










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkWater SA
versão On-line ISSN 1816-7950
versão impressa ISSN 0378-4738
Water SA vol.36 no.3 Pretoria Abr. 2010
Evaluation of oxidising disinfectants to control Vibrio biofilms in treated seawater used for fish processing
MNNN Shikongo-NambabiI; B KachigundaII; SN VenterIII,*
IDepartment of Food Science and Technology, University of Namibia, Windhoek, Namibia
IIDepartment of Crop Science, University of Namibia, Windhoek, Namibia
IIIDepartment of Microbiology and Plant Pathology, University of Pretoria, Pretoria 0002, South Africa
ABSTRACT
Marine fish-processing plants often use seawater during their operations. Chlorination and UV are commonly used for disinfection of this water but may not be effective in preventing biofilm formation within the water distribution network. These biofilms negatively impact water quality and could lead to contamination of fish products. During a recent study, Vibrio alginolyticus strains were detected on processed hake. The presence of most Vibrio spp. on fish products is of consumer safety concern and needs to be minimised. Water treatment strategies effective for seawater disinfection but with minimal negative effect on fish quality are required. In this study the effectiveness of chlorine, ozone and hydrogen peroxide (H2O2) in the inhibition of mature biofilms or biofilm formation in natural seawater was investigated. Two V. alginolyticus strains (V590 and V595) isolated from hake fish as well as the type strains of V. alginolyticus LMG 4409 and V. parahaemolyticus LMG 2850 were used. Chlorine was ineffective as experiments showed that strains V590, V595 and V. parahaemolyticus LMG 2850 could form biofilms even in the presence of 4 mg/ℓ of chlorine. When ozone was used, biofilm initiation and formation were completely inhibited for only 2 strains of V. alginoluticus, i.e. LMG 4409 and V590, at 1.6 mg/ℓ or 0.8 mg/ℓ ozone, respectively. Hydrogen peroxide performed the best of all the disinfectants evaluated in this study. Inhibition of biofilm formation was observed for all strains at 0.05% H2O2. The mature biofilms were more resistant to H2O2 but were all eliminated at 0.2% concentrations. This study indicated that H2O2 is the most effective biocide to prevent biofilm formation in seawater distribution networks and could potentially be used as an alternative or supplementary disinfectant of seawater in marine fish-processing plants.
Keywords: V. alginolyticus, V. parahaemolyticus, biofilms, H2O2, disinfection, seawater
Introduction
The use of seawater instead of freshwater during marine fish processing is an economical alternative, especially in arid countries such as Namibia which are often faced with severe freshwater shortages. The water is typically used for activities such as cooling of the product or washing of surfaces and apparatus in the plant. Seawater may, however, contain human pathogenic bacteria including Vibrio species such as V. parahaemolyticus, V. cholerae, V. vulnificus and V. alginolyticus (Thompson et al., 2004; Wekell et al., 1994). The water, therefore, requires treatment and disinfection before being used for the processing of fresh fish. Currently chlorination and UV treatment are mostly commonly used to disinfect the water.
A recent study at a fish-processing plant showed that the quality of hake deteriorated along the processing line (author's own observation). Vibrio species were isolated at the intermediate stages of processing. None of these species were isolated from the fish which had just been delivered to the factory for processing. From the study it was clear that Vibrio alginolyticus strains were introduced by the treated seawater used during processing. Indications were that the major source of these bacteria was not inefficient treatment of the raw water but the subsequent formation of biofilms in the distribution network in spite of the presence of residual chlorine. V. cholerae, (Faruque et al., 2006; Mueller et al., 2007), V. vulnificus (Joseph and Wright, 2004). V. alginolyticus (Kogure et al., 1998) and V. parahaemolyticus (Enos-Berlage et al., 2005) are well known to form biofilms.
