










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.36 n.4 Pretoria Jul. 2010
The use of liver histopathology, lipid peroxidation and acetylcholinesterase assays as biomarkers of contaminant-induced stress in the Cape stumpnose, Rhabdosargus holubi (Teleostei: Sparidae), from selected South African estuaries
N RichardsonI,*; AK GordonII; WJ MullerII; BI PletschkeIII; AK WhitfieldIV
IDepartment of Ichthyology and Fisheries Science, Rhodes University, PO Box 94, Grahamstown 6140, South Africa
IIUnilever Centre for Environmental Water Quality, Institute for Water Research, Rhodes University, PO Box 94, Grahamstown 6140, South Africa
IIIDepartment of Biochemistry, Microbiology and Biotechnology, Rhodes University, PO Box 94, Grahamstown 6140, South Africa
IVSouth African Institute for Aquatic Biodiversity, Private Bag 1015, Grahamstown 6140, South Africa
ABSTRACT
Three biomarkers of contaminant-induced stress (liver histopathology, the lipid peroxidation (LPx) assay and the acetylcholinesterase (AChE) assay) were adapted for application to the estuarine-dependent marine fish Rhabdosargus holubi (Steindachner, 1881). Specimens of R. holubi were collected using a seine net from 3 temporarily open/closed estuaries in the Eastern Cape, South Africa, each impacted by different anthropogenic activities. The East Kleinemonde estuary has a housing settlement on the banks in the lower reaches and some agriculture in its catchment. The Old Woman's estuary has a golf course adjacent to its lower and middle reaches and is crossed by a national road in its upper reaches. The Mtana estuary is virtually pristine, with limited cattle grazing occurring along the banks of the estuary and some subsistence agriculture in the catchment. According to the biomarker results from this study, R. holubi from the East Kleinemonde were in good health, as reflected by low LPx and high AChE levels. The liver histopathology did, however, suggest possible previous exposure to stress (increased melanomacrophage centres, increased perivascular connective tissue and severe vacuolation). Overall, liver histopathology results did not differ significantly between estuaries. Fish from the Old Womans recorded significantly inhibited AChE and increased LPx, while fish from the Mtana exhibited significantly increased LPx only, suggesting possible exposure to anticholinesterase contaminants in the former estuary and some form of oxidative stress in the latter. Although water samples were collected from each of the 3 estuaries and analysed for polychlorinated biphenyls, organochlorines, organophosphorous pesticides and pyrethroids, none of these chemicals were detected. As pesticide residues in water are highly variable, both temporally and spatially, future studies should focus on measuring tissue burdens of organisms in order to identify the contaminant stressor. This study has shown that while chemical analyses of water provide a 'snap-shot' of water quality at the time of sampling, biomonitoring can integrate past exposures to stress and is thus useful for identifying potential situations of concern that require further detailed investigation.
Keywords: biomonitoring, pollution, estuaries, AChE, LPx, histopathology
Introduction
Biomarkers can be sensitive indicators of sub-cellular stress in organisms exposed over short and longer periods to a range of pollutants (Adams et al., 2000). Several international studies have shown that biomarkers are useful tools in the monitoring of estuarine biotic 'health' and therefore give an indirect measure of possible environmental pollution or degradation (Cavanagh et al., 2000; Kirby et al., 2000; Fulton and Key, 2001; Lund Amado et al., 2006; Valavanidis et al., 2006; Sole et al., 2006). South African monitoring programmes in estuaries have focused on the use of chemical analyses of water and sediments to determine contaminant loadings (Schulz, 2001; Scharler and Baird, 2003; Bollmohr et al., 2007), and on fish community composition and structure to determine estuarine health (Harrison and Whitfield, 2006).
Chemical analyses of water and sediment cannot provide direct indications of the effect of contaminants on the biota of an estuary. This is because contaminant concentrations are not constant over time, toxicity is caused by the bioavailable fraction of the toxicant only (not the total measured concentration), and processes such as bioaccumulation can lead to an underestimation of toxic risk (Adams and Tremblay, 2003). Community indices have been criticised for only indicating severe stress that has already occurred (a change in the community structure), and do not provide any indication of the type of stressor that caused the change or early warnings of potential effects (Adams et al., 2005). Biomarkers have therefore been proposed as a method of detecting contaminant induced suborganism level stress in the biota of an estuary before population- or community-level responses become apparent (Adams et al., 2005).
The aim of this pilot study was to determine the feasibility of using 3 biomarkers, at different sub-organismal levels, to detect pollutant-induced stress in the Cape stumpnose, Rhabdosargus holubi, in 3 temporary open/closed estuaries (TOCEs), each impacted by different anthropogenic activities (Richardson, 2008). TOCEs were selected for this biomarker study as, when closed, fish are not able to migrate in or out of the estuary, thereby ensuring that any measured effects in fish can be attributed to the circumstances within that particular estuary. In addition, the selected estuaries are in close proximity to each other, in the same biogeographical zone (warm temperate), and are similar in terms of hydrology and mouth morphology.
Rhabdosargus holubi was identified as a potential bioindicator species for South African estuaries due to its abundance and wide distribution on both the south and east coasts. This species is dependent on estuaries as nursery areas during its juvenile phase (therefore only juvenile individuals can be used as bioindicators of estuarine health), with the subadults and adults restricted to the marine environment (Blaber, 1974a). The Cape stumpnose uses estuaries in a similar manner to a number of other important angling species in South African waters and therefore the management of estuaries using R. holubi as an indicator species could have positive outcomes for other estuary-associated marine taxa. Aspects of the biology and ecology of this easily identified species have been well studied (Blaber, 1973a; 1973b; 1974a; 1974b; 1974c; Whitfield, 1984; Cowley et al., 2001) and it has been successfully used in other pollution-related experiments (De Kock and Lord, 1988).
