve. Material (the filtrand) retained by the second sieve was washed into a beaker and the contents allowed to settle. Most of the supernatant was poured off and the remaining sediment, plus a small amount of remaining water, was returned to the four test tubes and centri-fuged. The supernatant, after centrifugation, was pipetted off and zinc sulphate (ZnSO47H2O, specific gravity 1.3) was added to the deposit to float out the eggs. The supernatant containing the eggs was removed and repeatedly washed with water to remove the zinc sulphate. The final pellet containing helminth eggs was suspended in physiological saline solution (0.9% NaCl) for storage at 6°C until use. Positive samples from the non-sanitation workers and those from the pit emptiers were kept separate. Egg suspensions from individual Ascaris-positive stool samples were combined for each of the two exposure groups, resulting in two stock suspensions (one for pit emptiers, one for non-sanitation workers). These two suspensions were then each further split into two fractions for the two experiments, resulting in a total of four helminth egg stock suspensions.
Four household disinfectants were chosen for this study, denoted as products A to D. The first three products were chosen on the basis of their wide use in the target communities; the last was added because it was identified by franchise operators as being supplied to pit emptiers for cleaning of PPE, tools and spills. The active ingredient in both Product A and Product B was sodium hypochlorite. Although the active ingredient for Products A and B was the same, Product A also contained soap-based detergents. It was more viscous than Product B, so was considered more likely to remain in contact with a surface rather than running off quickly as might be expected from Product B. Carbolic acid was the active ingredient in Product C. The chemical composition of Product D, a thick green pine-scented gel, was unknown. For all experiments, tap water was used as the control. Egg suspensions were exposed to the disinfectants at three concentrations - the recommended dilution as indicated on the product packaging, a 50% dilution (1:1 dilution with water) and undiluted (neat) disinfectant. This study was divided into three parts: Part 1: Experiment 1 which simulated the wiping of surfaces, Part 2: a trial experiment necessary for determining a suitable contact time to be used in Experiment 2, and Part 3: Experiment 2 which entailed soaking the Ascaris eggs in different concentrations of each disinfectant, representing soaking sludge spills in disinfectant. Each combination of disinfectant product, dilution and exposure time in each part of the study was triplicated. Furthermore, each of Experiment 1 and 2 was duplicated, once with helminth eggs from pit emptiers and once with eggs from non-sanitation workers.
Assessment of viability of helminth eggs in stock suspension
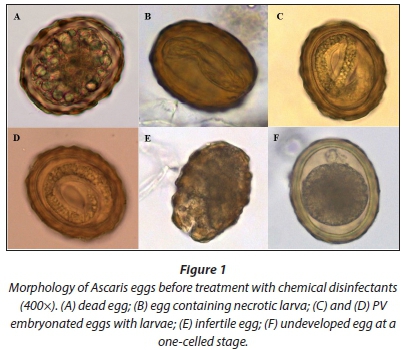
Egg viability and counts were assessed before each run of the experiment to establish the state of the eggs at the time of the experiment and to confirm the approximate number of eggs per 1 mL helminth egg stock suspension to be inoculated into each Petri dish. This was done as follows: 2 drops of well-mixed suspension were pipetted onto a glass microscope slide, covered with a 22 x 40 mm coverslip and viewed using a compound microscope. Eggs were assessed as either potentially viable (PV) or non-viable (NV), based on morphology. Potentially viable eggs included undeveloped eggs (at different cell stages), embryonated eggs with an immotile larva visible in the egg and embryonated eggs with a motile larva (Moodley et al., 2008). Non-viable eggs included mechanically broken eggs, eggs with globules inside, hatched eggs or larvae, embryonated eggs with a necrotic larva, and infertile eggs (Moodley et al., 2008). The total number of eggs per 2 drops of suspension was counted under the microscope and representative images captured. This was repeated for 3 slides and the mean count used to estimate the total number of eggs for the total number of drops (i.e. 20) in a 1 mL suspension. Eggs were assessed prior to each experiment as numbers and viability varied each time. The disinfectant solutions were prepared fresh on the day of use (Table 1).
In each experiment, the number of PV eggs recovered at the end of the experiment, relative to the number of eggs inoculated, was calculated as a percentage (Eq. 1).
Percentage recovery of PV eggs = (total PV eggs recovered/ total PV eggs inoculated) x100 (1)
Experiment 1: Assessment of helminth egg inactivation during cleaning/disinfection of surfaces
In Experiment 1, helminth eggs were exposed to disinfectant treatments on two different surfaces - glass (150 mm Petri dishes) and plastic (145 mm Petri dishes), based on unpublished observations that adhesion of eggs differed on different surfaces (Maya-Rendon, 2006). Products A, B and C were tested at the recommended dilutions (1.2%, 1.2% and 1.25%, v/v, respectively) and 50% dilution. Product D was used at the recommended dilution (25%) only as its high viscosity made it difficult to dissolve at 50% dilution. The pH of each disinfectant at each tested concentration was measured. The egg suspension was inoculated into either the glass or plastic Petri dishes, in 1 mL aliquots. The Petri dishes were agitated to spread the egg suspension evenly across the surface, and were left to dry for 30 min.
