










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkWater SA
versão On-line ISSN 1816-7950
versão impressa ISSN 0378-4738
Water SA vol.38 no.2 Pretoria Jan. 2012
ARTICLES
The response of microalgal biomass and community composition to environmental factors in the Sundays Estuary
Daisy Kotsedi; Janine B Adams*; Gavin C Snow
Department of Botany, Nelson Mandela Metropolitan University, PO Box 77000, Port Elizabeth 6031, South Africa
ABSTRACT
The Sundays Estuary is permanently open to the sea and experiences regular freshwater inflow in the form of agricultural return flows with large supplies of nutrients. The objectives of this study were to measure microalgal biomass and community composition and relate these to freshwater inflow, water quality and other environmental variables. These data can then be used in setting the ecological water requirements of the estuary. Surveys in August 2006, March 2007, February, June and August 2008 showed that salinity less than 10, expressed in practical salinity units, mostly occurred from 12.5 km from the mouth in the middle reaches of the estuary, which was also where the highest water column chlorophyll a (>20 μg.ℓ-1) was found. The study showed that different groups of microalgae formed phytoplankton blooms during individual sampling sessions. These included blooms of green algae (August 2006), flagellates (March 2007), dinoflagellates (June 2008) and diatom species (February and August 2008). The estuary was then sampled over 5 consecutive weeks from March to April 2009 to identify environmental factors that support different microalgal bloom species. Phytoplankton blooms were found during Weeks 1, 4 and 5 from the middle to the upper reaches of the estuary. It was shown that diatoms occurred in blooms during warm, calm conditions whereas wind-mixing and reduced temperature, as a result of a cold front during 17 to 19 March 2009, promoted the dominance of flagellates throughout the estuary although they were present at all times. Dominant diatom species (Cylindrotheca closterium, Cyclotella atomus and Cyclostephanus dubius) indicated brackish, nutrient-rich water. Nanoplankton (2.7 - 20 μm) was dominant during each week sampled and contributed 55 - 79% to the phytoplankton biomass. Maximum benthic chlorophyll a was found 12.5 km from the mouth. This study is the first to show successive chlorophyll a blooms consisting of different phytoplankton groups in an estuary, an indication of the eutrophic state of the system.
Keywords: Sundays Estuary, phytoplankton, microphytobenthos, chlorophyll a, environmental factors
Introduction
The Sundays River Estuary is an important recreational and tourist attraction, and now forms part of the Addo Elephant National Park. The estuary ranks 39 out of the top 50 South African estuaries (Turpie et al., 2002) in terms of biodiversity conservation importance. The estuary is permanently open, increasing its importance, as only 18% of South African estuaries have a permanently open connection with the sea. The Sundays Estuary does not have extensive intertidal sand banks, mudflats or salt marshes. Microalgae contribute 95% to primary production as there is little suitable habitat for macrophytes (Adams et al., 2008).
Phytoplankton play a significant role in nutrient cycling; they serve as a primary food source and are used as indicators of change in ecological conditions because they are sensitive to environmental perturbations (Lucas et al., 1999; Paerl et al., 2006). Benthic microalgae also play an important role in estuaries because they stabilise the sediment surface through growth and the production of extracellular polymeric substances (Underwood et al., 1995; Maclntyre et al., 1996); regulate nutrient fluxes/cycles and gas exchange (Sündback et al., 2003; Sündback et al., 2004); and serve as a highly nutritious food source for various benthic and pelagic fauna (Kibirige and Perissinotto, 2003; Nozais et al., 2005). The diversity of microphytobenthos species also provide numerous, sensitive indicators of environmental change and the specific conditions of their habitat.
As microalgae are an important source of primary production in the Sundays Estuary, studies of biomass and species composition are necessary to understand the functioning of the estuary. Phytoplankton in the Sundays Estuary were first investigated by Archibald (1981), with a focus on the taxonomy of diatoms, and subsequently Hilmer (1990) showed that the recurrent formation and decay of flagellate blooms was closely related to the spring-neap tide cycle. Blooms formed when semi-closed circulation increased the residence time of the water during stratified periods, usually at neap tides. Phytoplankton blooms have been recorded in the upper reaches of the estuary, when there was a water residence time of 7 neap tidal cycles and 3 spring tidal cycles (MacKay and Schumann, 1990). Phytoplankton from groups such as flagellates, dinoflagellates, and euglenoids have been recorded in the Sundays Estuary (Hilmer, 1990). Although diatom species have been shown to be continually present, they generally occur at low densities (Archibald, 1981; Hilmer, 1990) and high densities have been recorded shortly after flooding events (Jerling, 1994). Dinoflagellates have been found to be the dominant phyto-plankton community (Hilmer, 1990; Jerling, 1994), particularly in the vertically stratified middle reaches of the estuary, and cyanophytes have only been recorded in the upper half of the estuary (Hilmer, 1990; Archibald, 1981). Most blooms identified by Hilmer (1990) were caused by the dinoflagellate Katodinium rotundatum and a chlorophyte Micromonas pusilla.
Currently, estuarine research in South Africa has shifted its focus to temporarily open/closed estuaries (Gama et al., 2005; Thomas et al., 2005; Froneman, 2006; Perissinotto et al., 2006; Anandraj et al., 2008; Whitfield et al., 2008) with only a few recent studies focusing on permanently open estuaries. However, the Sundays Estuary has received great research attention in the recent past. The aim of the present study was to determine the microalgal biomass and community composition of the Sundays Estuary, a permanently open estuary. The objective was to gain insight into the response of microalgae to environmental conditions which can be useful in the freshwater inflow management of this important type of estuary in South Africa.
Materials and methods
Study area
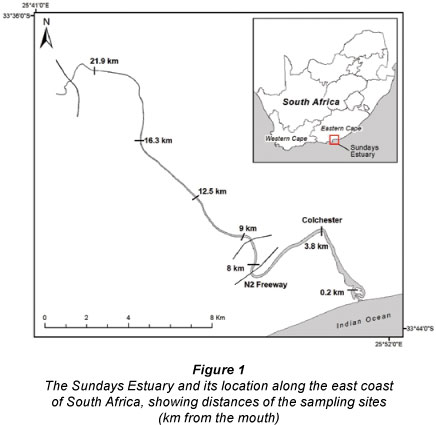
The Sundays River originates from the Sneeuberge and flows in a south to south-easterly direction. It passes through Graaff Reinet, Kirkwood and Addo in the fertile Sundays River Valley (Jerling, 1994). The river empties into the Indian Ocean at Algoa Bay, northeast of Port Elizabeth, through the Sundays Estuary (33° 43' S; 25° 51' E), and is permanently open to the sea (Fig. 1). Natural river flows are artificially augmented to ameliorate the poor water quality, in order to satisfy urban demands to Port Elizabeth and the irrigation demands in the catchment. In addition, some of the transferred water is allocated to freshening and flushing out of the Sundays River and there are high return flows from irrigation schemes. As a result, natural flows to the estuary have increased and are unlikely to decrease with the implementation of future schemes and water allocations in the catchment (Ninham Shand, 2008). Due to the strong inputs of freshwater resulting in large supplies of nutrients, the Sundays Estuary has a water column-based food web (Adams et al., 2008). The estuary has an intertidal zone mostly less than 5 - 6 m in width (Reddering and Esterhuysen, 1981) and there are no extensive intertidal sand banks, mud flats or salt marshes.