Knowledge of contaminant concentrations in most South African estuaries is limited. Orr et al. (2008) have reported the presence of heavy metals in the Kariega estuary sediments and to a lesser extent in the East Kleinemonde and Riet estuary sediments. Although no analyses of pesticide contamination have occurred in the Eastern Cape estuaries investigated during this study, Schulz (2001), Bollmohr et al. (2007) and Bollmohr et al. (2009) have shown that pesticides are periodically washed into estuaries along the Western Cape coast. Since one of the estuaries in this study is situated adjacent to a golf course that uses pesticides, the acetylcholinesterase activity (AChE) assay was chosen as a biomarker. Dipping of cattle belonging to rural subsistence farmers is undertaken by state veterinarians on a frequent basis in the Eastern Cape and these cattle often wade through estuaries. The potential impact of this on the ichthyofauna of estuaries was investigated using 2 broad biomarkers of exposure to contaminants, the lipid peroxidation (LPx) assay and liver histopathology.
Lipid peroxidation occurs as result of oxidative stress damage to the lipid components of cellular membranes (Kelly et al., 1998). This reaction occurs, to a limited degree, during natural immunity processes (e.g. phagocytosis) but is caused to a much greater extent by xenobiotics. The LPx assay measures malondialdehyde, a by-product of LPx, and can be used as a quantitative measurement of the extent of oxidative damage an organ has undergone. The liver was chosen for the LPx analyses in order to allow for comparison with the results from the histopathological investigations.
Histopathological analyses and LPx assay of the liver can provide a good indication of the overall health of an animal, including fish. The liver is a key organ of overall homeostasis, in terms of nutrition, defence against toxicants and reproductive development (Hinton and Lauren, 1990). Examples of redox cycling compounds that have been shown to affect LPx levels include quinones, certain dyes, bipyridyl herbicides, metals, aromatic nitro compounds and certain pesticides (Kelly et al., 1998; Dorval et al., 2005; Valavanidis et al., 2006).
Histological alterations in the liver have been shown to provide definite biological end points of exposure to a number of contaminants and have been successfully applied in several biomonitoring programs in Europe and North America (Malins et al., 1988; Kohler, 1989; Hinton and Lauren, 1990; Stentiford et al., 2003). Laboratory and field-based studies have established relationships between contaminant exposure and toxicopathic lesion formation (Naigaga, 2002; Stentiford et al., 2003; Van Dyk, 2003).
The AChE enzyme is found in the brain and muscle tissue of fish and controls a large proportion of physiological and behavioural responses in the animal; thus any changes to these regulatory abilities could be potentially detrimental to fish (Pan and Dutta, 1998; Kirby et al., 2000). This assay is regarded as a good biomarker and has been successfully used in fish to detect exposure to a range of neurotoxic compounds (Kirby et al., 2000; Roex et al., 2003; Eder et al., 2004; Lau et al., 2004). Until recently, the AChE assay was considered to be a specific biomarker of exposure to pesticides and insecticides; however recent studies have shown that AChE activity is also inhibited by metals, organochlorines, herbicides and surfactants (Guilhermino et al., 1998; Guilhermino et al., 2000; Corsi et al., 2003; Roy et al., 2006; Bervoets et al., 2009, Mochida et al., 2009; Modesto and Martinez, 2010).
Study sites
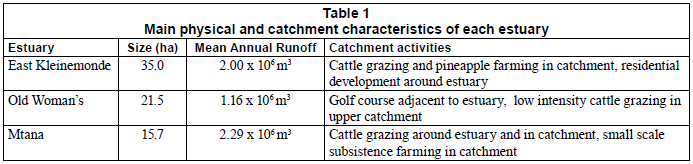
The 3 TOCEs chosen for this study, the East Kleinemonde (EK), Old Woman's (OW) and Mtana (MTN), are of similar morphology and are located within the same biogeographical zone in the Eastern Cape Province (Fig. 1). The estuaries differ according to the types of anthropogenic impacts occurring within their catchments (Table 1). The EK is characterised by housing developments along the banks of the lower estuary reaches and pineapple and cattle farms in the catchment area. The OW catchment has a golf course along the banks of the lower and middle reaches and is traversed by a national road in the upper reaches. Some low intensity cattle grazing occurs in the catchment of this estuary. The MTN catchment is virtually undeveloped although limited subsistence farming is recorded in certain areas. Low intensity cattle grazing occurs throughout the catchment and along the banks of the estuary.
Materials and methods
Sampling methods
At the time of sampling, all 3 estuaries were closed and had been for 4 months, thereby ensuring that the fish in a particular estuary had been exposed to the conditions in that particular estuary for at least that period. One hundred juvenile R. holubi of similar size (9.3 ± 1.6 cm standard length) were collected during the course of a morning between 18 and 25 April 2005 from the lower reaches of each estuary. Fish were caught using a 30 m x 1.7 m x 15 mm bar mesh seine net fitted with a 5 mm bar mesh cod end and then held in the estuary in a submerged, covered container that allowed free exchange of water. All fish were processed within 3 h of capture. For each individual, the spinal cord was severed before the brain and liver were excised. Dissecting tools were rinsed in 70% ethanol, washed in deionised water and dried with paper towel between each dissection. The brain and half of the liver were immediately frozen in liquid nitrogen and stored at -70 ºC until further processing for AChE and LPx assays could be conducted. The remaining half of the liver was placed in 10% buffered formalin for 72 h and then transferred to 70% alcohol for later histopathological analysis (Richardson, 2008).
Histopathological analyses
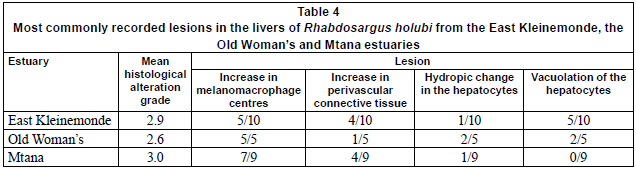
Liver tissues obtained from 24 individuals (10 from the EK, 5 from the OW and 9 from the MTN) were embedded in paraffin wax, sectioned into 5 µm slices and stained using haematoxylin and eosin. Each slide was graded between 0 and 4 (Table 2) according to methods adapted from Van Dyk et al. (2007). Grading involved examining the liver slides under the microscope in order to identify a range of lesions such as melanomacrophage centres, increases in perivascular tissue, hydropic change and vacuolation in the hepatocytes. Each type of change recorded was assigned a specific score (Table 2).
