










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkWater SA
versión On-line ISSN 1816-7950
versión impresa ISSN 0378-4738
Water SA vol.39 no.1 Pretoria ene. 2013
First results on bathymetry, stratification and physicochemical limnology of a small tropical African reservoir (Malilangwe, Zimbabwe)
Tatenda DaluI, II, *; Zane ThackerayIII; Rio LeuciIII; Bruce CleggIV; Lenin Dzibakwe ChariI, II; Tamuka NhiwatiwaI
IDepartment of Biological Sciences, University of Zimbabwe, PO Box MP167, Mt. Pleasant Harare, Zimbabwe
IIDepartment of Zoology and Entomology, Rhodes University, PO Box 94, Grahamstown 6140, South Africa
IIIEnvironmental Mapping and Surveying, PO Box 201155, Durban North, KwaZulu-Natal 4016, South Africa
IVMalilangwe Wildlife Reserve, Private Bag 7085, Chiredzi, Zimbabwe
ABSTRACT
The study provides a 9-month record of Malilangwe Reservoir water chemistry periodicity, for the period between February and October 2011. Malilangwe Reservoir is a small (211 ha), shallow (mean depth 4.54 m) reservoir situated in the southeastern lowveld of Zimbabwe. The reservoir has not spilled in nearly 11 years, which makes it a unique system as most reservoirs of comparable size spill annually. This is the first bathymetric and limnological study of the reservoir where the morphology and physicochemical quality of the water body were examined. The reservoir was not strongly stratified during the hot-wet and hot-dry season with oxygen depletion of < 2 mg£-1 DO being observed in the bottom layers (<6 m depth). Nutrient concentrations varied throughout the seasons. The reservoir exhibited marked seasonal fluctuations in water level, which decreased by over 149 cm between February and October. The N:P ratio rose to as high as 10.9 and generally reflected high levels of phosphorus in the reservoir. There were significant differences (p<0.05) in Secchi depth transparency between the study sites. Differences observed in water quality were due to water level fluctuations, with poor water quality conditions being experienced during the hot-dry season and the cool-dry season when water levels were low. The reservoir was classified as being mesotrophic. Therefore, there is a risk of eutrophication, especially since the reservoir is currently merely a sink for nutrients.
Keywords: Bathymetry, limnology, water level fluctuation, stratification, Malilangwe
Introduction
Limnology in the tropics has only recently developed past the stage of exploration (Lewis, 2000). Despite the large number of small storage reservoirs and lakes, baseline limnological information is available for relatively few of these (Hart, 1999; Nhiwatiwa and Marshall, 2007; Quarcoopome et al., 2008). The baseline limnological information that exists on small reservoirs and lakes is often insufficient, and is often based on once-off or short term studies, while the limnology of large and medium-sized African reservoirs has been extensively studied (Moss and Moss, 1969; McLachlan, 1974; Dorgeoloh et al., 1993; Mustapha, 2008). Seasonally comprehensive data are seldom available for a complete year, let alone spanning sufficient time to encompass the inherent hydrological variability of the region (Hart, 1999). There is a large density of small reservoirs in most parts of the continent, with Zimbabwe having about 14 000 small reservoirs (Nhiwatiwa and Marshall, 2007), with various impacts affecting these aquatic ecosystems. These impacts include changing seasonal flow patterns of streams and rivers, which in turn affect aquatic species migration (Jackson and Marmulla, 2001; McCartney and Sally, 2005), and water level fluctuations which influence physicochemical characteristics of reservoirs and lakes (Nhiwatiwa and Marshall, 2007).
Water quality deterioration in reservoirs and lakes usually comes from excessive nutrient inputs and organic pollution causing eutrophication, acidification and heavy metal contamination (Djukic et al., 1994; Mustapha, 2008). The effects of these inputs into the reservoir not only affect the socioeconomic functions of the reservoir negatively, but also contribute to a loss of structural biodiversity in the reservoir. The changes in physicochemical characteristics provide valuable information on water quality, sources of variation, and impacts on the functioning and biodiversity of the reservoirs (Mustapha, 2008).
The majority of reservoirs in tropical Africa are shallow, polymictic and found in regions where evaporation approaches or exceeds precipitation, with water typically being derived from rivers (Mustapha, 2009). These reservoirs are usually filled up with water in the rainy season, but become greatly reduced during the dry season as a result of evaporation, water withdrawal and municipal water usage. Many tropical African reservoirs have short water residence times, small sizes, large watersheds, high shoreline development ratios and large water fluctuations due to seasonal influences. There are problems in managing small reservoirs because most of them were constructed when ecological and environmental considerations were regarded to not be as important as agricultural and commercial production (Mustapha, 2009).
The most important environmental input quantities during daytime in natural shallow water bodies are the incoming short and long wave radiation, air temperature and wind speed (Kastev et al., 2010; Jacobs et al., 1997). Long-wave radiation is absorbed at the top of the water body, along with a large part of the incoming short-wave radiation. During the day this leads to a stable stratification in small natural water bodies. High wind speed may cause a strong forced mixing to occur, which leads to more-or-less isothermal water temperatures, which can cause water bodies to become polymictic. During night-time, the most important forcing function is long-wave radioactive cooling which triggers the formation of a mixing layer, starting from the water-atmosphere interface. Depending on the weather regime, the water temperature behaviour in shallow waters is very dynamic and complex (Jacobs et al., 1997). Given the advances in understanding of the structure and energetics of mixed layers, it is surprising that in tropical lakes even the most fundamental information, such as persistent thermocline depth and stability of stratification, is limited and inadequate (Nhiwatiwa and Marshall, 2006).
Water quality is the sum total of the physicochemical properties of a water body and generally gives information on the nutrient status, productivity and sustainability of a water body. Large-scale blooms of cyanobacteria in Malilangwe Reservoir have raised concerns about water quality. The aim of this study was to investigate the physical and chemical characteristics of the Malilangwe Reservoir, in order to assess water quality and to determine the impacts of water level fluctuations on water quality.