Cellulose sponge cloths (cut into pieces of 97 x 84 mm each) were used for the wiping simulation. They were rinsed and wrung once before use to remove any loose fibres or chemical residues. A single piece of cellulose sponge cloth was soaked in each of the disinfectant solutions and was wrung once. It was then wiped across the surface of the inoculated Petri dish in a clockwise motion, folded once diagonally, wiped again in an anticlockwise motion, folded again diagonally and wiped a third time in a clockwise motion. The cloth was then placed in a beaker containing 50 mL of ammonium bicarbonate (NH4HCO3) solution to facilitate the dissociation of eggs from the cloth (Moodley et al., 2008). In the first few trials, the Petri dishes were examined after wiping, using a dissecting microscope, to determine whether any eggs remained on the dishes. In these instances, the Petri dishes were filled with tap water to ensure that any residual eggs did not dry out before examination. Each cloth used for wiping was washed individually over the sieves, as previously described. The cloth was washed and rubbed slowly on both sides as well as on the edges to ensure that all eggs were removed and washed through the 100 μm sieve onto the 20 μm sieve, where the filtrand was thoroughly rinsed and transferred into two conical plastic 15 mL Falcon test tubes. Recovery of helminth eggs was performed as previously described for suspensions of stool samples. The final pellet was examined using a compound microscope (at 100x, and 400x) and viability assessed according to morphology, using the categories described previously. The entire pellet was analysed to ensure that, as far as possible, all eggs were counted and assessed.
Trial experiment: Determining a suitable contact time for soaking experiment
The contact times tested in this trial were 1 h and 12 h, using all disinfectants at 50% dilution (representing the median concentration of all being tested). A total of 6 plastic Petri dishes were used per disinfectant, with 3 being processed and examined immediately and 3 being processed and the final egg suspension incubated for a period of 28-30 days before examination. For this trial, only the egg stock suspension from non-sanitation workers was used. This was based on preliminary observations from Experiment 1 which suggested that results were similar for the two stock suspensions. The egg suspension was pipetted onto the Petri dishes in aliquots of 1 mL, to which was added 50 mL of the detergent solution. Petri dishes were left to stand for either 1 h or 12 h, after which the contents of each dish were processed as previously described. Half the samples were examined immediately, while the remainder were suspended in water and incubated at 28°C for 28-30 days to determine whether eggs developed further.
Trial experiment: Evaluating suspension media for helminth eggs for soaking experiment
A second trial experiment was conducted to evaluate media suitable for mixing with helminth eggs for Experiment 2, simulating soaking small sludge spills. The intention was to identify a medium which would simulate the protective effect of sludge in shielding helminth eggs from disinfectant action. Two candidate suspension media: (i) human faeces from a single donor (a 9-year-old child), and (ii) mealie-meal (a maize-based flour product), were tested as a locally relevant alternative to the mashed potato used by another research group (Foutch, 2014). Detailed methods and results are not given since neither human faeces nor the mealie meal was deemed suitable (as egg recovery was poor) and eggs were suspended in physiological saline instead.
Experiment 2: Inactivation of helminth eggs by soaking in disinfectant solutions
Helminth eggs (1 mL stock suspension) were inoculated onto plastic Petri dishes. The disinfectants were added to the helminth eggs and allowed to soak. Based on the trial experiment, a contact time of 1 h was selected. Although the longer contact time (12 h) yielded a greater reduction in PV eggs, the shorter contact time was more practical in a household context. Based on the egg recovery and viability from the initial experiments, both Products C and D were excluded from Experiment 2. The criterion used for disinfection was less than 10% PV eggs remaining after treatment as 90% mortality is considered significant (Ayçiçek et al., 2001). Products A and B were used at the recommended dilution, 50% dilution and undiluted. Water was used as the control. Inoculated Petri dishes were left to stand for 1 h, after which the egg suspension from each Petri dish was processed as described for Experiment 1. The final pellets were again divided equally, one for immediate examination and one for examination post-incubation.
Statistical analysis
Data were arcsine-transformed and tested for normality using a one-sample Kolmogorov-Smirnov test on IBM SPSS Statistics (Version 22). The Shapiro-Wilk test was used on the transformed data to test for normality of the residuals of the nested ANOVA. Levene's test was used to test for homogeneity of variance of residuals. A nested ANOVA was run on the arcsine-transformed data per replicate of each experiment using R (Version 3.0.2). Tukey's multiple comparisons post-hoc test was used to compare means of egg viability across and between different treatments. An independent samples i-test was performed to compare results obtained using eggs isolated from pit emptiers with those obtained using eggs isolated from non-sanitation workers in each experiment. A second independent samples i-test was performed to compare the mean percentage of PV eggs recovered between samples immediately analysed and those analysed after incubation.
RESULTS
Of the 50 samples collected from non-sanitation workers, 11 (22.0%) were positive for helminth eggs. Of the 46 samples collected from the pit emptiers, 3 (6.5%) samples were positive. This was an unexpected finding and points to the possibility of more significant infection routes than occupational exposure to faecal sludge. Upon microscopy, Ascaris eggs from the two populations differed markedly; those from the non-sanitation worker group contained a greater proportion of undeveloped eggs when compared with those from the pit-emptier group. However, eggs from the latter were in better condition.
Observations of eggs before and after treatment
Ascaris eggs responded differently when exposed to the four selected disinfectants. Morphological characteristics of the eggs were most altered in treatments which involved prolonged exposure to disinfectants. The most significant changes were observed after treatment with sodium hypochlorite-based disinfectants (Products A and B). Product C stained the eggs dark brown, making it difficult to assess egg morphology, even at high magnification. Product D had little effect on the morphology of the eggs, but did act on the suspension medium, deteriorating much of the fine particulate faecal matter which remained in the egg suspensions during preparation.
The figures below show Ascaris egg morphology as observed prior to treatment with chemical disinfectants (Fig. 1) and after being treated with sodium hypochlorite-based disinfectants (Fig. 2).

