










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkWater SA
On-line version ISSN 1816-7950
Print version ISSN 0378-4738
Water SA vol.36 n.3 Pretoria Apr. 2010
Ecological impacts of small dams on South African rivers Part 2: biotic response – abundance and composition of macroinvertebrate communities
Sukhmani K Mantel*; Nikite WJ Muller; Denis A Hughes
Unilever Centre for Environmental Water Quality – Institute for Water Research (UCEWQ-IWR), Rhodes University, PO Box 94, Grahamstown, South Africa
ABSTRACT
This paper investigates the cumulative impacts of small dams on invertebrate communities in 2 regions of South Africa – the Western Cape and Mpumalanga. Previous research found reduced discharge, increased total dissolved salts, and a decrease in average score per taxon (ASPT; collected using SASS4 methods) at sites with high density of small dams in their catchment. These changes in ASPT are investigated using the invertebrate abundance data available in the River Health Programme. Multivariate analyses found differences in invertebrate communities in rivers with high densities of small dams in their catchment in foothill-gravel streams (in both Western Cape and Mpumalanga) and in foothill-cobble streams (in Western Cape only). Opportunistic taxa that are tolerant of pollution, and capable of exploiting various habitats, and those that prefer slower currents increased in numbers, while other taxa that are sensitive to pollution and disturbance declined in numbers. Some regional differences were noted possibly reflecting climatic differences between the regions. Since the results of this study are correlative, it highlights the need for a systematic (by sites and seasons) and detailed (at species level) collection of data to verify the results of cumulative effects of small dams. This can further the development of a framework for small-dam construction and management that will limit their impact on river catchments.
Keywords: cumulative impacts, reduced low flows, environmental water quality, Ephemeroptera, Trichoptera
Introduction
The Serial Discontinuity Concept stresses the importance of longitudinal, lateral and vertical connectivity of rivers to the health and biodiversity conservation of a riverine ecosystem. The predictions of the concept regarding the impacts of regulation on riverine biodiversity have been tested in various rivers (Stanford and Ward, 2001). Impoverishment of riverine fauna, replacement by lentic species, and possible recovery of fauna with distance from the impoundment, particularly for macroinvertebrates below a confluence with an unregulated tributary, has been established (e.g. Hauer et al., 1989; Stanford and Ward, 1989; Bournaud et al., 1996; Pringle et al., 2000). The serial discontinuity concept is primarily relevant to large dams (>3x106 m3 or >15 m height; World Commission on Dams, 2000) with deep or hypolimnial releases, which contrasts with small dams and weirs that are generally surface releasing. However, the principle of the accumulation of the impacts of dams is one that this paper investigates in terms of the ecological cumulative impacts of small dams.
Individual small dams and the associated changes in ecological communities below them have received attention for facilitating the spread of pest species (e.g. black-flies: Palmer and O'Keeffe, 1995), acting as barriers to migratory species (e.g. amphidromous shrimps: Benstead et al., 1999), reducing macroinvertebrate abundance and diversity (Ogbeibu and Oribhabor, 2002, but see Cortes et al., 1998 for variation in impacts by dam usage), and reducing fish densities (Lessard and Hayes, 2003). Authors of these studies have linked these changes to the variation in water quality, water quantity, particle size of organic matter, and habitat modification, amongst other factors. The permanence of changes in the ecosystem variables was investigated by Doyle et al. (2005) for 5 small dams that were removed from US rivers. They found that macroinvertebrate communities, as an ecosystem response variable, were more resilient in comparison to riparian vegetation; however, some invertebrate species, including some endangered unionid mussels, recovered only partially following the removal of dams.
In South African rivers, research on the effects of changes in discharge on riverine macroinvertebrates have focused on two aspects – flow reduction due to a series of impoundments (all large, except one small dam), and flow increase and regime change due to inter-basin transfers. Palmer and O'Keeffe (1995) found that no flow below dams during low-flow periods led to the absence of black-flies below a small dam in an Eastern Cape river. Additionally, the dams shifted the distribution of some blackfly species (many of which are pest species) upstream. On the other hand, increased flow volumes due to inter-basin transfer have led to the dominance of a livestock pest species Simulium chutteri over other simuliid species (O'Keeffe and De Moor, 1988; Rivers-Moore et al., 2007). This summary of South African research on the impacts of dams on rivers highlights the gap in our knowledge on the effects of small dams. This paper thus focuses on the impacts on riverine fauna due to an accumulation of small dams in catchments which we refer to as 'cumulative impacts of small dams'.
South Africa records approximately 500 000 small farm dams (DWAF, 1986), which are primarily associated with agriculture and livestock farming. The present research has investigated their potential cumulative impacts on South African rivers and the analyses are presented as 2 papers. Part 1 of this series looked at the impacts of small dams (defined to be <3x106 m3 storage capacity; World Commission on Dams, 2000) on water quantity and quality (Mantel et al., 2010). We found significant reductions in base flows at sites with high small-dam density in 2 South African regions – Western Cape and Mpumalanga. Additionally, we quantified the impacts on the water quality in terms of an increase in total dissolved salts and a decrease in average score per taxon (ASPT, a measure of river health based on field collection of invertebrate families; Chutter, 1998; Dickens and Graham, 2002) in these 2 regions. Fig. 1 shows the change in SASS4 score and ASPT for 6 range classes of small-dam densities in these 2 regions. While the index values for the lower 3 range classes (corresponding to SDLo) do not show a trend, there is a clear trend of reduction in SASS4 score and ASPT values in the Western Cape and for SASS4 score in Mpumalanga for the range classes corresponding to SDHi. For this reason, this value has been used as a threshold to differentiate between sites with low and high small-dam densities in Mantel et al. (2010). This paper investigates the significant reduction in the ASPT index at sites with high small-dam densities found in Mantel et al. (2010), in terms of changes in the macroinvertebrate communities relevant to river health. The macroinvertebrate changes are investigated in the light of our ecological knowledge about invertebrate taxa regarding their distribution and preferences for habitat, flow and water quality. These research results can assist in determining environmental water requirements in the context of legislation protecting South African water resources (National Water Act, No. 36 of 1998), its environment (National Environmental Management Act, No. 107 of 1998), and its biodiversity (Biodiversity Act, No. 10 of 2004).
