










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkWater SA
versión On-line ISSN 1816-7950
versión impresa ISSN 0378-4738
Water SA vol.41 no.4 Pretoria jul. 2015
http://dx.doi.org/10.4314/WSA.V41I4.06
ARTICLES
Water use characteristics of a bambara groundnut (Vigna subterranea L. Verdc) landrace during seedling establishment
Tendai Polite Chibarabada*; Albert Thembinkosi Modi; Tafadzwanashe Mabhaudhi
Crop Science, School of Agricultural, Earth and Environmental Sciences, University of KwaZulu-Natal, Private Bag X01, Scottsville 3209, Pietermaritzburg, South Africa
ABSTRACT
Bambara groundnut is slow to establish and this has negative implications for total water use. Consideration of bambara groundnut as a water-efficient crop for dry areas will benefit from an understanding of water use efficiency and water use characteristics during establishment. We investigated whether there is an association between seed-coat colour of bambara groundnut seeds and water use efficiency. A secondary objective was to determine physiological and metabolic responses of bambara groundnut to terminal water stress during seedling establishment. Bambara groundnut seedlings showed reduced seedling canopy growth, increased root length and root: shoot ratio, as well as increased total soluble sugars and tissue water potential with decreasing water availability - traits associated with drought avoidance mechanisms. Consequently, seedling water use efficiency improved with decreasing water availability. Acclimation to water stress was observed under terminal stress; previously stressed seedlings responded better than previously non-stressed seedlings. Dark-coloured seeds had higher levels of phenolics which may have led to their enhanced performance during seedling emergence as well as improved drought tolerance and seedling WUE.
Keywords: drought, physiological, metabolic, growth, water use efficiency
INTRODUCTION
Uniform and early crop establishment are intricately related to crop yield (Tekrony and Egli, 1991). An uneven crop stand limits the crop's ability to fully utilize resources such as light and water. This leads to yield losses from which farmers cannot recover (Mabhaudhi and Modi, 2010). Bambara groundnut is slow to establish (Makanda et al., 2009; Mabhaudhi and Modi, 2013). This leads to significant non-productive water losses through soil evaporation during the establishment stage (Mabhaudhi, 2012), which may have negative implications on total water use. As such, consideration of bambara groundnut as a water-use efficient crop for dry areas must take into account water use efficiency (WUE) during seedling establishment. To date, studies that have determined the WUE of bambara groundnut have looked at it wholly with none determining WUE during the establishment stage.
Water use efficiency is the yield of harvested products achieved from water made available to the crop (Blum, 2005). Water use efficiency in bambara groundnut has been determined by Azam-Ali et al. (2004) and Mabhaudhi et al. (2013). They found WUE values ranging between 0.09 and 0.1 kg.m-3. These were higher than WUE values of 0.0025 kg.m-3 (Nageswara Rao et al., 1993) and 0.0018 kg.m-3 (Songsri et al., 2013) reported for its exotic counterpart Arachis hypogaea. This implies that bambara groundnut is a more suitable crop than Arachis hypogea during periods of drought. However, these values are for total crop WUE. With the reported slow establishment of bambara groundnut and significant unproductive water loss during seedling establishment, it can be hypothesized that total crop water use could be improved if crop establishment was improved. An initial step to investigate this issue is to quantify water use efficiency at the establishment stage. Water use efficiency has been reported to increase with decreasing water supply (Nageswara Rao et al., 1993; Mabhaudhi et al., 2013; Songsri et al., 2013). Genotypic (Nageswara Rao et al., 1993; Songsri et al., 2013) and seed colour variation (Mabhaudhi et al., 2013) in landraces has also been shown to influence WUE.
Several drought-tolerance mechanisms have been shown to contribute to high WUE in bambara groundnut. These include drought escape (Vurayai et al., 2011; Mabhaudhi and Modi, 2013) and avoidance mechanisms (Jorgensen et al., 2010; Vurayai et al., 2011; Mabhaudhi and Modi, 2013). There is scant information on accumulation of plant metabolites in bambara groundnut in response to drought. Plants accumulate a range of osmotically-active metabolites under water stress, including total soluble sugars and proline, in order to maintain water relations under osmotic stress (Nazarli and Faraji, 2011). Vurayai et al. (2011) and Zondi (2013) showed that proline was higher in water-stressed plants than in non-stressed plants, indicating that proline plays a role in drought tolerance of bambara groundnut. There is, however, no information in the literature on accumulation of soluble sugars in bambara groundnut during water stress, or on the plant's antioxidant defence systems in response to water stress at the seedling stage.
Understanding physiological and metabolic responses of bambara groundnut at the establishment stage will aid in understanding water-use characteristics of bambara groundnut and how they influence WUE. Previous research, using plain bambara groundnut seed colours, showed an association between seed colour and drought tolerance, with darker-coloured bambara groundnut seeds being more drought tolerant compared to light-coloured seeds (Sinefu, 2011; Mabhaudhi and Modi, 2013). Mabhaudhi and Modi (2013) further attributed this to the presence of phenolic compounds in dark-coloured seeds acting as a defensive mechanism against stress.
This study is a sequel to previous studies (Sinefu, 2011; Mabhaudhi and Modi, 2013; Mabhaudhi et al., 2013; Zondi, 2013) and investigates whether there is also an association between colour of speckles in bambara groundnut seeds, drought tolerance mechanisms and water use efficiency. A secondary objective was to determine physiological (chlorophyll content index, leaf water potential and photosynthetic efficiency) and metabolic (total antioxidant capacity, total pheno-lics and total soluble sugars) responses of bambara groundnut to water stress during the seedling establishment stage.
MATERIALS AND METHODS
Plant material
Bambara groundnut seed was obtained from Capstone Seeds, Mooi River, in a mixed seed colour batch. Thereafter, seeds were characterized into 4 distinct selections based on seed coat colour, namely: plain red, plain cream, cream with brown speckles (brown-speckled) and cream with black speckles (black-speckled) (Fig. 1). The basis for using seed coat colour as a selection criterion has been described elsewhere (Sinefu, 2011; Mabhaudhi and Modi, 2013; Mabhaudhi et al., 2013).
Controlled environment
The experimen: was conducted in a growth tunnel at the University of KwaZulu-Natal's Controlled Environment Facility (CEF). The environmental conditions in the tunnel were 33/27°C (day/night) temperatures, 60-82% relative humidity (RH) and natural day length; the environment in the tunnels is representative of a warm tropical climate (Modi, 2007).