Experiment 1: Disinfection of surfaces with household disinfectants
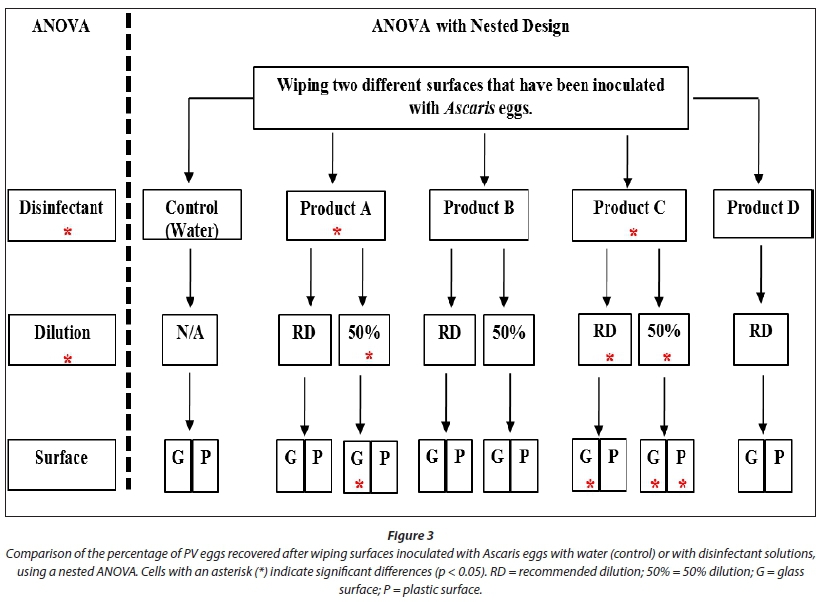
This experiment simulated the disinfection of surfaces contaminated by fleeting contact with faecal sludge, e.g., contact with a contaminated glove or boot. The percentage of PV eggs recovered after treatment was compared between the two helminth egg stock suspensions, representing the two populations of origin, using a i-test. There was no significant difference (p = 0.191), so data were combined for further analysis. The overall egg count per inoculum was assessed prior to each batch of tests run per day. The inoculated number ranged from 785-1 168 eggs/mL. The percentage of PV eggs recovered from the control treatment (water) was in the range 83-91%, thus any variation from this range was due to disinfectant action. Independently, the application of a disinfectant significantly reduced the percentage of PV eggs recovered (p < 0.005) and had a similar effect when applied at different concentrations (disinfectant*concentration; p < 0.005). However, when the effect of different surfaces was included in the model (i.e. product*concentration*surface), no significant difference was seen (Fig. 3).
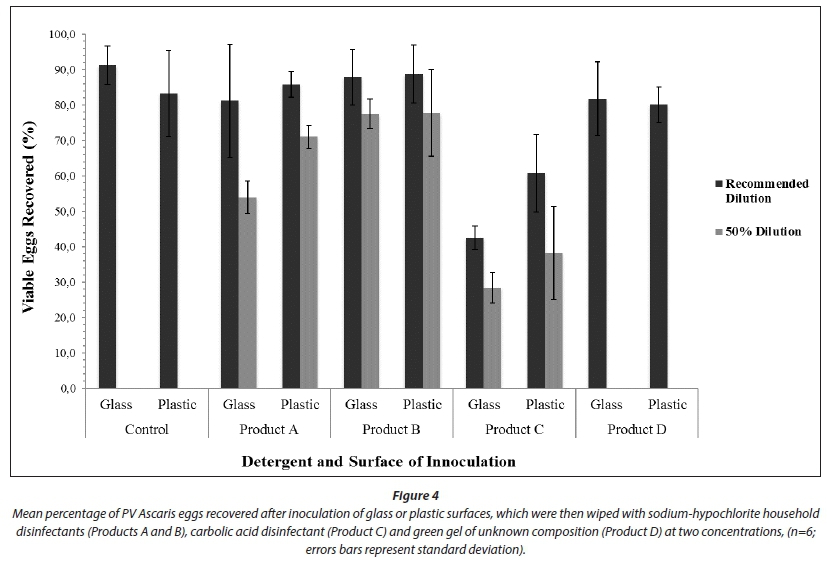
When analysed as a nested ANOVA (Fig. 3), Product A significantly reduced the percentage of PV eggs recovered (p < 0.05), both alone and at a 50% dilution (p < 0.005). Product B had no significant effect on the percentage of PV eggs recovered (Table 1). Product C significantly reduced the percentage of PV eggs recovered, alone (p < 0.005), at both concentrations (for both, p < 0.005) and on glass Petri dishes (for both, p < 0.005) (Fig. 3). However; it was ineffective in lifting eggs off surfaces (glass more so than plastic); thus the apparent reduction in percentage PV eggs recovered was due to poor egg recovery and not egg inactivation (Figs 3 and 4). Product D had no significant effect on percentage of PV eggs recovered, alone or in combination with surfaces tested. All treatments yielded a decrease in the percentage of PV eggs recovered (Fig. 1). However, recovery was too low to be considered as successful disinfection (as >10% PV eggs were recovered after treatment).
Trial experiment: Testing duration of exposure of Ascaris eggs to disinfectants
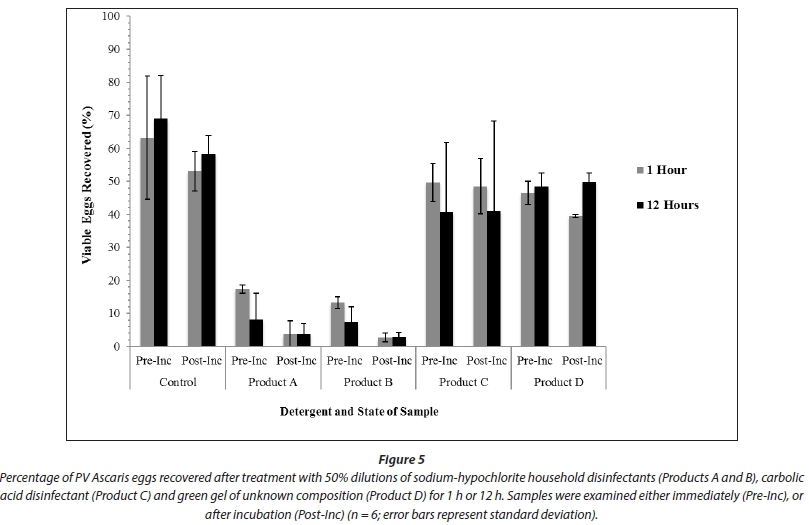
The total egg count assessed per inoculum was 1 577 eggs/ mL. Figure 5 shows the percentage recovery of PV eggs after 1 h and 12 h exposure to 50% dilution of each disinfectant on a plastic surface. Recovery of PV eggs from the control ranged from 53% before, to 69% after incubation, proving that the egg stock suspensions still contained viable eggs. Eggs treated with Products A and B both showed a very marked decline in egg recovery (p < 0.005 for differences before and after disinfectant treatment), with a shift in egg state from PV to NV after further incubation. Eggs appeared decorticated and the chitinous layer was removed upon prolonged exposure to these disinfectants. There was also a decrease in percentage of PV eggs recovered when comparing the samples examined immediately upon completion of the experiment with those incubated and then examined. This suggests that embryonated eggs with healthy larvae or undeveloped eggs shifted towards a non-viable state (necrotic or dead). Many developed, viable eggs containing motile larvae hatched upon exposure to Products A and B at 50% concentrations. Products C and D significantly reduced the percentage of PV eggs recovered relative to the control (p = 0.006 and 0.0153, respectively); however, the percentages were too high for disinfection to be considered successful (> 10% PV eggs recovered). On this basis, Products C and D were excluded from the second experiment.
