










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkWater SA
versão On-line ISSN 1816-7950
versão impressa ISSN 0378-4738
Water SA vol.37 no.2 Pretoria Abr. 2011
Local and regional factors influencing zooplankton communities in the connected Kasseb Reservoir, Tunisia
Ikbel SellamiI,*; Jannet ElloumiI; Asma HamzaII; Mohammed Alaoui MhamdiIII; Habib AyadiI
IUniversité de Sfax, Faculté des Sciences de Sfax, Département des Sciences de la Vie. Unité de recherche LR/UR/05ES05 Biodiversité et Ecosystèmes Aquatiques. Route soukra Km 3.5 - BP 1171 - CP 3000 Sfax, Tunisie
IIInstitut National des Sciences et Technologie de la Mer, Centre de Sfax BP 1035 Sfax 3018, Tunisie
IIIUniversité Sidi-Mohammed-Ben-Abdallah, Département de Biologie, Laboratoire LAMEC, Faculté des sciences, BP 1796, Atlas, Fès, Maroc
ABSTRACT
Associations between zooplankton community structure and abiotic (temperature, dissolved oxygen, turbidity, nutriments) and biotic factors (chlorophyll a and phytoplankton community) were examined, in Kasseb Reservoir, northern Tunisia. Samples were taken bimonthly from July to December 2002 at 3 sampling stations (deepest station: Station 1, Brik River: Station 2 and M'Zaz Stama River: Station 3). From our results it is evident that zooplankton exhibit seasonally and spatially heterogeneous distribution. The highest density of zooplankton was recorded in September at a depth of 5 m (10.8 × 103 ind·ℓ-1). At Station 1 cyclopoid copepods (65% of total abundance) were the most abundant group followed by Cladocera (21% of total abundance). At Station 2 (93% of total abundance) and Station 3 (98% of total abundance) cyclopoid copepods were numerically dominant throughout the study period. Canonical correspondence analysis (CCA) was used to estimate the influence of abiotic and biotic factors in structuring the zooplankton assemblage. Zooplankton abundance was negatively correlated with turbidity (r= -0.381, P <0.05). The results also suggest that both local (environmental parameters, competition, and predation) and regional (hydrologic connections and dispersal) factors have a significant effect on both species richness and community structure of zooplankton in Kasseb Reservoir. The presence of zooplankton species con sidered to be indicators of eutrophic status confirmed the high trophic levels of Kasseb Reservoir.
Keywords: Kasseb Reservoir, hydrologic connections, local and regional factors, zooplankton, heterogeneous distribution
Introduction
The Kasseb Reservoir is an important feature of Tunisia, and was built to serve a multitude of purposes, such as hydroelectric production, irrigation, and, principally, drinking water supply (30% of the population of Tunis city). The Kasseb Reservoir is directly connected to Ghdir El Goulla Reservoir (see Sellami et al., 2009).
In freshwater ecosystems the filter-feeding zooplankton play an important role in the production of the clear-water phase (Lair and Ayadi, 1989; Edmondson, 1991; Lampert and Sommer, 1997; Stella et al., 2007). These organisms are important in the structuring and dynamics of aquatic environments, as is their fundamental role in aquatic food chains (Cadjo et al., 2007) and nutrient cycling (Lansac-Tôha et al., 1997; Velho, 2000; Gillooly and Dodson, 2000). The factors affecting community composition and species diversity are often divided into 2 general categories: local processes (e.g., environmental heterogeneity and species interactions) and regional processes (e.g., dispersal, connectivity) (Ricklefs, 1987; Mouquet and Loreau, 2003; Holyoak et al., 2005). Some important local factors influencing zooplankton species richness and abundance have been identified: lake area and primary productivity (Dodson, 1991; 1992; Dodson et al., 2000), water quality (Jeppesen et al., 2000; Cottenie et al., 2001), lake depth (Keller and Conlon, 1994), latitude (Hebert and Hann, 1986), acidity (Brezonik et al., 1984; Locke, 1992), nutrients (Leibold, 1999; Jeppesen et al., 2000), toxins (Yan et al., 1996), climate (Stemberger et al., 1996), predation and competition (Brooks and Dodson, 1965; Shurin, 2000; Fernández-Rosado and Lucena, 2001; Isari et al., 2007; Larson et al., 2009). Connectivity can influence several community properties, such as local and regional diversity, and secondary productivity (e.g., Cottenie et al., 2001; Gonzalez and Chaneton, 2002; Cottenie and De Meester, 2004; Vanschoenwinkel et al., 2007). The connectivity of ecosystems is expected to influence dispersal rates and induce passive dispersal within meta-communities (Doi et al., 2010).
Abiotic and biotic processes commonly also cause lakewide distributions of plankton to be highly variable and heterogeneous, resulting in considerable patchiness (Folt and Burns, 1999). Heterogeneous distributions of zooplankton originate from various processes, some of which can be attributed to internal factors; that is, they are related to the activity and swimming behaviour of the animals (Folt and Burns, 1999). Other causes are linked to external factors, which force organisms to be transported passively, such as by wind-driven currents (George and Edwards, 1976; George and Winfield, 2000; Rinke et al., 2006; 2009).
To our knowledge, no published research has previously addressed these issues in Mediterranean ecosystems that have been artificially connected by human alteration. We therefore conducted a study to test the hypothesis that anthropogenic influence on the connectivity of ecosystems would affect zooplankton community dynamics. In Kasseb Reservoir, the artificially connected system, zooplankton communities were related to different local and regional variables, in order to determine the important structuring variables. We studied the vertical and horizontal heterogeneity of zooplankton commu nities in this reservoir, in order to define the principal factors responsible for zooplankton distribution.
Materials and methods
Study site
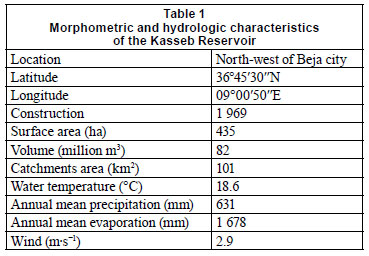
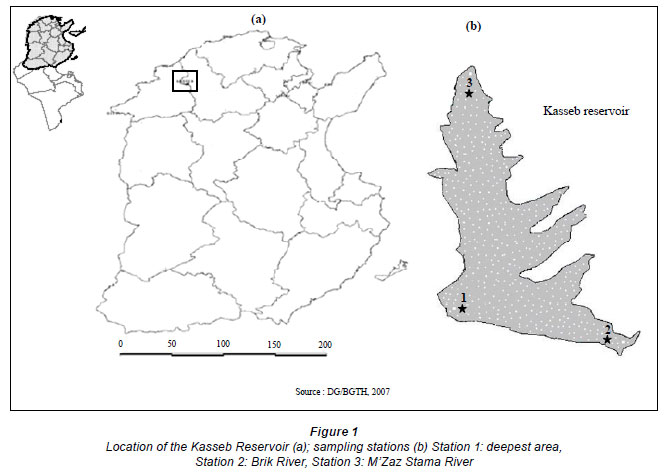
Kasseb Reservoir (K) is situated 18 km north-west of Beja city (between 36º45'30'N and 09º00'50'E) (Fig. 1). This reservoir receives water through the Brik (north- eastern part of Kasseb Reservoir, 27.9 km2) and M'Zaz Stama Rivers (western part of Kasseb Reservoir, 73.1 km2), the catchments of which are densely populated, with the rivers mainly used for agricultural purposes. Morphometric and other basic characteristics are shown in Table 1.