Study area
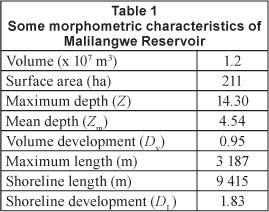
Malilangwe Wildlife Reserve is located in the Chiredzi District (20°58' S, 31°47' E; 21°02' S, 32°01' E) of the south-eastern lowveld of Zimbabwe (Fig. 1). Malilangwe Reservoir was impounded in 1964 and is used for water supply in the reserve. The reservoir is situated on the Nyamasikana River, a tributary of the Chiredzi River which in turn flows into the Runde River.
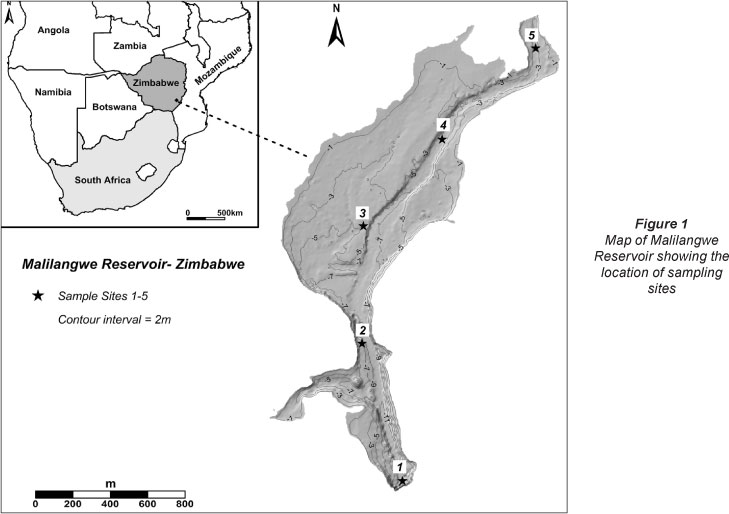
It is a gravity section masonry dam with a surface area of 211 hectares and maximum volume of 1.2 x 107 m3 at full capacity. Some morphometric characteristics of the reservoir are listed in Table 1. Flanked by rocky hills on most sides, the impoundment has a rocky substrate with a few sandy bays. It is poorly vegetated with few marginal plants, including Azolla filiculoides (Lam), Ludwigia stolonifera (Guill and Perr) Raven, Panicum repens (Lam), Schoenoplectus corymbosus (Roth ex Roem and Schult) Raynal, Potamogeton spp., Phragmites mauritianus (Kunth) and Cyperus sp. (Dalu et al., 2012a). The fish communities include predators, omnivores, detritivores, micro-and macrophages (Barson et al., 2008; Dalu et al., 2012b,c).
Methods
Sampling was carried out in the reservoir, throughout 3 seasons (hot-wet, cool-dry and hot-dry) between February and October 2011. All water samples were collected around midday so as to standardise sampling and reduce diurnal biases (Nhiwatiwa, 2004). Five sites were selected along the reservoir length and sampling was done during the last week of each month. A 10 I
Ruttner water sampler was used to collect water samples from the water column, at 1 m intervals from the bottom of the lake to the water surface. The collected water samples were placed in 500 mt polythene bottles and stored in ice before being analysed within 24 h in the laboratory.
Bathymetric survey
The Malilangwe Trust's Harrier bass boat was utilised to undertake the bathymetric survey. The vessel was equipment with a CEESTAR dual-frequency digital survey echo-sounder, 30 kHz and 200 kHz, capable of 0.01% of depth accuracy. The depth sounding positions were fixed using a NOVATel RTK GPS with DGPS capability, which yields real-time horizontal and vertical accuracy of <0.1 m. This accuracy was achieved due to real-time correction information being transmitted via satellite and from various terrestrial base stations. Six soundings were collected per second along predetermined survey lines. These were 25 m apart in the east-west direction and 50 m apart in the north-south orientation. Special manoeuvring had to be undertaken around zones with high tree densities and a few areas close to the dam edge could not surveyed as they were too shallow for the boat to navigate. A land survey of the lake edge allowed for accurate extrapolation of elevation data across these problematic zones.
Side-scan sonar survey
An Imagenex Yellowfin side-scan sonar (260/330/770 kHz) was used to acoustically image the reservoir floor. Survey lines were run parallel to the long axis of the dam and were positioned 35 m apart to produce 100% overlap between successive scan runs, whilst utilising a scan range of 40 m (80 m total swath). This allowed for a comprehensive side-scan sonar mosaic of the dam bottom to be produced. Extremely shallow areas (<1.5 m water depth) and zones with dense submerged trees proved problematic for surveying. Complete coverage of the dam was thus not possible.
Stratification
The seasonal pattern of stratification was determined by measuring temperature and dissolved oxygen concentrations at 1 m intervals using an oxygen meter (LDO HQ20, HACH). Measurements were done once a month throughout the three seasons (hot-wet, cool-dry and hot-dry; February to October), at the deepest point of the reservoir (Site 1). It should be noted that the sampling strategy did not consider spatial variations due to time limitations, hence one site only was selected. The diurnal variations in temperature and dissolved oxygen concentration were determined by taking readings at 2 h intervals over a 24 h period for each of 3 different seasons, February (hot-wet), June (cool-dry) and October (hot-dry).
Basic water quality measurements
Water samples were collected at each site and analysed for pH, conductivity, total dissolved solutes, temperature and dissolved oxygen (DO), using a pH, conductivity and DO meter (HACH, LDO, Germany). Water transparency was measured using a Secchi disk. Chemical oxygen demand (COD), nitrogen, nitrates and total and reactive phosphorus were determined using standard methods from HACH (2007) and Eaton et al. (2005). A Kruskall Wallis test (p<0.05) was carried out to test for statistically significant differences in physicochemical characteristics between sampling stations (H0: no difference between 5 sampling points). The analysis was done for the whole study period, February to October 2011, using SysStat ver. 12 (SYSTAT, 2007).