Methods
Mantel et al. (2010) found significant reductions in ASPT for foothill-gravel streams in both Mpumalanga and Western Cape, and in foothill-cobble streams in the Western Cape only, at high small-dam densities (Table 1; Fig. 1). For the present paper, data for the invertebrate communities and the river health indices were obtained from the South African River Health Programme (RHP, 2006), in order to investigate the changes in these communities. Analyses were conducted in 2 provinces – Western Cape and Mpumalanga – as these 2 regions comprised the majority of information in the RHP database. The Western Cape, located on the southwest coast of South Africa, has a temperate climate and receives winter rainfall, while Mpumalanga, located in the eastern part of the country, has a subtropical climate with summer rainfall. Both regions have an abundance of small dams (14 257 natural and man-made water bodies including small dams in the Western Cape and 10 040 of them in the Mpumalanga region; Chief Directorate of Surveys and Land Information, 1999), which are associated with vineyards and other cultivation in the Western Cape, and with stock farming, cultivation and exotic timber plantations in Mpumalanga. The data used and the analyses conducted were as follows:
Quantification of potential dam impacts
Part 1 of this series of 2 papers (Mantel et al., 2010) detailed the potential impact from small dams (< 3x106 m3; WCD, 2000) for each RHP-sampled location, in terms of the number of small dams in the catchment of each RHP sampling point included in the analyses, after standardising the density of dams to the square root of catchment area in km2 in order to reduce catchment size bias. Sites with low small-dam density in their catchment are referred to here as SDLo and those with high density as SDHi, where the low level for small-dam density refers to <5 dams per square-root catchment area (in km2) of the sampling location (refer to Fig. 1 and Mantel et al., 2010 for an explanation of the derivation of this threshold). Data used to calculate the catchment area were obtained from Midgley et al. (1994) and the information on the location and the area of small dams was acquired from a dams GIS coverage produced by the Chief Directorate of Surveys and Land Information (1999).
Biological data
The RHP reports family-level macroinvertebrate data and river health indices determined using the rapid river health monitoring protocol, the South African Scoring System (SASS; Chutter, 1998; Dickens and Graham, 2002). SASS is an index based on the sensitivity of macroinvertebrate families to pollution and disturbance. The invertebrate data collected using the SASS4 methodology (Chutter, 1998) is reported as 6 range classes of abundance: 0 (taxon absent), 1 (1 individual present), A (2 to 10 individuals), B (10 to 100 individuals), C (100 to 1 000 individuals), D (>1 000 individuals). For the present analyses, these ranges were replaced by the median value for each range (i.e. 0, 1, 6, 55, 550, respectively, for the classes 0, 1, A, B and C, respectively) and the range class D was replaced by 1 000 since an upper limit for this range cannot be identified. The RHP records the number of morphospecies found in the sample for 3 groups (Baetidae, Hydropsychidae and Trichoptera Case Caddis). To avoid the bias generated by the link between presence of different morphospecies groups, e.g. the presence of Baetidae 3 species means that Baetidae 1 and 2 species are also present, we have combined the morphospecies groups into 1 category for each taxon for the analyses, namely 'Baetidae All Species', 'Hydropsychidae All Species' and 'Trichoptera Case Caddis All Species'. For the RHP, the taxa are collected from 3 broadly-defined biotopes (stones, gravel/sand/mud and aquatic vegetation). A separate score is acquired for each of these biotopes, and a final SASS score is obtained for the sample by summing the SASS score for the taxa collected. The number of taxa (family level) and average score per taxon (ASPT, which is the SASS score divided by the number of taxa) are also calculated using the SASS4 (South African Scoring System, Version 4) methodology. The SASS4 index is equivalent to those used in the UK (Biological Monitoring Working Party (BMWP); Armitage et al., 1983) and in Australia (Stream Invertebrate Grade Number Average Level, SIGNAL; Chessman et al., 1997). Part 1 of this series of papers (Mantel et al., 2010) analysed the differences between the various macroinvertebrate indices for low vs. high densities of small dams (SDLo and SDHi as defined above). We had limited the samples analysed by Part 1 to those where water quality data was reported, since we were analysing differences in the water quality and the indices at varying densities of small dams. In order to increase the dataset for analyses in this paper (Part 2), we have included all the samples collected for the RHP sites used in Part 1 at the 2 small dam densities (SDLo and SDHi).
Analysis of dam impacts on macroinvertebrate communities
Statistical testing (Student's t-test or Mann-Whitney U test in cases where the assumptions of normality and homoscedacity were not met; Statistica, StatSoft, 2003) was conducted separately for foothill-gravel and foothill-cobble streams to avoid the bias of inherent sub-regional variation found by other researchers in South Africa (De Moor 2002; Dallas 2004). Sub-regions and geomorphological zones are distinguished by the stream gradient and the dominance of various substrate types, such as boulders, bedrock, cobble, etc. (Dallas, 2000). Analysis of Similarity (ANOSIM) was conducted on the fourth-root transformed data using the Bray-Curtis Similarity Measure to assess changes in the communities associated with low vs. high density of small dams using PRIMER (Clarke and Gorley, 2001) software. The output of ANOSIM is the test statistic R, which can vary from –1 to 1 (Clarke and Gorley, 2001). A value of zero for R indicates a true null hypothesis (no difference between treatments). An R > 0 indicates that intra-treatment variation among replicates (i.e. variation within each of the groups SDLo and SDHi) is less than inter-treatment variation (i.e. between the groups SDLo and SDHi), while the opposite is true for R < 0. Non-metric Multi Dimensional Scaling (nMDS) and Simper (Similarity Percentages – species contributions) were conducted for cases with significant results for ANOSIM.