When observing the overall effect of the disinfectant alone (independent of exposure time or whether the sample was examined immediately or incubated before examination), no significant difference in percentage viability of recovered eggs was observed (p = 0.600). The combined effects of disinfectant with contact time and point of analysis of the sample also showed no overall significant difference in percentage of PV eggs recovered (p = 0.812 and 0.810, respectively). The contact time selected for the second experiment was 1 h, because substantial reductions in recovery of PV eggs were seen for treatments with Products A and B after 1 h, and because a 12-h soaking time would probably be impractical to achieve in a household environment.
Experiment 2: Disinfection of Ascaris eggs by soaking with disinfectants
This experiment simulated disinfection of small faecal sludge spills by soaking with disinfectant. As per Experiment 1, Experiment 2 was conducted with Ascaris eggs obtained from pit emptiers (inoculum used, 413 eggs/mL) and, separately, with eggs from non-sanitation workers (inoculum 1414 eggs/ mL). The total egg count per inoculum was 914 ± 708 eggs/mL. No significant difference in percentage egg recovery was found between the two groups (p = 0.653), so the percentages from the two replicate experiments were combined for further analysis.
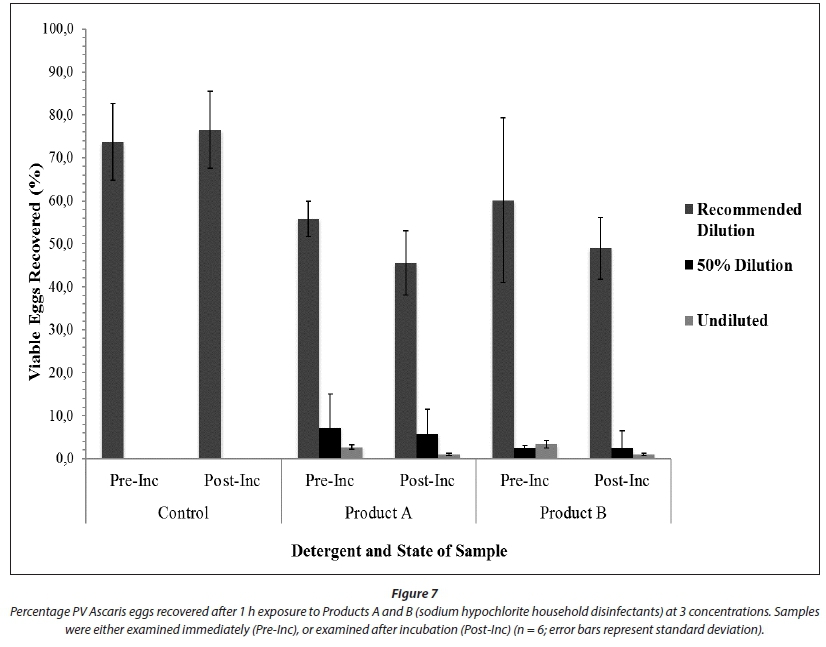
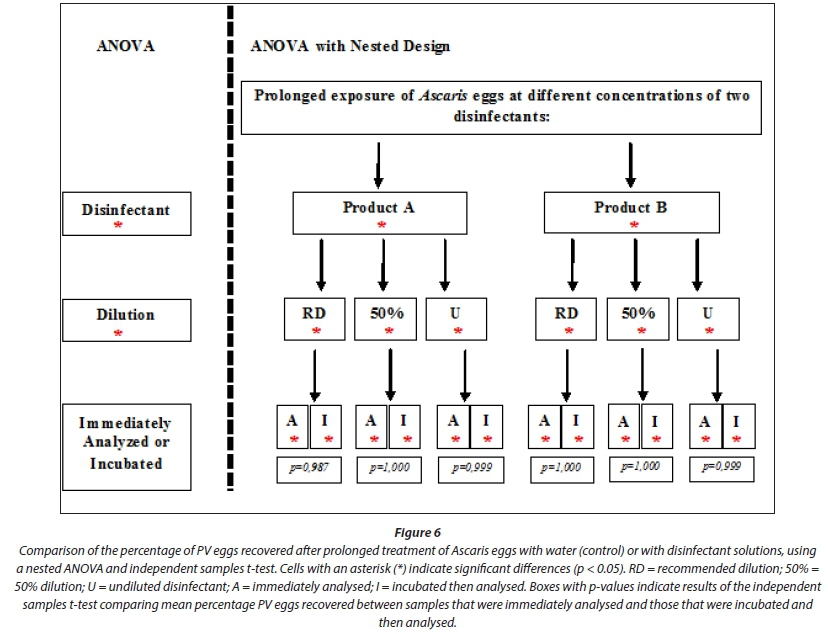
Percentage recovery for the control ranged from 73% to 77% (Fig. 7). When compared alone, the disinfectants significantly reduced the percentage of PV eggs recovered (p < 0.005). There was also a significant difference when the dilution factor was included in the model (p < 0.005). The point at which the sample was examined (either immediately or post-incubation) had no significant effect on the percentage of PV eggs recovered (p = 0.687) (Fig. 6). Independently, Product A significantly reduced the percentage of PV eggs recovered (p < 0.005) (Fig. 6). When combined with the recommended dilution, 50% dilution and undiluted disinfectant, the percentage of PV eggs recovered was significantly reduced by all combinations of factors (disinfectant*concentration; p < 0.005, p < 0.005 and p < 0.005, respectively). A significant reduction in egg viability was seen for both immediately analysed samples and samples that were incubated and then analysed (disinfectant*concentration*point of analysis; Fig. 6). Product B alone, and when coupled with the three dilutions and the point of analysis of the sample, had a significant effect on the percentage of PV eggs recovered (p < 0.005, p < 0.005 and p < 0.005 respectively).