Results
Bathymetry, side-scan sonar mosaic and reservoir bottom-type classification
The results of the bathymetric survey are presented in Fig. 2. The zero-datum level used in Fig. 2 was the water level at the time of survey (3.76 m below dam spill level). Depth of the reservoir is represented as negative values and in metres. Classification of the reservoir's bottom types was undertaken by the simultaneous interpretation of the side-scan sonar mosaic and bathymetric data. Ten major bottom types, as well as numerous submerged trees and high-relief boulders, were identified (Fig. 3). The overall morphology of the reservoir bottom is dominated by a submerged main channel that extends along the entire main axis of the dam, a central shallow alluvial basin with minor channels and a constricted steep-sided neck area with a secondary tributary channel (entering from the west) near the dam wall.
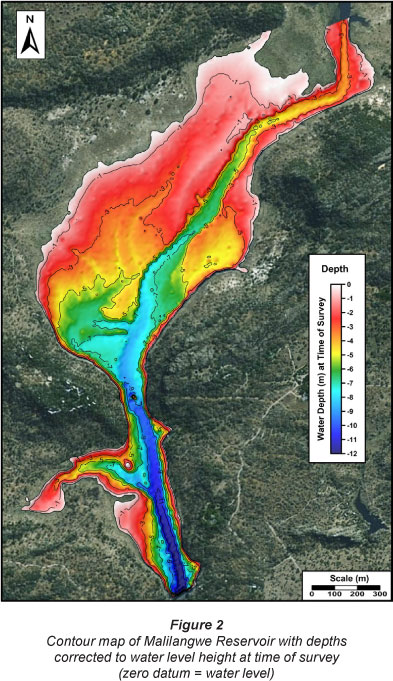
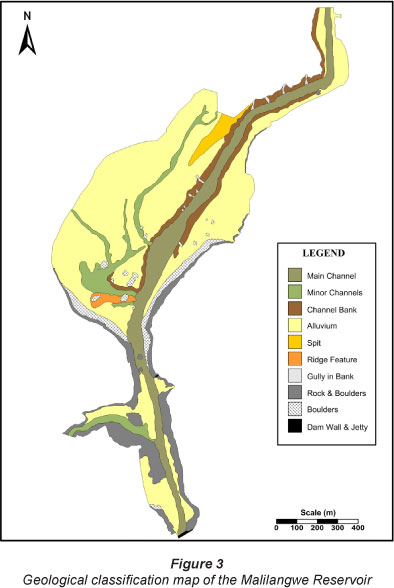
The base of the main channel occurs at a depth of 3 m at the northern limit of the survey area (main tributary input) and extends to a maximum depth of 12 m near the dam wall. This equates to a total elevation drop of 9 m over a horizontal distance of 2 990 m. The main channel incises the central flooded alluvial basin to a depth of 4 m at its northern input and exits the southern part of the basin at a depth of 8 m. The sides of this main channel (banks) are relatively steeply sloped throughout this length, except in the northern and the southern part of the central basin. The shallow central basin is divided by the main channel into an extensive western region, with minor incised channels, and a smaller eastern portion. A split occurs in the northern part of the central basin and is clearly evident at depth of 2 m. During periods of lower water level this split would be emergent. The south-western part of the central basin is punctuated by a minor ridge feature that extends east-west at a depth of 6.0-7.5 m. Its orientation (at a right angle to the normal flow path in the area) and structure appears odd relative to the more sinuous nature of the dam bottom, which is dominated by alluvial/fluvial processes. Origins of the ridge may thus be other than alluvial/fluvial and it is suggested that the ridge may represent an igneous dyke intrusion. Both the south-western and south-eastern margins of the central basin are steep sided, to a depth of 5.5-6.5 m, as a result of being formed by rock exposures. The rock margins extend further north on the eastern side of the central basin (Figs. 2 and 3).
The constricted neck approaching the dam wall forms the deepest part of the impoundment (9-12 m). A secondary, well-developed channel meets the main central channel from the west at a depth of 10.5 m. This channel descends 9 m (with a water depth of 1.5 m at the western limit) over a horizontal distance of 550 m. This area is characterised by steep rocky sides, with the exception of shallow regions at the western extremity of the secondary channel and the southwestern edge between the secondary channel and the dam wall. Within the first main constriction, between the central basin and the neck, a massive rock extends vertically from the dam floor, at a depth of 9 m, to a height of 2.6 m above the water surface. This large rock originated from the surrounding sandstone cliffs. A low-relief rock island (0.7 m maximum height above the water line at the time of survey) was located just north-west of the confluence between the secondary and main channels within this neck. This rock exposure is clearly evident to a depth of 6 m. A second constriction with steep rocky sides and a deep channel occurs at the southern limit of the neck, and this is where the dam wall is situated (Figs. 2 and 3). The reservoir's morphology and bottom composition still reflect the original fluvial and alluvial system that existed prior to the impoundment of this waterway.