Results
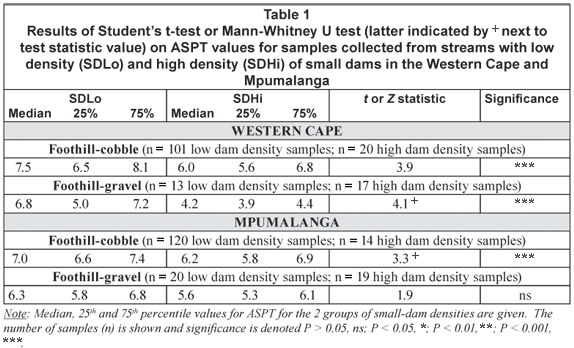
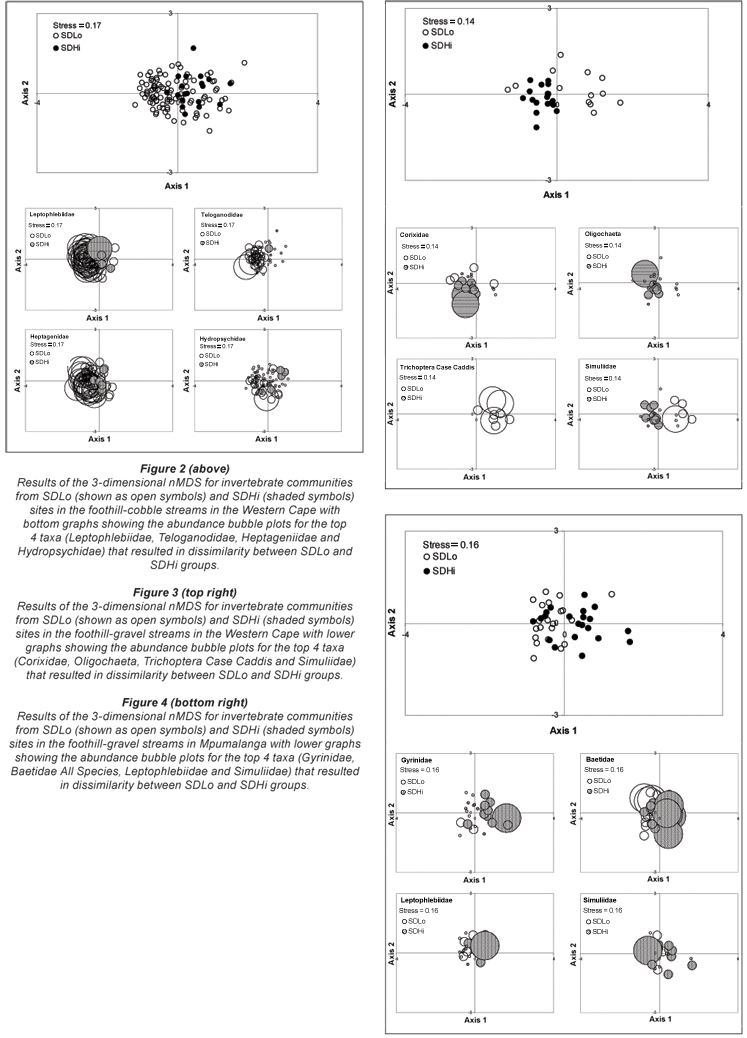
Changes in abundances of invertebrate families analysed using ANOSIM found significant differences in the foothill-cobble invertebrate communities in the Western Cape region for low vs. high small-dam density samples (Global R = 0.154, P = 0.014; nSDLo = 101; nSDHi = 20). Simper analysis found 54% dissimilarity between the invertebrate communities for high vs. low small-dam density groups in the foothill-cobble streams in the Western Cape. The first 2 axes of the 3-dimensional nMDS output with bubble plots (with the size of the bubble representing the density of the taxon in the sample) for the top 4 taxa resulting in 18% cumulative dissimilarity between the low and high density groups is shown in Fig. 2. A decrease in Leptophlebiidae, Teloganodidae, Heptageniidae and Hydropsychidae All Species at high density sites is visible. Analysis of invertebrate communities collected from the foothill-gravel streams in the Western Cape also found significant differences between low and high small-dam density samples (Global R = 0.507, P = 0.001; number of samples, nSDLo = 13; nSDHi = 17) and Simper analysis found 63% dissimilarity between the communities. The first 2 axes of the 3-dimensional nMDS output with bubble plots for the top 4 taxa resulting in 18% cumulative dissimilarity shows an increase in Corixidae and Oligochaeta along with a decrease in Trichoptera Case Caddis All species and Simuliidae at sites with high small-dam density in these streams (Fig. 3).
For the Mpumalanga region, no significant difference was detected in invertebrate communities between sites with low vs. high small-dam density in their catchments in the foothill-cobble streams (Global R = 0.15, P = 0.054; number of samples, nSDLo = 120; nSDHi = 14), although ASPT was significantly different. For foothill-gravel streams, ASPT was borderline on significance (P = 0.06) and the test for community differences found significant results (global R = 0.12, P = 0.002; nSDLo = 21; nSDHi = 21). Simper analysis revealed 55% dissimilarities between the high and low small-dam density groups in the foothill-gravel streams. The main 2 axes of the 3-dimensional nMDS plots for the top 4 taxa resulting in 15% cumulative dissimilarities between the small dam groups in the foothill-gravel streams is shown in Fig. 4 (with overall increased abundances of Gyrinidae, Baetidae All Species, Leptophlebiidae and Simuliidae at high density sites).