Experimental design
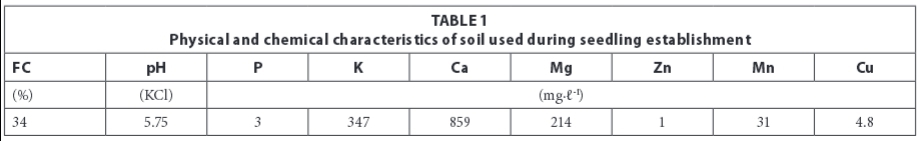
Seedling establishment was done using 128-unit seedling trays. The seedling trays were filled with a silt-loam soil (Table 1). The experimental design was a split-plot design with water regimes as the main factor and bambara groundnut landrace selections as the sub-factor, arranged in a randomized complete block design. The experiment was replicated 4 times. There were 3 water regimes: 25%, 50%, and 75% (control) of field capacity (FC). The four bambara groundnut landrace selections were plain red, plain cream, brown-speckled and black-speckled. One seed was planted per planting station. After planting, all trays were watered up to 75% of field capacity for 10 days to ensure maximum emergence. Thereafter, the different watering regimes were imposed by allowing soil water depletion until trays had attained their corresponding field capacities. Three extra trays that were treated in the same way as experimental trays were also established for the purpose of destructive sampling.
Watering regimes
Field capacity of the media was determined following the gravimetric field capacity test (Mabhaudhi, 2009). Three small drained pots (representing 3 replicates) were used. Each pot was filled with media. Thereafter, water was added to the pots until saturation was achieved. Pots were then left to drain for 12 h and thereafter mass of soil was measured hourly until a constant mass was reached. At this point it was assumed that the soil was now at field capacity. Following this, the soil was taken out, put in labelled brown paper bags and wet mass determined. Thereafter, brown bags with soil in them were dried in an oven at 80°C for 72 h, after which dry mass of the soil was measured. Gravimetric field capacity (FC) was then calculated as follows:

where: θm = gravimetric field water capacity
θw = wet mass of soil
θd = dry mass of soil
Water was applied based on gravimetric water content (1 g = 1 mℓ) every 2 days by weighing trays to determine the amount of water used by the plants and then refilling the trays to their corresponding field capacities. Water added at each irrigation event was recorded in order to determine water use at the end of the experiment. Irrigation was withdrawn from all of the trays, except the control, from 21 days after planting (DAP) until termination of the experiment (at 28 DAP).
Data collection
Growth and physiology
Visual counts of emerged seedlings were taken daily from 1 DAP up to 10 DAP. A seedling was considered to have emerged when the cotyledon had fully emerged. Mean emergence time was calculated using the formula of Bewley and Black (1994):

where: MET = mean emergence time, ƒ = number of newly germinated seeds at a given time (day), and x = number of days from date of sowing.
Thereafter, seedling height was measured weekly (from 7 DAP) until the end of seedling establishment (21 DAP). Seedling height was measured using a 30-cm ruler from the base of the main stem to the base of the longest secondary stem. Seedling leaf number was determined weekly (from 14 DAP up to 21 DAP) by visually counting leaves that were fully expanded and with more than 50% green leaf area. Each trifoliate leaf was counted as 1 leaf. Following that, 3 seedlings per replicate per landrace selection were destructively sampled and total leaf surface area per seedling was measured using an LI-3000C Portable Leaf Area Meter connected to a LI-3050C Transparent Belt Conveyer (LI-COR, USA). Upon termination of the experiment (at 21 DAP), seedling parameters (seedling length, root and shoot length, root: shoot ratio and seedling dry mass) were determined.
Chlorophyll content index (CCI) was measured weekly (from 7 DAP up to 21 DAP) using the SPAD-502Plus Chlorophyll Meter (Konica Minolta, USA) on the adaxial surface of fully expanded, fully exposed and actively photosynthesizing leaves. In order to determine plant photosynthetic efficiency, chlorophyll fluorescence (CF) was measured weekly (from 7 DAP up to 21 DAP) using a Pocket PEA-Chlorophyll fluorescence system (Hansatech Instruments, United Kingdom). Chlorophyll fluorescence was measured on the adaxial surface of young, fully expanded and fully exposed green leaves. Before measuring CF a sample area of the targeted leaf was covered with a lightweight leaf clip (Hansatech Instruments, UK) for 20 min to exclude light and allow for dark adaptation. Weekly measurements of CCI and CF were taken before an irrigation event. After 21 DAP, when irrigation was withdrawn (except the control), measurements of CCI and CF were taken daily until 27 DAP when the experiment was terminated. Measurements of CCI and CF were routinely taken at midday.
On the 21st day after planting, before irrigation was withdrawn, measurements of pre-dawn leaf water potential (PDLWP) and midday stem water potential (MSWP) were taken using the 3005F01 Portable Plant Water Status Console (Soilmoisture Equipment Corp, USA). Measurements were taken on a healthy, mature, fully-exposed terminal leaf. A random order of destructive sampling within each watering regime and among landrace selections was followed for each replicate. Pre-dawn leaf water potential readings were taken from 3.30 am to 5 am (before sunrise), based on the assumption that before sunrise the plant is at equilibrium with soil water potential and that PDLWP is hence a more sensitive indicator of soil water availability (Obso, 2006). At midday, stem water potential was preferred because it has been shown to be less susceptible to fluctuations in environmental pressures than leaf water potentials, and is therefore more representative of the actual level of stress (McCutchan and Shackel 1992; Chone et al., 2001; Williams and Araujo, 2002). Midday stem water potential was measured between 12 noon and 1.30 pm.
Before cutting, the selected leaf was wrapped with a moist cloth and secured using cling wrap to minimize further transpiration of the shoot which would alter the resultant pressure reading. Thereafter, the leaf petiole was cut using a surgical blade; a uniform length of 5 cm per cut petiole was maintained. The petiole was then quickly placed through the chamber lid and secured tightly with the excised edge of the petiole facing outside and the bagged leaf inside the chamber. The chamber was sealed and then slowly pressurized with nitrogen gas at 10 kPA-s-1. During pressurisation, a x16 magnifying glass was used to carefully observe the excised surface of the petiole for the appearance of a drop of water (sap). As soon as the drop appeared the corresponding pressure on the chamber gauge was recorded and leaf water potential was expressed as the negative of the corresponding pressure. For MSWP the same procedure as above was followed except that the targeted leaves were covered with a static shield bag (Soilmoisture Equipment Corp, USA) 2 h prior to measurements being taken.