LPx determination
Liver samples were assayed according to the methods by Ringwood et al. (2003). Each section of liver was weighed, added to cold 50 mM potassium phosphate buffer solution (pH 7.0) based on 4 times the gram weight of the sample, homogenised using Teflon Eppendorf micropestles, and then centrifuged for 5 min at 13 000 g at 4ºC. Sample supernatant (50 µℓ) was then added to 700 µℓ of 10 mM 0.375% (w/v) thiobarbituric acid and 7 µℓ of 2% (w/v) butylated hydroxytoluene and incubated in a heating block at 100ºC for 15 min. The samples were centrifuged at 13 000 g for 5 min at 22 ± 2ºC. A 100 µℓ sample of supernatant was placed in the well of a 96-well microtiter plate and read at 532 nm using a microtiter plate reader (PowerWavex, Bio-Tek Instruments Inc, USA). A standard curve was prepared using serial dilutions of malondialdehyde tetraethylacetal (MDA) in a concentration range of 6.25 to 3 200 µM MDA. Each sample and standard was analysed in quadruplicate and the results were expressed as the median ± 95% confidence intervals (Richardson, 2008).
AChE activity determination
Methods for the determination of AChE activity in the brain tissue were optimised for R. holubi based on methods described by McLoughlin et al. (2000). In brief, each brain sample was homogenised on ice in 30 µℓ cold phosphate buffer saline (PBS) (pH 8.0) containing 1% (v/v) Triton X-100, using Teflon Eppendorf micropestles. The homogenate was diluted with 270 µℓ PBS and centrifuged for 20 min at 13 000 g at 4ºC. A 50 µℓ sample of this supernatant was diluted with 950 µℓ cold PBS containing 0.1% (v/v) Triton X-100 in preparation for the assay. Enzyme activity was determined by adding 100 µℓ of 8 mM 5,5'-dithio-bis (2-nitrobenzoic acid) and 50 µℓ of each sample's supernatant to a well in a 96-well microtiter plate kept on ice. The microtiter plate was incubated at 30˚C in a microtiter plate reader (PowerWavex, Bio-Tek Instruments Inc, USA) for 3 min. An aliquot of 50 µℓ of 16 mM acetylthiocholine iodide was added to each well, after which a reading was taken at 405 nm every 30 s for 10 min (McLoughlin et al., 2000). Enzyme activity was calculated using the following equation:

where an extinction coefficient (ε) value of 1.36 x 104 ℓ·mol-1·cm-1 was used, and where l is the pathlength in cm, Δ abs is the change in absorbance per minute and df is the dilution factor (Ellman et al., 1961).
Fresh standards were prepared daily using commercial AChE (1 mg·mℓ-1) at a concentration range between 0 mg·mℓ-1 and 0.125 mg·mℓ-1 to produce a standard curve. Specific standard concentrations were included in subsequent plates to ensure consistently accurate readings. Each sample and standard was analysed in quadruplicate (Richardson, 2008).
Protein content determination
Liver and brain sample protein content was determined according to the method described in Bradford (1976). For the liver samples, a dilution was made using 2 µℓ of the original liver homogenate (containing 4 times the gram weight of the liver in millilitres of 50 mM potassium phosphate buffer, pH 7.0) and 198 µℓ of 50 mM potassium phosphate buffer (pH 7.0). A portion of this diluted sample (20 µℓ) was placed in the well of a 96-well microtiter plate and 230 µℓ of Bradford reagent added. The plate was allowed to stand for 5 min at 22 ± 2ºC whereafter a reading was taken at 595 nm.
For the brain tissue, 20 µℓ of brain sample homogenate (containing 50 µℓ of the original homogenate diluted with 950 µℓ cold PBS containing 0.1% (v/v) Triton X-100) was placed in a 96-well microtiter plate well. Bradford reagent (230 µℓ) was added and the plate was allowed to stand for 5 min at 22 ± 2ºC, whereafter a reading was taken at 595 nm.
Standards were prepared using commercial Bovine Serum Albumin (BSA) and ranged between 0 and 0.4 mg·mℓ-1. This standard curve was used to calculate the protein content in each liver and brain sample. Final malondialdehyde concentrations were expressed as nmol MDA produced·h-1·mg protein-1. For the brain tissue, sample AChE activities (µmol·mℓ-1·min-1) were divided by the sample's protein content (mg·mℓ-1) and multiplied by 1 000 to present AChE activity in nmol·min-1·mg protein-1 (Richardson, 2008).
Water analyses
Subsurface (<1 m) water samples were collected from the lower, middle and upper reaches of each estuary during the course of 1 morning and were immediately sent, on ice, to the South African Bureau of Standards (SABS) for analysis. Single determinations were carried out by employing the method as described in the Official Methods of Analysis of AOAC International manual (AOAC, 1995). Quantitative analyses were conducted by gas chromatography-electron capture detector for organochlorine pesticides (OCs), pyrethroids and polychlorinated biphenyls (PCBs). Organophosphorous pesticide (OPs) concentrations were determined using gas chromatography - flame photometric detection methods. The presence of pesticide residues were confirmed using gas chromatography - mass spectrometry. This multi-residue analysis was undertaken to test for a range of pesticides and pyrethroids (listed in Table 3) as these are the most commonly detected contaminants in South African estuaries (Schulz, 2001; Bollmohr et al., 2007; Bollmohr et al., 2009). Under the conditions of the analyses employed by the SABS, the lowest limit of detection was 0.5 µg·ℓ-1.