The presence of Vibrio spp. in the seawater used during processing of the fish poses a potential health hazard to consumers and should be minimised and controlled. Chlorine, or chloramines, are typically used to control bacterial biofilms in freshwater distribution systems (DeQueiroz and Day, 2007; Momba et al., 2002), but many studies have described the ineffectiveness of chlorine in controlling biofilm formation (Chu et al., 2003; Momba et al., 1998). There is, however, little information in the literature on alternative disinfectant strategies to control biofilm formation in seawater systems. The type and level of disinfectant to be used are, however, restricted, as high concentrations of disinfectants have been shown to cause discolouration of the fish due to oxidation of the myoglobin (Kim et al., 2000).
The aim of this study was to evaluate 3 different oxidising disinfectants to control biofilm formation by selected Vibrio isolates in seawater distribution systems. For this purpose chlorine, ozone and hydrogen peroxide were tested at a range of concentrations. Improvement in the treatment, disinfection and microbiological quality of the seawater used for fish processing would help to ensure the safety of the final product and protect consumers.
Materials and methods
Sampling and analysis of treated seawater
Samples of seawater used in a fish-processing facility in Walvis Bay, Namibia, were collected to evaluate the current treatment system. Samples were taken from the raw water, after flocculation and chlorination, after UV treatment at 300 J/cm2 at 254 nm (UVPure, Cape Town) and at 2 points within the factory after distribution through the network. Thiosulphate (0.04% w/v) was added to all samples to neutralise the residual chlorine. Samples were refrigerated during transport and storage. After serial decimal dilutions samples were plated in duplicate onto nutrient agar (NA) (Oxoid), seawater agar (SWA) (Atlas, 2006) and thiosulphate citrate bile salts sucrose agar (TCBS) (Farmer III and Hickmann-Brenner, 1991). NA and TCBS plates were incubated at 37ºC while SWA plates were incubated at 22ºC.
The studies to evaluate the ability of Vibrio strains to form biofilms and to determine the effect of selected disinfectants on the inhibition and control of biofilm formation were conducted using either natural or artificial seawater. The artificial seawater (ASW) consisted of water in which 0.4 M NaCl, 0.1 M MgSO4·7H2O, 0.02 M KCl and 0.02 M CaCl2.·2 H2O had been dissolved (Farmer III and Hickman-Brenner, 1991).
Bacterial cultures and inoculum
Four bacterial isolates were included in this study. Two of the isolates were Vibrio alginolyticus strains previously isolated from the processing plant. Isolate V590 was isolated from hake after filleting by the Baader machine, and V595 was isolated from the finished product. These strains were identified using the API 20E (Biomérieux) test and amplification of the collagenase gene according to the method of di Pinto et al. (2006). The type strains of V. alginolyticus (LMG 4409) and V. parahaemolyticus (LMG 2850) were also included for comparative purposes. Both these strains were initially isolated from food- poisoning incidents related to seafood.
Strains were cultured individually on seawater agar (SWA) by incubation overnight at 37ºC. This culture was then used to inoculate 250 mℓ sterile nutrient broth (NB) containing 3% NaCl and incubated unshaken at 37°C for 24 h. The cell suspension (75 mℓ) was harvested by centrifugation at 4 000 r/min for 30 min and washed twice with 0.85% (w/v) NaCl. The pellet was re-suspended in 15 mℓ 0.85% NaCl to obtain an inoculum containing ±107cfu/mℓ.
Biofilm formation in artificial seawater
The ability of the selected bacteria to form biofilms in seawater was evaluated using a Pedersen's device (Pedersen, 1982). The device contained 20 clean microscope glass slides (7.6 x 2.6 cm) and was connected by means of silicone tubing via a peristaltic pump to a reservoir containing artificial seawater. Sterile artificial seawater (ASW) instead of natural seawater was used for these experiments due to logistical constraints of transporting and storing large volumes of water. The ASW was supplemented with bacteriological peptone (Biolab) and nutrient broth (NB) (Biolab) at 1 g/ℓ each. The effluent was collected, disinfected and discarded. The system was disinfected by perfusing the systems for 24 h with tap water containing 2.5 g/ℓ residual chlorine at a rate of 500 mℓ/h. Afterwards, the chlorine was neutralised by allowing 12 ℓ of sterile tap water containing 1.0 mg/ℓ sodium thiosulphate to run through the systems for another 24 h. The system was then rinsed with 10 ℓ of sterile tap water. Sterility was assessed by plating 0.1 mℓ of the effluent onto TCBS agar and incubating the plates at 37ºC for 24 h.