Field collection
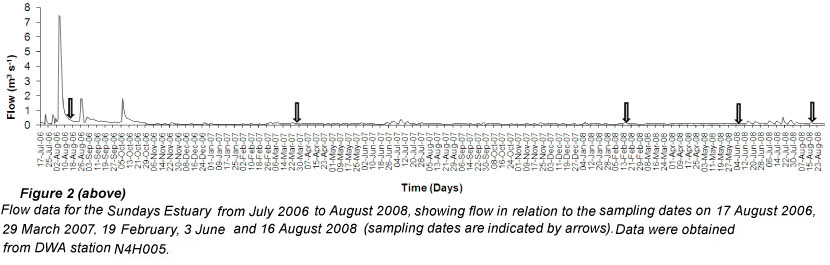
Rainfall records of the nearby city of Port Elizabeth indicate that rainfall has generally been highest during winter, between May and August, and the lowest in summer, between December and February. Samples were collected on 19 February, 3 June and 16 August 2008 in order to cover a range of freshwater inflow scenarios; from low river flow during summer to high flow in winter. There were 7 sites in total, which were 0.2, 3.8, 8.0, 9.9, 12.5, 16.3 and 21.9 km from the mouth (Fig. 1). Data sets from previous research (conducted on 17 August 2006 and 29 March 2007) similar to this study were incorporated into the current study.
For the short-term study, samples were collected on 13, 19, 26 March, then 2 and 9 April 2009 (referred to as Weeks 1 - 5) from approximately 09:00 to 14:00 at 5 sites along the Sundays Estuary. The sites were situated 3.8, 8, 9.9, 12.5 and 16.3 km from the mouth. Spring tides occurred in Weeks 1, 3 and 5 ' and sampling started during the ebbing tide on these dates at approximately 09:00. Neap tides occurred during Weeks 2 and 4 and sampling started during the flooding tide.
The physico-chemical and biological data collected on the first 5 sampling trips (August 2006 to August 2008) are indicated in Table 1. All variables listed in Table 1 were measured during the short-term study in March/April 2009.
Water quality variables
Water quality variables including temperature and salinity were recorded at each site using a 650 MDS YSI multiprobe. Additionally, rainfall data was obtained from Weather SA (http://www.weathersa.co.za). Water samples from the surface and bottom depths at each site were collected for nutrient analyses, and were filtered with a 0.45 μm pore size syringe through Millipore filters (Millex-HV Hydrophilic PVDF) and then stored in polyethylene sampling bottles. Samples were stored in a 'cooler box' (portable ice chest) in the field and stored in a freezer until further analysis. Samples were sent to the Council for Scientific and Industrial Research (CSIR) in Stellenbosch and analysed by accredited analytical laboratories using an auto-analyser (detection limit for dissolved nitrate, nitrite and total ammonium was 0.17 μM, for soluble reactive phosphorus (SRP) 0.32 μM and for dissolved reactive silicate 0.36 μM) according to the methods described in CSIR (2002). Dissolved inorganic nitrogen (DIN) is a combination of nitrate, nitrite and ammonium and dissolved inorganic phosphorus (DIP) is soluble reactive phosphorus. Samples were collected for nutrients on 29 March 2007, 19 February, 3 June and 16 August 2008, and during the short-term study in March/April 2009.
Phytoplankton biomass (chlorophyll a)
Water samples were collected using a 500 mℓ pop-bottle at the surface (0 m), 0.5, 1, 2, 3 and 4 m (if possible) depths. The samples were gravity-filtered under vacuum through 1.2 μm Whatman GF/C filters. The filter papers were placed into glass vials containing 10 mℓ of 95% ethanol (Merck 4111) and left overnight for chlorophyll a extraction at 1 - 2°C. After extraction, chlorophyll a spectrophotometric determinations were carried out according to Nusch (1980). Absorbances, before and after acidification of the extracts with 0.1 N HCl, were read using a UV/VIS spectrophotometer at 665 nm.
For the short-term study, size-fractionated chlorophyll a was measured. Phytoplankton samples were serially filtered through a 20 μm nitex screen mesh, 2.7 μm Whatman GF/D filter paper and, finally, through a 0.7 μm Whatman GF/F filter paper which might allow small cyanobacterial cells through. During the short-term study, a maximum acid ratio of 1.7 was used instead of an after-acid reading. The incorrect concentration for HCl was used in the after-acid readings; therefore a maximum acid ratio of 1.7 was used with the readings without acid. For the short-term study the following equation was used:
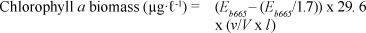
where:
Eb665 = absorbance at 665 nm before acidification
Ea665 = absorbance at 665 nm after acidification
v = volume of solvent used for the extraction (mℓ)
V = volume of the sample filtered (ℓ)
l = path of spectrophotometer cuvette (cm)
Phytoplankton community composition
Water samples (500 mℓ) for phytoplankton enumeration and identification were collected at the surface (0 m) and at depths of 0.5, 1.0 and 2.0 m, and were preserved with 1.5 mℓ of 1% (v/v) glutaraldehyde until counts were ready to be done. Eight drops of Rose Bengal were added to 60 mℓ of the preserved samples and the samples were put into Utermohl settling chambers and allowed to settle for 24 h. Counts and identification of microalgal groups (i.e. flagellates, dinoflagellates, diatoms, cyanophytes and chlorophytes) were performed using the Zeiss IM 35 inverted microscope at 630x magnification. The number of phytoplankton cells from the field of view was calculated according to a formula by Snow (2007).
Benthic microalgal biomass (chlorophyll a)
Four replicate intertidal and subtidal benthic samples were collected from each site. Intertidal samples from the uppermost 2 sites of the estuary were not collected due to the steepness of the banks. The surface sediment (<2 mm depth and 20 mm pre-marked area) was scraped just above the water level at low tide for intertidal samples. Subtidal samples were collected from each site using a 20 mm internal diameter corer attached to an extension pole and the top 1 cm sediment was scraped from the core. The samples were stored in vials in the 'cooler box' (portable ice chest) until they could be frozen. The samples were freeze-dried and 4 mℓ of 95% ethanol (Merck 4111) was added to approximately 0.2 g then stored for 24 h at 0°C. After the chlorophyll a extraction, the samples were filtered through Whatman GF/C filters and the extracts were analysed before and after the addition of 2 drops of 0.1 N HCl, using a spectro-photometer at 665 nm according to Nusch (1980).
For the short-term study only subtidal benthic biomass was measured as the objective of this study was to investigate the deposition of water column microalgae.
Benthic diatom composition
Benthic samples were collected according to methods described by Round (1981) and Bate et al. (2004). The samples were digested using 10 mℓ of saturated KMnO4 and 10 mℓ of 10 M HCl were added. Processed samples were stored in 2 mℓ Eppendorf safe-lock tubes until mounting. Permanent light microscopy slides were made with the digested diatom samples. NaphraxTM was used as a mounting agent and 2 drops were used. Diatom frustules were examined and counted using a Zeiss Axioplan light microscope with Differential Interference Contrast (DIC) optics. Using a television camera (The Imaging Source DFK 41F02), images of the dominant species were visualised using the Imaplan V 2.06 image analysis programme (IMATEC Elektronische Bildanalysesysteme GmbH ©2004). Diatom valves were counted in each sample using 1 000x magnification until the obvious dominant was established. In most cases this represented approximately 200 frustules. At least 1 digital image of every taxon was captured. The dominants were those species that were clearly present in the greatest number and the sub-dominants were those that had a frequency >10% but not dominant.