Stratification
Seasonal patterns of temperature and dissolved oxygen stratification
Thermal stratification was established during the hot-wet and hot-dry seasons and part of the cool-dry season (June-August) (Fig. 4c). During the hot-wet season (February), the mean water temperature at the water surface was 31.2ºC, but in the cool-dry season (July) the mean water temperature was 20ºC, with a mean bottom water temperature of 26.9ºC and 18.6ºC in February and July, respectively. Dissolved oxygen stratification was similar to that of thermal stratification. Oxygen concentrations dropped to >4 mgt-1 at 5 m depth during the February to March and August to October periods (Fig. 4b). As the air temperature got cooler and the solar radiation input decreased during the cool-dry season, the surface water began to cool, which resulted in surface water and the thermocline cooling down to the temperature of the hypolimnion. The reservoir at this point was not stratified. In this state the reservoir could be easily be mixed, even by light wind, resulting in complete mixing or turnover at the beginning of the cool-dry season. This gave the bottom water an opportunity to become aerated. As the surface water heated again, towards the end of the cool-dry season, the reservoir reached a state of thermal homogeneity and stratification set in, with the reservoir returning to the summer stratification state (Fig. 4c). The changes which occurred were related to air temperature and wind speed. Air temperature increased from 25.2ºC in February to 27.3ºC in March, before decreasing to a low of 16.8ºC in July. Air temperature then increased from July to October, reaching 26.5ºC. Wind speed followed a similar trend as temperature with a value of 1.1 kmh-1 being recorded in February, and decreasing to a low of 0.5 kmh-1 in May and June. Wind speed then increased to a value of 2.3 kmh-1 during October (Fig. 4a).
Diurnal patterns of temperature and dissolved oxygen stratification
Hot-wet season
The first 24-h sampling exercise was carried out on 24 February 2011 (during the hot-wet season) when the water level was high and there was inflow of water into the reservoir. The reservoir was thermally stratified during the day (10:00-18:00) but this stratification broke down at night (20:00-08:00) (Fig. 5c). The thermal stratification was characterised by very small temperature gradients. Stratification followed the air temperature trend, wherein air temperature decreased from 30.8ºC at 16:00 to a low of 20ºC at 04:00, before increasing to a high of 32.8ºC at 16:00 the next day (Fig. 5a). The dissolved oxygen stratification pattern was more pronounced and the concentration of oxygen fell to <2 mgl-1 (at depth = 8 m) from approx. 9 mgl-1 (at depth = 0 m). Oxygen concentrations in the entire water column fell after the breakdown of stratification (22:00-08:00), for a short period (Fig. 5b). Below 5 m depth, the reservoir was anoxic (<4 mgl-1) during the day.
Cool-dry season
The next 24-h sampling exercise was carried out on 15 June 2011 (cool-dry season). The reservoir water level was low as a result of evaporation and drawdown. There was weak thermal stratification in the reservoir during the day (10:00-16:00). Surface water temperatures decreased from a peak of 28ºC during the day (10:00-16:00) to a low of 19ºC during the night (Fig. 6b). Lower air temperatures were observed during the cool-dry season (10-24ºC) compared to February. Low air temperatures of about 10ºC were recorded during the early morning hours (04:00-06:00) (Fig. 6a). Stratification broke down after 16:00 and the water body remained well-mixed until 10:00. There was no oxygen stratification in the reservoir and oxygen concentrations ranged between 8 and 9 mgl-1, indicating that the water was oxygen saturated (Fig. 6b).
Hot-dry season
The last 24-h sampling exercise was carried on 27 October 2011 (during the hot-dry season) when the water was very low and there was no inflow into the reservoir. The reservoir was thermally stratified during the day (11:00-17:00) with stratification breaking down during the night (18:00-10:00). The water was warmer during the day with surface water temperatures in the range of 34-35ºC, and cooler during the night with surface water temperatures ranging from 25-26ºC (Fig. 7c). This pattern was related to air temperatures, which reached 30.4ºC around midday, with lower air temperatures being observed in the early morning (22-24ºC), between 02:00 and 06:00 (Fig. 7a). There was weak oxygen stratification during the same period and very low oxygen concentrations (<3-6 mgl-1) were recorded during the hot-dry season.
Oxygen concentrations were below 4 mgt-1 between 16:00 and 18:00 at the water surface, whilst at 5 m depth the oxygen concentration was < 4 mgt-1 for the rest of the study period (Fig. 7b).
Water chemistry
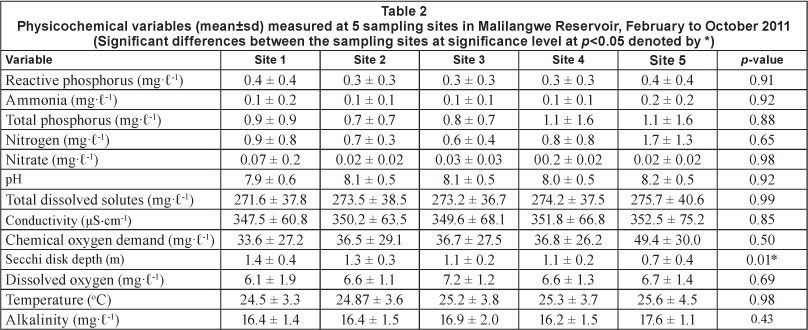
Table 2 summarises the mean values of the environmental variables measured in the Malilangwe Reservoir for the study period. Water levels decreased at an average rate of 18.63 cm per month at all sites, with the greatest decrease being recorded at all sites during March (25 cm) and the smallest decrease being recorded during June (9 cm). From February to October a total of 149 cm of water had been lost from the reservoir due to drawdown and evaporation.
The pH fluctuated during the 9 months of sampling. At the start of the sampling campaign in February (during the hot-wet season), pH at all 5 sites ranged from 7.32 to 8.18. There was not much difference in March and April but there was an increase in July at all of the study sites (pH = 8.84-9.04). The pH values dropped again in August as the cold season ended, being in the range of 7.72-7.82, before increasing again in September to around 8.00-8.52. The pH dropped once again in October, to its lowest values for the entire study period at 4 of the 5 sites. At this time, the pH was in the range 7.10-7.56 (. 8a).