At the start of each experiment ASW was allowed to flow through the Pedersen's device for 1 h. After inoculation with 5 mℓ of the cell suspension the flow of ASW was maintained at 500 mℓ/h. Slides were withdrawn at 24 h intervals up to 96 h. Each slide was washed in sterile water, transferred to a 100 mℓ screw-cap bottle containing 10 mℓ sterile 0.85% (w/v) NaCl solution supplemented with 0.01 mg/l sodium thiosulphate, and sonicated (Integral systems) for 5 min. The suspension was diluted, plated in duplicate on TCBS agar plates and incubated at 37ºC for 24 h, after which the culturable counts were determined. The degree of biofilm formation was calculated as the density of bacteria per cm2 (Momba et al., 2002).
Disinfectants
The disinfectant concentrations used during this study corresponded to levels previously indicated as suitable for food processing. Previous studies showed that H2O2 could be used at a higher concentration than the other 2 disinfectants without negatively affecting the product quality (Kim et al., 2000). Chlorine concentrations were prepared by adding varying amounts of commercial sodium hypochlorite solution (3.5% m/v) to 250 mℓ NSW to give final concentrations of 0.2, 0.4, 0.6, 1.0, 1.5, 2.0, 2.5, 3.0 or 4.0 mg/ℓ free residual chlorine. The chlorine concentrations were measured using N, N-diethyl-p-phenelenediamine (DPD) Tablets No1 (The Tintometer (Ltd), England) and a Lovibond colour comparator. Ozone was generated using the Ozone Air and Water System, (Bulkmatech, Cape Town) to give ozone concentrations of 0.4, 0.8, 1.0, 1.6 and 2.0 mg/ℓ. Ozone concentrations were measured by the indigo colorimetric method (Standard Methods, 1998). H2O2 concentrations were prepared by adding varying amounts of 35% H2O2 (Merck) to 250 mℓ NSW for final concentrations of 0.05, 0.08, 0.1, 0.2, 0.3, 0.4 and 0.5%.
Inhibition of biofilm formation
Chlorine, ozone or H2O2 solutions were prepared at the concentrations described above in screw-top bottles containing 250 mℓ sterile NSW. Seawater without any disinfectant added served as the control. Microscope slides were inserted in all the bottles. After inoculation with 1 mℓ of the test culture suspension, the bottles were kept at 22ºC. Slides were withdrawn after 72 h. All slides were washed in sterile ASW and examined for the presence of biofilms through culturable-count determination as described above.
Inhibition of mature biofilms
Chlorine, ozone or H2O2 were prepared at the concentrations described above in 250 mℓ sterile NSW. Seawater without any disinfectant added was used as a control. Four slides containing mature (72 h) monoculture biofilms of the selected strains were obtained from the Pedersen's device systems described above. The slides were rinsed in sterile ASW and then immersed into the bottles, swirled and allowed to stand at room temperature. A slide was withdrawn from each bottle after 1 h. Slides were examined for the presence of biofilms by determining the culturable counts using TSBC agar incubated at 37ºC for 24 h.
Statistical analysis
The experimental set-up was a randomised complete block design. The statistical analysis was conducted by ANOVA using Genstat Release 7.2. The mean, the least significant difference, and coefficient of variation were calculated to determine the significance in responses among treatments.