Environmental variables
In this study, samples were taken bimonthly from 3 stations in Kasseb Reservoir, between July and December 2002. Two replicates were taken at each depth/station. The 3 stations were selected as follows: Station 1 located at the deepest area, Station 2 (Brik River) and Station 3 (M'Zaz Stama River) located at nearly 25 and 35 km, respectively, from the deepest area. Water samples were collected at the surface, -5 m, -10 m, -20 m (near the bottom) at Station 1 and close to the surface at Station 2 and Station 3, with a 1 ℓ Van Dorm bottle, simultaneously for physical and chemical analyses (Fig. 1).
Water temperature was measured with a mercury glass thermometer. The dissolved oxygen concentration was measured by means of Winkler methods (Rodier, 1984). The turbidity was measured with a portable turbidimeter. Total nitrogen was assayed after oxidation to the nitrate form in an alkaline medium, using potassium persulphate. The nitrate assay was based on the formation, in a concentrated sulphuric acid medium, of a phenol disulphonic acid derivative (D'Elia et al., 1977). The total phosphorus concentrations were determined after mineralising the samples using the colorimetric protocol of Murphy and Riley (1962).
Biological variables
Phytoplankton
For phytoplankton cell counts (Fathalli, 2004), water was taken using a 1 ℓ Van Dorn bottle and fixed with Lugol solution (Schwoerbel, 1986) for counting, according to Utermöhl (1958). The taxonomic identification was done according to Bourrelly (1966; 1968; 1985), Baker (1991; 1992) and Shameel (2001). Sub-samples (0.5 ℓ) for quantification of chlorophyll a were filtered using Whatman GF/C filters (0.45 µm pore size filter and 25 mm- diameter) and the quantity of pigments was deter mined using a fluorometric method (Welschmeyer, 1994) after a methanol extraction (Herbland et al., 1985).
Zooplankton
Zooplankton samples were collected by filtering 50 ℓ (with a 1 ℓ Van Dorn bottle) through a Juday plankton net (mesh size 55 µm), preserved with 4% formalin and coloured with Bengal Pink. Two replicates were taken at each depth/station. The samples were only collected during the day - diel vertical migrations of zooplankton were not considered in this study. Therefore the abundance of the zooplankton may be underestimated. The zooplankton were identified and counted under a Leica binocular microscope, in Dolffus chambers. The taxonomic identification was carried out according to Amoros (1984), Margaritora (1985), Korovchinsky (1992), Dussart (1969) and Stella (1982). The zooplankton density was expressed as the number of individuals per sample volume (ind·ℓ-1). The zooplankton community structure was studied by calculating the species diversity index H' (bits ind·ℓ-1) (Shannon and Weaver, 1949); it is the most popular index (Andronikova, 1993).
This index was calculated from the density of zooplankton species:
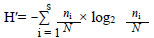
where:
ni is the density of i species
N is the density of the entire community, respectively.
Statistical analysis
Canonical correspondence analysis (CCA) was applied to physical (water temperature and dissolved oxygen), chemical (total nitrogen and total phosphorus) and biological parameters (chlorophyll a, phytoplankton and zooplankton) assessed over 12 observations. Simple log (x + 1) transformation was applied to data in order to correctly stabilise the variance (Frontier, 1973). Pearson's correlation coefficient was calculated to determine the association between the physico-chemical variables and the zooplankton community (Zar, 1999). A multiple regression was applied in order to investigate what variables may predict zooplankton abundance. Additionally, an independent 1-way ANOVA was run to assess the effect of the temporal and spatial variation on zooplankton density.
Results
Environmental parameters
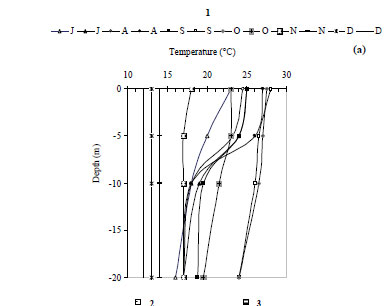
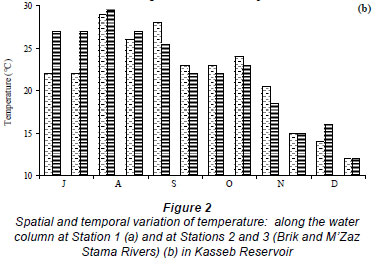
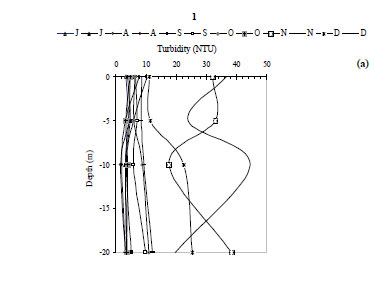
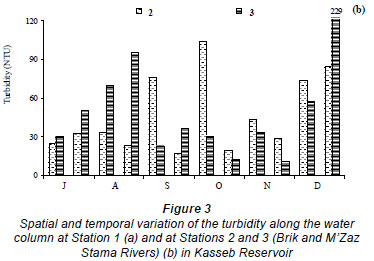
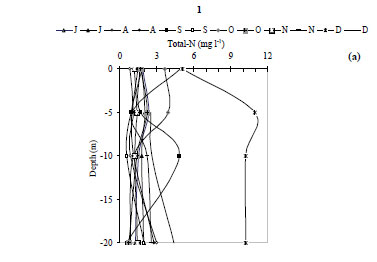
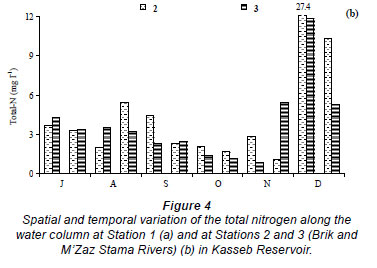
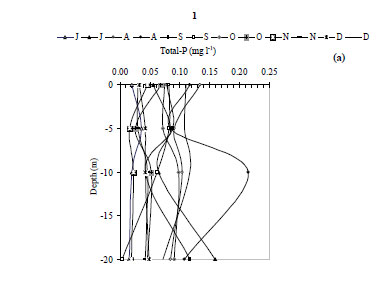
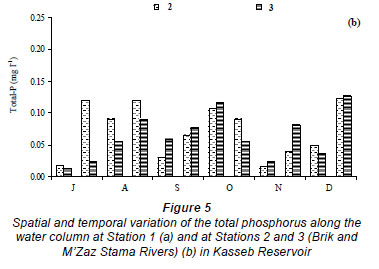
The water temperature in the Kasseb Reservoir varied from 12ºC, in December along the water column, to 28ºC, in September at the surface (mean ± s.d. = 19.6 ± 5.1ºC) (Fig. 2a), with spatial and temporal fluctuations typical to those of a warm monomictic reservoir. The temperature of the water column increased markedly from July to September. At other stations the water temperature showed a maximum in August (Station 3; 29.5ºC). The minimum values were recorded in December (12ºC) for all stations (Fig. 2b). The values for dissolved oxygen concentration measured at the surface were 8.8 ± 1.5 mg·ℓ-1 (at Station 1), 8.4 ± 0.5 mg·ℓ-1 (at Station 2) and 7.6 ± 0.3 mg·ℓ-1 (at Station 3). In the deepest area, turbidity ranged from 1.6 NTU, in August at a depth of 10 m, to 44.6 NTU in December at a depth of 10 m (mean ± s.d. = 10.8 ± 10.7 NTU) (Fig. 3a). However, for Station 3 the maximum value of 229 NTU was reported in December (Fig. 3b). The total nitrogen concentrations varied from 0.5 mg·ℓ-1 in September at a depth of 10 m to 10.9 mg·ℓ-1 in December at a depth of 5 m (mean ± s.d. = 2.5 ± 3.4 mg·ℓ-1) (Fig. 4a). The highest total nitrogen concentration was recorded in December at Station 2 (27.4 mg·ℓ-1) (Fig. 4b). The highest values for total nitrogen concentration were probably the result of inputs arriving at the reservoir from the surface of each catchment. At Station 1, the total phosphorus concentrations fluctuated between 0.003 mg·ℓ-1 in September at a depth of 20 m and 0.2 mg·ℓ-1 in August at a depth of 10 m (mean ± s.d. = 0.06 ± 0.04 mg·ℓ-1) (Fig. 5a). For the other 2 stations, the maximum total phosphorus concentration was registered in December at Station 3 (0.1 mg l-1) (Fig. 5b). Significant differences between stations were found for dissolved oxygen (F = 3.91, df = 35, P = 0.05) and turbidity (F = 3.76, df = 35, P = 0.05). Temperature (F = 5.28, df = 11, P = 0.05), total nitrogen (F = 5.53, df = 11, P = 0.05) and total phosphorus (F = 6.04, df = 11, P = 0.05) differed significantly between months (Table 2).