Ionic conductivity also fluctuated throughout the sampling period. In February, the conductivity ranged from 308.91 uS-cm-1 (at Site 1) to 346.00 uS-cm-1 (at Site 5). There was a sudden drop in March to between 180.33 uS-cm-1 (at Site 5) and 208.64 uS-cm-1 (at Site 1) as a result of rainfall that occurred during the month. This was followed by a significant increase in April, to a range of around 352.00-367.00 uS-cm-1. From that time onwards, conductivity levels continued to rise, reaching the highest values in October, when all sites were above 400.00 US-cm-1 (Site 1 = 427.63 uS-cm-1; Site 2 = 427.67 uS-cm-1; Site 3 = 433.00 uSc-m-1; Site 4 = 437.33 uS-cm-1 and Site 5 = 441.00 uS-cm-1) (Fig. 8c). Total dissolved solid (TDS) concentrations initally followed the same trend as conductivity (as these two variables are correlated), for February through April, when the highest concentrations for the study period were recorded (TDS = 342.75-352.91 mg-l-1). Thereafter, TDS concentrations decreased in May, to around 275-280 mg-l-1 for all study sites, before rising slightly from June to July to around 290-295 mg-l-1. Concentrations dropped again in August to 230-233 mg-l-1, before increasing to 251-254 mg-l-1 in October. Site 1 recorded the lowest and Site 5 the highest concentration of total dissolved solutes during the study (Fig. 8b).
Chemical oxygen demand (COD) was constant during the first 2 months (February and March) of the study and increased slightly in April before decreasing to its lowest values in June for Site 1 (19.20 mg-t-1) and in July for Sites 2-4 (Site 2 = 18.00 mg-t-1, Site 3 = 13.40 mg-t-1, Site 4 = 31.75 mg-t-1 and Site 5 = 9.50 mg-t-1). COD then increased at all study sites during August (Site 1 = 79.22 mg-t-1, Site 2 = 110.00 mg-t-1, Site 3 = 104.00 mg-t-1, Site 4 = 97.74 mg-t-1 and Site 5 = 100.87 mg-t-1) before decreasing again in October. Site 1 had the lowest while Site 5 had the highest COD for the entire study period (Fig. 8d).
Secchi disk (SD) transparency was lowest at Site 5 (below 1 m) and highest at Site 1 during the study period. Secchi disk transparency increased from February through May (Site 1 = 2.00 m, Site 2 = 1.80 m and Site 3 = 1.50 m) before decreasing in October (Fig. 8e). Secchi disk readings were relatively constant for Site 4. The highest SD water transparency levels were recorded in August (Fig. 8e). Total suspended solids (TSS) concentrations increased from February to August, and decreased to their lowest concentrations in October at Sites 1 through 4 (Fig. 8g). Turbidity levels were highest at Site 2 (114.67 mg-t-1) and lowest at Site 3 (4.6 mg-t-1). Turbidity levels decreased in August before increasing in September and finaling decreasing again in October (Fig. 8h).
Nutrients
Site 1 had the highest reactive phosphorus (RP) concentrations whilst Site 3 had the lowest concentrations. RP concentrations increased from February to March before dropping to their lowest concentrations in April. RP concentrations then increased in June before decreasing in July. From July to October, RP concentrations increased to their highest levels (Site 2 = 0.90 mg-t-1, Site 3 = 0.77 mg-t-1, Site 4 = 0.83 mg-t-1 and Site 5 = 0.76 mg-t-1), although the highest RP concentration, of 1.28 mg-t-1 was recorded at Site 1 in September (Fig. 9a). Total phosphorus (TP) concentrations were constant between February and March (at Site 1 through Site 4), decreasing in April and then increasing in June, before decreasing in July (Site 1 = 0.19 mg-t-1, Site 2 = 0.18 mg-t-1, Site 3 = 0.12 mg-t-1, Site 4 = 0.09 mg-t-1 and Site 5 = 0.10 mg-t-1). Ammonia concentrations increased from July to October at Site 2 (2.07 mg-t-1), Site 3 (1.77 mg-t-1) and Site 5 (2.13 mg-t-1). At Site 1 and Site 4, ammonia concentrations increased to a high in September (Site 1 = 2.94 mg-t-1 and Site 4 = 1.67 mg-t-1) before decreasing in October (Fig. 9c).
The highest ammonia concentrations at Site 1 were recorded in February (0.46 mg-t-1) when the rest of the sites had their lowest concentrations (at Sites 2-5 ammonia was undetectable). Ammonia concentrations at Sites 2 through 4 increased from February to a high in March. Site 1 had unde-tectable ammonia concentrations in April. Ammonia concentrations ranged from undetectable to 0.20 mg-t-1 between April and October at Sites 1-3, while ammonia concentrations increased at Site 5 during July (0.29 mg-t-1). Ammonia concentrations at Site 4 increased from 0.06 mg-t-1in April to 0.40 mg-t-1 in May before decreasing to 0.05 mg-t-1 in June (Fig. 9b). Nitrate concentrations decreased from February to March and then increased in April (Site 1 = 0.07 mg-t-1, Site 2 = 0.06 mg-t-1, Site 3 = 0.08 mg-t-1, Site 4 = 0.07 mg-t-1 and Site 5 = 0.06 mg-t-1). Nitrate concentrations decreased to undetect-able levels during July at all sites, and then increased during October (Site 1 = 0.03 mg-t-1, Site 2 = 0.04 mg-t-1, Site 3 = 0.02 mg-t-1 and Site 5 = 0.03 mg-t-1) (Fig. 9e).
Total nitrogen (TN) concentrations increased from February to March before becoming nearly constant. Concentrations at Site 5 decreased in June (to undetectable levels) and August (0.33 mg-t-1), and increased in July (2.56 mg-t-1) and September (2.96 mg-t-1). Sites 1, 2 and 4 recorded their highest TN concentrations in May (2.58 mg-t-1), June (1.22 mg-t-1) and September (1.21 mg-t-1) respectively (Fig. 9d). A slight variation in the N:P ratio was observed at all of the study sites and N:P ratios were below 1 for all sites during September and October, with the exception of Site 5 (1.37). The highest N:P ratio recorded was 56.96 for Site 5 , 6.37 for Site 2 and 2.18 for Site 1 =, while the lowest ratios were too low to calculate at Site 4 and Site 3. During the cool-dry season, Site 4 had an N:P ratio of 2.96 (May), Site 5 = 24.73 (July), Site 2 = 6.84 (July) and Site 3 = 5.52 (July) (Fig. 9f).