Statistical analyses
Normality of the data from each biomarker analysis was tested using the Kolmogorov-Smirnov test (P < 0.05). All the data were non-parametric and therefore the Kruskal-Wallis test was performed, and multiple comparisons of the data sets between estuaries were undertaken using the Mann Whitney U-test. Comparisons were conducted between each pair of estuaries to determine for significant differences in biomarker responses in fish from the 3 estuaries. A Bonferroni adjusted level of significance of P<0.02 was used when determining these differences (Zar, 1999). All analyses were conducted using STATISTICATM Version 7.0.
Results
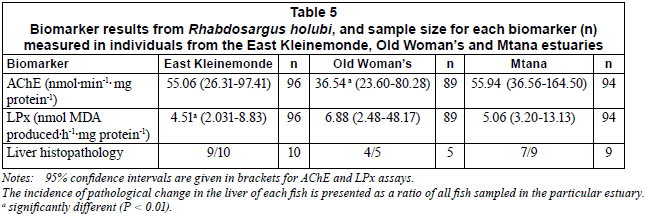
None of the data were normally distributed; consequently the results are presented as medians with 95% confidence intervals. The 3 estuaries did not differ significantly in terms of the results from the histological analyses performed (Kruskal-Wallis, H=0.6378, df=2, 24, P=0.727). The predominant lesions found in fish from the EK were increased melanomacrophage centres, increased perivascular connective tissue and severe vacuolation (Table 4). Nine out of ten fish from the EK showed pathological changes (Table 5). Four out of five fish from the OW (Table 5) showed pathological changes with the most common lesions being increased melanomacrophage centres, cellular swelling due to hydropic change and severe vacuolation (Table 4). Seven out of nine fish from the MTN showed pathological changes (Table 5), with the predominant lesions being increased melanomacrophage centres and perivascular connective tissue (Table 4). The mean scores for histological alterations as determined from the grading system were 2.9, 2.6 and 3.0 for the EK, OW and MTN estuaries, respectively (Table 4).
The LPx levels in liver samples from the 3 estuaries were found to be significantly different (Kruskal-Wallis, H=23.76, df=287, P<0.01). Levels in fish from the MTN and OW were significantly different (higher) to those in the EK (EK/MTN: Mann Whitney U test, z=-3.557, P<0.001. EK/OW: Mann Whitney U test, z=-4.329, P<0.001), while the OW and MTN were not significantly different from one another (Mann Whitney U test, z=2.206, P=0.03) (Table 5).
Significant differences in brain AChE activity were measured from the 3 estuaries (Kruskal-Wallis, H=44.09, df=293, P<0.001). While fish from the EK and MTN did not have significantly different AChE activities (Mann Whitney U test, z=-1.476, P=0.14), fish from the OW had significantly lower AChE activities than fish from both the EK (Mann Whitney U test, z=4.781, P<0.001) and the MTN (Mann Whitney U test, z=-6.409, P<0.001) (Table 5).
Water samples collected from the upper, middle and lower reaches in all 3 estuaries showed pesticide concentrations below the detection limit of the tests employed. Unfortunately, no historical water chemistry data exists for these 3 estuaries either.
Discussion
The estuaries investigated during this study were all temporarily open/closed systems. The last mouth-opening event was 4 months prior to sampling, consequently migration of R. holubi into or out of the estuaries during this time was prevented. Thus, the fish investigated during this study had all inhabited their respective estuaries for at least 4 months prior to sampling and were exposed to any pollution events during this period. Although toxicological analysis of water samples from the lower, middle and upper reaches of the study estuaries failed to reveal the presence of polychlorinated biphenyls, organochlorines, organophosphorous pesticides or pyrethroids (Richardson, 2008), this does not mean that these contaminants were absent from these systems in the prior 4 months. Since pesticide residues are highly variable, both temporally and spatially (Bollmohr et al., 2009), future studies should focus on analysing regular water samples from the study estuaries, as well as measuring tissue burdens of organisms over an extended period in order to identify contaminant stress.
Liver histopathology of fish from the 3 estuaries did not conclusively distinguish between the anthropogenic activities (and associated potential pollutant stressors) present within each catchment. An increase in melanomacrophage centres can be indicative of previous exposure to toxicants but may also be a result of parasitic infection (Hinton and Lauren, 1990; Dezfuli et al., 2007). A macroscopic inspection of each fish was conducted prior to removal of the liver and only 1 sample from the OW was found to have parasites in the mesenteries surrounding the stomach and this specimen was not used for analysis. Parasites can, however, also be present on the gills and in the muscle tissue of fish and these areas were not investigated due to time constraints in the field. A study conducted using flounder (Platichthys flesus) from 4 estuaries in England found that melanomacrophage centres were more prevalent in samples collected from contaminated estuaries compared to uncontaminated estuaries (Stentiford et al., 2003). Although the increase in melanomacrophage centres recorded in the present study could possibly suggest exposure to pollutants, further intensive contaminant analysis of the 3 estuaries is required to confirm this.
An increase in connective tissue in the parenchyma was found in fish from all 3 estuaries (Table 4) and, according to Hibiya (1982), this lesion is a result of phagocyte infiltration to a focal area of necrosis. Van Dyk (2003) conducted a histopathological study on Clarias gariepinus collected from 3 sites: 2 were considered polluted and the 3rd was a site that was considered to be in pristine condition. The researcher found an increase in connective tissue in the parenchyma in 60% of the fish caught in one of the polluted sites and 35% of the fish caught from the 2nd polluted site. None of the fish collected from the control site had an increase in perivascular tissue. The above results, coupled with findings from the present study, suggest that this type of lesion may be a potentially useful indicator of exposure to xenobiotics in R. holubi.