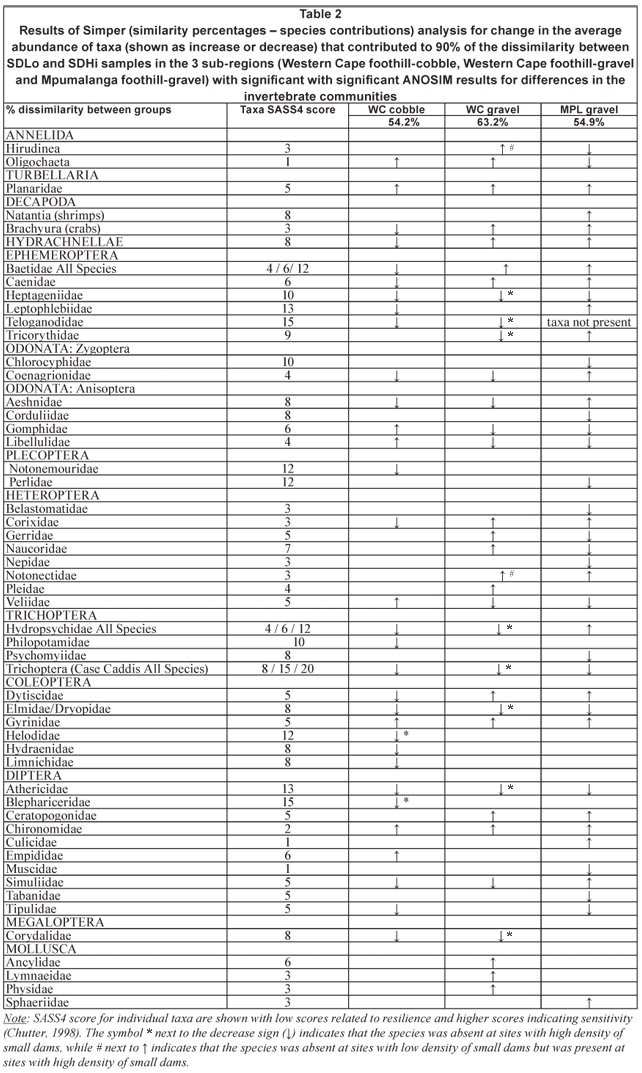
Table 2 summarises the trends (increase or decrease) in the average abundance for the taxa contributing to 90% of the dissimilarity in the invertebrate communities between sites with low and high small-dam density for the 3 group comparisons where ANOSIM found significant differences. In general, the abundance of taxa with low SASS4 scores (signifying species that are less sensitive to pollution and disturbance, and primarily belonging to Turbellaria, Mollusca and Heteroptera) increased, and those with high SASS4 score (i.e. more sensitive species, mostly Odonata, Trichoptera and some Ephemeroptera) declined. Additionally, there were no regional or sub-regional differences in the response of some taxa, as indicated by consistent increases or decreases in the 3 groups with significant differences in the invertebrate communities (see Table 2), e.g. increases in Planaridae, Gyrinidae and Chironomidae, and decreases in Heptageniidae, Trichoptera (Case Caddis All Species), Elmidae/Dryopidae and Athercidae. In comparison, regional and sub-regional differences were noted for some taxa that were absent from collections at sites with high small-dam density; 8 taxa in the foothill-gravel streams of Western Cape were absent at high density sites, while in the foothill-gravel streams of Mpumalanga and in the foothill-cobble streams of the Western Cape either none or only two of the taxa were missing at these sites.
Discussion
River flows (floods, droughts, high pulses, base-flows, etc.) serve different functions in moulding the available physical habitat. They act as cues triggering various life history stages of riverine organisms and thus play a part in dictating which organisms are found where and when (Petts, 1984; Uys and O'Keeffe, 1997; Richter et al., 1997; Bunn and Arthington, 2002). Alteration of a river's flow regime through the building of large dams can lead to impacts on animal and plant communities via the disruption of longitudinal and lateral connectivity, loss of thermal and flow cues, changed habitat and hydraulics, and the establishment of exotic species (Bunn and Arthington, 2002). Part 1 of this series of paper (Mantel et al., 2010), investigating the cumulative impacts of small dams in South Africa, proposed that the link between reduced water quantity during low flow periods and the increased concentrations of water quality variables (particularly through an increase in total dissolved salts) found in the rivers of 2 regions (Western Cape and Mpumalanga) was the main reason for the changes noted in ASPT. This paper documents the changes in the invertebrate communities and uses the available information on their sensitivities to pollution and disturbance and their habitat preferences to support this hypothesis. Note that although we refer to decrease in discharge or changed water quality (investigated in Mantel et al., 2010) at various locations in this paper, the changed parameters could in fact be various interlinked factors such as a specific component of the flow, or the resultant change in available biotopes. For example, in a comprehensive study of 52 sites on Swedish rivers with hydroelectric dams, Malmqvist and Englund (1996) found that various species of mayflies responded to different flow parameters, including peak flow, reduced discharge and constancy of flow. Jowett's (2003) study of 4 common invertebrates of New Zealand streams (3 mayfly and 1 net-spinning caddisfly species) found specific near-bed hydraulic ranges that these organisms preferred. In South African rivers, Dallas (2007) noted that samples from stones biotopes had higher scores for SASS4, number of taxa and ASPT relative to vegetation and sand samples and, additionally, the taxa with high SASS sensitivity scores were found more frequently on stones, particularly taxa in the orders of Ephemeroptera, Plecoptera and Trichoptera. This suggests that changes in the availability of different biotopes might influence community structure, although streambed drying would probably be more influential than available habitat for macroinvertebrates (Chadwick and Huryn, 2007). Future studies would need to tease out these various relationships for South African streams in order to fully comprehend the reasons behind the taxon-specific abundance changes noted at reduced discharge sites in our study.
Response of Ephemeroptera, Diptera and predators
Our study noted increased abundance for taxa that are capable of exploiting various biotopes and currents (e.g. Planaridae, Gyrinidae, chironomids), and for those that prefer slower currents and stagnant waters (such as Natantia shrimps, Notonectidae, Mollusca), at sites with high small-dam density. Note that the habitat and flow preferences for South African taxa have been derived from Chutter (1970), Schael and King (2005) and Thirion (2007). The top 4 taxa groups that changed in abundance in our study primarily belonged to the orders Ephemeroptera, Diptera and Trichoptera.