Benthic algal community composition
For the short-term study the entire benthic community was counted and identified. Sediment samples (surface 1 mm) were collected using a 20 mm internal diameter corer and placed in a 50 mℓ vial with about 20-25 mℓ unfiltered estuarine water in the field. The sampling method does combine surface sediment and unfiltered water from above the sediment core so samples represent a mix of benthic and pelagic taxa. Samples were preserved with 1.5 mℓ of 1% (v/v) glutaraldehyde solution in the field and stored at 5°C in a darkened environment. For enumeration purposes, samples were re-suspended and stained with Rose Bengal. The stained samples were allowed to stain for 24 h. A drop was placed on slides with coverslips on top then identified under a light microscope at 400x magnification. Counts and identification of microalgal groups (i.e. flagellates, dinoflagellates, diatoms, cyanophytes and chlorophytes) were performed using the Zeiss IM 35 inverted microscope at 630x magnification. Relative abundances of the different groups were reported.
Data analysis
All statistical analyses were completed using the MINITAB Version 15 (Minitab, Inc.) statistical package. The data collected (Table 1) were categorised as follows: algal counts, biomass and physico-chemical from February 2008 to August 2008; algal and physico-chemical data for 5 weeks in 2008. The data (phytoplankton counts) from 2006 to 2007 represented means and were excluded from the current analysis, but were used for long-term comparisons. Data for each sampling trip were analysed and comparisons were also made for all trips combined.
All data were tested for normality using the Kolmogorov-Smirnov test for normality. The null hypothesis of normality was rejected for most response variables and a Johnson transformation was performed to normalise such data. The Johnson transformation applies to a broader range of data types than the alternative Box-Cox transformation. It is equally robust for data sets with negative values and selects an appropriate transformation function for each data set because of its wider range of transformation functions (Farnum, 1996; Chou et al., 1998). The maximμM p-value for selection of an appropriate bounded (SB), lognormal (SL) or unbounded (SU) transformation distribution was 0.10.
Levene's test for homogeneity of variance, with a confidence interval of 95%, was used to test the equality of variance across variables. Following a positive test, a general linear model (GLM) analysis of variance (without interactions) was conducted on the transformed data to test the effect of the different predictors (distance from estuary mouth, sampling time and depth) on the response variables at a = 0.05 (although transformed data were used in the analyses, for purposes of clarity, means and standard errors of the actual untransformed data are presented in some of the results). Pearson product moment correlation was performed to test the relationship between variables. Contour plots for chlorophyll a and phy-toplankton cell counts were produced using Grapher (Golden software) Version 8. Contour XY Data Map was used for the plots. Different scales were used for the ;y-axis for different sampling dates so that the patterns were visible. In some cases, very high values were recorded but only for a single sampling date. Mean values are expressed as mean ± standard error of the mean.
Results
Long-term study
Water quality variables
Flows of 7.5 and 7.4 m3s-1 were recorded at Department of Water Affairs (DWA) Station N4H005 on 4 and 5 August 2006 (Fig. 2). These high flows occurred prior to the 17 August 2006 sampling session as a result of rainfall (128 mm and 64 mm on 2 and 3 August 2006, respectively). The highest mean monthly flow of 1.29 m3s-1 also occurred in August 2006. For the other sampling sessions flow was less than 0.15 m3s-1 (Fig. 2). Data from the environmental water requirements study on the Sundays Estuary (Taljaard et al., 2008) was also used to identify the flow into the estuary at the time of sampling, based on the plots of the salinity gradient. The estuary in August 2006 was in a freshwater-dominated state which is characterised by flow greater than 15 m3s-1. In March 2007, and February, June and August 2008, a freshwater front was present in the upper and middle reaches. This was classified as a small transition state where flow is between 0.5 to 2 m3s-1 (Taljaard et al., 2008).
Both vertical and horizontal salinity gradients were observed during all sampling trips. Salinity, expressed in practical salinity units (PSU), was highest at the mouth (15 ± 0.1 to 28.2 ± 0 across the study period from 2006 to 2008) and decreased towards the head of the estuary and with depth (0.6 to 1.6 at 22 km from the mouth) (Fig. 3a - e). Vertically averaged salinity of less than 10 PSU occurred from 12.5 km upstream of the mouth, and this was the region where the highest water colμMn chlorophyll a was found (Fig. 4a - e). Freshwater intrusion (salinity <5) occurred up to 8 km and 13 km from the mouth in August 2006 and March 2007, respectively. In 2008 freshwater intrusion was evident at 11 to 12.5 km from the mouth. Salinity differed significantly with time (df = 4; F = 13.47; p <0.05; R2 = 84.3%). Salinity in June 2008 (16.7 ± 1.7) was significantly higher than August 2006 and March 2007 (7.7 ± 2.3 and 12.2 ± 2.6, respectively).
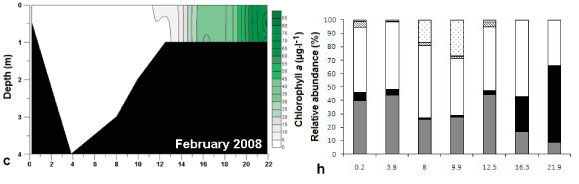
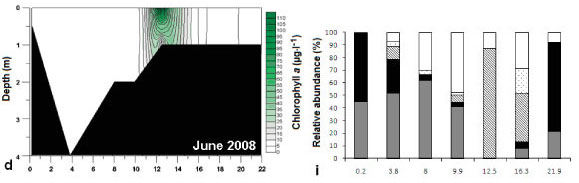
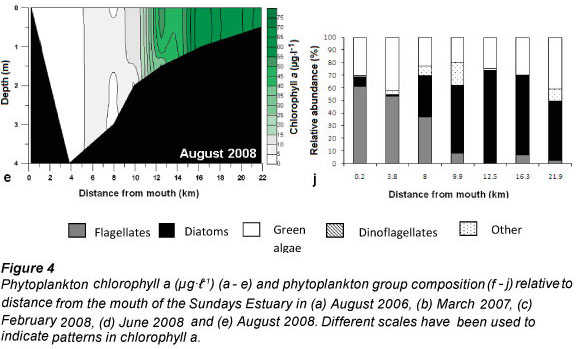
Generally, there was a decrease in temperature from the lower reaches to the upper reaches of the estuary (Fig. 3f - j). There was a significant difference in the overall mean temperature during the different sampling periods (df = 4; F = 167.88; p <0.05; R2 = 81.4%), where August 2006 was lower than the rest of the trips and February 2008 had the highest temperature. Temperature varied significantly with distance from the mouth of the estuary (df = 19; F = 0. 37; p <0.05; R2 = 50.3%).
Nutrients
Average dissolved inorganic nitrogen (DIN) at each site ranged from 0.1 μM to 134.6 μM during the 2007 and 2008 sampling sessions, typically increasing with distance from the mouth of the estuary. Dissolved inorganic phosphorus (DIP) also increased with distance from the mouth and ranged from below detectable limits to 8.8 μM. The DIN:DIP ratio was <16 in 2008, which suggests that microalgal growth was N-limited, but this was only likely in February 2008 when DIN concentrations as low as 0.1 μM were measured.
Phytoplankton biomass (chlorophyll a)
Water colμMn chlorophyll a (Fig. 4a- e) was significantly different (df = 4; F = 2.76; p <0.05, R2 = 9.4 %) for the different sampling trips (February 2008 to August 2008). Chlorophyll a was significantly higher in March 2007 and August 2008 (31.5 ± 12.6 and 24.08 ± 5.56 μg.ℓ1) than in August 2006 and June 2008 (14.15 ± 1.45 and 8.16 ± 4.37 μg.ℓ-1). Chlorophyll a was always higher in low-salinity water and decreased with water depth (r = -0.53 and -0.27 respectively; p <0.05, n = 75).