Discussion
The study area falls within the sub-tropical region where seasonality is defined by rainfall. This affects water levels in water bodies, especially small water bodies, resulting in large decreases in water level during the long dry season (April to October) as a result of evaporation and abstraction (drawdown). Shoreline development (DL) is an important aspect that reflects the potential for development of littoral communities, which are usually areas of high biological productivity. The DL in the Malilangwe reservoir was estimated at 1.94. This relatively large ratio meant that the shoreline was more crenulated than a circular water body of the same surface area, and, hence, has a greater potential for littoral community development. Despite a calculated water residence time of 4.4 years, the Malilangwe Reservoir has not spilled for the past 11 years. This is all the more remarkable when compared to other water bodies of similar size, such as Munwahuku Reservoir which spills multiple times annually (Nhiwatiwa and Marshall, 2006).
The bathymetric and side-scan sonar survey of the Malilangwe Reservoir revealed the original fluvial and alluvial state of this area prior to impoundment. Water volumes in the reservoir were calculated only for the area covered by the hydrographic survey; the upper portions of the main tributary leading in to the dam were not surveyed. These portions, therefore, did not form part of the water volume estimates. The reservoir volumes calculated for different water levels are therefore slightly less than the actual reservoir volume. The difference between the calculated volumes for the survey area and the actual reservoir capacity will become progressively greater as the dam water level drops.
Mixing in shallow and stratified reservoirs varied both daily and seasonally depending on solar irradiance and wind regimes. Nhiwatiwa and Marshall (2006) and Graham et al. (2008) showed that daily discontinuities in solar radiation and wind regimes are the main causes of stratification changes. The pronounced daily variations in atmospheric heat content, evaluated through the range of air temperature values, are related to nocturnal heat loss from the water, so that the instantaneous heat content of the water does not reflect the reality of heat exchanges (Arcifa et al., 1990). Temperatures were naturally higher during daytime in the hot-dry season than in the cool-dry season. Weak diurnal thermal and oxygen stratification was observed during the cool-dry season, mainly as a result of lower temperatures.
Deoxygenation of bottom waters was observed during the hot-wet and hot-dry seasons, while the whole water column was well oxygenated during the cool-dry season. The spatial variability of oxygen depletion (defined as <2 mgl-1 of dissolved oxygen) that the reservoir exhibited could be partly due to respiration as a result of decomposing organic matter and the nature of the soils (Chapman et al., 1998). Seasonal importation of allochthonous organic material during the rainy season is a characteristic feature of most tropical reservoirs, and Malilangwe Reservoir is no exception. The reduced rate of oxygen depletion during the cool-dry season is mainly due to complete mixing of the water column. Temperature is a major determinant of the rates of biological and biochemical processes such as decomposition (Nhiwatiwa and Marshall, 2006).
Dissolved oxygen (DO) depletion during the hot-wet and hot-dry seasons is a result of demand exceeding supply. Greater oxygen demand is due to the increase in activity of aquatic animals, and a greater abundance of aquatic plants and decaying organic matter which consume DO. Large fish populations in the reservoir have a faster metabolic rate as water temperatures increase with the change of season (from the cool-dry to hot-dry season), which increases their oxygen requirements during the warm weather periods. As a result, more oxygen is needed by the fish during the hot-wet and hot-dry seasons. Another factor leading to low DO levels during the hot-dry season could be decomposition by bacteria, a process that further reduces DO in the water column. The development of hypolimnetic anoxia also reduces the biologically-available habitat, reducing the volume of the water body in which the majority of aquatic organisms exist and severely impacting less mobile benthic organisms. Fish usually avoid water layers containing <2-3 mgl-1 oxygen concentration (Abd El-Monem, 2008) and the same is true for other aquatic organisms. About 40% of the Malilangwe Reservoir volume can be oxygen deficient (with the anoxic layer extending up to 4 m below the water surface), creating conditions of oxygen stress during the hot-wet and hot-dry seasons and greatly reducing available habitat.
Thermal and oxygen stratification regimes have implications for nutrient exchange between sediments and water. Cowan and Boynton (1996) observed that sediment oxygen consumption rates increase with increasing temperature until bottom-water dissolved oxygen concentrations fell below <3 mgl-1, at which point sediment oxygen consumption rates become limited. This partly explains the depletion of oxygen that occurred in Malilangwe Reservoir. With regards to nutrient cycling, high DO concentrations in the hypolimnion inhibit sediment release of ammonia, by enhancing nitrification of ammonia to nitrate (and thus nitrogen assimilation into bacterial biomass). However, fluxes of ammonia were elevated at high temperatures, and when coupled with increasing hypolimnetic anoxic conditions (<5 mgl-1) very large releases of ammonia occur (Beutel et al., 2008). Phosphate fluxes were small except in areas of hypoxic and anoxic bottom waters. It is likely that nutrient exchange is rapid and frequent in small reservoirs even though they are characterised by vertical gradients associated with thermal and oxygen stratification (Nhiwatiwa and Marshall, 2006). In Malilangwe Reservoir, there were short-term fluctuations in some isopleths of temperature and dissolved oxygen indicating that there is dynamism among the different layers, even during stratification. This is likely to result in partial mixing of the water column and may allow nutrient exchanges between the epilimnion and the hypolimnion.