Ephemeroptera are one of the most diverse orders of aquatic invertebrates in streams across the world. Our study found increases in some taxa (Baetidae, Caenidae) and decreases in others (Teloganodidae and Heptageniidae). The changes observed are supported by Brittain and Saltveit's (1989) review on the effects of river regulation on mayflies, with decreases in Heptageniidae and increases in Baetidae due to flow reduction. Brittain and Saltveit (1989) suggest dietary and habitat reasons for the observed patterns, as the increase of certain baetids below dams can be linked to greater algal growth (as a food source and predation refuge), while heptageniids that prefer clean rocks for attachment might decrease in numbers below dams. Similarly, reduced baseflows below a 4 m high dam, used for diverting water, led to periphyton growth and increased numbers of ephemeropterans that fed on algae or that were deposit collectors (Cortes et al., 1998). Although we lack information on algal growth below small dams in the present study, it is expected that a combination of reduced flows, decreased scouring and increased nutrient concentrations would result in greater algal growth (Biggs and Close, 1989; Palmer and O'Keeffe, 1990).
The decrease in simuliid abundance below a high number of small dams in the Western Cape streams is probably correlated with reduced flow as the results are similar to findings of reduced number, and at times absence, of simuliids below a small dam during low flow periods on the Buffalo River, South Africa (Palmer and O'Keeffe, 1995). Simuliids use their cephalic fans to filter organic matter from water currents and they are, therefore, rheophilic (Crosskey, 1990). The relationship between flow and simuliid densities has been observed in a US stream where simuliid densities were higher during years with non-scouring elevated flow compared to years of low flow (Schlosser and Ebel, 1989). Palmer and O'Keeffe (1995) also reported that the smallest of the 4 dams on the Buffalo River (10 m high Maden Dam; 0.32x106 m3 storage capacity) had the greatest impact on blackfly species distribution, by shifting the distribution of blackfly species from foothills to upstream areas, particularly for pest species of blackflies. In the present study, data was mostly available at family level, which obscured observation of any pattern of changes in species distribution.
The present study found a downward trend in the abundances of predatory species (Gomphidae, Libellulidae, Tipulidae), as well as a decrease in the abundances and the diversity of trichopteran species (e.g. decreased abundance, and absence at some sites, for Hydropsychidae and case-building caddisflies). Altered hydraulics, reduced habitat diversity (due to bed 'armouring') and decreased availability of sediment for burrowing and building cases can lead to declines in abundance and diversity of gomphid odonates and certain net-spinning hydropsychid trichopterans (e.g. Hauer and Stanford, 1982; Hauer et al., 1989; Schlosser and Ebel, 1989; Donnelly, 1993; but see Parker and Voshell, 1983 for increased production of Trichoptera below a large dam, and Cortes et al., 1998 for increased abundance of some shredder and net-spinning species below a small dam). A review of macroinvertebrate responses to impoundments by Petts (1984) found similar numbers of studies documenting increasing and decreasing trends in caddisfly abundance below dams and he commented that the variation was due to the river's hydrological characteristics and the dam's water release depth. Predators can be important in structuring both the abundance and the diversity of the prey community and the strength of these interactions can be altered by dams due to reduced flow variability and resultant changes in available habitat and water quality (Allan, 1995; Power et al., 1996). Assuming that certain predatory species are absent at sites with high small-dam density and that this is not a chance result due to the time of sampling, their absence could be an additional reason for the observed invertebrate community change in this study. On the other hand, it is noteworthy that for some animals and plants, farm dams can represent important refuges during low flow and drought periods (e.g. Samways, 1989; Brainwood and Burgin, 2006). Future studies will need to investigate the importance of abiotic forces (such as hydraulics, geomorphology and water quality) relative to biotic forces (predation, competition, food availability) for macroinvertebrate communities below small dams. Insight into the permanence of changes resulting from small dams is provided by studies investigating the recovery rates of ecological attributes following the removal of small dams (Stanley et al., 2002; Doyle et al., 2005). Individual species can recover following dam removal, depending on their life cycle and the position of the dam in the landscape (upstream vs. lowland), and, generally, the populations from the same stream are the primary source of re-colonisers, particularly for species that do not have a flying adult stage (Hughes, 2007).
Regional variation in invertebrate response
Regional differences in the community response (in terms of absence of taxa at sites with high density of small dams) were noted in the present study that are possibly linked to climatic differences and additionally to the magnitude of water quantity change in the regions (i.e. greater reductions in the Western Cape than in Mpumalanga were noted by Mantel et al., 2010). Heptageniidae and Tricorythidae, 2 ephemeropteran taxa with high sensitivity, noted by their SASS4 taxa score of 10 to 15, were absent at sites with high small-dam density in the foothill-gravel rivers of Western Cape. However, in Mpumalanga, Heptageniidae showed a decrease, although not complete absence, in foothill-gravel streams with high density of small dams, while Trichorythidae increased in abundance in these streams. Similarly, 2 trichopteran taxa (Hydropsychidae and cased caddisflies) that completely disappeared at sites with high small-dam density in the foothill-gravel streams of Western Cape, showed either increased or decreased abundance in Mpumalanga, but not complete absence. A confounding reason for the differences in taxon response could be the time of collection since a previous study has found seasonal variation in invertebrate communities in Western Cape rivers (Dallas, 2004). In our study, the Western Cape samples were collected during all 4 seasons, although all seasons were not sampled equally and samples from winter and autumn were sparse in our database; therefore, dominance of samples from some seasons in the database could result in bias.
The absence of Trichoptera in foothill-gravel streams with high small dam density in the Western Cape is of particular concern as the Cape Floral Kingdom has the highest diversity of species in addition to the largest number of endemic families and genera of Trichoptera in South Africa (De Moor 2006). Ecologists have debated whether absence of species affects ecosystem functionality. Cardinale et al. (2000 p. 176) suggest that 'there may be no single, generalisable relationship between the number of species and the functioning of ecosystems because the relative contributions of species diversity and species composition to ecological processes change with environmental context'. Davies et al. (1994) suggest that the biota of dry-land rivers, as in South Africa, are more opportunistic than those from systems with higher rainfall, because the fauna and flora in these systems evolved in an environment that is highly variable, particularly due to the very low conversion ratio of rainfall to runoff. This opportunism and flexibility is evident from the results of gut contents of 12 macroinvertebrate species collected from various sites (including above and below dams) and seasons from a turbid South African river (Buffalo River). The authors found a wide overlap in the diet of Ephemeropteran and Trichopteran species, despite different functional morphology and behaviour (Palmer et al., 1993a; b), suggesting that ecosystem functioning in this river was somewhat more resilient to the loss of some species. The relationship of ecosystem functioning with species diversity and the effect of high density of small dams needs further investigation in the South African context.