Phytoplankton community composition and distribution
For this study phytoplankton groups with a relative abundance greater than 10% of the total algal count were considered to be dominant. Phytoplankton community composition is presented in Figs. 4f - j. The following phytoplankton groups were identified: flagellates, diatoms, dinoflagellates, chlorophytes (green algae) and cyanophytes (blue-green algae). For the purposes of this study, organisms that were considered to be flagellates have 1 or more flagella and do not fall within the phytoplankton groups already mentioned. The group 'other' included planktonic organisms (mostly zooplankton, e.g. ciliates) not belonging to the above-mentioned groups.
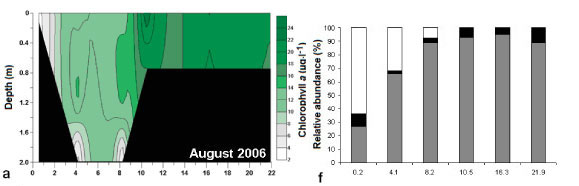
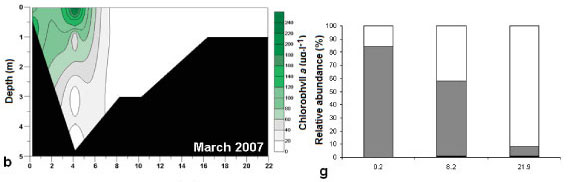
During all sampling trips the highest phytoplankton cell density occurred from 12.5 km to 21.9 km from the mouth. Different groups of microalgae formed phytoplankton blooms (defined as chlorophyll a greater than 20 μg.ℓ-1) during the different sampling trips (Figs. 4f - j). These included blooms of green algae (August 2006), flagellates (March 2007), dinoflagellates (June 2008) and diatom species (February and August 2008).
In August 2006 green algae were dominant at 0.2 km from the mouth of the estuary with mean cell numbers of 9 042 cellsm.ℓ-1 and making up 64% of the composition (Fig. 4f). The dominant green alga, Diogenes, are freshwater alga (Prescott, 1970). Flagellates were dominant from 4.1 km and towards the head of the estuary (8 449 to 9 319 cellsm.ℓ-1).
An opposite trend was observed in March 2007 where green algae were dominant (5 741 cells mℓ-1 to 41 804 cells.mℓ-1) towards the head of the estuary (Fig. 4g). By contrast flagellate density decreased from the mouth to the head of the estuary (from 31 324 to 3 400 cells.mℓ-1). A freshwater flagellate from the genus Chlamydomonas (Prescott, 1970) was dominant in the lower reaches.
In February 2008 a diatom bloom of Cyclotella atomus Hustedt was present in the upper reaches of the estuary with 11 028 ± 40 (± SE) and 55 738 ± 0 cells.mℓ-1 at 16.3 and 21.9 km from the mouth (Fig. 4h), respectively, contributing to the high chlorophyll a (34.4 ± 0.9 μg.ℓ-1 and 73.6 ± 3.3 μg.ℓ-1 respectively) found at these sites (Fig. 4c). This group respectively contributed 26% and 57% to the total community composition at these sites. Diatom blooms also occurred in August 2008 (Fig. 4j) and the bloom species were identified as C.atomus Hustedt, Cyclostephanus dubius (Fricke) Round and Stephanodiscus Hantzshia Grunow. At sites 9.9, 12.5, 16.3 and 21.9 km from the mouth diatom cell densities of 12 502 ± 3 460 cells.mℓ-1, 66 886 ± 0 cells.mℓ-1, 58 525 ± 2 787 cells.mℓ-1 and 35 885 ± 1 742 cells.mℓ-1 were recorded, respectively. Diatoms contributed 53, 74, 63 and 47% to the total phytoplankton composition at the mentioned sites, respectively.
Phytoplankton chlorophyll a peaked 12.5 km from the mouth in June 2008 (Fig. 4d) and was associated with a dense unidentified dinoflagellate bloom (11 705 ± 7 246 cells.mℓ-1) with relative abundance of 88% (Fig. 4i). Dinoflagellate density was still relatively high upstream of the bloom site (3 583 ± 1 407 cells.mℓ-1 at 16.3 km from the mouth).
Benthic microalgal biomass (chlorophyll a)
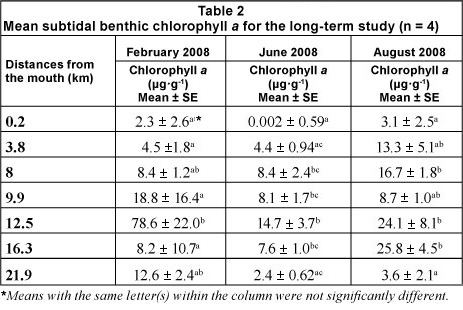
During the sampling trips (February, June and August 2008), benthic microalgal samples from the intertidal areas were not collected at sites 16.3 and 21.9 km from the mouth due to the channel-like morphology of the estuary and the presence of dense reed beds. Mean subtidal benthic chlorophyll a was significantly different at each sampling site (df = 6; p <0.05) during each trip (F = 3.44, 6.43 and 7.14; R2 = 35.8, 54.7 and 57.7% for February, June and August 2008 respectively). Intertidal benthic chlorophyll a was also found to be significantly different (df = 4; p <0.05) for all sampling trips (F = 13.59, 16.29 and 4.47; R2 = 72.6, 76.3 and 42.2% for February, June and August 2008, respectively). Benthic chlorophyll a at 12.5 km from the mouth was significantly higher than at other sites during most of the sampling trips (Tables 2 and 3). Maximum phytoplankton biomass was always found at the sites where the highest benthic microalgal biomass was recorded.
Short-term study Nutrients
During the short-term study (13 March - 9 April 2009), DIN ranged from 21.4 μM to 179.1 μM and typically increased with distance from the mouth of the estuary. Dissolved inorganic phosphorus increased with distance from the estuary mouth but concentrations were particularly low, ranging from 0.2 μM to 1.5 μM. The low DIP concentrations relative to DIN resulted in high DIN:DIP values (>34), suggesting that microalgal growth was P-limited during this period.
Phytoplankton biomass (chlorophyll a)
Chlorophyll a concentrations were significantly different (df = 4; F = 5.52; p <0.05; R2 = 52.5%) for the different weeks.
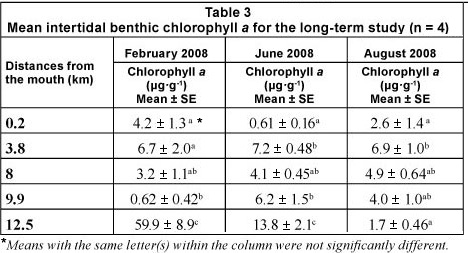
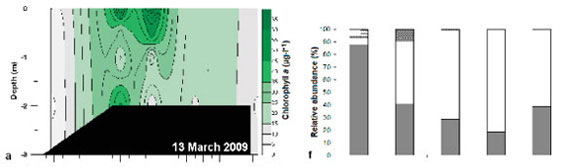
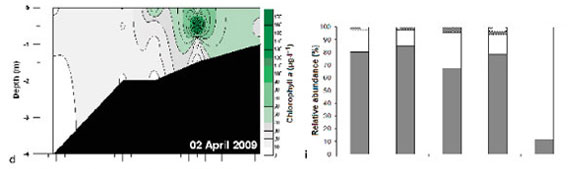
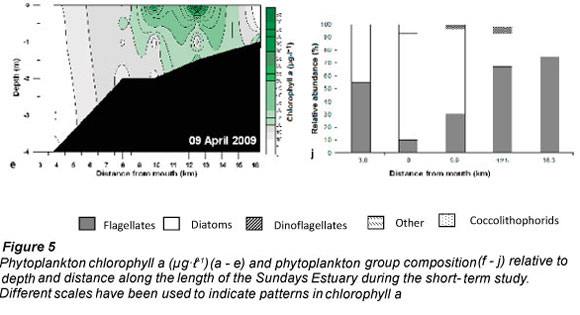
Chlorophyll a concentrations in Weeks 1, 4 and 5 (21 ± 3.8; 30 ± 7.2 and 17 ± 4.1 μg.ℓ -1) were significantly higher than in Weeks 2 and 3 (9.4 ± 1.5 and 11.7 ± 1.1 μg.ℓ -1). There was a strong cold front (17 to 24°C) from 17 to 19 March 2009 which mixed the water colμMn, resulting in a decrease in chlorophyll a (<20 μg.ℓ -1). The highest chlorophyll a biomass (>20 μg.ℓ -1) occurred in the middle to upper reaches of the estuary during Weeks 1, 4 and 5 (Fig. 5a - e).