Metabolic responses
In order to determine metabolic responses of bambara groundnut during soil water depletion, starting from 21 DAP irrigation was withdrawn from all trays except the control. Plant leaf tissue was sampled destructively daily at midday for laboratory analysis of plant metabolites associated with stress acclimation (total antioxidant capacity, total phenolics and total sugars).
Determination of total antioxidant capacity
Total antioxidant capacity in leaf tissue of bambara groundnut landrace selections was determined using the ferric reducing ability of plasma (FRAP) assay as described by Benzie and Strain (1996). 0.1 g of ground freeze-dried leaf tissue was mixed with 5 ml 1N perchloric acid and homogenized using an Ultra-Turrax (Model T25D, IKA, Germany) for 30 s. The homogenate was centrifuged at 5 000 g for 10 min at 4°C using a Sorvall RC 5C centrifuge (Sorvall, Newtown, CT, USA). 900 μℓ freshly-prepared FRAP reagent was pipetted into cuvettes. 30 μℓ of sample solution was added to the 900 μℓ FRAP and mixed. Cuvettes were incubated for 10 min making sure that the contents of each cuvette reacts for the same period of time. Absorbance was then recorded at 593 nm using a UV-1800 UV-Vis spectrophotometer (Shimadzu, North America). Concentration of FESO4 was in turn plotted against concentrations of the standard antioxidants.
Determination of total phenolics content
Total phenolics in leaf tissue of bambara groundnut landrace selections was determined using the method of Pérez-Conesa et al. (2009). 0.1 g sample of freeze-dried leaf tissue was mixed with 1 mℓ of 1 M HCL, vortexed for 1 min and incubated at 37°C for 30 min. After incubation, 1 mℓ NaOH (2 M in 75% methanol) solution was used for alkaline hydrolysis and the resulting mixture vortexed for 1 min and incubated at 37°C for 30 min. The samples were vortexed before mixing with 1.0 mℓ of 0.75 M metaphosphoric acid and centrifuged at 5 000 r-min-1 (2 510) for 10 min. The supernatants were collected, transferred into a 10 ml volumetric flask and the pellets re-suspended in 1.0 mℓ of acetone:water (1:1, v/v), vortexed for 1 min and centrifuged at 5 000 r.min-1 (2 510) for 10 min. Both extracts were combined and made up to 10 mℓ with acetone:water (1:1, v/v). Total phenolics were determined using Folin-Ciocalteu reagent. Briefly, 5 mℓ of nanopure water, 1 ml of sample and 1 ml of Folin-Ciocalteu reagent were added to a 25 mℓ volumetric flask and allowed to stand for 5-8 min at room temperature. Thereafter, 10 mℓ of a 7% sodium carbonate solution was added, followed by the addition of 8 mℓ of nanopure water placed to volume. The solution was vortexed thoroughly and allowed to stand at room temperature for 2 h before being filtered through a Whatman 0.45 μm polyfilter prior to determination of total phenolics at 750 nm absorbance using a UV-1800 UV-Vis spectrophotometer (Shimadzu, North America). Gallic acid monohydrate was used as the standard to prepare the calibration curve. The results were expressed as milligrams of gallic acid equivalents (GAE) per gram DM of leaf tissue.
Total soluble sugars determination
For total soluble sugars determination 5 g of freeze-dried plant sample was sent to Cedara Feed Laboratory, Pietermaritzburg. Samples taken at 21 (the last day of watering), 24 (3 days after withdrawing irrigation) and 27 DAP (6th day after withdrawing irrigation and day of termination) were submitted for analysis.
Seedling water use efficiency
The amount of water used (WU) to establish seedlings across the different watering regimes was converted from ml to mm (depth) using conversion factors described by Allen et al. (1998) where:

At the end of seedling establishment (21 DAP), seedling bio-mass was also determined. Using these measurements, seedling water use efficiency was then determined as follows:
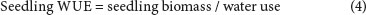
where: WUE = water-use efficiency in g-mm-1, biomass = seedling biomass (above and below ground) in g, water use = amount of water (in mm) applied to the respective seedling trays.
Statistical analyses
Data collected were subjected to analysis of variance (ANOVA) using GenStat Version 14 (VSN International, United Kingdom) at the 5% level of significance. Means of significantly different variables were separated using Duncan's test in GenStat at the 5% level of significance.
RESULTS
Seedling establishment
Emergence
Figure 2 represents total emergence of landrace selections as all trays were watered to 75% FC to ensure maximum emergence (hence watering regime was not a factor affecting differential emergence). Results of seedling emergence showed highly significant differences (P<0.001) among landrace selections, time (DAP) and the interaction between the two. The highest (80%) and lowest (63%) final emergence was observed in the plain red and black-speckled landrace selections, respectively. The brown-speckled and plain-cream landrace selections had a final emergence of 73% and 67%, respectively. All landrace selections started emerging at 5 DAP. Results of mean time to emergence showed no differences (P>0.05) among landrace selections.

Growth and physiology
Leaf number showed no significant differences (P>0.05) among landrace selections. The differences among watering regimes and time (DAP) were highly significant (P<0.001). The interaction among landrace selections and watering regimes over time was not significant (P>0.05). At the end of the 21-day seedling establishment experiment, the highest leaf number (4) was observed in the 75% FC treatment while the lowest number of leaves (3) was observed in the 25% FC treatment.
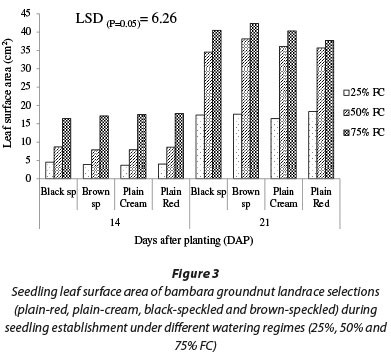
Results for seedling leaf surface area were synonymous with results for leaf number (Fig. 3). Generally, increasing water availability from 25% FC to 50% FC increased leaf surface area by 109%; further increasing water availability to 75% FC increased leaf surface area by 27%. Across all watering regimes and landrace selections, leaf surface area increased with time and the highest leaf surface area (42.31 cm2) was observed in the non-stressed (75% FC) brown-speckled landrace selection, while the lowest (16.43 cm2) was observed in the severely stressed (25% FC) plain-cream landrace selection.