Although significant changes were seen in the percentage of PV eggs recovered between the control and treatments, the recommended dilutions yielded PV egg recovery above 10% and therefore could not be considered as providing safe disinfection. Both Products A and B, at dilutions of 50%, yielded PV egg recoveries of < 10%, and therefore can be considered to provide successful disinfection (Fig. 7). The undiluted disinfectant yielded even lower recoveries, with less than 5% PV eggs recovered after incubation. The number of eggs recovered from both 50% and undiluted treatments were minimal, with developed eggs being highly decorticated.
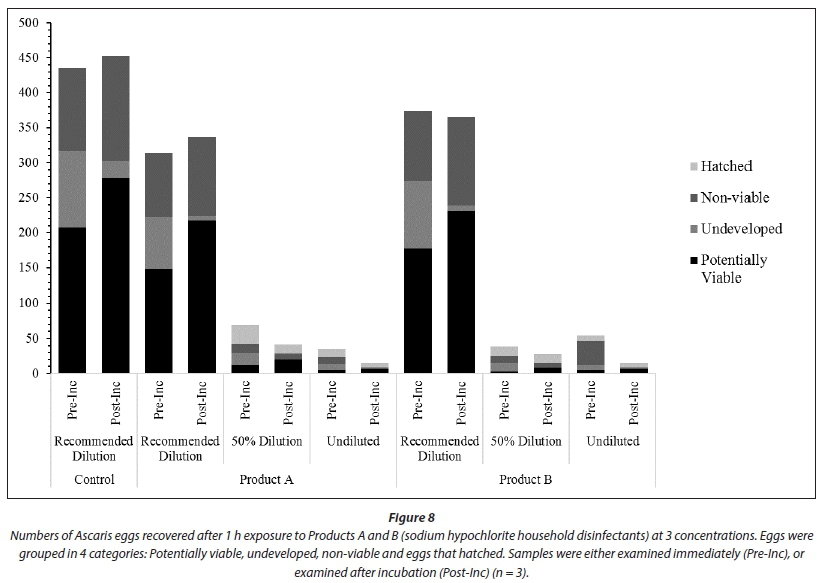
Marked shifts in egg morphology were observed after incubation of eggs, from an undeveloped state to a potentially viable state (Fig. 8). Larvae within developed eggs were highly motile. Eggs treated with the recommended dilution for the two disinfectants exhibited the greatest shift, indicating that further development occurred after being treated with a chemical disinfectant. Further development was also evident at 50% dilution. However, there was also an increase in percentage of inactivated or NV eggs recovered (Fig. 7). Hatching of eggs occurred at 50% dilution and in undiluted treatments, both before and after incubation (Fig. 8). Larvae died quickly after hatching and were therefore classified as NV. No significant difference was therefore found between the egg viability of samples immediately analysed and those incubated and then analysed (Fig. 6; p > 0.05 for all factor combinations). At 50% dilution and in undiluted treatments with Products A and B, the percentage of PV eggs recovered was much lower than from the recommended dilution.
pH of disinfectants
The pH values of all four disinfectants at all dilutions were basic, with undiluted Product A being the most basic and Product D at the recommended dilution being the least basic (Table 1). The pH was considered as a factor influencing inactivation of Ascaris eggs in this study.
DISCUSSION
The results from Experiment 1 (Fig. 4) indicate that wiping a contaminated surface does not inactivate helminth eggs. Whilst wiping does facilitate transfer of eggs from the contaminated surface to the cloth, the limited exposure of the eggs to the disinfectant on the cloth is not sufficient for inactivation. Egg recovery from plastic and glass surfaces was high, except in the case of Product C on a glass surface. Upon examination of the glass Petri dishes treated with Product C, many residual eggs were found. When used on a cloth to wipe glass surfaces, Product C remained behind on the glass. This concurs with observations by Capizzi and Schwartzbrod (2001) that Ascaris suum eggs displayed hydrophobic properties when mixed with an emulsified octane suspension, causing the eggs to adhere to hydrocarbon droplets. Eggs also adhered to glass powder (Capizzi and Schwartzbrod, 2001). Since Product C is oil-based and Ascaris eggs are hydrophobic, it is likely that eggs adhered to droplets of Product C which remained behind when wiping the glass. This reduced the percentage of PV eggs recovered (Fig. 4) but not as a result of egg inactivation. Products A and B, at both concentrations and on both surfaces, did not meet the disinfection criterion of < 10% PV eggs remaining. Product D reduced the percentage of PV eggs recovered, but also did not meet the disinfection criterion.