Results
Microbial quality of treated seawater
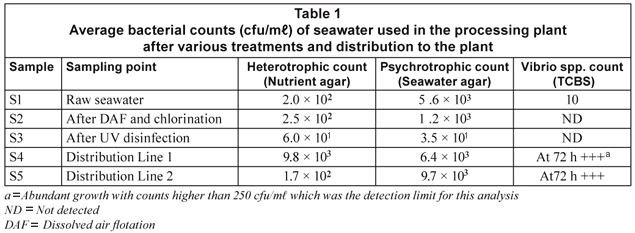
The aerobic heterotrophic, psychrotrophic and Vibrio culturable counts were determined for the treated seawater used in the Namibian fish-processing plant and are presented in Table 1. The heterotrophic and psychrotrophic counts for the untreated seawater (S1) were 2.0 x 102 (cfu/mℓ) and 5.6 x 103 (cfu/mℓ), respectively, and only 10 cfu/mℓ of presumptive Vibrio colonies was detected. Similar heterotrophic and psychrotrophic results were observed after dissolved air flotation ((DAF) and chlorination. At this point of the treatment no Vibrio ssp. were detected. The water leaving the plant after UV disinfection had very low numbers of both heterotrophic (60 cfu/mℓ) and psychrotrophic bacteria (35 cfu/mℓ). No Vibrio spp. were detected in this sample. Inside the processing plant the value of both the heterotrophic bacteria and the psychrotrophic bacteria had risen sharply to the similar levels detected in the raw water. Growth of Vibrio spp. above the detection limit of 250 cfu/mℓ was also observed on the TSBC plates after 72 h of incubation (Table 1).
Ability of Vibrio isolates to form biofilms in seawater
All Vibrio strains included in this study were able to form biofilms on glass slides in artificial seawater (ASW) supplemented with bacteriological peptone and nutrient broth (Fig. 1). The average bacterial densities in the biofilms peaked at between 107 and 109 cfu/cm2 after 72 h, changing only slightly over the next 24 h.

Effect of disinfectant on biofilm formation
The development of biofilms in the presence of different pre-determined concentrations of disinfectant was measured. After 72 h of exposure to the chlorine concentrations biofilm formation was observed for almost all of the strains (Fig. 2a). At the lower chlorine concentrations (1.5 mg/ℓ and below) there was no significant reduction in bacterial densities; however, there was a significant difference in the mean bacterial counts between the control without chlorine and the highest chlorine concentration (4 mg/ℓ) used (LSD=1.24) (Fig. 2a). The bacterial densities of strains V590, V595 and LMG 2850 dropped to log 3.73, 2.33 and 2.88, respectively, while V. alginolyticus type strain (LMG 4409) did not form biofilm at a chlorine concentration of 4 mg/ℓ.


Biofilm formation varied substantially between the different Vibrio strains (Fig. 2b) at the different concentrations of ozone used. There was a significant interraction between Vibrio strains and the disinfectant (p<0.001). For the V. alginoluticus isolates, strains V590 and LMG 4409, bioflim formation was completely inhibited at 0.8 mg/ℓ and 1.6 mg/ℓ, respectively. Biofilm formation was, however, still observed for V. parahaemolyticus (LMG 2850) and one of the V. alginolyticus fish isolates (V595) after 72 h of exposure to 2.0 mg/ℓ ozone (Fig. 2b). Statistically, the overall response of V595 was similar to that of V. parahaemolyticus LMG 2850. Their responses were, however, significantly different from that of V. alginolyticus LMG 4409 and V590. No formation of biofilms could be detected after 24 h at any of the H2O2 concentrations tested.
Effect of disinfectants on mature biofilms
Chlorine had a limited impact on the bacterial levels of all 3 V. alginolyticus biofilms after 1 h of exposure (Fig. 3a) when compared to the control value. Vibrio parahaemolyticus LMG 2850 was more sensitive to chlorine, with a significant reduction in bacterial numbers. After 1h of exposure to 4 mg/ℓ, the bacterial density dropped from 8.03 x 107 to 9.43 x 104 cfu/cm2 (Fig. 3a). At 4 mg/ℓ the reactions of the type strains, V. parahemolyticus LMG 2850 and V. alginolyticus LMG 4409, to chlorine were not significantly different, but they differed from those of the 2 fish isolates that were more resistant to chlorine. The ozone treatments resulted in a typical 1 log reduction in the bacterial levels, independent of the ozone concentration used (Fig. 3b) and these reductions were not statistically significant (p=0.166).