Spatial and temporal distribution of phytoplankton
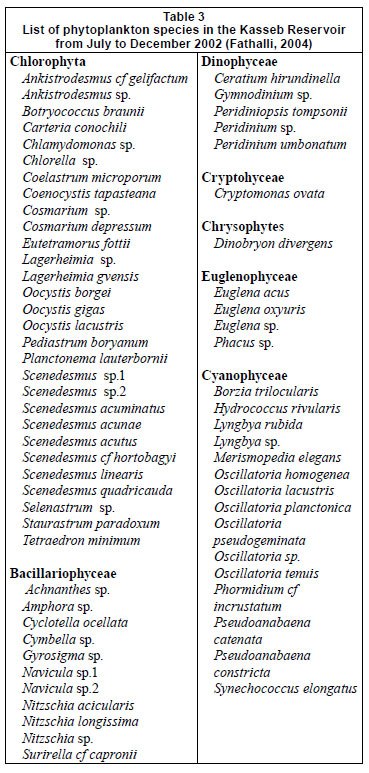
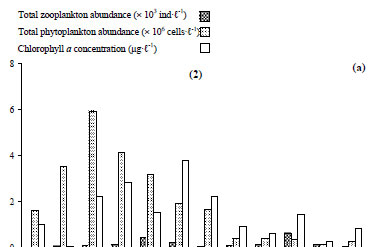
For the 7 groups of phytoplankton, 66 species were identified from the samples at the 3 stations (Table 3). Numerous phytoplankton species were distributed among the major taxonomic categories as follows: Chlorophyta 29, Bacillariophyceae 11, Dinophyceae 5, Chrysophytes1, Cryptohyceae 1, Cyanophyceae 15 and Euglenophyceae 4. The contribution to total cell phytoplankton density was: Bacillariophyceae 59%, Chlorophyta 22%, cyanobacteria 15% and other taxa 4% (Fathalli, 2004). The phytoplankton community was more abundant at Station 2 than at other stations. At Station 2 total density varied from 0.1 × 106 in early December to 5.9 × 106 cells·ℓ-1 in early August (mean ± s.d. = 1.9 × 106 ± 1.8 × 106 cells·ℓ-1) (Fig. 7a). Bacillariophyceae (75% of the total phytoplankton abundance) were numerically dominated by the species Cyclotella ocellata (64.6% of the total abundance).
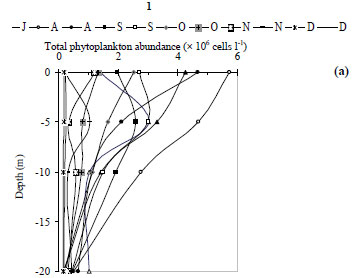
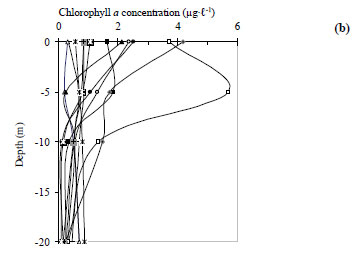
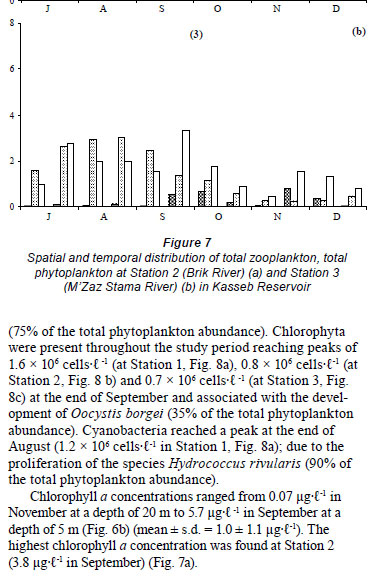
At Station 1, the total phytoplankton abundance ranged from 0.1 × 106 cells·ℓ-1 in July at a depth of 20 m to 5.7 × 106 cells·ℓ-1 in early August in the surface (Fig. 6a) (mean ± s.d. = 1.3 × 106 ± 1.3 × 106 cells·ℓ-1). Bacillariophyceae was the dominant group, accounting for 73% of the total phytoplankton abundance (Fig. 8a). The most abundant species was Cyclotella ocellata (79.2% of the total abundance). At Station 3, the total phytoplankton density ranged from 0.2 × 106 at end- November to 3.0 × 106 cells l·ℓ-1 at end- August (mean ± s.d. = 1.4 × 106 ± 1.0 × 106 cells·ℓ-1) (Fig. 7b). Bacillariophyceae (67% of the total phytoplankton, Fig. 8c) were largely dominated by the species Cyclotella ocellata(75% of the total phytoplankton abundance). Chlorophyta were present throughout the study period reaching peaks of 1.6 × 106 cells·ℓ-1 (at Station 1, Fig. 8a), 0.8 × 106 cells·ℓ-1 (at Station 2, Fig. 8 b) and 0.7 × 106 cells·ℓ-1 (at Station 3, Fig. 8c) at the end of September and associated with the development of Oocystis borgei (35% of the total phytoplankton abundance). Cyanobacteria reached a peak at the end of August (1.2 × 106 cells·ℓ-1 in Station 1, Fig. 8a); due to the proliferation of the species Hydrococcus rivularis (90% of the total phytoplankton abundance).