Phytoplankton size-fractionated chlorophyll a
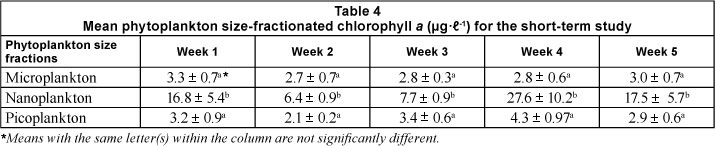
One-way ANOVA showed that the size fractions of chlorophyll a differed significantly (p <0.05) within the sampling trips. The nanoplankton size fraction was significantly higher during each week than the other 2 size fractions and contributed between 55 and 79% to the phytoplankton biomass (Table 4).
Benthic microalgal biomass (chlorophyll a)
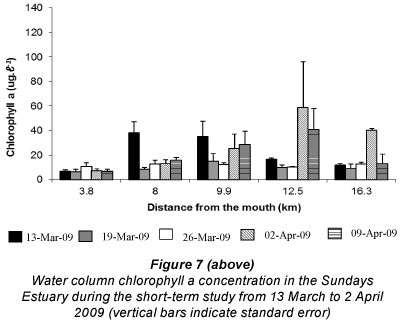
Subtidal benthic chlorophyll a increased from the mouth towards the upper reaches of the estuary (Fig. 6), following the pattern of the water column biomass (Fig. 7). There was a peak in the water column chlorophyll a in Weeks 4 and 5 at 12.5 km from the mouth, which can also be seen in the subtidal benthic biomass. Benthic and water column chlorophyll a were not collected in the same manner so, in order to determine if high benthic chlorophyll a concentrations were caused by water column chlorophyll a, average values per site were used. Benthic and water column microalgal biomass from Weeks 4 and 5 were positively correlated (r = 0.92 and 0.95 respectively; p <0.05; n = 5). The highest subtidal benthic biomass occurred in the same region of the estuary where water column chlorophyll a concentrations were high.
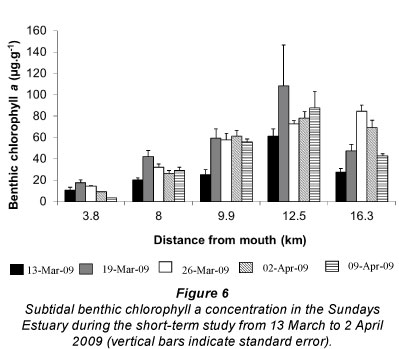
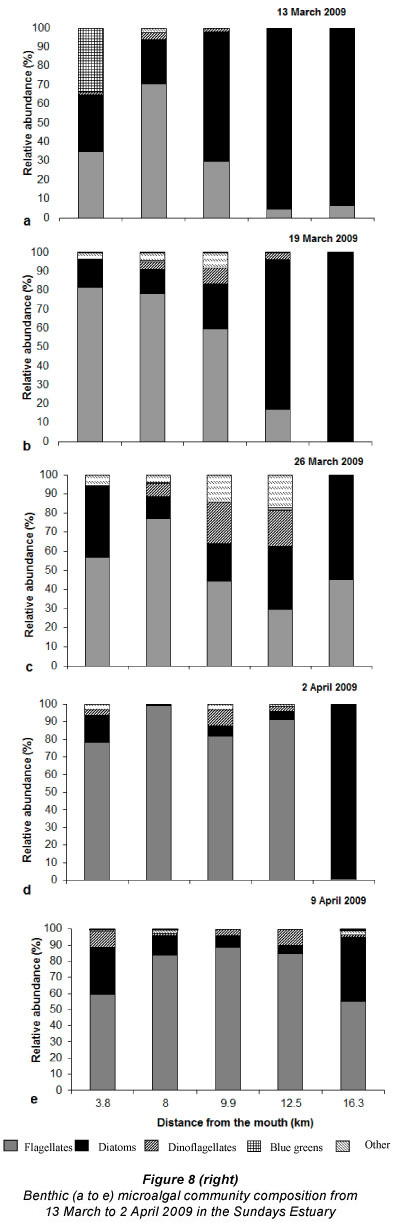
Benthic algal community composition
The patterns of species composition for the benthos and water column were similar to one another during all weeks, possibly indicating deposition of cells from the water column to the benthos (Fig. 8a - e and Fig. 5f - j). In Weeks 1 and 2 flagellates were dominant in the lower to upper reaches whereas diatoms became dominant in the upper reaches. This pattern was also observed for the phytoplankton composition in the same week. Flagellates were dominant in the water column throughout the estuary in Week 3. They were also present in the benthos but not as dominant as the flagellates in the water column (29 - 74% dominance). During Week 4 the benthic diatoms at 16.3 km from the mouth constituted 99% of the composition, similarly to water column diatoms (88%). In Week 5 flagellates dominated the benthos throughout the estuary, as well as the water column, except at 8 and 9.9 km from the mouth. No significant correlations were found when comparing relative abundances of microalgal groups from the benthos and water column (flagellates, dinoflagellates, other plankton) for all the weeks and sites. However, diatom relative abundance was significantly correlated between water column and sediment surface (r =0.50; p <0.05; n = 25).
Discussion
Long-term study
The objectives of this study were to determine the microalgal biomass and community composition in the Sundays Estuary. Results show a unique case where the chlorophyll a blooms in the estuary consist of multiple microalgal groups. High chlorophyll a concentrations (>20 μgℓ-1) in the shallow upper reaches of the Sundays Estuary have previously been found where nitrate concentrations were greater than 14 μM, water residence time was 6 - 7 tidal cycles and where there was marked salinity stratification (Hilmer and Bate, 1991; Adams and Bate, 1999; Scharler, 2000). Scharler (2000) recorded a bloom in the middle and upper reaches of the Sundays Estuary and recorded a mean chlorophyll biomass of 22 μgℓ-1 at nitrate concentrations of 33 μM in the upper reaches. Similarly, in the Great Fish Estuary consistent freshwater inflow carrying a high nutrient load supported high (>20 μgℓ-1) phytoplankton biomass (Allanson and Read, 1995). High (>20 μgℓ-1) chlorophyll a levels are a persistent feature in the upper reaches of the Sundays Estuary. This has been found to be the most productive region, which was also evident in this study, and the occurrence of phytoplankton blooms indicated nutrient uptake from the water column. Water residence time is important and allows for phytoplankton bio-mass to increase (Adams and Bate, 1994; Gameiro and Brotas, 2010). However, residence time is not easy to measure, which is a problem since realistic simulation of phytoplankton populations depends on accurate interpretations of mixing processes in estuaries (Cloern, 1991).