There was a significant reduction in bacterial counts of the mature biofilms of all the Vibrio isolates (Fig. 3c) as the H2O2 concentration increased (p<0.001). Of all the strains tested, V. parahaemolyticus strain LMG 2850 was the most sensitive and the mature biofilm could be inhibited after 1 h of exposure to 0.08% H2O2. The V. alginolyticus type strain (LMG 4409) biofilms were inhibited at 0.1% H2O2. The 2 V. alginolyticus stains isolated from the facility (V590 and V595) were more difficult to remove and were only inhibited after 1 h exposure to 0.2% H2O2 (Fig. 4c).
Discussion
For many marine fish-processing facilities the use of treated seawater during operations is a viable economic alternative to the use of freshwater. The microbial quality of the water should be well managed as it may have a negative impact on the quality and safety of the final product. During an investigation into the deterioration of the microbial quality of hake during processing, V. alginolyticus strains most likely introduced by the treated seawater used during processing were isolated (author's own observation). Although certain strains of V. alginolyticus have been shown to be pathogenic, it is their close relationship to V. parahaemolyticus, a pathogen widely associated with food-borne infections and outbreaks linked to seafood (Thompson et al., 2004) that is of even greater concern. The ability of these V. alginolyticus strains to survive and grow in treated seawater and the subsequent contamination of the hake may, therefore, be indicative of a similar behaviour of V. parahaemolyticus, should this pathogen be present. Control of this route of contamination is, therefore, of great importance for the efforts to minimise potential health hazards to consumers.
The first focus of the study was to survey the quality of the seawater used at the facility and to investigate whether it could serve as a source of contamination. Results showed that chlorination combined with UV irradiation drastically reduced both the mesophilic and psychrotrophic bacteria in seawater. Although the quality of the water directly after these treatments was very good, it deteriorated during distribution and some Vibrio spp. could be detected. It was clear from these data that the residual chlorine was ineffective in inhibiting biofilm formation in the distribution network. This was not unexpected as biofilm formation is common in water distribution networks (September et al., 2007) and detachment of the biomass can lead to deterioration of the microbial quality of the water.
Under a defined set of conditions of temperature, pH and a limited supply of nutrients, the 4 Vibrio isolates selected for this study were able to form monoculture biofilms on glass slides in the Pedersen device. This was not unexpected as biofilm formation by V. alginolyticus has previously been demonstrated by Kogure et al. (1998). The biofilm formation of the fish isolates (V595 and V590) was similar to that of the V. parahaemolyticus (LMG 2850) and V. alginolyticus (LMG 4409) type strains. These results support the hypothesis that the Vibrio species detected on hake fish could have originated from bacteria released from biofilms that formed in the water distribution network after the initial treatment. It also showed that there was little difference in the overall behaviour of the V. alginolyticus strains isolated from the facility and the V. parahaemolyticus type strain. All isolates were quite similar in terms of their ability to form biofilms and their resistance to specific disinfectants.
Chlorine is not very effective against biofilms formed by either atypical V. alginolyticus isolates or the V. parahaemolyticus type strain LMG 2850 in seawater. The effectiveness of chlorine against microorganisms in freshwater depends on a number of factors including the residual concentration, contact time, temperature, pH, and aggregation (Obi et al., 2008). Not much attention has, however, been given to the possible additional inhibitors of chlorine that might exist in water with high salt concentrations such as seawater. From the historical data kept at the factory it is clear that a residual chlorine concentration of 0.2 mg/ℓ was constantly maintained in the system. These conditions might have selected for strains with an ability to tolerate high chlorine concentrations as was previously demonstrated by Ridgeway and Olson (1982). From the responses of the V. parahaemolyticus type strain it can be deduced that this bacterium will behave similarly in the seawater distribution system and may, therefore, contaminate the final product whenever present.