Seasonal changes in water quality were also investigated. Conductivity was characterised by fluctuations between the cool-dry season and the hot-dry and hot-wet seasons. The lower conductivities measured during the wet season occur because of rainfall, during the few contiguous months of the year during which precipitation occurs. This results in a high dilution factor as river inflows enter the reservoir. Later in the year, after inflows have ceased, ionic concentrations increase as water levels drop. This concentration effect has been observed and reported in other studies (Nhiwatiwa and Marshall, 2007; Moss and Moss, 1969; Osborne et al., 1987). The temporal and vertical patterns of pH in the reservoir were mediated by processes of photosynthesis (production) and respiration (consumption). The high pH observed during the cool-dry season (June to July) could be attributed to photosynthetic uptake of CO2 by prolific algal blooms observed during the same period, while decomposition and respiration tended to decrease pH during the hot-wet and hot-dry seasons.
Secchi disk transparency is an important feature of water quality and has important ecological implications. Secchi disk transparency ranged from 0.2-1.8 m in Malilangwe Reservoir and was comparable to the characteristic modal range of 0.1-1.6 m recorded in 2 small reservoirs in the Manyame catchment (Nhiwatiwa and Marshall, 2007), as well as in Oyun Reservoir, Nigeria (Mustapha, 2008). Secchi disc transparency was low during the rainy season (February to March) and, in particular, at Site 5, throughout the study. The rainfall season is naturally characterised by high runoff resulting in sediment-laden inflows into the reservoir. Site 5 had riverine characteristics and was the point where the major inflows came into the reservoir. Higher water transparencies, observed in the dry season, reflected suspended particles settling to the bottom of the reservoir. The range of water transparency (0.2-1.8 m) indicates that depth of light penetration is generally adequate for the aquatic organisms (plankton and fish) that thrive in the pelagic regions of Malilangwe Reservoir (Mustapha, 2008).
The river inflows were clearly linked to higher phosphorus concentrations during the hot-wet season. These higher concentrations are most likely linked to the resultant algal blooms that occur during the cool-dry season and which contribute to the high level of productivity in the reservoir. Additional nutrients were also likely to be released during turnover, contributing to the overall increase in productivity (Mustapha, 2008). The overall decline in phosphorus concentrations, similarly, is linked to uptake by algae and macrophytes and, during the hot-dry season, retention in the hypolimnion. The decrease in water levels in the reservoir corresponded with an increase in phosphorus concentration, but this relationship was confounded by the concurrent effects of turnover.
Nitrogen concentrations were generally low. This finding supports the suggestion that nitrogen does not accumulate in tropical lakes and reservoirs due to the occurrence of anoxic hypolimnia (Bootsma and Hecky, 2003). Anoxia and warm temperatures promote rapid denitrification and enhance phosphorus mobilisation in tropical lakes, leading to the greater prevalence of nitrogen limitation in the tropics (Bootsma and Hecky, 2003).
Nitrogen to phosphorus (N:P) ratios observed in the reservoir were relatively unchanged during the study period. According to Smith (1979), nitrogen is limiting when the N:P ratio is less than 10:1 and phosphorus is limiting when N:P is greater than 13:1. Thus, in the Malilangwe Reservoir, nitrogen was found to be the limiting factor, as shown by the relatively low N:P ratio, of less than 10.9, at all study sites. Nevertheless, the trophic status of the reservoir is likely to change in the short- to medium-term to eutrophic, as the reservoir continues to accumulate nutrients. Nutrient loadings from the catchment are currently low; thus, the reservoir has not yet become eutrophic.
Acknowledgements
This study was made possible through the financial support of the Malilangwe Trust Research Grant and German Academic Exchange Service (DAAD - A/10/02914). Special thanks go to Esther S Jairos (University of Pretoria, RSA) for guidance and for providing most of the relevant literature used. Warm thanks go to Clemence Chakuya and Patrick Mutizamhepo of the University of Zimbabwe; Philemon Chivambu, Pandeni Chitimela, and Pamushana Lodge guides of the Malilangwe Wildlife Reserve, who assisted in the field. Our appreciation also goes to Elizabeth Munyoro and the technical staff of the Department of Biological Sciences, University of Zimbabwe, for all their technical support during the study.
References
ABD EL-MONEM AM (2008) Impact of summer thermal stratification on depth profile of phytoplankton productivity, biomass, density and photosynthetic capacity in Lake Nasser (Egypt). Jordan J. Biol. Sci. 1 173-180. [ Links ]
ARCIFA MS, MESCHIATTI AJ and GOMES EAT (1990) Thermal regime and stability of a tropical shallow reservoir: Lake Monte Alegre, Brazil. Rev. Hydrobiol. Trop. 23 271-231. [ Links ]
BARSON M, MULONGA A and NHIWATIWA T (2008) Investigation of a parasitic outbreak of Lernaea cyprinacea Linnaeus (Crustacea: Copepoda) in fish from Zimbabwe. Afr. J. Zool. 43 175-183. [ Links ]
BEUTEL M, HORNE A, TAYLOR W, LOSEE R and WHITNEY R (2008) Effects of oxygen and nitrate on nutrient release from profundal sediments of a large, oligo-mesotrophic reservoir, Lake Mathews, California. Lakes Reservoir: Res. Manage. 24 18-29. [ Links ]