Hydropic change in a marine flatfish (Pleuronectes vetulus) and in 3 estuary-associated species (Platichthys flesus, Pomatoschistus minutus and Zoarces viviparous) was described in 2 northern hemisphere studies and in both cases was shown to be correlated with exposure to hydrocarbons (Stehr et al., 1998; Stentiford et al., 2003). In addition, reduced occurrences of hydropic change in the flounder, Pleuronectes americanus, occurred when improved sewage management practices in Massachusetts resulted in decreases in heavy metal and organic compounds within the sewage effluent entering receiving waters (Moore et al., 1996). A few R. holubi from each estuary in this study showed evidence of hydropic change in their hepatocytes. However, quantification of the natural level of occurrence of hydropic change in R. holubi is required in order to ascertain the extent of exposure to contaminants.
Vacuolation was only found in fish from the EK and OW (Table 4). Vacuolation has been shown to occur as a result of exposure to xenobiotics. For example, Wester et al. (1990) exposed Oryzias latipes to various concentrations of bis(trin-butyltin)oxide and di-n-butyltindichloride, and found a dose-dependent increase in glycogen-related vacuolation in the liver in response to exposure to both of these chemicals. In a similar study, Crestani et al. (2007) investigated the effects of short-term exposure to herbicide clomazone on liver histology and vacuole formation in silver catfish, Rhamdia quelen. These authors reported vacuolation of hepatocytes for various exposure durations and both the above studies suggest that vacuolation may be indicative of short-term toxicant exposure.
The fish from the OW and MTN had significantly higher LPx levels compared to the EK (Table 5), suggesting that these fish experienced oxidative stress. Dorval et al. (2005) investigated LPx levels using the white sucker (Catostomus commersoni) and found that the fish from 3 polluted sites showed significantly higher LPx levels in the liver tissue than those taken from the control site. These authors also found that plasma AChE levels were depressed when compared to a control site, probably as a consequence of pesticides in the water at the polluted sites. Similar results were reported by Sole et al. (2006), who found significant increases in LPx in Pomatoschistus minutus following the release of pesticide-containing water from an upstream dam. Pomatoschistus minutus was sampled throughout the year, and increased LPx levels were only reported during periods of high rainfall or when water was released from the upstream dam, coinciding with a significant decrease in AChE activity (Sole et al., 2006). Interestingly, Sole et al. (2006) noted that LPx levels were relatively uniform (low standard errors) during low levels of LPx activity, a pattern that was mirrored in the samples from the EK, with high standard errors during periods of high LPx levels; this was also the pattern noted in the data from the OW (Table 5) in this study.
The brain AChE levels in fish from the OW were 35% lower than the values obtained for fish from the EK and MTN (Table 4). Varo et al. (2003) consider inhibitions greater than 20% to be indicative of exposure to acetylcholinesterase inhibiting chemicals. The per cent inhibitions recorded during the present study are less than those found in related studies which used similar techniques (Coppage and Braidech, 1976; Minier et al., 2000; Kirby et al., 2000; De la Torre et al., 2002; Sole et al., 2006). However, all of those studies were undertaken in heavily industrialised rivers, possibly explaining the high inhibition levels recorded. In contrast, the OW is not heavily influenced by industrial land-use, but could possibly be impacted by pesticides during spray-drift and rainfall runoff from the nearby golf course.
When considered in combination, the biomarker results from the EK suggest that the R. holubi in this estuary were in good health at the time of sampling. The overall score for the EK in terms of the histology was less than 3, despite a high (9 out of the 10 samples) occurrence of pathological changes to the liver. The AChE levels were high and the LPx levels were low, suggesting that the brain and liver tissues were not affected by anticholinesterase compounds or oxidative stressors respectively. The predominant lesions that were recorded using the histopathology were an increase in melanomacrophage centres and an increase in perivascular tissue. Both of these pathologies suggest previous exposure to stressors as they are indicative of cellular repairing mechanisms. The stressors may have been of anthropogenic (e.g. chemical) or natural origin (e.g. parasites). The low LPx and absence of other histopathological signs of stress, such as a degeneration of the parenchyma, suggests that the liver was no longer undergoing stress at the time of sampling (Richardson, 2008).
The biomarker results suggest fish from the OW were stressed. Four of the five histological samples of fish from the OW showed signs of pathological changes to the liver. In addition, there was large AChE inhibition and high LPx levels in the fish sampled from this estuary. Although the golf course appears to be the principal potential pollution source (since there are no other anthropogenic activities in the catchment), analyses of the water samples did not detect the presence of pesticides. Future studies should investigate both the water quality of precipitation run-off from the golf course into the estuary and monitor the accumulation of contaminants in the body tissues of fish within the system.
The fish from the MTN estuary exhibited increased LPx levels, but AChE activities remained high. In addition, 2 fish from this estuary scored a grade of 4 for the histological analyses. The grade of 4 indicates a degeneration of the liver parenchyma and is indicative of fish that have been strongly affected by hepatic disease. This, combined with the LPx data, suggested that some fish from the MTN were exposed to xenobiotics. The causes of these biomarker responses are unknown since there are limited land-use activities (primarily small-scale subsistence crop farming and cattle grazing) within this catchment. The fact that the AChE levels were unaffected implies that the pollutant is probably not an anticholinesterase agent. However, the only possible source of contaminants to the estuary was from biocide-dipped cattle belonging to local inhabitants. These cattle were regularly observed wading through the estuary as well as feeding on vegetation adjacent to the estuary. Cattle are routinely immersed in a biocide solution to prevent parasites (especially ticks) by the state veterinary services. Dipping occurs every 2 to 3 weeks in winter and every 1 to 2 weeks in summer. Plunge dips are used and the dip solution is Amitraz (chemical name N,N'-[(methylimino) dimethyllidyne] di-2,4-xylidine) (Pretorius, YEAR). Further water chemistry and sediment sampling of the MTN estuary, as well as toxicological examinations of the particular pesticide used on the cattle in the area, would be required in order to determine the causes of the biomarker responses measured.