The pH of Products A, B, C and D at the recommended dilutions was 12.3, 11.8, 8.5 and 8.2, respectively. Pecson and Nelson (2005) showed that a pH of 11, combined with a temperature of 38-40°C was needed to achieve 99% inactivation of helminth eggs suspended in an ammonium chloride solution. Prolonged exposure of up to 90 days was required for such inactivation (Ghiglietti et al., 1997; Pecson and Nelson, 2005). Bina et al. (2004) reported that lime treatment resulted in successful elimination of pathogenic bacteria; however, pH of 11-12 over a period of 5 days did not inactivate helminth eggs. The present investigation was conducted at room temperature (25°C) without prolonged exposure to the detergents in Experiment 1, and only 1 h exposure in Experiment 2. In the light of other studies, the inefficacy of the recommended dilutions of all four disinfectants to inactivate Ascaris eggs adequately by simple wiping of the contaminated surface is not unexpected. However, the observation that wiping with disinfectant removed eggs from the contaminated surface onto the wiping cloth is promising, since cloths can be disinfected by soaking in disinfectant after wiping contaminated surfaces.
One hour exposure of a simulated sludge spill to both Products A and B (sodium hypochlorite-based), at concentrations of 50% and above, successfully inactivated Ascaris eggs (Experiment 2; Figs 7 and 8). The recommended dilutions did not meet the disinfection criterion of 10% or less PV eggs remaining. The pH of Products A and B, at 50% dilution and undiluted, was extremely basic (Table 1). Exposure to the high pH and oxidizing properties of these products for relatively short periods at room temperature successfully decreased the percentage of PV eggs recovered. Fukuzaki (2006) highlighted the complementary roles of HOCl and -OCl in inactivating bacteria by oxidation from both outside and inside the cell. Biological molecules containing C=C double bonds and amino groups are especially susceptible to oxidation. A concentrated sodium hypochlorite solution with pH > 12.5 results in the degradation of the proteins, fatty acids and other constituents of the cell wall and membrane (Fukuzaki, 2006). This could account for the observed decortication of eggs.
The pH of the stomach is strongly acidic, but pH of the gastrointestinal tract is in the range 6.5-7.5, with the small intestine of humans having pH 7.4 (Fallingborg, 1999). Ascaris eggs can be expected to be adapted to an increase in pH to an optimum within the neutral range. However, further increase into the basic range is detrimental. Although tested under different conditions (varying temperatures, drying, long exposure times, varying suspension media, and treatment with alkalising agents such as lime and urea), high pH has been consistently linked to inactivation of Ascaris eggs (Maya et al., 2012; Pecson et al., 2007; Katakam et al., 2014). The combination of high pH and oxidizing properties of Products A and B probably account for the disinfection of Ascaris reported here.
The sodium hypochlorite-based disinfectant solutions decorticated Ascaris eggs (and removed the chitin layer) at concentrations of 50% and above, resulting in the innermost egg layer appearing almost membranous (Fig. 2). Many of the PV eggs which were recovered post-treatment subsequently hatched (Fig. 8). Hatching of Ascaris eggs occurs due to the combination of rapid L3 larval movement, digestive action by chitinases and proteases in the perivitelline fluid and altered permeability of the lipid layer (Geng et al., 2002; O'Connor, 1951). It is likely that chemical removal of the outermost layers of the Ascaris eggs combined with movement of the developed larvae (L3), facilitated hatching of the eggs. This is supported by O'Connor's (1951) description that extra-corporeal hatching of Ascaris eggs may be attributed to the escape of the larvae due to mechanical or chemical destruction of the egg shell and not to biological hatching. Hatching of an Ascaris egg outside the small intestine of the primary host results in disintegration of the larva (Pawlowski, 1982). Hatching outside the host may thus be considered as successful inactivation.
CONCLUSIONS
This study tested the effects of four common chemical disinfectants on helminth eggs. The criterion used for successful disinfection was < 10% potentially viable eggs remaining after treatment. None of the four disinfectants met this criterion when used to wipe contaminated surfaces due to the very short exposure time of the eggs to the four disinfectants at both dilutions. Wiping of a surface did, however, successfully transfer eggs from the surface to the wiping cloth which could then be soaked in one of the detergents at the optimum dilution required to inactivate the eggs.
In Experiment 2 (where spills were soaked), Products A and B, both hypochlorite-based, were effective in inactivating Ascaris eggs at 50% dilution and undiluted, reducing the percentage of potentially viable eggs recovered to 5-10%. Hypochlorite-treated Ascaris eggs appeared highly decorticated (with the chitin layer removed as well) at these concentrations, facilitating hatching of the eggs which rendered the emerging larvae non-viable. It can therefore be recommended that heavily contaminated surfaces or small sludge spills be disinfected by soaking with either Product A or Product B, either undiluted or at 50% dilution. Surfaces that are lightly contaminated and cannot be soaked for a prolonged period of time can be wiped with a disinfectant-saturated cloth and the cloth should then be soaked for an hour or more in either Product A or B at concentrations at or above 50%. Such disinfectants may also be poured onto sludge spills. Although commonly used in low-cost households and supplied to pit workers, both Product C and D were ineffective for disinfection of helminth eggs on surfaces, by wiping and with soaking. It is noted that although the ideal situation would be for disinfectants to kill 100% of parasite eggs, in reality, this is impractical for poor householders to achieve. Thus lowering their risk of infection to below 10% with minimal effort and cost to them is a worthwhile alternative.
Numerous studies have focused on the inactivation of helminth eggs. However, most have investigated the effects of various environmental factors and treatments on eggs in faecal sludge, rather than on suspensions of eggs in water or other solutions, which would indicate the direct effects of treatment on eggs without protection by an organic matrix. Further research is required to ascertain the effect of hypochlorite-based disinfectants on helminth eggs that are suspended in faecal sludge and to investigate the role of the organic matter in the protection of eggs against disinfection. It may also be of value to investigate the efficacy of other inactivation methods, such as liming and solar disinfection, in disinfecting contaminated surfaces and small sludge spills for use in the development of safe practices for the manual emptying of pit latrines.