Hilmer (1990) explained the distribution of phytoplankton chlorophyll a in connection with the hydrodynamic features of the estuary. In the lower reaches of the estuary the biomass was lower because the bathymetry is uneven and deep and promotes turbulent mixing. As a result, phytoplankton experience rapid changes in salinity and flushing time (1 to 2 tidal cycles), which would result in death and rapid flushing out to sea (Hilmer, 1990). The middle and upper reaches of the estuary are shallower and have longer residence time and semi-closed circulation occurs (Jerling, 1994; MacKay, 1988). These findings correspond with the findings of the current study.
This study showed that in the Sundays Estuary blooms could consist of dinoflagellates, diatoms, green algae and flagellates. Under low to medium flow conditions, indicated by salinity profiles, diatoms and dinoflagellates were dominant, whereas under high flow conditions flagellates and green algae occurred. A flood occurred in the Eastern Cape region on the 2 August 2006 and sampling took place on 17 August. This event accounted for the dominance of the freshwater flagellates at the head of the estuary and the decreasing gradient in cell density towards the mouth. A freshwater flagellate (Chlamydomonas sp.) was the dominant phytoplankton species in March 2007. Only during this sampling session was the chlorophyll a maximμM (145.2 and 237.4 μgℓ-1 at 0.2 and 4.1 km respectively) found in the lower reaches of the estuary. High rainfall (38 to 40 mm) that occurred before sampling (4 and 5 of March), and an increase in flow, most likely transported the flagellate to the lower reaches of the estuary. The diatom cell numbers ranged between 11 028 and 66 886 cells.mℓ-1 in the middle to upper reaches of the estuary during the February and August 2008 sampling trips. Jerling and Wooldridge (1995) also recorded a bloom (13 000 cells.mℓ-1) of centric diatoms in the middle and upper reaches of the Sundays Estuary after a flood in November 1989.
Past studies have shown that dinoflagellates form dense blooms during neap tides when strong vertical salinity stratification has developed; the difference in surface and bottom water salinity is greater than 5 (Margalef, 1978; Hilmer and Bate, 1991). In this study a dinoflagellate bloom (18 951 cells.mℓ-1) occurred in June 2008 in the surface waters at 12.5 km from the mouth. The dinoflagellate bloom contributed to the high chlorophyll a biomass (99.5 μgℓ-1) and this was supported by a strong positive correlation between dinoflagellate cell numbers with chlorophyll a (r = 0.97; p <0.05; n = 25).
The highest benthic microalgal biomass found in the present study was located 12.5 km from the mouth during all sampling trips. This site was turbid, the intertidal area consisting of compacted clay, whereas the subtidal sediment was sandy and the water column was strongly stratified. It is possible that the deposition of phytoplankton cells in this area contributed to high benthic chlorophyll a. Furthermore, low flows possibly resulted in increased sediment stability which in turn supported high benthic microalgal biomass. A study in the Gamtoos Estuary showed similar results in that flows of less than 1 m3s-1 resulted in the development of epipelic diatom biofilms and, subsequently, high microphytobenthos biomass (Snow, 2007). Sandy silts and exposed habitats usually support a lower microalgal biomass than sheltered sites that are dominated by fine cohesive sediment (MacIntyre et al., 1996; Underwood and Kromkamp, 1999). The results of the current study were also similar to findings in the Kwelera Estuary where the distribution of microphytobenthic biomass was found to be strongly influenced by position along the longitudinal gradient (Walker, 2003). Adams and Bate (1994) also found that the chlorophyll a concentrations were highest in the middle reaches of the Swartkops Estuary compared to the mouth and upper reaches. Deductions from Walker (2003) were that the more turbulent hydrological conditions, together with loose sediment in the mouth area, did not support microphytob-enthos biomass, and low levels were consistently found in the sandy sediments near the mouth, whereas higher concentrations were found in muddier sites during the current study. Furthermore, during this study, cattle and benthic cyanobacterial mats were observed in the intertidal zone at 12.5 km from the mouth, which could have also contributed to the high benthic chlorophyll a levels at this site.
Short-term study
The short-term study was conducted in order to determine the short-term variability of phytoplankton composition and biomass in response to physical, chemical and climatic factors. Carstensen et al. (2004) stated that blooms develop from the active growth of phytoplankton populations, which must be accompanied by the input of new nutrients to the surface water. For this study on the Sundays Estuary high chlorophyll a was mainly associated with flagellates and diatoms, and the diatom species Cyclotella atomus, Cyclostephanus dubius and Cylindrotheca closterium made up most of the diatom blooms observed in the middle to upper reaches of the estuary. The dominant diatom species found during the short-term study corresponded with previously reported bloom species that have been identified for the Sundays Estuary in 2008 (long-term study) and similar to the findings of Jerling and Wooldridge (1995) on the centric diatom bloom in the middle and upper reaches. Cylindrotheca closterium blooms occurred on 13 March 2009 at 8 and 9.9 km from the mouth. This species has never been recorded in blooms in the Sundays Estuary before. Diatoms have been shown to be the most favourable phyto-plankton group in well-mixed, nutrient-rich waters (Margalef, 1978; Lassen et al., 2004; Domingues et al., 2005).
Although phytoplankton succession was not strongly exhibited in this study, diatoms and flagellates were the most abundant groups, succeeding each other in terms of dominance. Diatom blooms were mostly found in the middle to upper reaches whereas flagellates were found in the estuary at all times, and along the entire length of the estuary.
Nanoplankton contributed on average 55 - 79% to the total phytoplankton biomass in the weeks sampled. Results from this study are also supported by the findings of Revelante and Gilmartin (1978), Lassen et al. (2004) and Verity and Borkman (2010), in which nanophytoplankton were found to be the dominant size class in estuaries. Nanoplankton have also been found to be dominant in South African temporarily open/closed estuaries (Froneman, 2000; Thomas et al., 2005). They are able to out-compete larger species for nutrients and light because of their small size and large surface to volμMe ratio (van den Hoek et al., 1995). Nanophytoplankton are able to propel themselves to zones of greater light intensity and can therefore achieve much higher biomass than microphytoplankton because of their competitive advantage during periods of decreased light (Thomas et al., 2005).
Grange et al. (2000) indicated that the continuous inflow of freshwater in permanently open estuaries ensures that the phytoplankton size structure remains constant throughout the season. Phytoplankton size structure in the Sundays Estuary throughout the sampling sessions was consistently maintained and dominated by the nanoplankton size group. In contrast, the size structure of the phytoplankton community is highly variable in temporarily open/closed estuaries due to the sporadic inflow of freshwater. For instance, in the Kasouga Estuary nano- and microphytoplankton were dominant when the mouth was open, but during the closed mouth phase picophytoplank-ton were dominant (Froneman, 2002).
Results from the long-term study were also supported by the findings from the short-term study. During the 2 studies phytoplankton blooms (>20 μgℓ-1) were recorded 8 km upstream of the mouth. High benthic biomass was found in the same area of the estuary where high water column chlorophyll a occurred. The short-term study also showed that the patterns of phytoplankton and benthic species composition were similar for specific sites. The sampling method does combine surface sediment with water collected from above the core, to capture any flocculated material, so it is likely that the micro-algae are a combination of the benthic and pelagic communities. Dinoflagellates and flagellates were present in the benthic samples, which are typical water column species that depend on flagella for movement (Tomas, 1997).