There were no significant differences (P>0.05) among landrace selections with respect to plant height. The interaction between watering regimes and time (DAP) as well as the interaction among landrace selections and watering regimes over time (DAP) were however highly significant (P<0.001). At the end of the experiment, the tallest seedlings (~17 cm) were observed in the 75% FC treatment while the shortest seedlings (~14 cm and ~11 cm) were observed in the moderately (50% FC) and severely stressed (25% FC) watering regimes, respectively. Landrace selections behaved differently within the watering regimes but in all watering regimes plant height increased with time.
Results for chlorophyll content index showed no significant differences (P<0.05) among landrace selections, watering regimes, time (DAP) and the interaction among the three. At 21 DAP, the plain-red and brown-speckled landrace selections had the highest and lowest CCI, respectively, across all water regimes. Chlorophyll content index did not vary significantly over time.
Chlorophyll fluorescence varied significantly (P<0.05) among landrace selections (Fig 4). Watering regimes showed no significant differences (P>0.05) (Fig 4). The interaction between landrace selections and watering regimes over time was not significant (P>0.05) (Fig 4). Based on means of landrace selections, CF was, respectively, highest and lowest in the plain-red (0.81) and plain-cream landrace selections (0.80). Results over time (DAP) showed that during the first week CF was 0.80 and then peaked at 0.81 14 DAP, only to decrease to 0.79 at 21 DAP.

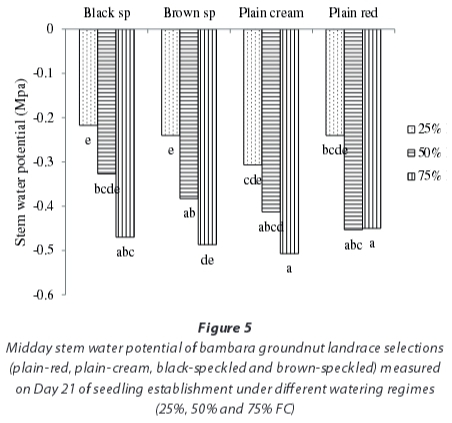
With respect to pre-dawn leaf water potential, no significant differences (P>0.001) were observed among landrace selections, watering regimes and the interaction between the two. Results for midday stem water potential also showed no significant differences (P>0.05) among landrace selections (Fig 5), but among watering regimes were, however, highly significantly different (P<0.001) (Fig 5). The interaction between landrace selections and watering regimes was not significant (P>0.05) (Fig 5). Midday stem water potential was close to zero (-0.286 MPa) in the water-stressed watering regime (25% FC) and more negative (-0.44 MPa) in the no-stress watering regime (75% FC). For the moderately-stressed watering regime (50%) FC, though slightly lower (-0.42 MPa), this was statistically similar to the no-stress watering regime (75% FC).
Seedling parameters
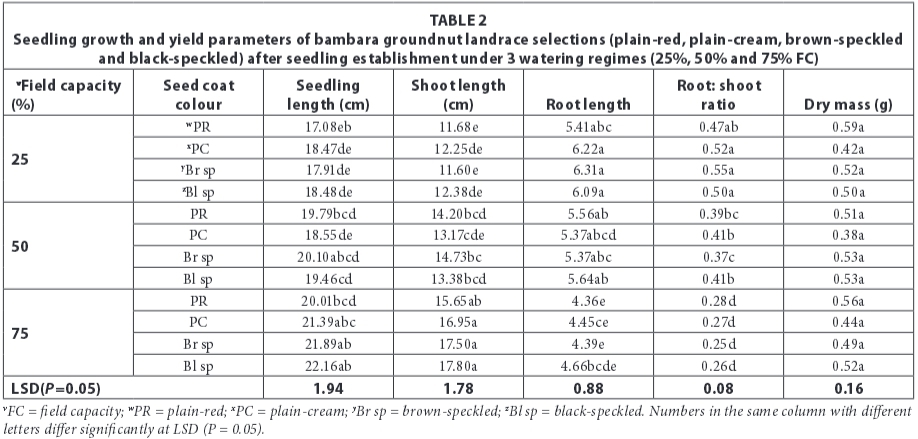
A consistent pattern was observed in results for seedling growth parameters (seedling length, root and shoot length and root: shoot ratio). Landrace selections were not significantly different (P>0.05), watering regimes were highly significantly different (P<0.001) and the interaction between landrace selections and watering regimes was not significant (P>0.05) (Table 2). Seedling length was longest (21.36 cm) in the no-stress watering regime (75% FC) and shortest (17.98 cm) in the stressed water regime (25% FC). Shoot lengths also followed a similar pattern where longest shoots (16.90 cm) were observed in the no-stress water treatment (75% FC) and the shortest shoots observed in the stressed water regime (25 % FC). Results for root length were the inverse of seedling and shoot lengths, whereby the longest roots (6 cm) were observed in the stressed water regime (25% FC) and the shortest roots (4.50 cm) observed in the no-stress water regime (75% FC). Root length in the moderately-stressed watering regime (50%) was 5.50 cm. Consequently, the trend in root:shoot ratio was such that 25% FC (0.5) > 50% FC (0.35) > 75% FC (0.27). Results for seedling dry mass showed no significant differences (P>0.001) among landrace selections, watering regimes and the interaction between the two (Table 2).