ACKNOWLEDGEMENTS
Mrs Valerie Kelly is thanked for initial screening of faecal samples, and for her guidance and support. This study was funded by the Water Research Commission (WRC Project K5/2134).
REFERENCES
AWASTHI S, BUNDY DAP and SAVIOLI L (2003) Helminthic infections. Brit. Med. J. 327 431-433. http://dx.doi.org/10.1136/bmj.327.7412.431 [ Links ]
AYÇİÇEK H, YARSAN E, SARIMEHMETO ĞLU HO, TANYÜKSEL M, GlRGlNKARDE$LER N and ÖZYURT M (2001) Efficacy of some disinfectants on embryonated eggs of Toxocara canis. Turk. J. Med. Sci. 31 35-39. [ Links ]
BETHONY J, BROOKER S, ALBONICO M, LOUKAS A and HOTEZ PJ (2006) Soil-transmitted helminth infections: Ascariasis, trichuriasis and hookworm. Lancet 367 1521-1532. http://dx.doi.org/10.1016/S0140-6736(06)68653-4 [ Links ]
BINA B, MOVAHEDIAN H and KORD I (2004) The effect of lime stabilization on the microbial quality of sewage sludge. Iran. J. Environ. Health Sci. Eng. 1 (1) 34-38. [ Links ]
BROWNELL SA and NELSON KL (2006) Inactivation of single-celled Ascaris suum eggs by low-pressure UV radiation. Appl. Environ. Microbiol. 72 (2) 2178-2184. http://dx.doi.org/10.1128/AEM.72.3.2178-2184.2006 [ Links ]
CAPIZZI-BANAS S, DELOGE M, REMY M and SCHWARTZBROD J (2004) Liming as an advanced treatment for sludge sanitisation: Helminth egg elimination- Ascaris eggs as model. Water Res. 38 3251-3258. http://dx.doi.org/10.1016/j.watres.2004.04.015 [ Links ]
CAPIZZI S and SCHWARTZBROD J (2001) Surface properties of Ascaris suum eggs: hydrophobic potential and Lewis acid-base interactions. Colloids Surf. B: Bioinf. 22 99-105. http://dx.doi.org/10.1016/s0927-7765(01)00145-x [ Links ]
DE SOUZA GSMB, RODRIGUES LA, DE OLIVEIRA WJ, CHERNICHARO CAL, GUIMARÄES MP, MASSARA CL and GROSSI PA (2011) Disinfection of domestic effluents by gamma radiation: Effects on the inactivation of Ascaris lumbri-coides eggs. Water Res. 45 5523-5528. http://dx.doi.org/10.1016/j.watres.2011.08.008 [ Links ]
DE VELASQUEZ TA, MARTINEZ JL, MONJE-RAMIREZ I and ROJAS-VALENCIA N (2004) Destruction of helminth (Ascaris suum) eggs by ozone. Ozone: Sci. Eng. 26 359-366. http://dx.doi.org/10.1080/01919510490482188 [ Links ]
ESREY SA, POTASH JB, ROBERTS L and SHIFF C (1991) Effects of improved water supply and sanitisation on ascariasis, diarrhoea, dracunuliasis, hookworm infection, schistosomiasis and trachoma. Bull. World Health Org. 69 (5) 609-621. [ Links ]
FALLINGBORG J (1999) Intraluminal pH of the human gastrointestinal tract. Dan. Med. Bull. 46 (3) 183-196. [ Links ]
FOUTCH G (2014) Personal communication, 1 July 2014. Dr Gary Foutch, Oklahoma State Univeristy, United States of America. [ Links ]
FUKUZAKI S (2006) Mechanisms of actions of sodium hypochlorite in cleaning and disinfection processes. Biocontrol Sci. 11 (4) 147-157. http://dx.doi.org/10.4265/bio.11.147 [ Links ]
GENG J, PLENEFISCH J, KOMUNIECKI PR and KOMUNIECKI R (2002) Secretion of a novel developmentally regulated chitinase (family 19 glycosyl hydrolase) into the perivitelline fluid of the parasitic nematode, Ascaris suum. Mol. Biochem. Parasitol. 124 11-21. http://dx.doi.org/10.1016/S0166-6851(02)00155-X [ Links ]
GHIGLIETTI R, GENCHI C, MATTEO LD, CALCATERRA E and COLOMBI AM (1997) Survival of Ascaris suum eggs in ammonia-treated wastewater. Bioresour. Technol. 59 195-198. http://dx.doi.org/10.1016/S0960-8524(96)00147-2 [ Links ]
HEASELGRAVE W and KILVINGTON S (2011) The efficacy of simulated solar disinfection (SODIS) against Ascaris, Giardia, Acanthamoeba, Naegleria, Entamoeba and Cryptosporidium. Acta Tropica 119 138-143. http://dx.doi.org/10.1016/j.actatropica.2011.05.004 [ Links ]
HEIJNEN M, CUMMING O, PELETZ R, CHAN GKS, BROWN J, NAKER K and CLASEN T (2014) Shared sanitation versus individual household latrines: A systematic review of health outcomes. PLoS ONE 9 (4) e93300. doi:10.1371/journal.pone.0093300. http://dx.doi.org/10.1371/journal.pone.0093300 [ Links ]
JOHANNESSEN A, ROSEMARIN A, THOMALLA F, SWARTLING AG, STRENSTROM TA and VULTURIUS G (2014) Strategies for building resilience to hazards in water, sanitation and hygiene (WASH) systems: The role of public private partnerships. Int. J. Disaster Risk Reduction 10 102-115. http://dx.doi.org/10.1016/j.ijdrr.2014.07.002 [ Links ]
KATAKAM KK, MEJER H, DALSGAARD A, KYVSGAARD NC and THAMSBERG SM (2014) Survival of Ascaris suum eggs in liquid manure at different ammonia concentrations and temperatures. Vet. Parasitol. 204 249-257. http://dx.doi.org/10.1016/j.vetpar.2014.05.017 [ Links ]