The biomarker methods used during this study were successfully implemented to provide a preliminary assessment of whether anthropogenic activities in the 3 systems were having an effect on the health of resident R. holubi or not. The histology methodology applied to R. holubi in this study provided a quantitative assessment of the lesions observed, although further investigation is required to fully understand the relationship between the histological responses in R. holubi and chemical exposure. The AChE and LPx assays were relatively straightforward to perform and many (~ 35 samples/day) samples could be analysed simultaneously. Other assays related to the liver tissue such as the glutathione assay, the superoxide dimutases assay or measuring metallothionein levels could further assist in determining more precisely which group of chemicals were responsible for the lesions seen in the fish livers, particularly in the EK.
The fact that the chemicals tested for in water from each of the estuaries were below the test detection limits may be due to the time of sampling, as there had been no recent rainfall event to wash any land-based chemicals into the estuaries. Other factors to consider are the short half-life of some pesticides and the influence of salinity on the bioavailabilty of pesticides. Furthermore, the chemicals may have come out of solution and bound to the sediments (De la Torre et al., 2002; Bollmohr et al., 2009), or were present in concentrations below the detection limit of the analyses. These possibilities highlight the shortcomings of using water sample chemical analyses to detect pollution, as the results provide limited insight into the longerterm effects of harmful chemicals on the biota of an ecosystem. Chemical analyses provide a 'snap shot' of the water quality at the time of sampling (Bollmohr et al., 2007) and do not indicate medium or long-term ecosystem health. Biomonitoring has the advantage of integrating past pollution events and can be used to determine the effect of longer-term pollution problems on the aquatic biota (McCarthy and Shugart, 1990). However, most biomarkers lack ecological relevance as they cannot be linked to effects at population or community levels, and consequently should not replace community-level indicators, but rather complement such investigations.
Acknowledgements
Funding from the South African Institute for Aquatic Biodiversity, the Unilever Centre for Environmental Water Quality at Rhodes University, the National Research Foundation and the Water Research Commission is gratefully acknowledged. The staff and students of the Department of Ichthyology and Fisheries Science, Dr JC Van Dyk and Jenny Grewar are gratefully acknowledged for their help during this study.
References
ADAMS SM, GREELEY MS and RYON MG (2000) Evaluating effects of contaminants on fish health at multiple levels of biological organisation: extrapolating from lower to higher levels. Hum. Ecol. Risk Assess. 6 (1) 15-27. [ Links ]
ADAMS SM and TREMBLAY LA (2003) Integration of chemical and biological tools in environmental management and regulation. Australas. J. Ecotoxicol. 9 157-164. [ Links ]
ADAMS SM, RYON MG and SMITH JG (2005) Recovery in diversity of fish and invertebrate communities following remediation of a polluted stream: investigating causal relationships. Hydrobiol. 542 77-93. [ Links ]
AOAC International (1995) Official Methods of Analysis of AOAC International (16th edn.) Cunniff PA (ed.) Vol. 1, Chapter 10. AOAC International, Gaithersburg. [ Links ]
BERVOETS L, VAN CAMPENHOUT K, REYNDERS H, KNAPEN D, COVACI A and BLUST R (2009) Bioaccumulation of micropollutants and biomarker responses in caged carp (Cyprinus carpio). Ecotoxicol. Environ. Saf. 72 720-728 [ Links ]
BLABER SJM (1973a) Temperature and salinity tolerance of juvenile Rhabdosargus holubi (Steindachner) (Teleostei: Sparidae). J. Fish Biol. 5 593-598. [ Links ]
BLABER SJM (1973b) Population size and mortality of the marine teleost Rhabdosargus holubi (Pisces: Sparidae) in a closed estuary. Mar. Biol. 21 219-225. [ Links ]
BLABER SJM (1974a) The population structure and growth of juvenile Rhabdosargus holubi (Steindachner) (Teleostei: Sparidae) in a closed estuary. J. Fish Biol. 6 (4) 455-460. [ Links ]
BLABER SJM (1974b) Field studies of the diet of Rhabdosargus holubi (Pisces: Teleostei: Sparidae). J. Zool. Lond. 173 407-417. [ Links ]
BLABER SJM (1974c) Osmoregulation in juvenile Rhabdosargus holubi (Steindachner) (Teleostei: Sparidae). J. Fish Biol. 6 797-800. [ Links ]
BOLLMOHR S, DAY JA and SCHULZ R (2007) Temporal variability in particle-associated pesticide exposure in a temporarily open estuary, Western Cape, South Africa. Chemosphere 68 (3) 479-488. [ Links ]
BOLLMOHR S, VAN DEN BRINK PJ, WADE PW, DAY JA and SCHULZ R (2009) Spatial and temporal variability in particlebound pesticide exposure and their effects on benthic community structure in a temporarily open estuary. Estuar. Coast. Shelf Sci. 82 50-60. [ Links ]
BRADFORD MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72 (1-2) 248-254. [ Links ]
CAVANAGH JE, BURNS KA, BURNSKILL GJ, RYAN DAJ and AHOKAS JT (2000) Induction of hepatic cytochrome P-450 1A in Pikey Bream (Acanthopagrus berda) collected from agricultural and urban catchments in far North Queensland. Mar. Pollut. Bull. 41 (7-12) 377-384. [ Links ]