From the 5-week sampling period in 2009 no single factor could be identified as the driver or trigger of phytoplank-ton blooms. However, it was shown that diatoms occurred in blooms during warm, calm conditions, whereas wind-mixing and reduced temperature (due to the cold front) promoted the dominance of flagellates throughout the estuary and they were present at all times. Both DIN and DIP generally increased with distance from the mouth of the estuary but DIP possibly limited microalgal growth based on the high DIN:DIP ratios, particularly in the lower reaches. It is likely that flow supported high phyto-plankton chlorophyll a in the middle reaches of the estuary but as DIP decreased towards the mouth of the estuary, being taken up by phytoplankton, a threshold DIP concentration was reached and chlorophyll a was generally lowest in the lower reaches. Nutrients were evenly distributed on 26 March 2009 and DIP was <1.0 μM, possibly limiting the growth of phytoplankton.
Conclusion
This study has contributed to an understanding of microalgal responses in a nutrient-rich permanently open estuary. The results from this study were used to provide specialist input on the microalgal component for the determination of the ecological water requirements for the Sundays Estuary. This study showed that water column chlorophyll a was frequently >20 μg.ℓ-1, and phytoplankton density often > 10 000 cells.mℓ-1, indicative of an extremely productive estuary.
The consistently high nutrient-rich river flow keeps the build up of organic material low so hypoxic and anoxic events do not occur often. However, a reduction in river flow and the subsequent build up of sand in the mouth area are likely to increase the residence time of water in the estuary and lead to more frequent symptoms of eutrophication; i.e. anoxia associated with dense phytoplankton blooms. If phosphorus was better managed in the catchment of the Sundays River, it is likely that the frequency of phytoplankton blooms and the risks associated with eutrophication could be reduced. Records of long-term datasets need to be kept so that the range of the natural variation between and within estuaries on a seasonal and interannual basis can be established. By doing so, the different types of estuaries can be better understood and efficiently managed. For the current study, difficulties were experienced in terms of using raw data from studies previously completed on the Sundays Estuary for comparison purposes.
Acknowledgements
Funding for this study was obtained from NMMU, Vodacom and the National Research Foundation (NRF). Thanks are due to the NMMU Department of Botany for providing facilities and equipment, Mrs Patricia Smailes for assisting with the taxonomic identification of the diatoms and Mr Pascal Tabot for assisting with the statistical analyses.
References
ADAMS JB and BATE GC (1994) The freshwater requirements of estuarine plants incorporating the development of an estuarine decision support system. WRC Report No. 292/1/94. Water Research Commission, Pretoria. [ Links ]
ADAMS JB and BATE GC (1999) Primary producers. Estuarine microalgae. In: Allanson BR and Baird D (eds.) Estuaries of South Africa. Cambridge University Press, Cambridge. 91-101. [ Links ]
ADAMS JB, WOOLDRIDGE T, COWLEY P, SNOW GC, STRYDOM N, TALJAARD S, VAN NIEKERK L, WHITFIELD AK and WHITTINGTON P (2008) Ecological Water Requirements study, Intermediate level, Sundays Estuary. Report prepared for SANParks by the IECM of the Nelson Mandela Metropolitan University. IECM Report No. C151.1. 115 pp. excl. appendices. [ Links ]
ALLANSON BR and READ GHL (1995) Further comment on the response of Eastern Cape Province estuaries to variable freshwater inflows. S. Afr. J. Aquat. Sci. 21 56-70. [ Links ]
ANANDRAJ A, PERISSINOTTO R, NOZAIS C and STRETCH D (2008) The recovery of microalgal production and biomass in a South African temporarily open/closed estuary, following mouth breaching. Estuar. Coast. Shelf Sci. 79 599-606. [ Links ]
ARCHIBALD REM (1981) An Investigation into the Taxonomy of the Diatoms Bacillariophyta of the Sundays and Great Fish Rivers with Ecological Observations in Sundays. Ph.D. Thesis, Rhodes University, Grahamstown. [ Links ]
BATE GC, SMAILES PA and ADAMS JB (2004) Benthic diatoms in the rivers and estuaries of South Africa. WRC Report No. TT 234/04. Water Research Commission, Pretoria. 32 pp. [ Links ]
CARSTENSEN J, CONLEY DJ and HENRIKSEN P (2004) Frequency, composition, and causes of summer phytoplankton blooms in a shallow coastal ecosystem, the Kattegat. Limnol. Oceanogr. 49 (1) 190-201. [ Links ]
CHOU Y, POLANSKY AM and MASON RL (1998) Transforming nonnormal data to normality in statistical process control. J. Qual. Technol. 30 133-141. [ Links ]
CLOERN JE (1991) Tidal stirring and phytoplankton bloom dynamics in an estuary. J. Mar. Res. 49 203-221. [ Links ]
COUNCIL FOR SCIENTIFIC AND INDUSTRIAL RESEARCH (CSIR) (2002) Analytical Methods Manual. Estuarine and marine waters, sediment and biological tissue. CSIR Technical Report ENV/S-I 97011. CSIR, Stellenbosch, South Africa. 133 pp. [ Links ]
DOMINGUES RB, BARBOSA A and GALVÁO H (2005) Nutrients, light and phytoplankton succession in a temperate estuary (the Guadiana, south-western Iberia). Estuar. Coast. Shelf Sci. 64 249-260. [ Links ]
FARNμM NR (1996) Using Johnson Curves to describe non-normal process data. Qual. Eng. 9 329-336.