Failure of chlorine to inhibit biofilm formation and mature biofilms was not due to the effect of pH. The pH of the NSW was 7.4, a level at which both HOCl and OCl- exist in various proportions (LeChevallier et al., 2004). In this study the CT (concentration x exposure time) value for the highest concentration used (4 mg/ℓ) was 288 after 72 h exposure, a value that was much higher than the 15 to 150 CT values recommended for drinking water (Debore and Von Gunten, 2008). The use of chlorine at concentrations higher than 4 mg/ ℓ was not considered, due to the potential effect that higher chlorine levels might have on costs, acceptability of the final product, corrosion in the plant and the potential to generate possible carcinogens (Gopal et al., 2007; Wang et al., 2007).
The ability of ozone to inhibit biofilm formation varied between the strains tested. Again V. alginolyticus V595 was the most resistant to ozone, and biofilm formation was not inhibited at 2 mg/ℓ ozone. Although biofilm formation could be inhibited for V alginolyticus V590 at an ozone concentrations of 0.8 mg/ℓ, none of the mature biofilms was inhibited at the highest concentrations of ozone used (2 mg/ℓ) in this study. Higher ozone concentrations to remove mature biofilms are, however, not recommended. Reports have shown that at high concentrations ozone reacts with organic matter in water, generating nutrients that could stimulate bacterial attachment to surfaces and formation of biofilms (Clark et al., 1994). Ozone is also not stable for long periods and may not provide the level of residual disinfectant required to inhibit existing biofilms (Khadre et al., 2001; Guzel-Seydim et al., 2004)
Hydrogen peroxide (H2O2) was very effective in inhibiting biofilm formation at a concentration of 0.05% (500 mg/ℓ) H2O2. Mature biofilms of all 4 strains tested could be killed at concentrations between 0.08% and 0.2% H2O2. This suggested that H2O2 at higher concentrations could be used to remove existing mature biofilms from seawater distribution systems during shock dose treatments. During this study H2O2was more effective in killing bacteria than during a study done by Kim et al. (2000) on channel catfish carried out in freshwater. The discrepancy found in H2O2 action between seawater and freshwater could be ascribed to different disinfection environments implying that mineral ions present in seawater are essential in maximising the action of H2O2 against bacteria. Pedahzur et al., (1995) found such a synergistic effect between silver ions and H2O2 when inactivating E. coli in phosphate buffer.
Part of the success of H2O2 was that it could be used at a higher concentration than the other 2 disinfectants without negatively affecting the product quality. Kim et al., (2000) investigated the effectiveness of 0.7% (7 000 mg/ℓ) H2O2 on reducing bacterial counts on catfish fillets and found no significant differences between controls and H2O2 treated fillets with regard to appearance, colour, and odour scores. The levels of H2O2 used by Kim et al. (2000) were 10 times higher than the concentrations used in this study.
Conclusion
This study has shown that although seawater could be an alternative source of water for marine fish-processing plants the treatment and the quality of the water needs to be carefully managed. The conditions in this seawater distribution network have selected for V. alginolyticus strains that can form biofilms in the presence of a residual chlorine concentration of 0.2 mg/ℓ. Once present in the system these bacteria may be released from the biofilm and would contaminate the fish during processing. The presence of V. alginolyticus on its own is not of great health concern as it is rarely associated with cases of diarrhoea or gastroenteritis. V. alginolyticus is, however, closely related to the common food-borne pathogen, V. parahaemolyticus, and may be predicative of the growth, behaviour and survival of this seafood pathogen in the water system. The current study has confirmed that this is the case as there was little difference between the behaviour of strains representing these 2 species in the experiments conducted. This finding also emphasised the need for control of biofilm growth in distribution systems even though it may not currently pose a significant health threat.