COPPAGE DL and BRAIDECH TE (1976) River pollution by anticholinesterase agents. Water Res. 10 (1) 19-24. [ Links ]
CORSI I, MARIOTTINI M, SENSINI C, LANCINI L and FOCARDI S (2003) Cytochrome P450, acetylcholinesterase and gonadal histology for evaluating contaminant exposure levels in fishes from a highly eutrophic brackish ecosystem: the Orbetello Lagoon, Italy. Mar. Pollut. Bull. 46 203-212. [ Links ]
COWLEY PD, WHITFIELD AK and BELL KNI (2001) The surf zone ichthyoplankton adjacent to an intermittently open estuary, with evidence of recruitment during marine overwash events. Estuar. Coast. Shelf Sci. 52 339-348. [ Links ]
CRESTANI M, MENEZES C, GLUSCZAK L, SANTOS MIRON DD, SPANEVELLO R, SILVEIRAA, GONCALVES FF, ZANELLA R and LORO VL (2007) Effect of clomazone herbicide on biochemical and histological aspects of silver catfish (Rhamdia quelen) and recovery pattern. Chemosphere 67 (11) 2305-2311. [ Links ]
DE KOCK AC and LORD DA (1988) Kinetics of the uptake and elimination of polychlorinated biphenyls by an estuarine fish species (Rhabdosargus holubi) after aqueous exposure. Chemosphere 17 (12) 2381-2390. [ Links ]
DE LA TORRE FR, FERRARI L and SALIBIAN A (2002) Freshwater pollution biomarker: response of brain acetylcholinesterase activity in two fish species. Comp. Biochem. Physiol. 131 (Part C) 271-280. [ Links ]
DEZFULI BS, PIRONI F, SHINN AP, MANERA M and GIARI L (2007) Histopathology and ultrastructure of Platichthys flesus naturally infected with Anisakis simplex S.L. larvae (Nematoda; Anisakidae). J. Parasitol. 93 (6) 1416-1423. [ Links ]
DORVAL J, LEBLOND V, DEBLOIS C and HONTELA A (2005) Oxidative stress and endocrine endpoints in white sucker (Catostomus commersoni) from a river impacted by agricultural chemicals. Environ. Toxicol. Chem. 24 (5) 1273-1280. [ Links ]
EDER KJ, LEUTENEGGER CM, WILSON BW and WERNER I (2004) Molecular and cellular biomarker responses to pesticide exposure in juvenile chinook salmon (Oncorhynchus tshawytscha). Mar. Environ. Res. 58 809-913. [ Links ]
ELLMAN GL, COURTNEY D, ANDRES V and FEATHERSTONE R (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 7 88-95. [ Links ]
FULTON MH and KEY PB (2001) Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorous insecticide exposure and effects. Environ. Toxicol. Chem. 20 (1) 37-45. [ Links ]
GUILHERMINO L, BARROS P, SILVA MC and SOARES AMVM (1998) Should the use of inhibition of cholinesterases as a specific biomarker of organophosphate and carbamate pesticides be questioned? Biomarkers 3 (2) 157-163. [ Links ]
GUILHERMINO L, LACERDA MN, NOGUEIRAAJA and SOARES AMVM (2000) In vitro and in vivo inhibition of Daphnia magna acetylcholinesterase by surfactant agents: possible implications for contamination biomonitoring. Sci. Total Environ. 247 137-141. [ Links ]
HARRISON TD and WHITFIELD AK (2006) Application of a multimetric fish index to assess the environmental condition of South African estuaries. Estuaries Coasts 29 (6B) 1108-1120. [ Links ]
HIBIYA T (1982) An Atlas of Fish Histology: Normal and Pathological Features. Kodansha Ltd. Tokyo. 90 pp. [ Links ]
HINTON DE and LAUREN DJ (1990) Liver structural alterations accompanying chronic toxicity in fishes: potential biomarkers of exposure. In: McCarthy JF and Shugart LR (eds.) Biomarkers of Environmental Contamination. Lewis Publishers, Florida. 17-57. [ Links ]
KELLY SA, HAVRILLA CM, BRADY TC, HARRIS ABRAMO K and LEVIN ED (1998) Oxidative stress in toxicology: Established mammalian and emerging piscine model systems. Environ. Health Perspect. 106 (7) 357-384. [ Links ]
KIRBY MF, MORRIS S, HURST M, KIRBY SJ, NEALL P, TYLOR T and FAGG A (2000) The use of cholinesterase activity in flounder (Platichthys flesus) muscle tissue as a biomarker of neurotoxic contamination in UK estuaries. Mar. Pollut. Bull. 40 (9) 780-791. [ Links ]
KOHLER A (1989) Regeneration of contaminant-induced liver lesions in flounder - experimental studies towards the identification of cause-effect relationships. Aquat. Toxicol. 14 203-232. [ Links ]
LAU PS, WONG HL and GARRIGUES P (2004) Seasonal variation in antioxidative responses and acetylcholinesterase activity in Perna viridis in eastern oceanic and western estuarine waters of Hong Kong. Cont. Shelf Res. 24 1969-1987. [ Links ]
LUND AMADO L, DA ROSA CE, MEIRELLES LEITE A, MORAES L, VAZ PIRES W, LEAES PINHO GL, MARTINEZ GASPAR MARTINS C, BERTEAUX ROBALDO R, MAIA NERY LE, MONSERRAT JM, BIANCHINI A, ELIAS MARTINEZ P and GERACITANO LA (2006) Biomarkers in croakers Micropogonias furnieri (Teleostei: Sciaenidae) from polluted and non-polluted areas from the Patos Lagoon estuary (Southern Brazil): evidences of genotoxic and immunological effects. Mar. Pollut. Bull. 52 (2) 199-206. [ Links ]
MALINS DC, McCAIN BB, LANDAHL JT, MYERS MS, KRAHN MM, BROWN DW, CHAN SL and ROUBAL WT (1988) Neoplastic and other diseases in fish in relation to toxic chemicals: an overview. Aquat. Toxicol. 11 43-67. [ Links ]
McCARTHY JF and SHUGART LR (1990) Biological markers of environmental contamination. In: McCarthy JF and Shugart LR (eds.) Biomarkers of Environmental Contamination. Lewis Publishers, Florida. 3-14. [ Links ]
McLOUGHLIN N, YIN D, MALTBY L, WOOD RM and YU H (2000) Evaluation of sensitivity and specificity of two crustacean biochemical biomarkers. Environ. Toxicol. Chem. 19 (8) 2085-2090. [ Links ]