Irrigation withdrawal
Physiological responses
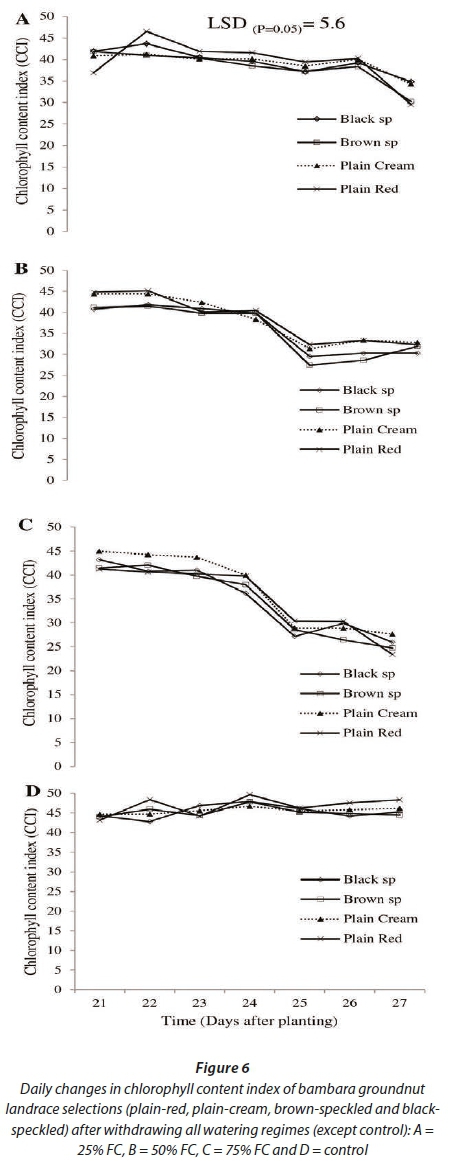
Following withdrawal of irrigation, results for CCI showed significant differences (P<0.05) among landrace selections, while highly significant differences (P<0.001) were observed among watering regimes and time (Fig 6). However, the interaction among lan-drace selections and watering regimes over time (DAP) was not significant (P>0.05) (Fig. 6). Results for CCI showed huge variability among landrace selections across all watering regimes. The plain-cream and brown-speckled landrace selections respectively had the highest (39.98) and lowest (38.40) CCI. A significant decline in CCI (~42 - ~28) was first observed in the previously no-stress water regime (75% FC) 3 days after withdrawing irrigation. In the previously moderately-stressed water regime (50% FC), a significant reduction in CCI (~42 - ~30) was observed 4 days after withdrawing irrigation. In the previously severely-stressed (25% FC) water regime a significant decrease in CCI (~40 - ~32) was observed 6 days after withdrawing irrigation. The control treatment maintained the highest (> 40) CCI, compared to the treatments where irrigation was withdrawn.
After withdrawing irrigation, results for chlorophyll fluorescence showed highly significant differences (P<0.001) among landrace selections, watering regimes, time (DAP) and the interaction of the three (Fig 7). Comparing means for landrace selections, the highest (0.71) and lowest (0.69) CF was observed in the plain-red and black-speckled landrace selections, respectively. Chlorophyll fluorescence was relatively constant in the control treatment while it decreased significantly (from ~0.80 -0.57) in he previously no-stress (75% FC) water regime 3 days after withdrawing irrigation. At the end of the experiment (7 days after withdrawing irrigation) chlorophyll fluorescence had decreased to -0.48 in the previously severely-stressed water regime (25% FC), ~0.5 0 in the ρreviously moderately-stressed water regime and ~0.35 in the previously no-stress water regime (75% FC).

Metabolic responses
Results for TAOC showed highly significant differences (P<0.001) among watering regimes over time (DAP) (Fig. 8). Landrace selections showed no significant (P>0.05) variation and neither did the interaction among watering regimes and landrace selections over time (DAP) (Fig. 8). For all treatments, TAOC increased following withdrawal of irrigation. Total antioxidant capacity reached its peak on different days across the watering regimes. The previously not-stressed (75% FC) and moderately-stressed water regimes (50% FC) attained peak TAOC (~0.90 mg-g-1 DW) at 24 DAP, corresponding to 3 days after withdrawing irrigation. The previously severely-stressed water regime (25% FC) reached peak TAOC (~0.80 mg-g-1 DW) 25 DAP (4 days after withdrawing irrigation). The control treatment did not show much fluctuation for the duration of the experiment; it was constant between 0.20 mg-g-1 DW and 0.30 mg.g-1 DW.
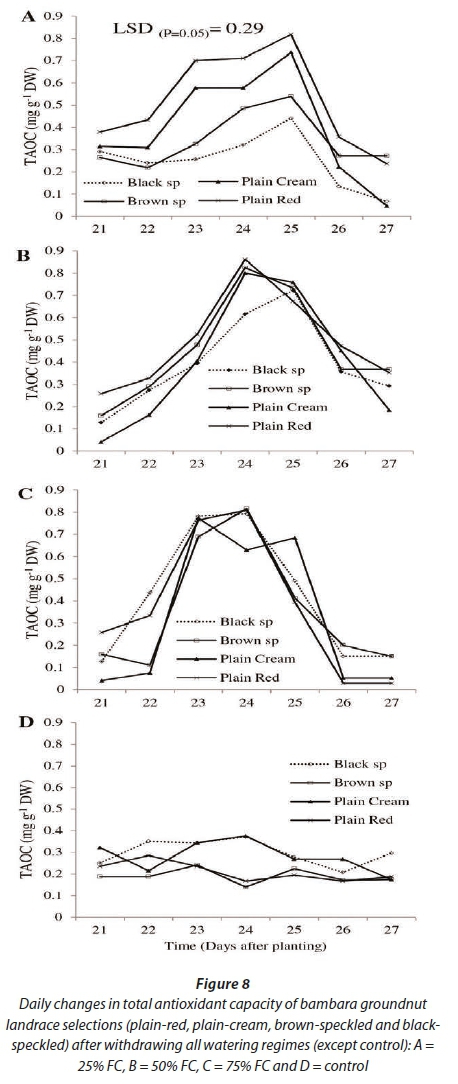
It is worth noting that at 21 DAP TAOC was different across watering regimes; the trend for TAOC was 25% FC > 50% FC > 75% FC. With respect to landrace selections, generally plain-red had the highest TAOC under moderate and severe stress while the plain-cream landrace selection had the lowest. Differences among landrace selections were more pronounced in the previously severely-stressed watering regime (25%), with plain-red landrace selection being superior and black-speckled landrace selection being inferior in this regard.