Evaluation of 3 oxidising disinfectants showed that chlorine and ozone are ineffective in preventing biofilm formation and in removing mature biofilms formed by Vibrio species in seawater at their permissible concentrations. The only disinfectant that showed some promise was H2O2. Bench-scale experiments indicated that it would be possible to control biofilm formation at a concentration of 0.05% (500 mg/ℓ) H2O2 and that existing biofilms could be removed by shock doses of 0.2%. This still needs to be investigated with larger-scale experiments run over a longer period of time. Results published by Kim et al., (2000) strongly support the notion that the proposed levels of H2O2 would not have any negative effect on the quality of the processed hake, but further studies would be required to confirm these conclusions.
Acknowledgements
We would like to thank the Ministry of Education of Namibia for funding this research, the Department of Food Science and Technology for making the research facilities available, Dr P Chimwamurombe for his valuable contribution in editing the manuscript, Mr Benjamin Vries, the technician at the workshop at Neudamm Campus, University of Namibia, for assisting in maintenance and repair of the research apparatus, and the fish-processing plant in Walvis Bay, Namibia for permitting us to use their facility and for providing us with information on water treatment and disinfection methods used.
References
ATLAS RM (2006) Handbook of Microbiological Media for the Examination of Food (2nd edn.). Taylor and Francis, Baco Raton, United States of America. 314 pp. [ Links ]
CHU C, LU C, LEE C and TSAI C (2003) Effects of chlorine on the growth of biofilm in water pipes. J. Environ. Sci. Health A. 38 1377-1388. [ Links ]
CLARK RM, LYKINS BW, BLOCK JC, WYMER LJ and REASORER DJ (1994) Water quality changes in simulated distribution systems. J. Water SRT-Aqua 43 236-277. [ Links ]
DEBORE M and VON GUNTEN U (2008) Reactions of chlorine with inorganic and organic compounds during water treatment − Kinetics and mechanisms: A critical review. Water Res. 42 13-51. [ Links ]
DEQUEIROZ GA and DAY DF (2007) Antimicrobial activity and effectiveness of a combination of sodium hypochlorite and hydrogen peroxide in killing and removing Pseudomonas aeruginosa biofilms from surfaces. J. Appl. Microbiol. 103 794-802. [ Links ]
DI PINTO A, CICCARESE G, FONTANAROSA M, TERIO V and TANTILLO G (2006) Detection of Vibrio alginolyticus and Vibrio parahaemolyticus in shellfish samples using collagenase-targeted multiplex-PCR. J. Food Safety 26 150-159. [ Links ]
ENOS-BERLAGE JL, GUVENER ZT, KEENAN C and McCARTER LL (2005) Genetic determinants of biofilm development of opaque and translucent Vibrio parahaemolyticus. Mol. Microbiol. 55 1160-1182. [ Links ]
FARMER III JJ and HICKMAN-BRENNER FW (1991) The genera Vibrio and Photobacterium. In: Ballows A, Strüper HG, Dworkin M, Harder W and Schleifer K-H (eds.) The Prokaryotes. Vol. III (2nd edn.) Springer-Verlag, Berlin, Germany. 2952-3011. [ Links ]
FARUQUE SM, BISWAS K, UDDE SMN, AHMAD QS, SACK DA, NAIR GB and MAKALANOS J (2006) Transmissibility of cholera: In vivo-formed biofilms and their relationship to infectivity and persistence in the environment. Proc. Natl. Acad. USA 103 6350-6355. [ Links ]
GOPAL K, TRIPATHY SS, BERSILLON JL and DUBEY SP (2007) Review. Chlorination byproducts, their toxicodynamics and removal from drinking water. J Hazard. Mater. 14 1-6. [ Links ]
GUZEL-SEYDIM ZB, GREENE AK and SEYDIM AC (2004) Use of ozone in the food industry. Lebensm-Wiss. U-Technol. 37 453-460. [ Links ]