MINIER C, LEVY F, RABEL D, BOCQUENE G, GODEFROY D, BURGEOT T and LEBOULENGER F (2000) Flounder health status in the Seine Bay. A multibiomarker study. Mar. Environ. Res. 50 373-377. [ Links ]
MOCHIDA K, ITO K, HARINO H, TANAKA H, ONDUKA T, KAKUNO A and FUJII K (2009) Inhibition of acetylcholinesterase by metabolites of copper pyrithione (CuPT) and possible involvement in vertebral deformity of a CuPT-exposed marine teleostean fish. Comp. Biochem. Physiol. 149 (Part C) 624-630. [ Links ]
MODESTO KA and MARTINEZ CBR (2010) Roundup® causes oxidative stressing liver and inhibits acetylcholinesterase in muscle and brain of the fish Prochilodus lineatus. Chemosphere 78 294-299. [ Links ]
MOORE MJ, SHEA D, HILLMAN RE and STEGEMAN JJ (1996) Trends in hepatic tumours and hydropic vacuolation, fin erosion, organic chemicals and stable isotope ratios in winter flounder from Massachusetts, USA. Mar. Pollut. Bull. 32 (6) 458-470. [ Links ]
NAIGAGA I (2002) Bioaccumulation and histopathology of copper in Oreochromis mossambicus. M.Sc. Thesis, Rhodes University, Grahamstown. [ Links ]
ORR KK, BURGESS JE and FRONEMAN PW (2008) The effects of increased freshwater inflow on metal enrichment in selected Eastern Cape estuaries, South Africa. Water SA 34 (1) 39-52. [ Links ]
PAN G and DUTTA HM (1998) The inhibition of brain acetylcholinesterase activity of juvenile largemouth bass Micropterus salmoides by sublethal concentrations of diazinon. Environ. Res. 79 (section A) 133-137. [ Links ]
PRETORIUS J (2007) Personal communication, July 2007. State Veterinarian, Grahamstown. [ Links ]
RICHARDSON N (2008) A preliminary investigation into the use of biomarkers and a fish community index to assess estuarine health in selected Eastern Cape estuaries. M.Sc. thesis, Rhodes University, Grahamstown. [ Links ]
RINGWOOD AH, HOGUET J, KEPPLER CJ, GIELAZYN ML, WARD BP and ROURK AR (2003) Cellular biomarkers (lysosomal destabilization, glutathione and lipid peroxidation) in three common estuarine species: a methods handbook. Marine Resources Research Institute, South Carolina Department of Natural Resources. 45 pp. [ Links ]
ROEX EWM, KEIJZERS R and VAN GESTEL CAM (2003) Acetylcholinesterase inhibition and increased food consumption rate in zebrafish, Danio rerio, after chronic exposure to parathion. Aquat. Toxicol. 64 451-460. [ Links ]
ROY S, CHATTORAJ A and BHATTACHARYAS (2006) Arsenic induced changes in optic tectal histoarchitecture and acetylcholinesterase-acetylcholine profile in Channa punctatus: Amelioration by selenium. Comp. Biochem. Physiol. 144 (Part C) 16-24. [ Links ]
SCHARLER UM and BAIRD D (2003) The influence of catchment management on salinity, nutrient stochiometry and phytoplankton biomass of Eastern Cape estuaries, South Africa. Estuar. Coast. Shelf Sci. 56 735-748. [ Links ]
SCHULZ R (2001) Rainfall-induced sediment and pesticide input from orchards into the Lourens River, Western Cape, South Africa: importance of a single event. Water Res. 35 (8) 1869-1876. [ Links ]
SOLE M, KOPECKA J and GARCIA DE LA PARRA LM (2006) Seasonal variations of selected biomarkers in Sand Gobies Pomatoschistus minutus from the Guadalquivir Estuary, Southwest Spain. Arch. Environ. Contam. Toxicol. 50 249-255. [ Links ]
STEHR CM, JOHNSON LL and MyERS MS (1998) Hydropic vacuolation in the liver of three species of fish from the US West Coast: lesion description and risk assessment associated with contaminant exposure. Dis. Aquat. Org. 32 (2) 119-135. [ Links ]
STENTIFORD GD, LONGSHAW M, LYONS BP, JONES G, GREEN M and FEIST SW (2003) Histopathological biomarkers in estuarine fish species for the assessment of biological effects of contaminants. Mar. Environ. Res. 55 (2) 137-159. [ Links ]
VALAVANIDIS A, VLAHOGIANNI T, DASSENAKIS M and SCOULLOS M (2006) Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 64 (2) 178-189. [ Links ]
VAN DYK JC (2003) Fish histopathology as a monitoring tool for aquatic health: a preliminary investigation. M.Sc. thesis, Rand Afrikaans University, Johannesburg. [ Links ]
VAN DYK JC, PIETERSE GM and VAN VUREN JHJ (2007) Histological changes in the liver of Oreochromis mossambicus (Cichlidae) after exposure to cadmium and zinc. Ecotoxicol. Environ. Saf. 66 432-440. [ Links ]
VARO I, NAVARRO JC, AMAT F and GUILHERMINO L (2003) Effect of dichlorvos on cholinesterase activity of the European sea bass (Dicentrarchus labrax). Pestic. Biochem. Physiol. 75 61-72. [ Links ]
WESTER PW, CANTON JH, VAN IERSEL AAJ, KRAJNC EI and VAESSEN HAMG (1990) The toxicity of bis(tri-n-butyltin)oxide (TBTO) and di-n-butyltindichloide (DBTC) in the small fish species Oryzias latipes (medaka) and Poecilia reticulata (guppy). Aquat. Toxicol. 16 53-72. [ Links ]
WHITFIELD AK (1984) The effects of prolonged aquatic macrophyte senescence on the biology of the dominant fish species in a southern African coastal lake. Est. Coast. Shelf Sci. 18 (3) 315-329. [ Links ]
ZAR JH (1999) Biostatistical Analysis. Prentice Hall, New Jersey. [ Links ]
Received 15 May 2009; accepted in revised form 14 July 2010.
* To whom all correspondence should be addressed. +2746 622 2364; fax: +2746 622 6564; e-mail: n.richardson@cesnet.co.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}