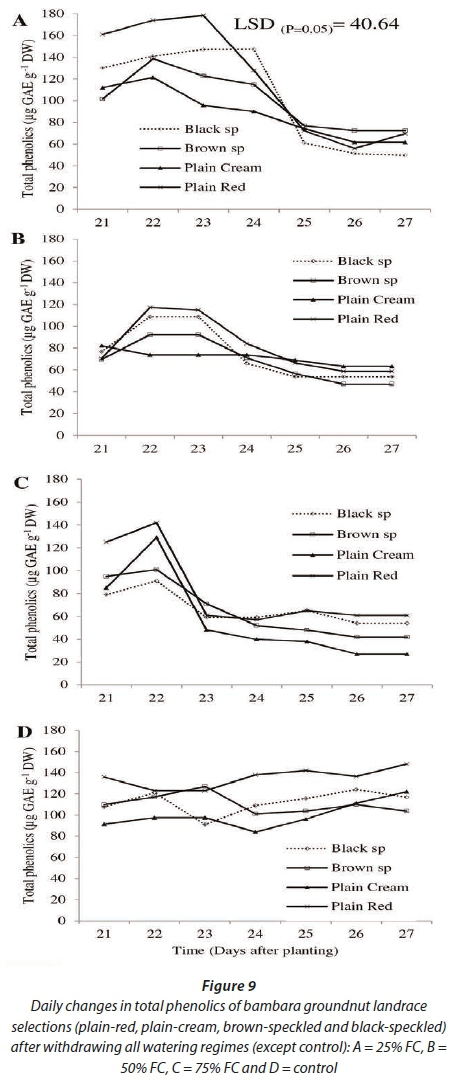
Total phenolics varied significantly (P<0.001) among water regimes, landrace selections, time (DAP) and the interaction of the three (Fig. 9). Across all water regimes, a noticeable decline in phenolics was observed at 23 and 24 DAP (2 and 3 days after withdrawing irrigation), although the patterns of decline were different. In the previously severely-stressed water regime (25% FC), the decline was gradual. A sharp decline was observed at 23 DAP (2 days after withdrawing irrigation) in the previously no-stress water regime (75% FC). In the control treatment, total phenolics were constant between 80 and 130 μg GAE-g-1 DW. At 21 DAP total phenolics were higher (~120 μg GAE-g-1 DW) in the severely-stressed watering regime (25% FC), compared to ~80 μg GAE-g-1 DW in the moderately-stressed watering regime (50% FC) and 100 μg GAE-g-1 DW in the no-stress watering regime (75% FC). Similar to TAOC, differences among landrace selections were more pronounced in the previously severely stressed water regime (25% FC). Again, landrace selections exhibited huge variability across water regimes, except for the plain-red landrace selection which showed consistently high total phenolics across all watering regimes.
There were highly significant differences (P<0.001) among watering regimes and time (DAP), with respect to total sugars (Fig. 10). Total sugars among landrace selections were not significantly different (P>0.05) and neither was the interaction among watering regimes and landrace selections over time (DAP) (Fig. 10). When irrigation was withdrawn, total sugars declined (~40% - ~5%) in the previously severely-stressed (25% FC) and moderately-stressed water regimes (50% FC). In the previously no-stress water regime (75% FC), total sugars went up from -15 % - ~30% during the period 21 to 24 DAP (3 days after withdrawing irrigation), and then declined from -30% - -3% by 28 DAP. Total sugars in the control treatment did not show much variation for the duration of the experiment (Fig. 10).

Seedling water use efficiency
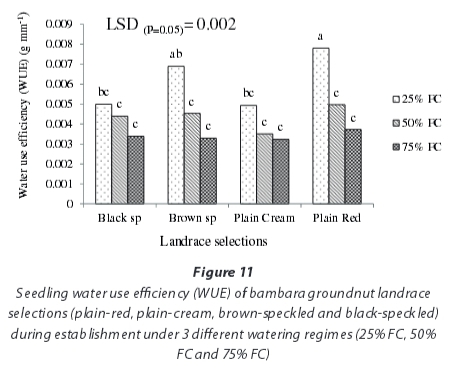
Seedling water use efficiency showed significant (P<0.05) variation among water regimes and landrace regimes. The interaction between water regimes and landrace selections was, however, not significant (P>0.05) (Fig. 11). Seedling water use efficiency was highest in the severely-stressed (25% FC) water regime (0.006 g-mm-1) and lowest in the non-stressed (75% FC) watering regime (0.003 g-mm-1) while it was 0.004 g-mm 1 in the moderately-stressed (50% FC) watering regime. All landrace selections showed improved WUE in response to decreasing water availability. At 25% FC, seedling WUE was statistically different to that at 50% FC and 75% FC; seedling WUE at 50% was statistically similar to that at 75% FC (Fig. 11). Based on means of landrace selections across water regimes, the trend in seedling WUE were such that plain-red (0.006 g mm-1) > brown-speckled (0.005 g-mm-1) > black-speckled (0.00427 g-mm-1) > plain-cream landrace selection (0.004 g-mm-1).
DISCUSSION
The objective of this study was to determine growth, physiology and water use characteristics of a bambara groundnut landrace differing in seed coat colour, during establishment under different water regimes. A secondary objective was to determine physiological and metabolic responses of bambara groundnut to terminal water stress.
With respect to seedling final emergence, results of this study concurred with the findings of Pillay (2003); Sinefu (2011) and Zondi (2013) where dark-coloured seeds had better emergence (80%) compared to the light-coloured seeds (63%). This was related to the tannins present in dark-coloured seeds which are polyphenols and act as antioxidants under stress conditions (Mabhaudhi and Modi, 2013). This study went further to determine the amount of phenolics present in leaf tissue of the different bambara groundnut landraces. True to expectation, high phenolic content and high total antioxidant capacity were observed in the plain-red landrace selection. Our observations of high phenolic content in the plain-red landrace selection concur with Mabhaudhi and Modi's (2013) hypothesis. This implies that the use of seed colour as a seed selection criterion has merit. Final emergence (> 63%) observed in this study was relatively higher compared to that reported by Legwaila et al. (2013) of < 42%. These differences may be attributed to different landraces used in the two studies; landraces originating from different locations typically show huge variability (Zondi, 2013). However, what is evident from both studies is the fact that bambara groundnut establishes poorly. This reduces crop water use efficiency as more water is lost through evapotranspiration during this period with minimal crop biomass gain. Mabhaudhi and Modi (2013) suggested this as a possible reason for low uptake of the crop by farmers and recommended that strategies to improve emergence should be explored. Improving seedling emergence will improve crop water use efficiency. In their study, Legwaila et al. (2013) managed to improve emergence in bambara groundnut landraces by hydro-priming them.