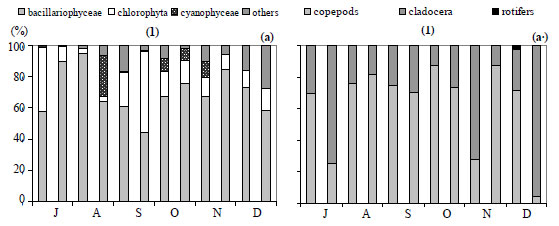

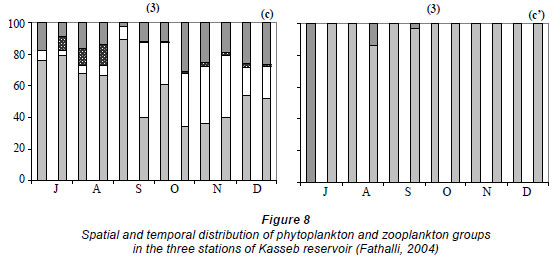
Chlorophyll a concentrations ranged from 0.07 µg·ℓ-1 in November at a depth of 20 m to 5.7 µg·ℓ-1 in September at a depth of 5 m (Fig. 6b) (mean ± s.d. = 1.0 ± 1.1 µg·ℓ-1). The highest chlorophyll a concentration was found at Station 2 (3.8 µg·ℓ-1 in September) (Fig. 7a).
Spatial and temporal distribution of zooplankton
A total of 4 cladocerans (Bosmina longirostris, Diaphanosoma brachyurum, Daphnia longispina, and Ceriodaphnia quadrangula), 3 copepods (Copidodiaptomus numidicus, Eudiaptomus sp., and Acanthocyclops sp.) and 1 rotifer (Keratella quadrata) were found in Kasseb Reservoir (Table 3). At Station 1, cyclopoid copepods (65% of total abundance) were the most abundant group (Acanthocyclops sp., 67.3% of total abundance), followed by Cladocera (21% of the total zooplankton abundance), which was represented by the species Bosmina longirostris (32.6% of the total zooplankton abundance). At Station 2 (93% of total abundance) and Station 3 (98% of the total zooplankton abundance) cyclopoid copepods (Acanthocyclops sp.) were numerically dominant throughout the study period. Calanoid copepods were absent at Station 2. The species Eudiaptomus sp., Ceriodaphnia quadrangula and Keratella quadrata were present only at Station 1. Significant differences between stations and months were found for the abundances of the following species: Acanthocyclops sp. (F = 12.7, df = 35, P = 0.001; F = 6.57, df = 11, P = 0.01), Eudiaptomus sp. (F = 6.26, df = 35, P = 0.001), Bosmina longirostris (F = 3.25, df = 35, P = 0.05), Diaphanosoma brachyurum (F = 6.76, df = 35, P = 0.01; = 3.01, df = 35, P = 0.01), Daphnia longispina (F = 5.22, df = 35, P = 0.01; F =3.59, df = 35, P = 0.01) and Ceriodaphnia quadrangula (F = 11.24, df = 35, P = 0.001; F = 4.91, df = 35, P = 0.05) (Table 4).
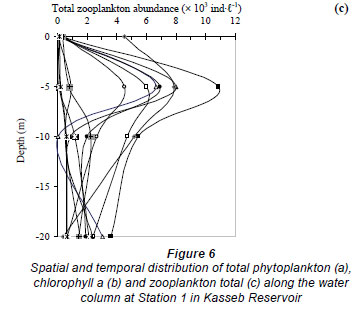
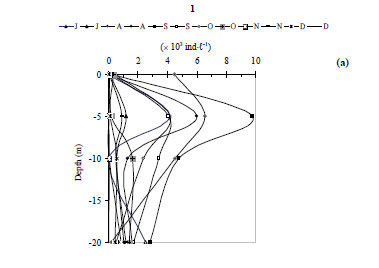
At Station 1, the total zooplankton abundance varied from 0.04 × 103 ind·ℓ-1 in July at a depth of 10 m to 10.8 × 103 ind·ℓ-1 in early September at a depth of 5 m (mean ± s.d. = 2.2 × 103 ± 2.6 × 103 ind·ℓ-1) (Fig. 6c). The zooplankton abundance was significantly positively correlated with water temperature (r = 0.779, P <0.01), and with dissolved oxygen (r = 0.578, P <0.05) and significantly negatively correlated with turbidity (r = - 0. 381, P <0.05). The zooplankton community was dominated by the cyclopoid Acanthocyclops sp. (58 - 83% of the total zooplankton abundance, H'= 0.57 bits ind·ℓ-1 to 1.59 bits ind·ℓ-1) (Fig. 8a', Fig. 9a, Fig. 10a), with the exception of late July, early November and late December, which were dominated by Cladocera: 75% (H'= 1.28 bits ind·ℓ-1), 72% (H'= 1.29 bits ind·ℓ-1) and 96% (H'= 0.47 bits ind·ℓ-1) of the total zooplankton abundance, respectively. The species Bosmina longirostris was responsible for the exceptional peak that occurred in late July, accounting for 56% of the total zooplankton abundance, whereas the species Daphnia longispina accounted for 74% and 97% of the total zooplankton abundance in early November and late December, respectively.
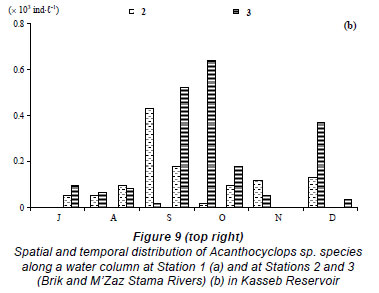
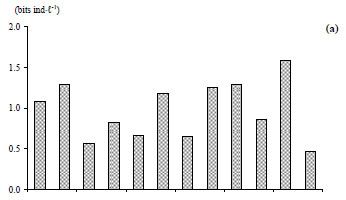
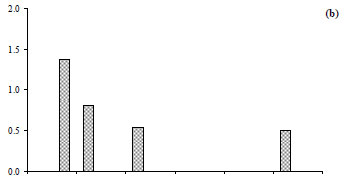
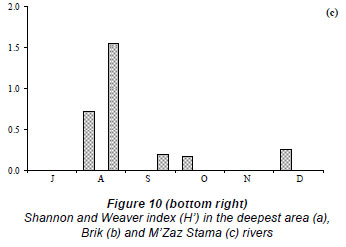
The total zooplankton abundance ranged between 0.01 × 103 ind∙ℓ-1 in early July and 0.6 × 103 ind∙ℓ-1 in late November (mean ± s.d. = 0.15 × 103 ± 0.18 × 103 ind∙ℓ-1) at Station 2 (Fig. 7a) and between 0.01 × 103 ind·ℓ-1 in early July and 0.8 × 103 ind·ℓ-1 in late November at Station 3 (mean ± s.d. = 0.24 × 103 ± 0.27 × 103 ind·ℓ-1) (Fig. 7b). At Station 2, the zooplankton community was largely dominated by Acanthocyclops sp. throughout the study period (Fig. 8b') (60 to 100% of total zooplankton abundance, H'= 0.00 bits ind·ℓ-1 to 1.37 bits ind·ℓ-1, Fig. 9b, Fig. 10b), with the exception of late December which was dominated by Daphnia longispina (accounting for 100% of the total zooplankton abundance). At Station 3, Acanthocyclops sp. (71 to 100% of the total zooplankton abundance, H'= 0.00 bits ind·ℓ-1 to 1.55 bits ind·ℓ-1, Fig. 10c) was the dominant species throughout the study period (Fig. 8c', Fig. 9b), with the exceptions of early July, which was dominated by the cladoceran Diaphanosoma brachyurum (100% of the total zooplankton abundance), and late November, which was dominated by the calanoid Copidodiaptomus numidicus (100% of the total zooplankton abundance).