Limitations of SASS
Our results highlight a limitation of macroinvertebrate indices that is well-known by practitioners. In the foothill-cobble streams in Mpumalanga, significant differences in ASPT were found (mean ASPT changed from 7.0 for low small-dam density samples to 6.2 for high small-dam density), although the community differences were not significant. Additionally, the situation was reversed for foothill-gravel streams in Mpumalanga where the decrease in ASPT at sites with high density of small dams was not significantly different from that at sites with low density of small dams, although the P value was borderline while the community was significantly different according to ANOSIM. The non-significance of ASPT value between high and low density sites could have resulted from the replacement of species by others that have similar SASS4 score. These results caution about taking indices at face value, since the overall index number can hide the fact that the invertebrate community might be different, in addition to other shortfalls of the SASS4 index, described in detail by De Moor (2002). Family level identification that forms the basis of SASS is especially of concern for a number of speciose families such as Chironomidae, Leptoceridae and Simuliidae.
Study limitations and conclusion
Our research results have some limitations to their interpretation, some of which have been noted in Part 1 of this series of 2 papers (Mantel et al., 2010). Most importantly, due to the correlative nature of our analyses, the confounding effects of other land-use impacts, such as agriculture and forestry, which are correlated with, and are sometimes the reason for, the building of small dams, cannot be disregarded and would need to be investigated in future studies. In this regard, there is a need to develop diagnostic indices that incorporate the sensitivity of various invertebrate taxa, preferably at the species level, so as to distinguish impacts of small dams from other anthropogenic impacts, as has been done for Australian and US invertebrates (Chessman and McEvoy, 1998; US EPA, 2000). Specific to the invertebrate data analysed in this paper, 2 major limitations stand out. Although invertebrate communities showed differences at the level of small-dam density, the data were at a coarse level (family level and abundance ranges), which might hide or possibly exaggerate the patterns that we observed. Additionally, there was large discrepancy in the number of replicates for the low and high density groups in foothill-cobble streams, which could have biased our results. However, this problem could not be resolved since we included all the available data. Future studies can also investigate seasonal variation in the invertebrate assemblage below high density small-dam sites if data are reported consistently. Another limitation of the data was that we were only able to investigate the downstream effects of small dams, while ignoring the upstream implications of these dams at the level of genes/species (reduced genetic flow due to isolation), population/community (e.g. downstream areas providing habitat for exotic species that might spread to upstream areas) and possibly ecosystem/landscape (e.g. build up of mercury in reservoirs) (Pringle, 1997). We suggest incorporation of small dam effects into the Serial Discontinuity Concept (Stanford and Ward, 2001) that presently only focuses on large dams.
South Africa faces various challenges in implementing the ecological Reserve (equivalent to the environmental water requirement, which refers to both the quantity and quality of water required to protect the aquatic ecosystems) due to our limited understanding of the links between water quantity and quality (Palmer et al., 2005). Biodiversity conservation of rivers, even in world-famous areas like the Kruger National Park, requires reassignment of land to allow protection of upper catchment areas. However, at present this is only a 'theoretical' exercise as land redistribution takes higher priority due to South Africa's political history (Roux et al., 2008). As noted in the introduction, with few exceptions most of the studies cited in this paper have investigated either large dam impact or individual small dams. This paper is the first to study the cumulative impacts of small dams in South Africa and the results highlight their potential impacts on South African water resources. Much more remains to be done, however, and the areas that need particular attention include quantifying the impacts using a systematic (by sites and seasons) and detailed (at species level) approach that will allow testing hypotheses of the cause and effect of these small dams.
Acknowledgements
We would like to acknowledge the Rhodes University Postdoc Fellowship which allowed Sukhmani Mantel to conduct this work. Staff of UCEWQ-IWR contributed helpful suggestions and comments that the authors are grateful for. Comments on a draft version of the paper by Dr. Ferdy de Moor helped improve the manuscript.
References
ALLAN JD (1995) Predation and its consequences. In: Allan JD, Stream Ecology: Structure and Function of Running Waters. Chapman and Hall, London, UK. 163-185. [ Links ]
ARMITAGE PD, MOSS D, WRIGHT JF and FURSE MT (1983) The performance of a new biological water quality score system based on macroinvertebrates over a wide range of unpolluted running-water sites. Water Res. 17 333-347. [ Links ]
BENSTEAD JP, MARCH JG, PRINGLE CM and SCATENA FN (1999) Effects of a low-head dam and water abstraction on migratory tropical stream biota. Ecol. Appl. 9 656-668. [ Links ]
BIGGS BJF and CLOSE ME (1989) Periphyton biomass dynamics in gravel bed rivers: the relative effects of flows and nutrients. Freshwater Biol. 22 209-231. [ Links ]