Water is vital to plant growth and development. It drives the turgor process responsible for cell division and cell expansion. Under water-limited conditions plant growth and development are compromised. Studies on the effect of water stress on bambara groundnut seedling growth have shown that seedling growth was affected by water availability; growth rate increased with increasing water availability (Mwale et al., 2007; Sinefu, 2011; Zondi, 2013). The results of this study confirmed those of these previous studies. However, our study also showed that physiological responses such as CCI and CF were not very sensitive to water stress. This behaviour suggests that reported drought avoidance in bambara groundnut landraces is also present at the seedling stage. Bambara groundnut seedlings were able to reduce plant growth (leaf number, leaf surface area and plant height) in order to maintain high tissue water potential under stress (Blum, 1996; 2005). Results of leaf water potential confirmed this as there were no significant differences observed across the water regimes with respect PLWP and MSWP. The results for total sugars was such that 25% FC > 50% FC > 75% FC at the end of the seedling establishment. It could be assumed that accumulation of TSS under water stress facilitated retention of water, protected photosynthetic apparatus and maintained ion homeostasis in stressed seedlings, contributing to high tissue water potential (Bohnert and Jensen, 1996). Results of MSWP could be related to the longer roots observed in the stressed treatments compared to the no-stress treatments, implying that there was enhanced soil water capture in the stressed treatment. The higher root:shoot ratios observed in the stressed water treatments suggests that the plant favoured root growth to shoot growth in order to manipulate the source and sink strength and hence utilize resources efficiently (Yordanov et al., 2000; Chaves et al., 2002).
Seedling WUE is defined as a ratio of biomass accumulation, expressed in total seedling biomass (g), to water added in mm. High WUE is largely a function of reduced water use and net improvement in plant production (Blum, 2005). This was the case in this study as results for the enumerator (total dry mass) were not statistically different among landrace selections and watering regimes. The differences in WUE were influenced by the denominator (water use), with WUE increasing with decreasing water supply. The high WUE could be attributed to the drought avoidance strategies (reduced leaf number, leaf surface area and plant height and sugars accumulation) observed in the stressed seedlings. Mabhaudhi et al. (2013) observed a similar behaviour when they determined whole WUE for the same species. With respect to landrace selections, WUE was higher in the plain-red landrace selection. This lends further credence to the hypothesis of Mabhaudhi et al. (2013) that dark-coloured seeds performed better and exhibited enhanced drought tolerance compared to light-coloured seeds.
Lastly, we determined the physiological and metabolic responses of bambara groundnut landrace selections to terminal stress following withdrawal of watering. Generally, the previously severely-stressed plants responded better to terminal stress, both physiologically and metabolically, compared to the previously non-stressed plants. This was as a result of 'acclimation' where plant adaption to any stress situation is mediated by defences. The effectiveness of these defences can be enhanced by pre-exposing plants to specific mild stimuli before imposing the full strength of the stress. This was the case for plants that had previously been subjected to 25% FC. Acclimation to water stress has been shown in Arabidopsis, where plants were able to alleviate the effect of water stress due to prior exposure to water stress at an early stage (Harb et al., 2012).
Maxwell and Johnson (2000) advanced that the ability of the plant to tolerate environmental stresses and the extent to which these stresses have damaged photosynthetic apparatus can be deduced from measuring CF. Based on the latter, our results indicate that the previously moderately- and severely-stressed plants were better able to tolerate terminal stress compared to the previously not-stressed plants. This supports the theory of acclimation to water stress. Our findings also support the assertion by Khaleghi et al. (2012) that CCI and CF could be used to evaluate the photosynthetic process under water stress and thus as a rapid technique for detecting plant tolerance to drought stress.
CONCLUSIONS
Major findings of this study are: (i) dark-coloured seeds performed better than light-coloured seeds, especially under stress conditions, (ii) drought avoidance strategies and, (iii) acclimation to water stress were present in bambara groundnut seedlings. Improved seedling WUE under water-limited conditions was associated with smaller seedling canopy size, longer roots, and higher root: shoot ratio as well as accumulation of solutes which allowed for maintenance of high tissue water potential. The red landrace selection may be recommended due to its high emergence rate and ability to tolerate drought. Future studies should determine WUE of bambara groundnut propagated using seedlings under field conditions so as to determine whether transplanting improves WUE.
ACKNOWLEDGEMENTS
The Water Research Commission of South Africa is acknowledged for initiating, funding, managing and directing this study through WRC Project no. K5/2274/4 'Determining water use of indigenous grain and legume food crops'.
REFERENCES
ALLEN RG, PEREIRA LS, RAES D and SMITH M (1998) Crop evapotranspiration: Guidelines for computing crop water requirements. FAO Irrigation and Drainage Paper no. 56. FAO, Rome. [ Links ]
AZAM-ALI SN, KHONGA E, WENZEL G, FLESSNER K and SESAY A (2004) Increasing the productivity of bambara groundnut (Vigna subterrânea (L.) Verdc.) for sustainable food production in semi-arid Africa. BAMFOOD Project Final Report. University of Nottingham, Nottingham. [ Links ]
BEWLEY JD and BLACK M (1994) Seeds: Physiology of Development and Germination. Plenum, New York. [ Links ]
BENZIE IFF and STRAIN JJ (1996) The ferric reducing ability of plasma (FRAP) as a measure of 'antioxidant power'. Anal. Biochem. 239 70-76. [ Links ]