Community analysis
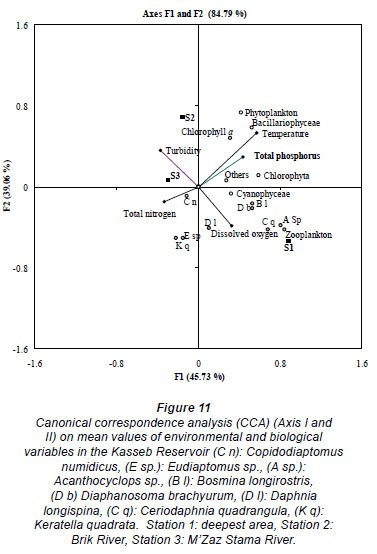
Canonical correspondence analysis (CCA) was performed for total zooplankton, total phytoplankton abundance and environmental variables (Fig. 11).
Discussion
Our study suggests that local and regional processes interact to produce patterns of species composition and diversity of zooplankton in Kasseb Reservoir. In this system of highly connected reservoirs, Kasseb Reservoir provides the water for the Ghdir El Goulla Reservoir (Sellami et al., 2009). Connections between reservoirs can have positive and negative impacts on zooplankton communities. The Kasseb Reservoir has low diversity (8 zooplankton species). The results of this study suggest that this system is structured according to the species-sorting paradigm (Leibold et al., 2004). Connectivity can promote high community diversity by providing new species from the regional pool, but can also limit local diversity by washing out species, setting up local communities for competitive exclusion, and otherwise providing a disturbance that can offset equilibrium conditions (Cottenie and DeMeester, 2005).
The highest zooplankton densities were found in early September at a depth of 5 m (10.8 × 103 ind·ℓ-1) for the deep southern part of the Kasseb Reservoir (Station 1), but in late November for the shallow north-eastern (Station 2) and western (Station 3) areas. A significant correlation was found between water temperature and zooplankton abundance at Station 1 (r= 0.779, P <0.01). This accords with previous studies (De Azevedo and Bonecker, 2003; Cherbi et al., 2008; Primo et al., 2009) concluding that temperature influences zooplankton distribution. By contrast, the lowest zooplankton density was found in the rainy period (December). It is likely that the rainfall washes out the zooplankton species from Kasseb to Ghdir El Goulla Reservoir. It has been reported for the floodplain of the Danube River that crustacean zooplankton species richness decreases in floodplain habitats as connection to the river increases (Baranyi et al. 2002). Many studies (Murugavel and Pandian, 2000; Guevara et al., 2009) have recognised that limnological variables associated with rainfall are of importance to the zooplankton community.
The greatest density and diversity of zooplankton was observed at Station 1, due to the relative stability of the wide reservoir body, with slower velocity of water flow relative to the 2 rivers (Stations 2 and 3). Similar results were found by Zhenbin et al. (2008). Moreover, the low density and diversity (4 species) of zooplankton observed at Stations 2 and 3 could be linked to high turbidity at these stations. Turbidity showed significant spatial differences (P <0.05). The highest values of turbidity observed for Stations 2 and 3 may be due to an increase in sediment load from surface runoff at these stations. Zooplankton abundance was negatively correlated with turbidity (r = -0.381, P <0.05). These findings confirm the results of Dejen el al (2004). Turbidity is a very important structuring variable (Scheffer, 1998) for zooplankton communities.
There was an imbalance between phytoplankton and zooplankton communities. In this reservoir, zooplanktonic organisms were probably able to feed on sources other than phytoplankton (microzooplankton and detritus). These results were in agreement with the observations of Pinto-Coelho et al. (2005) and Morgado et al. (2007), which showed detritus food chains to be a resource for zooplankton. The relationship between phytoplankton and zooplankton was illustrated by a comparison of further changes in density of both communities. Subsequently, each decrease in phytoplankton density was preceded by an increase in zooplankton density; reflecting a state of imbalance in this ecosystem. Similar results have been found in other studies (Abowei et al., 2008; Friedrich and Pohlmann, 2009; Mitra, 2009).
The highest zooplankton density was found at the upper layer (0 to 10 m) with a maximum at 5 m (10.8 × 103 ind·ℓ-1 in the beginning of September). The vertical distribution of zooplankton was similar to that reported in other studies (Cadjo et al., 2007). Such a pattern may have been related to the food availability in the upper layer, where phytoplankton was most abundant. These findings confirm the results of Cherbi et al. (2008). Thus, zooplankton is probably an important grazer of algae during periods of high zooplank ton abundance. This is supported by the findings of a study by Horn and Horn (2008).
There was no clear trophic gradient in Kasseb Reservoir; some horizontal differences in zooplankton abundance and community structure were observed. Copidodiaptomus numidicus was more common in the western part of Kasseb Reservoir. Eudiaptomus sp., Acanthocyclops sp., Bosmina longirostris, Diaphanosoma brachyurum, Daphnia longispina, Ceriodaphnia quadrangula and Keratella quadrata were dominant in the deep southern part of the reservoir. In a similar study conducted in Lake Pyhäselkä by Karjalainen et al. (1996a), Daphnia and Eudiaptomus were dominant in the southern pelagial zone.
The Cladocera community was characterised by the dominance of the smaller species in summer (Bosmina longirostris, Diaphanosoma brachyurum, and Ceriodaphnia quadrangula) and the larger ones in autumn (Daphnia longispina). Daphnia longispina abundance increased during autumn and winter and decreased in summer (Table 3). Similar results have been found for other freshwater systems (Primo et al., 2009).
The development of cyanobacteria in summer (the maximum was observed at the end of August: 1.2 × 106 cells·ℓ-1 at Station 1, the deepest station, Fig. 8a) led to a decrease in Daphnia longispina densities. On the contrary, smaller cladocerans avoided the typically abundant cyanobacteria and fed on smaller algal particles (Paranaguá et al., 2005). These associations were confirmed by the positive relationship between the species Bosmina longirostris, Diaphanosoma brachyurum, and Ceriodaphnia quadrangula and cyanobacteria on the first CCA axis (G1). By contrast, Daphnia longispina seems to exclude itself (G3) (Fig. 10a). Frequently, the replacement of large species by small ones occurs when cyanobacteria are dominant (Leonard and Pearl, 2005; Ferráo-Filho et al., 2009). The PEG model (Plankton Ecology Group, Sommer et al., 1986) reported the replacement of smaller cladoceran species by larger ones in summer (Sommer et al., 1986; Lair and Ayadi, 1989). Many authors confirm competitive mechanisms between Daphnia and Diaphanosoma (Horn and Horn, 1990) and between Daphnia and Bosmina (Urabe, 1990). On the other hand, the low density of Daphnia longispina observed in summer was probably the result of grazing by Chaoborus larvae and planktivorous fish. Many studies have established that planktivorous fish deplete the abundance of large cladocerans such as Daphnia through direct grazing (Pont et al., 1991, Couture et al., 2008).