BOURNAUD M, CELLOT B, RICHOUX P and BERRAHOU A (1996) Macroinvertebrate community structure and environmental characteristics along a large river: Congruity of patterns for identification to species or family. J. N. Am. Benthol. Soc. 15 232-253. [ Links ]
BRAINWOOD M and BURGIN S (2006) An exploration of the relationships between macroinvertebrate community composition and physical and chemical habitat characteristics in farm dams. Environ. Monit. Assess. 119 459-480. [ Links ]
BRITTAIN JE and SALTVEIT SJ (1989) A review of the effect of river regulation on mayflies (Ephemeroptera). Regulated Rivers: Res. Manage. 3 191-204. [ Links ]
BUNN SE and ARTHINGTON AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manage. 30 492-507. [ Links ]
CARDINALE BJ, NELSON K and PALMER MA (2000) Linking species diversity to the functioning of ecosystems: On the importance of environmental context. Oikos 91 175-183. [ Links ]
CHADWICK MA and HURYN AD (2007) Role of habitat in determining macroinvertebrate production in an intermittent-stream system. Freshwater Biol. 52 240-251. [ Links ]
CHESSMAN BC, GROWNS JE and KOTLASH AR (1997) Objective derivation of macroinvertebrate family sensitivity grades numbers for SIGNAL biotic index: application to the Hunter River System, New South Wales. Mar. Freshwater Res. 48 159-172. [ Links ]
CHESSMAN BC and McEVOY PK (1998) Towards diagnostic biotic indices for river macroinvertebrates. Hydrobiol. 364 169-182. [ Links ]
CHIEF DIRECTORATE OF SURVEYS and LAND INFORMATION (1999) Dams and Lakes of South Africa from 1:50 000 Topographical Map Blue Plates. Polygon coverage generated by Mike Silberbauer, The Institute for Water Quality Studies, Pretoria, South Africa. [ Links ]
CHUTTER FM (1970) Hydrobiological studies in the catchment of the Vaal Dam, South Africa. Part 1. River zonation and benthic fauna. Int. Rev. Ges. Hydrobiol. 55 445-494. [ Links ]
CHUTTER FM (1998) Research on the Rapid Biological Assessment of Water Quality Impacts in Streams and Rivers. WRC Report No. 422/1/98. Water Research Commission Pretoria, South Africa, 265 pp. [ Links ]
CLARKE KR and GORLEY RN (2001) PRIMER v5: User Manual/Tutorial. PRIMER-E Ltd., Plymouth, UK. [ Links ]
CORTES RMV, FERREIRA MT, OLIVEIRA SV and GODINHO F (1998) Contrasting impact of small dams on the macroinvertebrates of two Iberian mountain rivers. Hydrobiol. 389 51-61. [ Links ]
CROSSKEY RW (1990) The Natural History of Blackflies. John Wiley and Sons, UK. 711 pp. [ Links ]
DALLAS HF (2000) Ecological Reference Condition Project: Field Manual. Volume 1: General Information, Catchment Condition, Invertebrates and Water Chemistry. Report No. 10, Institute for Water Quality Studies, Department of Water Affairs and Forestry, Pretoria, South Africa, 34 pp. [ Links ]
DALLAS HF (2004) Seasonal variability of macroinvertebrate assemblages in two regions of South Africa: implications for aquatic bioassessment. Afr. J. Aquat. Sci. 29 173-184. [ Links ]
DALLAS HF (2007) The influence of biotope availability on macroinvertebrate assemblages in South African rivers: implications for aquatic bioassessment. Freshwater Biol. 52 370-380. [ Links ]
DAVIES BR, THOMS MC, WALKER KF, O'KEEFFE JH and GORE JA (1994) Dryland rivers: Their ecology, conservation and management. In: Calow P and Petts GE (eds.) The Rivers Handbook. Volume 2. Blackwell Scientific Publications, Oxford, UK. 484-511. [ Links ]
DE MOOR FC (2002) Shortcomings and advantages of using rapid biological assessment techniques for the purpose of characterizing rivers in South Africa. Verh. Internat. Verein. Limnol. 28 651-662. [ Links ]
DE MOOR FC (2006) Regional biogeographical differences in Trichoptera diversity in South Africa: observed patterns and processes. In: Bueno-Soria J, Barba-Álvarez R and Armitage B (eds.) Proc. 12th Internat. Symp. Trichop, 18-22 June 2006. The Caddis Press, Mexico. 211-218. [ Links ]
DICKENS CWS and GRAHAM PM (2002) The South African Scoring System (SASS) version 5 rapid bioassessment method for rivers. Afr. J. Aquat. Sci. 27 1-10. [ Links ]
DONNELLY TW (1993) Impoundment of rivers: sediment regime and its effect on benthos. Aquat. Conserv: Mar. Freshwater 3 331-342. [ Links ]
DOYLE MW, STANLEY EH, ORR CH, SELLE AR, SETHI SA and HARBOUR JM (2005) Stream ecosystem response to small dam removal: lessons from the heartland. Geomorphol. 71 227-244. [ Links ]
DWAF (DEPARTMENT OF WATER AFFAIRS AND FORESTRY, SOUTH AFRICA) (1986) Management of the Water Resources of the Republic of South Africa. Department of Water Affairs and Forestry, Pretoria, South Africa. [ Links ]
HAUER FR and STANFORD JA (1982) Ecological responses of hydropsychid caddisflies to stream regulation. Can. J. Fish. Aquat. Sci. 39 1235-1242. [ Links ]
HAUER FR, STANFORD JA and WARD JV (1989) Serial discontinuities in a Rocky Mountain river. II. Distribution and abundance of Trichoptera. Regulated Rivers: Res. Manage. 3 177-182. [ Links ]
HUGHES JM (2007) Constraints on recovery: using molecular methods to study connectivity of aquatic biota in rivers and streams. Freshwater Biol. 52 616-631. [ Links ]
JOWETT IG (2003) Hydraulic constraints on habitat suitability for benthic invertebrates in gravel-bed rivers. Rivers Res. Appl. 19 495-507. [ Links ]
LESSARD JL and HAYES DB (2003) Effects of elevated water temperature on fish and macroinvertebrate communities below small dams. Rivers Res. Appl. 19 721-732. [ Links ]
MALMQVIST B and ENGLUND G (1996) Effects of hydropower-induced flow perturbations on mayfly (Ephemeroptera) richness and abundance in north-Swedish river rapids. Hydrobiol. 341 145-158. [ Links ]