BLUM A (1996) Crop responses to drought and the interpretation of adaptation. Plant Growth Regul. 20 135-148. [ Links ]
BLUM A (2005) Drought resistance, water use efficiency and yield potential- are they compatible, dissonant or mutually exclusive? Aust. J. Agric. Res. 56 1159-1168. [ Links ]
BOHNERT HJ and JENSEN RG (1996) Strategies for engineering water stress tolerance in plants. Trends Biotechnol. 14 89-97. [ Links ]
CHAVES MM, PEREIRA JS, MOROCO J, RODRIGUES ML, RICARDO CPP, OSORIO ML, CARVALHO L, FARIA T and PINHEIRO C (2002) How plants cope with stress in the field. Photosynth. Growth. Ann. Bot. 89 907-916. [ Links ]
CHIBARABADA TP, MODI AT and MABHAUDHI T (2014) Seed quality characteristics of a bambara groundnut (Vigna subterranea L.) landrace differing in seed coat colour. S. Afr. J. Plant & Soil 31 219-226. [ Links ]
CHONE X, VAN LEEUWEN C, DUBOURDIEU D and GAUDILLERE JP (2001) Stem water potential is a sensitive indicator for grapevine water status. Ann. Bot. 7 477-483. [ Links ]
HARB A, KRISHNAN A, AMBAVARAM MMR and PEREIRA A (2010) Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 154 1254-1271. [ Links ]
JORGENSEN ST, LIU F, OUEDRAOGO M, NTUNDU WH, SARRAZIN J and CHRISTIANSEN JL (2010) Drought responses of two bambara groundnut (Vigna subterranea L. Verdc.) landraces collected from a dry and humid area of Africa. J. Agron. Crop Sci. 196 412-422. [ Links ]
KHALEGHI E, ARZANI K, MOALLEMI N and BARZEGAR M (2012) Evaluation of chlorophyll content and chlorophyll fluorescence parameters and relationships between chlorophyll a, b and chlorophyll content index under water stress in Olea europaea cv. Dezful. World Acad. Sci. Eng. Technol. 68 1154-1157. [ Links ]
LEGWAILA GM, KARIKARI SK, MOGAMISI K and MATHOWA T (2013) Response of three bambara groundnut landraces to presowing hydration in Botswana. Agric. Biol. J. N. Am. 4 430-434. [ Links ]
MABHAUDHI T (2009) Responses of landrace maize (Zea mays L.) to water stress compared with commercial hybrids. MSc Thesis, University of KwaZulu-Natal, South Africa. [ Links ]
MABHAUDHI T and MODI AT (2010) Early establishment performance of local and hybrid maize under two water stress regimes. S. Afr. J. Plant Soil 27 299-304. [ Links ]
MABHAUDHI T (2012) Drought tolerance and water use of selected South Africa landraces of Taro (Colocasia Esculenta L. Schott) and bambara groundnut (Vigna subterranea L. Verdc). PhD thesis, University of KwaZulu-Natal, South Africa. [ Links ]
MABHAUDHI T and MODI AT (2013) Growth, phenological and yield responses of a bambara groundnut (Vigna subterranea L. Verdc) landrace to imposed water stress under field conditions. S. Afr. J. Plant Soil 30 69-79. [ Links ]
MABHAUDHI T, MODI AT and BELETSE YG (2013) Growth, pheno-logical and yield responses of a bambara groundnut (Vigna subter-ranea L. Verdc) landrace to imposed water stress: ll. Rain shelter conditions. Water SA 39 191-198. [ Links ]
MAKANDA I, TONGOONA P, MADAMBA R, ICISHAHAYO D and DERERA J (2009) Evaluation of bambara groundnut varieties for off season production in Zimbabwe. Afr. Crop Sci. J. 16 175-183. [ Links ]
MAXWELL K and JOHNSON GN (2000) Chlorophyll fluorescence - a practical guide. J. Exp. Bot. 51 659-668. [ Links ]
MCCUTCHAN H and SHACKEL KA (1992) Stem water potential as a sensitive indicator of water stress in prune trees (Prunus domestica L. cv. French). J. Am. Soc. Hortic. Sci. 117 607-611. [ Links ]
MODI AT (2007) Growth temperature and plant age influence on nutritional quality of Amaranthus leaves and seed germination capacity. Water SA 33 369-371. [ Links ]
MODI AT and MABHAUDHI T (2013) Water use and drought tolerance of selected traditional and indigenous crops. WRC Report No. 1771/1/13, ISBN 978-1-4312-0434-2. Water Research Commission, Pretoria. [ Links ]
MWALE SS, AZAM-ALI SN and MASSAWE FJ (2007) Growth and development of bambara groundnut (Vigna subterranea) in response to soil moisture 1. Dry matter and yield. Eur. J. Agron. 26 345-353. [ Links ]
NAGESWARA RAO RC, WILLIAMS JH, WADIA KDR, HUBICK KT and FARQUHAR GD (1993) Crop growth, water use efficiency and carbon isotope discrimination in groundnut (Arachis hypogaea L.) genotypes under end of season drought conditions. Ann. Appl. Biol 122 357-367. [ Links ]
NAZARLI H and FARAJI F (2011) Response of proline, soluble sugars and antioxidant enzymes in wheat (Triticum aestivum L.) to different irrigation regimes in greenhouse condition. Cerietari Agron. Moldova 148 27-33. [ Links ]
PILLAY D (2003) Physiological and biochemical characterisation of four South African varieties of bambara groundnut (Vigna subterranea (L.) Verdc). MSc Thesis, International Master Programme at the Swedish Biodiversity Centre. [ Links ]
SINEFU F (2011) Bambara groundnut response to controlled environment and planting date associated water stress. MSc Thesis, University of KwaZulu-Natal, South Africa. [ Links ]
SONGSRI P, JOGLOY S, JUNJITTAKARN J, KESMALA T, VORASOOT N, HOLBROOK CC and PANANOTHAI A (2013) Association of stomatal conductance and root distribution with water use efficiency of peanuts under different soil water regimes. Aust. J. Crop Sci. 7 948-955. [ Links ]
TEKRONY DM and EGLI D (1991) Relation of seed vigour to crop yield: A review. Crop Sci. 31 816-822. [ Links ]
VURAYAI R, EMONGOR V and MOSEKI B (2011) Effect of water stress imposed at different growth and developmental stages on morphological traits and yield of bambara groundnuts (Vigna subterranea L. Verdc). Am. J. Plant Physiol. 6 17-27. [ Links ]
WILLIAMS LE and ARAUJO F (2002) Correlations among predawn leaf, midday leaf and midday stem water potential and their correlations with other measures of soil and plant water status in Vitis vinifera (L.). J. Am. Soc. Hortic. Sci. 127 448-454. [ Links ]
YORDANOV I, VELIKOVA V and TSONEV T (2000) Plant responses to drought, acclimation and stress tolerance. Photosynthetica 38 186. [ Links ]
ZONDI LZ (2013) Responses of bambara groundnut (Vigna subterranea L. Verdc) landraces to field and controlled environment conditions of water stress. MSc Thesis, University of KwaZulu-Natal, South Africa. [ Links ]
Received 25 May 2014
Accepted in revised form 3 June 2015
* To whom all correspondence should be addressed. +27 33 260 5442; Fax: +27 33 260 6094; e-mail: tendaipolite@gmail.com


{kind=link}
{kind=link}
{kind=link}