A positive relationship was shown between the species Acanthocyclops sp., Bosmina longirostris and Diaphanosoma brachyurum on the first CCA axis. Cyclopoids have been observed by some workers (Pinto-Coelho et al., 2005) at high trophic levels. Several studies have reported that Bosmina longirostris was associated with more eutrophic conditions (De Eyto, 2001; Jaramillo and Gaviria, 2003; Guevara et al., 2009). The species Diaphanosoma brachyurum was generally abundant at high trophic levels (Amoros, 1984).
Conclusion
The zooplankton community in Kasseb Reservoir showed spatial and temporal variations in density in relation to local and regional parameters. The phytoplankton community contributes to succession ofzooplankton community. Nevertheless, zooplankton dynamics can be affected by Chaoborus larvae and fish predation.
The presence of zooplankton species known to be indica tors of eutrophic status reflected the high trophic condition of Kasseb Reservoir. In addition, total phosphorus concentration (25-200 µg·ℓ-1) was within the range that is typically found in eutrophic reservoirs. It is therefore important to prevent the deterioration of Kasseb Reservoir, since it is an important drinking water supply for Tunisia.
Acknowledgements
We thank one anonymous reviewer for providing helpful suggestions for improvement. We also thank Mr Nabil Kallel, an English teacher in the Faculty of Science, for helping us in proofreading this article. The authors wish to thank A Fathalli for the phytoplankton data.
References
ABOWEI JFN, TAWARI CC and DAVIES OA (2008) Zooplankton of lower Sombreiro River, Niger delta, Nigeria. J. Fish. Int. 3 (4) 105-111. [ Links ]
AMOROS C (1984) Introduction pratique à la systématique des organismes des eaux continentales françaises. Crustacés cladocères. Bull. Soc. Linn. Lyon. 72-145. [ Links ]
ANDRONIKOVA IN (1993) Lake Classification with Respect to Biological Productivity, in Teoreticheskie voprosy klassifikatsii ozer (Theoretical Problems of Lake Classification), St. Petersburg: Nauka 51-72. [ Links ]
BAKER DR (1991) Interdiffusion of hydrous dacitic and rhyolitic melts and the efficacy of rhyolite contamination by dacitic enclaves. Contrib. Mineral. Petrol. 106 462-473. [ Links ]
BAKER DR (1992) Interdiffusion of geologic melts: calculations using transition state theory. Chem. Geol. 98 11-21. [ Links ]
BARANYI C, HEIN T, HOLAREK C, KECKEIS S and SCHIEMER F (2002) Zooplankton biomass and community structure in a Danube River floodplain system: effects of hydrology. Freshwater Biol. 47 473-82. [ Links ]
BOURRELLY P (1966) Les algues d'eau douce. Vol. 1: Les algues vertes. Editions N. Boubee et Cie, Paris. 512 pp. [ Links ]
BOURRELLY P (1968) Les algues d'eau douce. Vol. II: Les algues jaunes et brunes, Chrysophycees, Pheophycees, Xanthophycees et Diatomees. Editions N. Boubee et Cie Paris. 440 pp. [ Links ]
BOURRELLY P (1985) Les algues d'eau douce. Tome III: les algues bleues et rouges. Paris, France, Boubée et Cie. 606 pp. [ Links ]
BREZONIK PL, CRISMAN TL and SCHULZE RL (1984) Planktonic communities in Florida softwater lakes of varying pH. Can. J. Fish. Aquat. Sci. 41 46-56. [ Links ]
BROOKS JL and DODSON SI (1965) Predation, body size, and composition of zooplankton. Sci. 150 28-35. [ Links ]
CADJO S, MILETIC A and DJURKOVIC A (2007) Zooplankton of the Potpec reservoir and the saprobiological analysis of water quality. Desalination 213 24-28. [ Links ]
CHERBI M, LEK-ANG S, LEK S and ARAB A (2008) Distribution du zooplancton dans les lacs à climat méditerranéen. C. R. Biologies 331 692-702. [ Links ]
COTTENIE K and DE MEESTER L (2004) Metacommunity structure: synergy of biotic interactions as selective agents and dispersal as fuel. Ecol. 85 114-119. [ Links ]
COTTENIE K and DEMEESTER L (2005) Local interactions and local dispersal in a zooplankton metacommunity. 189-211. In: Holyoak M, Leibold MA and Holt RD (eds.) Metacommunities: Spatial Dynamics and Ecological Communities. The University of Chicago Press, Chicago, IL, USA. [ Links ]
COTTENIE K, MICHELS E, NUYTTEN N and DE MEESTER L (2001) Zooplankton community structure and environmental conditions in a set of interconnected ponds. Hydrobiologia 442 39-350. [ Links ]
COUTURE SC and WATZIN MC (2008) Diet of Invasive Adult White Perch (Morone americana) and their Effects on the Zooplankton Community in Missisquoi Bay, Lake Champlain. J. Great Lakes Res. 34 485-494. [ Links ]
D'ELIA CF, STEUDLER PA and CORWIN N (1977) Determination of total nitrogen in aqueous samples using persulfate digestion. Limnol. Oceanogr. 22 760-764. [ Links ]
DE AZEVEDO F and BONECKER CC (2003) Community size structure of zooplanktonic assemblages in three lakes on the upper River Paran'a floodplain, PR-MS, Brazil. Hydrobiologia 505 147-158. [ Links ]
DE EYTO E (2001) Chydorus sphaericus as a biological indicator of water quality in lakes. Verhandlungen Internationale Vereinigung für Theoretische und Angewente Limnologie 27 3358-3362. [ Links ]
DEJEN E, VIJVERBERG J, NAGELKERKE LAJ and SIBBING FA (2004) Temporal and spatial distribution of microcrustacean zooplankton in relation to turbidity and other environmental factors in a large tropical lake (L. Tana, Ethiopia). Hydrobiologia 513 39-49. [ Links ]
DODSON SI (1991) Species richness of crustacean zooplankton in European lakes of different sizes. Verhandlungen Internationale Vereinigung für Theoretische und Angewente Limnologie 24 1223-1229. [ Links ]
DODSON SI (1992) Predicting crustacean zooplankton species richness. Limnol. Oceanogr. 37 848- 856. [ Links ]
DODSON SI, ARNOTT SE and COTTINGHAM KL (2000) The relationship in lake communities between primary productivity and species richness. Ecol. 81 2662-2679. [ Links ]
DOI H, CHANG KH and NAKANO S (2010) Dispersal, connectivity, and local conditions determine zooplankton community composition in artificially connected ponds. Aquat. Biol. 10 47-55. [ Links ]