FRONEMAN PW (2000) Feeding studies on selected zooplankton in a temperate estuary, South Africa. Estuar. Coast. Shelf Sci. 51 543-552. [ Links ]
FRONEMAN PW (2002) Response of the plankton to three different hydrological phases of the temporarily open/closed Kasouga Estuary, South Africa. Estuar. Coast. Shelf Sci. 55 535-546. [ Links ]
FRONEMAN PW (2006) The importance of phytoplankton size in mediating trophic interactions within the plankton of a southern African estuary. Estuar. Coast. Shelf Sci. 70 693-700. [ Links ]
GAMA PT, ADAMS JB, SCHAEL DM and SKINNER T (2005) Phytoplankton chlorophyll a concentration and community structure of two temporarily open/closed estuaries. WRC Report No. 1255/1/05. Water Research Commission, Pretoria. 91 pp. [ Links ]
GAMEIRO C and BROTAS V (2010) Patterns of phytoplankton variability in the Tagus Estuary (Portugal). Estuar. Coasts 33 311-323. [ Links ]
GRANGE N, WHITFIELD AK, DE VILLIERS CJ and ALLANSON BR (2000) The response of two South African east coast estuaries to altered river flow regimes. Aquat. Conserv.: Mar. Freshwater Ecosyst. 10 155-17. [ Links ]
HILMER T (1990) Factors Influencing the Estimation of Primary Productivity in Small Estuaries. Ph.D. Thesis, University of Port Elizabeth. 154 pp. [ Links ]
HILMER T and BATE GC (1991) Vertical migration of a flagellate-dominated bloom in a shallow South African estuary. Bot. Mar. 34 113-121. [ Links ]
JERLING HL (1994) Feeding Ecology of Mesozooplankton in the Sundays River Estuary. Ph.D. Thesis, Department of Zoology, University of Port Elizabeth, Port Elizabeth. [ Links ]
JERLING HL and WOOLDRIDGE TH (1995) Relatively negative δ13C ratios of mesozooplankton in the Sundays River estuary, comments on potential carbon sources. S. Afr. J. Aquat. Sci. 21 71-77. [ Links ]
KIBIRIGE I and PERISSINOTTO R (2003) In situ feeding rates and grazing impact of zooplankton in a South African temporarily open estuary. Mar. Biol. 142 357-367. [ Links ]
LASSEN M.F, BRAMM ME, RICHARDSON K, YUSOFF F and SHARIFF M (2004) Phytoplankton community composition and size distribution in the Langat River Estuary, Malaysia. Estuaries 27 (4) 716-727. [ Links ]
LUCAS LV, KOSEFF JR, MONISMITH SG, CLOERN JE and THOMPSON JK (1999) Process governing phytoplankton blooms in estuaries. II: The role of horizontal transport. Mar. Ecol. Prog. Ser. 187 17-30. [ Links ]
MACINTYRE HL, GEIDER RJ and MILLER DC (1996) Micro- phytobenthos: The ecological role of the "Secret Garden" of unveg-etated, shallow-water marine habitats. I. Distribution, abundance and primary production. Estuaries 19 (2A) 186-201. [ Links ]
MACKAY HM (1988) Water Structures and Circulation in the Sundays River Estuary. M.Sc. Thesis, University of Port Elizabeth, Port Elizabeth. 214 pp. [ Links ]
MACKAY HM and SCHμMANN EH (1990) Mixing and circulation in the Sundays River Estuary, South Africa. Estuar. Coast. Shelf Sci. 31 203-216. [ Links ]
MARGALEF R (1978) Life-forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta 1 493-509. [ Links ]
NINHAM SHAND (2008) A Hydrological Study of the Sundays River Estuary. WRC Report No. K8/780/2. Water Research Commission, Pretoria. 7 pp. [ Links ]
NOZAIS C, PERISSINOTTO R and TITA G (2005) Seasonal dynamics of meiofauna in a South African temporarily open/closed estuary (Mdloti Estuary, Indian Ocean). Estuar. Coast. Shelf Sci. 62 325-338. [ Links ]
NUSCH EA (1980) Comparison of different methods for chlorophyll and phaeopigment determination. Archiv fur Hydrobiologie, Beiheft Ergebnise der Limnologie 14 14-36. [ Links ]
PAERL HW, VALDES LM, ADOLF JE and HARDING Jr. LW (2006) Anthropogenic and climatic influences on the eutrophication of large estuarine ecosystems. Limnol. Oceanogr. 51 448-462. [ Links ]
PERISSINOTTO R, IYER K and NOZAIS C (2006) Response of microphytobenthos to flow and trophic variation in two South African temporarily open/closed estuaries. Bot. Mar. 49 10-22. [ Links ]
PRESCOTT GW (1970) How to know: The Freshwater Algae. Wm. C. Brown Company Publishers, Dubuque. Lowa. 348 pp. [ Links ]
REDDERING JSV and ESTERHUYSEN K (1981) Sedimentation in the Sundays Estuary. Rosie Report No. 3. Department of Geology. University of Port Elizabeth. [ Links ]
REVELANTE N and GILMARTIN M (1978) Characteristics of microplankton and nanoplankton communities of an Australian coastal plain estuary. Aust. J. Mar. Freshwater Res. 29 9-18. [ Links ]
ROUND FE (1981) The Ecology of Algae. Cambridge University Press, London. 653 pp. [ Links ]
SCHARLER UM (2000) Response of Nutrient Status and Biotic Communities to Variable Freshwater Input Regimes in Eastern Cape Estuaries, South Africa, Including a Network of Analysis Approach. Ph.D. thesis, University of Port Elizabeth, Port Elizabeth. South Africa. 269 pp. [ Links ]
SNOW GC (2007) Contributions to the Use of Microalgae in Estuarine Freshwater Reserve Determinations. PhD thesis. Nelson Mandela Metropolitan University. South Africa. 249 pp. [ Links ]
SÜNDBACK K, MILES A, HULTH S, PIHL L, ENGSTROM P, SELANDER E and SVENSON A (2003) Importance of benthic nutrient regeneration during initiation of macroalgal blooms in shallow bays. Mar. Ecol. Prog. Ser. 246 115-126. [ Links ]
SÚNDBACK K, LINARES F, LARSON F and WULFF A (2004) Benthic nitrogen fluxes along a depth gradient in a microtidal fjord: The role of denitrification and microphytobenthos. Limnol. Oceanogr. 49 1095-1107. [ Links ]
TALJAARD S, VAN NIEKERK L and SONO S (2008) Ecological water requirements study: Sundays Estuary. Specialist Report: Hydrodynamics and water quality. Appendix B. CSIR, Stellen-bosch. 40 pp. [ Links ]
THOMAS CM, PERISSINOTTO R and KIBIRIGE I (2005) Phyto-plankton biomass and size structure in two South African eutrophic, temporarily open/closed estuaries. Estuar. Coast. Shelf Sci. 65 223-238. [ Links ]
TOMAS CR (ed.) (1997) Identifying Marine Phytoplankton. Florida Marine Research Institute, St. Petersburg. 858 pp. [ Links ]
TURPIE JK, ADAMS JB, JOUBERT A, HARRISON TD, COLLOTY BM, MAREE RC, WHITFIELD AK, WOOLDRIDGE TH, LAMBERTH SJ, TALJAARD S and VAN NIEKERK L (2002) Assessment of the conservation priority status of South African estuaries for use in management and water allocation. Water SA 28 191-206. [ Links ]
UNDERWOOD GJC, PATERSON DM and PARKER RJ (1995) The measurement of microbial carbohydrate exopolymers from intertidal sediments. Limnol. Oceanogr. 40 1243-1253. [ Links ]
UNDERWOOD GJC and KROMKAMP J (1999) Primary production of phytoplankton and microphytobenthos in estuaries. Adv. Ecol. Res. 29 94-155. [ Links ]
VAN DEN HOEK C, MANN DG and JAHNS HM (1995) Algae: An Introduction to Phycology. Cambridge University Press, United K ingdom. 623 pp. [ Links ]
VERITY P and BORKMAN D (2010) A decade of change in the Skidaway River Estuary. III. Plankton. Estuar. Coast. 33 513-540. [ Links ]
WALKER DR (2003) Plant and Algal Distribution in Response to Environmental Variables in Selected Eastern Cape Estuaries. Ph.D. Thesis, University of Port Elizabeth, Port Elizabeth. 176 pp. [ Links ]
WHITFIELD AK, ADAMS JB, BATE GC, BEZUIDENHOUT K, BORNMAN TG, COWLEY PD, FRONEMAN PW, GAMA PT, JAMES NC, MACKENZIE B, RIDDIN T, SNOW GC, STRYDOM NA, TALJAARD S, TERORDE AI, THERON AK, TURPIE JK, VAN NIEKERK L, VORWERK PD and WOOLDRIDGE TH (2008) A multidisciplinary study of a small, temporarily open/ closed South African estuary, with particular emphasis on the influence of mouth state on the ecology of the system. Afr. J. Mar. Sci. 30 453-473. http://dx.doi.org/10.4314/wsa.v38i23 Available on website http://www.wrc.org.za ISSN 0378-4738 (Print) = Water SA Vol. 38 No. 2 April 2012 ISSN 1816-7950 (On-line) = Water SA Vol. 38 No. 2 April 2012 [ Links ]
Received 6 June 2011;
Accepted in revised form 2 April 2012.
* To whom all correspondence should be addressed. imagem aqui +27 41 504-2429; fax: +27 41 583-2317; e-mail: Janine.Adams@nmmu.ac.za


{kind=link}
{kind=link}
{kind=link}