MANTEL SK, HUGHES DA and MULLER WJ (2010) Ecological impacts of small dams on South African rivers: Part I. Drivers of change - water quantity and quality. Water SA 36 (3) 351-360. [ Links ]
MIDGLEY DC, PITMAN WV and MIDDLETON BJ (1994) Surface Water Resources of South Africa 1990. Six Volumes. WRC Report No. 298/1/94. Water Research Commission, Pretoria, South Africa. [ Links ]
OGBEIBU AE and ORIBHABOR BJ (2002) Ecological impact of river impoundment using benthic macro-invertebrates as indicators. Water Res. 36 2427-2436. [ Links ]
O'KEEFFE JH and DE MOOR FC (1988) Changes in the physico-chemistry and benthic invertebrates of the Great Fish River, South Africa, following an interbasin transfer of water. Regulated Rivers: Res. Manage. 2 39-55. [ Links ]
PALMER RW and O'KEEFFE JH (1990) Downstream effects of impoundments on the water chemistry of the Buffalo River (Eastern Cape), South Africa. Hydrobiol. 202 71-83. [ Links ]
PALMER C, O'KEEFFE J, PALMER A, DUNNE T and RADLOFF S (1993a) Macroinvertebrate functional feeding groups in the middle and lower reaches of the Buffalo River, eastern Cape, South Africa. II. Dietary variability. Freshwater Biol. 29 441-453. [ Links ]
PALMER C, O'KEEFFE J and PALMER A (1993b) Macroinvertebrate functional feeding groups in the middle and lower reaches of the Buffalo River, eastern Cape, South Africa. I. Functional morphology and behaviour. Freshwater Biol. 29 441-453. [ Links ]
PALMER RW and O'KEEFFE JH (1995) Distribution and abundance of blackflies (Diptera: Simuliidae) in relation to impoundments in the Buffalo River, Eastern Cape, South Africa. Freshwater Biol. 33 109-118. [ Links ]
PALMER CG, ROSSOUW N, MULLER WJ and SCHERMAN PA (2005) The development of water quality methods within ecological Reserve assessments, and link to environmental flows. Water SA 31 161-170. [ Links ]
PARKER CR and VOSHELL JR (1983) Production of filter-feeding Trichoptera in an impounded and a free-flowing river. Can. J. Zool. 61 70-87. [ Links ]
PETTS GE (1984) Impounded Rivers: Perspectives for Ecological Management. John Wiley and Sons Ltd, UK. [ Links ]
POWER ME, DIETRICH WE and FINLAY JC (1996) Dams and downstream aquatic biodiversity: potential food web consequences of hydrologic and geomorphic change. Environ. Manage. 20 887-895. [ Links ]
PRINGLE C (1997) Exploring how disturbance is transmitted upstream: going against the flow. J. N. Am. Benthol. Soc. 16 425-438. [ Links ]
PRINGLE CM, FREEMAN MC and FREEMAN BJ (2000) Regional effects of hydrologic alterations on riverine macrobiota in the New World: Tropical-temperate comparisons. Biosci. 50 807-823. [ Links ]
RICHTER BD, BAUMGARTNER JV, WIGINGTON R and BRAUN DP (1997) How much water does a river need? Freshwater Biol.37 231-249. [ Links ]
RHP (RIVER HEALTH PROGRAMME) (2006) Rivers Database. Department of Water Affairs and Forestry, Pretoria, South Africa. URL: http://www.dwaf.gov.za/iwqs/rhp/naehmp.htm (Accessed October to December 2006). [ Links ]
RIVERS-MOORE NA, DE MOOR FC, MORRIS C and O'KEEFFE J (2007) Effect of flow variability modification and hydraulics on invertebrate communities in the great fish river (Eastern Cape province, South Africa), with particular reference to critical hydraulic thresholds limiting larval densities of Simulium chutteri lewis (Diptera, Simuliidae). Rivers Res. Appl. 23 201-222. [ Links ]
ROUX DJ, NEL JL, ASHTON PJ, DEACON AR, DE MOOR FC, HARDWICK D, HILL L, KLEYNHANS CJ, MAREE GA, MOOLMAN J and SCHOLES RJ (2008) Designing protected areas to conserve riverine biodiversity: Lessons from a hypothetical redesign of the Kruger National Park. Biol. Cons. 141 100-117. [ Links ]
SAMWAYS MJ (1989) Farm dams as nature reserves for dragonflies (Odonata) at various altitudes in the Natal Drakensberg Mountains, South Africa. Biol. Cons. 48 181-187. [ Links ]
SCHAEL DM and KING JM (2005) Western Cape River and Catchment Signatures. WRC Report No 1303/1/05. Water Research Commission, Pretoria, South Africa. [ Links ]
SCHLOSSER IJ and EBEL KK (1989) Effects of flow regime and cyprinid predation on a headwater stream. Ecol. Monogr. 59 41-57. [ Links ]
STANFORD JA and WARD JV (1989) Serial discontinuities in a Rocky Mountain river. I. Distribution and abundance of Plecoptera. Regulated Rivers: Res. Manage. 3 169-175. [ Links ]
STANFORD JA and WARD JV (2001) Revisiting the serial discontinuity concept. Regulated Rivers: Res. Manage. 17 303-310. [ Links ]
STANLEY EH, LUEBKE MA, DOYLE MW and MARSHALL DW (2002) Short-term changes in channel form and macroinvertebrate communities following low-head dam removal. J. N. Am. Benthol. Soc. 21 172-187. [ Links ]
STATSOFT (2003) Statistica, Version 6.1. StatSoft Inc, Oklahoma, USA. [ Links ]
THIRION C (2007) Module E: Macroinvertebrate Response Assessment Index in River EcoClassification: Manual for EcoStatus Determination (Version 2). WRC Report No TT 332/08. Joint Water Research Commission and Department of Water Affairs and Forestry Report, Pretoria, South Africa. [ Links ]
US EPA (2000) Stressor Identification Guidance Document. US Environmental Protection Agency, Washington DC, USA. URL: http://cfpub.epa.gov/caddis/ (Accessed 6 May 2009) [ Links ]
UYS MC and O'KEEFFE JH (1997) Variability in the invertebrate fauna of an intermittent stream over three consecutive midsummer periods. S. Afr. J. Aquat. Sci. 23 55-79. [ Links ]
WORLD COMMISSION ON DAMS (2000) Dams and Development: A New Framework for Decision-making. Earthscan Publications Ltd, London and Sterling, VA. URL: http://www.dams.org/report/contents.htm (Accessed 6 May 2009) [ Links ]
Received 22 December 2008; accepted in revised form 23 February 2010.
* To whom all correspondence should be addressed.
+2746 622 4014; fax: +2746 622 9427;
e-mail: s.mantel@ru.ac.za


{kind=link}
{kind=link}
{kind=link}