DUSSART B (1969) Les copépodes des eaux continentales d'Europe occidentale. Tome II : Cyclopoides et biologie quantitative. N. Boubée et Cie, Paris. 292 pp. [ Links ]
EDMONDSON WT (1991) The Uses of Ecology: Lake Washington and Beyond. University of Washington Press. [ Links ]
FATHALLI A (2004) Etude de la qualité des eaux de la retenue du barrage Kasseb: caractérisation physico-chimique, phytoplanctonique et évaluation de la taxinomie. Institut Nationale de Tunisie. 119 pp. [ Links ]
FERNÁNDEZ-ROSADO MJ and LUCENA J (2001) Space- time heterogeneities of the zooplankton distribution in La Concepción reservoir (Istán, Málaga; Spain). Hydrobiologia 455 157-170. [ Links ]
FERRÁO-FILHO AS, SOARES M CS, MAGALHáES VF and AZEVEDO SMFO (2009) Biomonitoring of cyanotoxins in two tropical reservoirs by cladoceran toxicity bioassays. Ecotoxicol. Environ. Saf. 72 479-489. [ Links ]
FOLT CL and BURNS CW (1999) Biological drivers of zooplankton patchiness. Trends Ecol. Evol. 14 300-305. [ Links ]
FRIEDRICH G and POHLMANN M (2009) Long-term plankton studies at the lower Rhine/Germany. Limnologica 39 14-39. [ Links ]
FRONTIER S (1973) Etude statistique de la dispersion du zooplancton. J. Exp. Mar. Biol. Ecol. 12 229-262. [ Links ]
GEORGE DG and EDWARDS RW (1976) The effect of wind on the distribution of chlorophyll a and crustacean plankton in a shallow eutrophic reservoir. J. Appl. Ecol. 13 667-691. [ Links ]
GEORGE DG and WINFIELD IJ (2000) Factors influencing the spatial distribution of zooplankton and fish in Loch Ness, UK. Freshwater Biol. 43 557-570. [ Links ]
GILLOOLY JF and DODSON SI (2000) Latitudinal patterns in the size distribution and seasonal dynamics of new world, freshwater cladocerans. Limnol. Oceanogr. 45 22-30. [ Links ]
GONZALEZ A and CHANETON EJ (2002) Heterotrophic species extinction, abundance and biomass dynamics in an experimentally fragmented microecosystem. J. Anim. Ecol. 71 594-602. [ Links ]
GUEVARA G, LOZANO P, REINOSO G and VILLA F (2009) Horizontal and seasonal patterns of tropical zooplankton from the eutrophic Prado Reservoir (Colombia). Limnologica 39 128-139. [ Links ]
HEBERT PDN and HANN BJ (1986) Patterns in the composition of arctic tundra pond microcrustacean communities. Can. J. Fish. Aquat. Sci. 43 1416-425. [ Links ]
HERBLAND A, Le BOUTEILLER A and RAIMBAULT P (1985) Size structure of phytoplankton biomass in the equatorial Atlantic Ocean. Deep Sea Res. 32 819-836. [ Links ]
HOLYOAK M, LEIBOLD MA and HOLT RD (2005) Metacommunities: Spatial Dynamics and Ecological Communities. Chicago University Press, Chicago. [ Links ]
HORN H and HORN W (2008) Bottom-up or top-down - How is the autotrophic picoplankton mainly controlled? Results of long-term investigations from two drinking water reservoirs of different trophic state. Limnologica 38 302-312. [ Links ]
HORN W and HORN H (1990) Long-term relationship between phyto and zooplankton in the meso-eutrophique reservoir Saidenbach. Arch. hydrobiol. Beih. Ergebn. Limnol. 33 749-762. [ Links ]
ISARI S, PSARRA S, PITTA P, MARA P, TOMPROU MO, RAMFOS A, SOMARAKIS S, TSELEPIDES A, KOUTSIKOPOULOS C and FRAGOPOULU N (2007) Differential patterns of mesozooplankters' distribution in relation to physical and biological variables of the northeastern Aegean Sea (eastern Mediterranean). Mar. Biol. 151 1035-1050. [ Links ]
JARAMILLO JC and GAVIRIA S (2003) Caracterización física, química y estructura de la comunidad zooplanctónica de un pequeńo lago tropical, lago Santander (Rionegro, Antioquia, Colombia). Caldasia 25 355-380. [ Links ]
JEPPESEN E, JENSEN JP, SøNDERGAARD M, LAURIDSEN T and LANDKILDEHUS F (2000) Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshwater. Biol. 45 201-218. [ Links ]
KARJALAINEN J, HOLOPAINEN AL and HUTTUNEN P (1996a) Spatial patterns and relationships between phytoplankton, zooplankton and water quality in the Saimaa lake system, Finland. Hydrobiologia 322 267-276. [ Links ]
KELLER W and CONLON M (1994) Crustacean zooplankton communities and lake morphometry in Precambrian shield lakes. Can. J. Fish. Aquat. Sci. 51 2424-2434. [ Links ]
KOROVCHINSKY NM (1992) Sididae and Holopediidae (Crustacea: Daphniiformes). Guides to the Identification of the Micro invertebrates of the Continental Waters of the World 3. SPB Academic Publishing, The Hague. 85 pp. [ Links ]
LAIR N and AYADI H (1989) The seasonal succession of planktonic events in lake Ayadat, France: a comparison with the PEG model. Arch. für Hydrobiol. 115 (4) 589-602. [ Links ]
LAMPERT W and SOMMER U (1997) Limnoecology. The Ecology of Lakes and Streams. Oxford University Press, New York. 382 pp. [ Links ]
LANSAC-TÔHA FA, BONECKER CC, VELHO LF M and LIMA AF (1997) Composição, distribuição e abundância da comunidade zooplanctônica. In: Vazzoler AEA de M, Agostinho AA, Hahn NS (eds.) A Planície de Inundação do Alto rio Paraná: Aspectos Físicos, Biológicos e Socioeconômicos. EDUEM: Nupélia, Maringá 117-156. [ Links ]
LARSON GL, HOFFMAN R, MCINTIRE CD, LIENKAEMPER G and SAMORA B (2009) Zooplankton assemblages in montane lakes and ponds of Mount Rainier National Park, Washington State, USA. J. Plankton Res. 31 (3) 273-285. [ Links ]
LEIBOLD MA (1999) Biodiversity and nutrient enrichment in pond plankton communities. Evol. Ecol. Res. 1 73-95. [ Links ]
LEIBOLD MA, HOLYOAK M, MOUqUET N, AMARASEKARE P, CHASE JM, HOOPES MF, HOLT RD, SHURIN JB, LAW R, TILMAN D, LOREAU M and GONZALEZ A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol. Lett. 7 601-13. [ Links ]
LEONARD J and PEARL HW (2005) Zooplankton community structure, microzooplankton grazing impact, and seston energy content in the St Johns River, Florida as influenced by the toxic cyanobacterium Cylindrospermopsis raciborskii. Hydrobiologia 537 89-97. [ Links ]