BOOTSMA HA and HECKY RE (2003) A comparative introduction to the biology and limnology of the African Great Lakes. J. Great Lakes Res. 29 3-18. [ Links ]
CHAPMAN LJ, CHAPMAN CA, CRISMAN TL and NORDLIE FG (1998) Dissolved oxygen and thermal regimes of a Ugandan crater lake. Hydrobiologia 385 201-211. [ Links ]
COWAN JLW and BOYNTON WR (1996) Sediment-water oxygen and nutrient exchanges along the longitudinal axis of Chesapeake Bay: Seasonal patterns, controlling factors and ecological significance. Estuaries 19 562-580. [ Links ]
DALU T, NHIWATIWA T and CLEGG B (2012a) Aquatic macro- phytes in a tropical African reservoir: diversity, communities and the impact of reservoir-level fluctuations. Trans. R. Soc. S. Afr. 67 117-125. [ Links ]
DALU T, CLEGG B, MARUFU L and NHIWATIWA T (2012b) The feeding habits of an introduced piscivore, Hydrocynus vittatus (Castelnau 1861) in a small tropical African reservoir. Pan-Am. J. Aquat. Sci. 7 85-92. [ Links ]
DALU T, NHIWATIWA T, CLEGG B and BARSON M (2012c) Impact of Lernaea cyprinacea Linnaeus 1758 (Crustacea: Copepoda) almost a decade after an initial parasitic outbreak in fishes of Malilangwe Reservoir, Zimbabwe. Knowledge Manage. Aquat. Ecosyst. 406 1-9. [ Links ]
DJUKIC N, MALETIN S, PUJIN V, IVANC A and MILAJONOVIC B (1994) Ecological assessment of water quality of Tisze by physico-chemical and biological parameters. Tisca Szeged 28 37-40. [ Links ]
DORGEOLOH WG, SEAMAN MT and GAIGHER IG (1993) The physical and chemical limnology of Skerkfontein Dam, eastern Orange Free State, South Africa. Water SA 19 177-186. [ Links ]
EATON A, CLESCERI LS, RICE EW and GREENBERG AE (2005) Standard Methods for the Examination of Water and Wastewater (21st edn.). American Public Health Association (APHA) and Water Environment Federation (WEF), USA. [ Links ]
GRAHAM JL, LOFTIN KA, ZIEGLER AC and MEYER MT (2008) Cyanobacteria in lakes and reservoirs: Toxin and taste and odour sampling guidelines (version 1.0). US Geological Survey Techniques of Water-Resources Investigations. Book 9, Chapter A7, Section 7.5. 6-8. [ Links ]
HACH (2007) DR 2800 Spectrophotometer: Procedures Manual. Catalog Number DOC022.53.00725 (2nd edn.). Hach, Colorado, USA. [ Links ]
HART RC (1999) On the limnology of Spioenkop, a turbid reservoir on the upper Thukela River, with particular reference to the structure and dynamics of its plankton community. Water SA 25 519-529. [ Links ]
JACKSON D and MARMULLA G (2001) The influence of dams on river fisheries. In: Marmulla G (ed.) Dams, fish and fisheries. Opportunities, challenges and conflict resolution. FAO Fisheries Technical Paper No. 419. FAO, Rome. 166 pp. [ Links ]
JACOBS AFG, JETTEN TH, LUCASSEN DC, HEUSINKVELD BG and NIEVEEN JP (1997) Diurnal temperature fluctuations in a natural shallow water body. Agric. For. Meteorol. 88 269-277. [ Links ]
KATSEV S, CROWE SA, MUCCI A, SUNDBY B, NOMOSATRYO S, HAFFNER GD and FOWLE DA (2010) Mixing and its effects on biogeochemistry in the persistently stratified, deep, tropical Lake Matano, Indonesia. Limnol. Oceanogr. 55 763-776. [ Links ]
LEWIS WM Jr. (2000) Basis for the protection and management of tropical lakes. Lakes Reservoir: Res. Manage. 5 35-48. [ Links ]
McCARTNEY MP and SALLY H (2005) Managing the environmental impact of large dams in Africa. In: Lankford BA and Mahoo HF (eds.) Proc. East Africa Integrated River Basin Management Conference, 7-9 March 2005, Sokoine University of Agriculture, Morogoro (Tanzania). [ Links ]
McLACHLAN AJ (1974) Recovery of the mud substrate and its associated fauna following a dry phase in a tropical lake. Limnol. Oceanogr. 19 74-83. [ Links ]
MOSS B and MOSS J (1969) Aspects of the limnology of an endorheic African lake (L. Chilwa, Malawi). Ecology 50 109-118. [ Links ]
MUSTAPHA MK (2008) Assessment of the water quality of Oyun Reservoir, Offa, Nigeria, using selected physico-chemical parameters. Turkish J. Fish. Aquat. Sci. 8 309-319. [ Links ]
MUSTAPHA MK (2009) Problems, challenges and management of small, shallow tropical African reservoirs: A case study of Oyun Reservoir, Offa, Nigeria. Int. J. Lakes Rivers 2 163-174. [ Links ]
NHIWATIWA T and MARSHALL BE (2006) Seasonal and diurnal stratification in two small Zimbabwean reservoirs. Afr. J. Aquat. Sci. 31 185-196. [ Links ]
NHIWATIWA T and MARSHALL BE (2007) Water quality and plankton dynamics in two small dams in Zimbabwe. Afr. J. Aquat. Sci. 32 139-151. [ Links ]
NHIWATIWA T (2004) The limnology and ecology of two small man-made reservoirs in Zimbabwe. MPhil dissertation, University of Zimbabwe, Harare. [ Links ]
OSBORNE PL, KYLE JH and ABRAMSKI MS (1987) Effects of seasonal water level changes on the chemical and biological limnology of Lake Murray, Papua New Guinea. Aus. J. Mar. Freshwater Res. 38 397-408. [ Links ]
QUARCOOPOME T, AMEVENKU FYK and ANSA-ASARE OD (2008) Fisheries and limnology of two Reservoirs in Northern Ghana. West Afr. J. Appl. Ecol. 12 1-18. [ Links ]
SMITH VH (1979) Nutrient dependence of primary productivity in lakes. Limnol. Oceanogr. 26 1051-1064. [ Links ]
SYSTAT (2007)Mystat: A student version of Systat 32-bit UNICODE English. Version 12.02.00. SYSTAT Software, Inc. [ Links ]
Received 21 March 2012;
Accepted in revised form 11 December 2012.
* To whom all correspondence should be addressed. S +27 46 603 8524; e-mail: dalutatenda@yahoo.co.uk


{kind=link}
{kind=link}
{kind=link}
{kind=link}