LIBERTI L, NOTARNICOLA M and PETRUZZELLI D (2002) Advanced treatment for municipal wastewater reuse in agriculture. UV disinfection: Parasite removal and by-product formation. Desalination 152 315-324. http://dx.doi.org/10.1016/S0011-9164(02)01079-2 [ Links ]
MARA D (1984) The Design of Ventilated Improved Pit Latrines. The International Bank for Reconstruction and Development/The World Bank, Washington DC. 1 pp. [ Links ]
MARA D, DRANGERT J-O, ANH NV, TONDERSKI A, GULYAS H and TONDERSKI K (2007) Selection of sustainable sanitation arrangements. Water Polic. 9 305-318. http://dx.doi.org/10.2166/wp.2007.009 [ Links ]
MAYA C, TORNER-MORALES FJ, LUCARIO ES, HERNÁNDEZ E and JIMÉNEZ B (2012) Viability of six species of larval and non-larval helminth eggs for different conditions of temperature, pH and dryness. Water Res. 46 4770-4782. http://dx.doi.org/10.1016/j.watres.2012.06.014 [ Links ]
MAYA-RENDON C (2006) Personal (verbal) communication (unpublished observation), 2006. C Maya-Rendon, Universidad Nacional Autónoma de México, Environmental Engineering Department. [ Links ]
MOODLEY CL, ARCHER C, HAWKSWORTH D and LEIBACH L (2008) Standard methods for the recovery and enumeration of helminth ova in wastewater, sludge, compost and urine diversion waste in South Africa. WRC Report no. TT 322/08, Water Research Commission, Pretoria. [ Links ]
MURRELL DK, ERIKSEN L, NANSEN P, SLOTVED H-C and RASMUSSEN T (1997) Ascaris suum: A revision of its early migratory path and implications for human ascariasis. J. Parasitol. 83 (2) 255-260. http://dx.doi.org/10.2307/3284450 [ Links ]
O'CONNOR GR (1951) Morphological and environmental studies on the hatching of ascarid eggs in vitro. J. Parasitol. 37 (2) 179-182. http://dx.doi.org/10.2307/3273450 [ Links ]
O'LORCAIN P and HOLLAND CV (2000) The public health importance of Ascaris lumbricoides. Parasitology 121 S51-S71. http://dx.doi.org/10.1017/s0031182000006442 [ Links ]
PARTNERS IN DEVELOPMENT (2013) Controlled study to establish pathways of exposure in the handling and desludging of onsite sanitation systems - a literature review. WRC Project K5/2134 (Deliverable 7; Reference Group Meeting 16 October 2014, unpublished). Water Research Commission, Pretoria. [ Links ]
PAWLOWSKI ZS (1982) Ascariasis: Host-pathogen biology. Rev. Infect. Dis. 4 (4) 806-814. http://dx.doi.org/10.1093/4.4.806 [ Links ]
PEBSWORTH PA, ARCHER CE, APPLETON CA and HUFFMAN MA (2012) Parasite transmission risk from geophagic and foraging behavior in Chacma baboons. Am. J. Primatol. 74 940-947. http://dx.doi.org/10.1002/ajp.22046 [ Links ]
PECSON B, BARRIOS JA, JIMÉNEZ BE and NELSON KL (2007) The effects of temperature, pH, and ammonia concentration on the inactivation of Ascaris eggs in sewage sludge. Water Res. 41 2893-2902. http://dx.doi.org/10.1016/j.watres.2007.03.040 [ Links ]
PECSON BM and NELSON KL (2005) Inactivation of Ascaris suum eggs by ammonia. Environ. Sci. Technol. 39 (20) 7909-7914. http://dx.doi.org/10.1021/es050659a [ Links ]
RANSOM BH and FOSTER WD (1919) Recent discoveries concerning the life history of Ascaris lumbricoides. J. Parasitol. 5 (3) 93-99. http://dx.doi.org/10.2307/3271160 [ Links ]
ROGERS RA (1956) A study of eggs of Ascaris lumbricoides var. suum with the electron microscope. Journal of Parasitology Archives 42 (2)97-108. http://dx.doi.org/10.2307/3274720 [ Links ]
RSA (Republic of South Africa) (1996) National Sanitation Policy. (White Paper). National Sanitation Task Team, Republic of South Africa. [ Links ]
SCHÜLE SA, CLOWES P, KROIDL I, KOWUOR DO, NSOJO A, MANGU C, RIESS H, GELDMACHER C, LAUBENDER RP, MHINA S, MABOKO L, LÖSCHER T, HOELSCHER M and SAATHOFF E (2014) Ascaris lumbricoides infection and its relation to environmental factors in the Mbeya region of Tanzania, a cross-sectional, population-based study. PLoS ONE 9 (3) e92032. doi:10.1371/journal.pone.0092032. [ Links ]
UNICEF/WHO (2009) Diarrhoea: Why Children Are Still Dying And What Can Be Done. World Health Organisation, Geneva, Switzerland. http://dx.doi.org/10.1371/journal.pone.0092032 [ Links ]
VINNERÄS B, HOLMQVIST A, BAGGE E, ALBIHN A and JÖNSSON H (2003) The potential for disinfection of separated faecal matter by urea and by peracetic acid for hygienic nutrient recycling. Bioresour. Technol. 89 155-161. http://dx.doi.org/10.1016/S0960-8524(03)00044-0 [ Links ]
WHO (1994) Bench Aids for the Diagnosis of Intestinal Parasites. World Health Organisation, Geneva, Switzerland. [ Links ]
Received 1 September 2015
Accepted in revised form 6 September 2016
* To whom all correspondence should be addressed. +27 (0)84 5871500; e-mail: archerc@ukzn.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}