LOCKE A (1992) Factors influencing community structure along stress gradients: zooplankton responses to acidification. Ecol. 73 903-909. [ Links ] MARGARITORA FG (1985) Cladocera. Fauna d'Italia, Bologna. 399 pp. [ Links ]
MITRA A (2009) Are closure terms appropriate or necessary descriptors of zooplankton loss in nutrient-phytoplankton-zooplankton type models? Ecol. Model. 220 611-620. [ Links ]
MORGADO F, QUINTANEIRO C, RODRIGUES E, PASTORINHO MR, BACELAR-NICOLAU P, VIEIRA L and AZEITEIRO UM (2007) Composition of the Trophic Structure of Zooplankton in a Shallow Temperate Estuary (Mondego Estuary, Western Portugal). Zool. Stud. 46 1 57-68. [ Links ]
MOUQUET N and LOREAU M (2003) Community patterns in sourcesink metacommunities. Am. Nat. 162 544-557. [ Links ]
MURPHY J and RILEY JP (1962) A modified single solution method for the determination of phosphate in natural waters. Annal. Chim. Acta. 27 31-36. [ Links ]
MURUGAVEL P and PANDIAN T J (2000) Effect of altitude on hydrology, productivity and species richness in Kodayar - a tropical peninsular Indian aquatic system. Hydrobiologia 430 33-57. [ Links ]
PARANAGUÁ MN, NEUMANN-LEITÃO S, NOGUEIRA-PARANHOS JD, SILVA TA and MATSUMURA-TUNDISI T (2005) Cladocerans (Branchiopoda) of a tropical estuary in Brazil. Braz. J. Biol. 65 107-115. [ Links ]
PINTO-COELHO R, PINEL-ALLOUL B, MÉTHOT G and HAVENS KE (2005) Crustacean zooplankton in lakes and reservoirs of temperate and tropical regions: variation with trophic status. Can. J. Fish. Aquat. Sci. 62 348-361. [ Links ]
PONT D, CRIVELLI A J and GUILLOT F (1991) The impact of 3-spined sticklebacks on the zooplankton of a previously fish-free pool. Freshwater Biol. 26 149-163. [ Links ]
PRIMO AL, AZEITEIRO UM, MARqUES SC, MARTINHO F and PARDAL MA (2009) Changes in zooplankton diversity and distribution pattern under varying precipitation. Estuar. Coast. Shelf Sci. doi:10.1016/j.ecss.01.019. [ Links ]
RICKLEFS RE (1987) Community diversity: relative roles of local and regional processes. Science 235 167-171. [ Links ]
RINKE K, BNER IH, PETZOLDT T, ROLINSKI S, KöNIG-RINKE M, POST J, LORKE A and BENNDORF JR (2006) How internal waves influence the vertical distribution of zooplankton. Freshwater Biol. 52 137-144. [ Links ]
RINKE K, HUBER AMR, KEMPKE S, EDER M, WOLF T, PROBST WN and ROTHHAUPT KO (2009) Lake-wide distributions of temperature, phytoplankton, zooplankton, and fish in the pelagic zone of a large lake. Limnol. Oceanogr. 54 (4) 1306-1322. [ Links ]
RODIER J (1984) Analyse de l'eau. Edition Dunod, Paris. 1135 pp. [ Links ]
SCHEFFER M (1998) Ecology of Shallow Lakes. Chapman and Hall, London. [ Links ]
SCHWOERBEL J (1986) Methoden der Hydrobiologie-Süβwasser-biologie Susswasserbiologie. Stuttgart: Gustav Fischer Verlag. 369 pp. [ Links ]
SELLAMI I, AYADI H, BOUAIN A, ALEYA L and ALAOUI MHAMDI M (2009) Distribution of zooplankton related to environmental factors in three interconnected reservoirs: Kasseb, Mornaguia and Ghdir El Goulla (North of Tunisia). Ann. Limnol. - Int. J. Lim. 45 107-117. [ Links ]
SHAMEEL M (2001) An approach to the classification of algae in the new millennium. Pakistan J. Mar. Biol. 7 (1-2) 233-250. [ Links ]
SHANNON CE and WEAVER G (1949) The Mathematical Theory of Communication. University of Illinois Press, Urbana, Chicago. [ Links ]
SHURIN JB (2000) Dispersal limitation, invasion resistance and the structure of pond zooplankton communities. Ecol. 81 3074-3086. [ Links ]
SOMMER U, GLIWICZ ZM, LAMPERT W and DUNCAN A (1986) The PEG-model of seasonal succession of planktonic events in fresh waters. Arch Hydrobiol. 106 433-471. [ Links ]
STELLA B, SEBASTIAN D, HERWIG S, GABRIELE T, MIRIAM R, ANGELIKA W, ACHIM W, CHRISTOPH J and MAREN S (2007) Water temperature and mixing depth affect timing and magnitude of events during spring succession of the plankton. Oecologia 150 (4) 643-654. [ Links ]
STELLA E (1982) Calanoidi (Crustacea, Copépoda, Cananoida), 14. Consiglio Nazionale Delle Ricerche Aq/1/140. Guide Per riconscimento delle specie animali delle acqua interne italiane. Impresa dalla stamperia valdonesa verona. 67 pp. [ Links ]
STEMBERGER RS, HERLIHY AT, KUGLER DL and PAULSEN SG (1996) Climatic forcing on zooplankton richness in lakes of the northeastern United States. Limnol. Oceanogr. 41 1093-1101. [ Links ]
URABE J (1990) Stable horizontal variation in the zooplankton community structure of a reservoir maintained by predation and competition. Limnol. Oceanogr. 35 1703-1717. [ Links ]
UTERMÖHL H (1958) Zur Vervollkommung der quantitativen phytoplankton methodik. Mitteilungen Internationale Vereinigung für Theoretische und Angewandte Limnology 9 1-38. [ Links ]
VANSCHOENWINKEL B, DE VRIES C, SEAMAN M and BRENDONCK L (2007) The role of metacommunity processes in shaping invertebrate rock pool communities along a dispersal gradient. Oikos 116 1255-1266. [ Links ]
VELHO LFM (2000) Estrutura e dinâmica de assembléia de tecamebas no plâncton da planície de inundação do Alto Rio Paraná. Tese de Doutorado, Universidade Estadual de Maringá, Paraná [ Links ].
WELSCHMEYER NA (1994) Fluorimetric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Ocean. 39 1985-1992. [ Links ]
YAN ND, KELLER W, SOMERS KM PAWSON TW and GIRARD RE (1996) Recovery of crustacean zooplankton communities from acid and metal contamination: comparing manipulated and reference lakes. Can. J. Fish. Aquat. Sci. 53 1301-1327. [ Links ]
ZAR JH (1999) Biostatistical Analysis. Prentice-Hall International, New Jersey. [ Links ]
ZHENBIN WU, AIFEN L, SHIYANG Z, SHUIPING C, XIAOHUI WU (2008) Short-term effects of drawing water for connectivity of rivers and lakes on zooplankton community structure. J. Environ. Sci 20 419-423. [ Links ]
Received February 2010; accepted in revised form 8 March 2011.
* To whomall correspondence should be addressed. +216 (97) 64 21 80; fax: +216 (74) 27 44 37; e-mail: sellamifss@yahoo.fr


{kind=link}
{kind=link}
{kind=link}