JOSEPH LA and WRIGHT AC (2004) Expression of Vibrio vulnificus capsular polysaccharide inhibits biofilm formation. J. Bact. 186 889-893. [ Links ]
KIM TJ, SILVA JL, CHAMUL RS and CHEN TC (2000) Influence of ozone, hydrogen peroxide or salt on microbial profile, TBARS, and color of channel catfish fillets. J. Food Sci. 65 1210-1213. [ Links ]
KHADRE MA, YOUSEF AE and KIM J-G (2001) Microbiological aspects of ozone applications in food: A review. J. Food Sci. 66 1242-1252. [ Links ]
KOGURE K, IKEMOTO E and MORISAKI H (1998) Attachment of Vibrio alginolyticus to glass surfaces is dependent on swimming speed. J. Bact. 180 932-937. [ Links ]
LeCHEVALLIER MW (2004) Inactivation (disinfection) process In: LeChevallier MW and Kwok-Keuk A (eds.) Water Treatment and Pathogen Control: Process Efficiency in Achieving Safe Drinking Water. WHO, IWA, London. 41-65. [ Links ]
MOMBA MNB, CLOETE TE, VENTER SN and KFIR R (1998) Evaluation of the impact of disinfection processes on the formation of biofilms in potable surface water distribution systems. Water Sci. Technol. 38 (8-9) 283-289. [ Links ]
MOMBA MNB, NDALISO S and BINDA MA (2002) Inhibition of Biofilm Regrowth in Potable Water Systems. WRC Report No. 1023/1/02. Water Research Commission, Pretoria, South Africa. [ Links ]
MUELLER RS, McDOUGALD D, CUSUMANO D, SODHI N, KJELLEBERG S, AZAM F and BARTLETT DH (2007) Vibrio cholerae strains possess multiple strategies for abiotic and biotic surfaces colonization. J. Bact. 189 5348-5360. [ Links ]
OBI CL, IGUMBOR JO, MOMBA MNB and SAMIE A (2008) Interplay of factors involving chlorine dose, turbidity flow capacity and pH on microbial quality of drinking water in small water treatment plants. Water SA 34 565-572. [ Links ]
PEDAHZUR R, LEV O, FATTAL B and SHUVAL HI (1995) The interaction of silver ions and hydrogen peroxide in the inactivation of E. coli: a preliminary evaluation of a new long acting residual drinking water disinfectant. Water Sci. Technol. 31 123-129. [ Links ]
PEDERSEN K (1982) Methods of studying microbial biofilms in flowing water systems. Appl. Environ. Microbiol. 43 6-13. [ Links ]
RIDGEWAY HF and OLSON BH (1982) Chlorine resistance patterns of bacteria from two drinking water distribution systems. Appl. Environ. Microbiol. 44 972-987. [ Links ]
SEPTEMBER SM, ELS FA, VENTER SN and BROZEL VS (2007) Prevalence of bacterial pathogens in biofilms of drinking water distribution systems. J. Water Health 5 219-227. [ Links ]
STANDARD METHODS (1998) Standard Methods for the Examination of Water and Wastewater (20th edn.). Part 2000. American Public Health Association/American Water Works Association/Water Environment Federation, Washington, DC. [ Links ]
THOMPSON FL, IIDA T and SWINGS J (2004) Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev. 68 403-431. [ Links ]
WANG W, YE B, YANG L, LI Y and WANG Y (2007) Risk assessment on disinfection by-products of drinking water of different water sources and disinfection processes. Environ. Int. 33 219-225. [ Links ]
WEKELL MM, MANGER R, KOLBURN K, ADAMS A and HILL W (1994) Microbiological quality of seafoods: viruses, bacteria and parasites. In: Shahidi F and Botta RJ (eds.) Seafoods: Chemistry, Processing Technology and Quality. (Ch. 11). Blackie Academic & Professional, Glasgow, Scotland. [ Links ]
Received 28 October 2009; accepted in revised form 23 February 2010.
* To whom all correspondence should be addressed.
+2712 420 4100; fax: +2712 420 3266;
e-mail: fanus.venter@up.ac.za


{kind=link}