










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.93 n.2 Pretoria 2022
http://dx.doi.org/10.36303/JSAVA.480
ORIGINAL RESEARCH
The effects of feeding and transport length on the welfare of white rhinoceroses (Ceratotherium simum simum) during long-distance translocations: a preliminary study
M LeiberichI, II, †; F PohlinI, II, III, †; EH HooijbergI, IV; M HofmeyrV, VI; D CooperVII; M ReubenVIII; LCR MeyerI, II
ICentre for Veterinary Wildlife Research, Faculty of Veterinary Science, University of Pretoria, South Africa
IIDepartment of Paraclinical Sciences, Faculty ofVeterinary Science, University of Pretoria, South Africa
IIIResearch Institute of Wildlife Ecology, Department of Interdisciplinary Life Sciences, University of Veterinary Medicine Vienna, Austria
IVDepartment of Companion Animal Clinical Studies, Faculty of Veterinary Science, University of Pretoria, South Africa
VGreat Plains Conservation and Rhinos Without Borders, Botswana
VIRhino Recovery Fund/Wildlife Conservation Network and Oak Foundation, United Kingdom
VIIEzemvelo KZN Wildlife, South Africa
VIIIDepartment of Wildlife and National Parks, Botswana
ABSTRACT
Translocation is a valuable conservation tool, but poses significant risks for the transported rhinoceroses. Interventions reducing these risks are required to ensure positive welfare during transportation. The aim of this study was to evaluate the effect of journey duration and feeding during the transport of white rhinoceroses (Ceratotherium simum simum). A total of 32 animals were transported by road during two events, five days apart. Fifteen rhinoceroses in the first transport event (37.0 ± 2.4 hr duration) were not fed, while 17 rhinoceroses in the second event (32.2 ± 1.5 hr duration) were offered lucerne. Blood samples were collected at capture and after transport for the evaluation of changes in serum clinical chemistry analytes. The Wilcoxon rank-sum test was used to compare differences between the groups.
In all rhinoceroses, transport resulted in changes in serum electrolyte, metabolite and enzyme concentrations, indicating a loss in total body water, nutritional shifts, stress and fatigue. Fed rhinoceroses, transported over a shorter time, displayed greater changes in osmolality (p < 0.006), serum sodium and chloride concentrations (p = 0.005 and = 0.001, respectively) indicating a greater degree of total body water loss than non-fed rhinoceroses. Feeding and a shorter transport duration reduced, but did not prevent, nutritional challenges. A greater increase in the muscle enzymes CK and AST (p = 0.027 and = 0.001, respectively), indicated greater fatigue in non-fed rhinoceroses transported over a longer time. Further work to distinguish the effects of feeding and journey duration is required to better understand the role feeding may play in mitigating welfare challenges during rhinoceros translocation.
Keywords: fluid shift, negative energy balance, stress, transport, translocation, white rhinoceros, Ceratotherium simum simum, animal welfare
Introduction
Wildlife translocation has been defined as the "deliberate movement of organisms from one site for release in another" (IUCN/SSC 2012). Translocations are a useful tool to reestablish populations, to improve the conservation status of a species and to fight the loss of biodiversity (IUCN/SSC 2012). However, translocations of wildlife are a complex, costly and high-risk endeavour (Berger-Tal et al. 2020). The success rate of translocations is variable and chronic stress-related complications possibly lead to translocation failures, especially after release and during the establishment phase (Dickens et al. 2009). The mortality rates resulting from rhinoceros translocations in South Africa and Namibia are estimated at 5% (Miller et al. 2016). In 2018, several translocations made tragic headlines when 11 out of 11 black rhinoceroses transported in Kenya died, and four out of six rhinoceroses translocated from South Africa to Chad died within months of release into their new habitat (Modise & Read 2018; Save the Rhino 2018). These losses not only attracted great criticism, but posed a great cost to the conservation organisations involved.
Stress was blamed as the primary cause of the failure of these translocations. While an acute stress response can benefit the adaptation process, chronic stress occurs when the physiological stress response system is overwhelmed and persisting or repeated exposure to stressors leads to pathology (Dickens et al. 2010; Linklater et al. 2010; Teixeira et al. 2007). The transport of domestic animals, and even more so, wild animals, inevitably involves several stressors such as unfamiliar handling, surroundings, noise, the movement of the transport vehicle, the extended time standing, as well as the deprivation of food and water (Dembiec et al. 2004; Dickens et al. 2009; Dickens et al. 2010; Fazio & Ferlazzo 2003; Gelling et al. 2009; Grandin 1997; Leadon et al. 2008; Montes et al. 2004; Teixeira et al. 2007).
Rhinoceros transports are carried out according to the guidelines developed by the Convention on International Trade in Endangered Species for the non-air transport of wild animals and plants (CITES 2013), and recommendations for the transport of rhinoceroses (Emslie et al. 2009; Morkel & Kennedy-Benson 2007). Mostly, animals are not fed or offered water during translocations, as anecdotally rhinoceroses do not drink from an unfamiliar water source, and affixed watering troughs can lead to injuries (CITES 2013). Furthermore, it is believed that wild caught animals do not eat offered feed and the potential risk of colic, and other gut complications associated with the tranquilising and immobilising drugs together with the feeding, is feared (Bapodra et al. 2014). Little is known about the stress responses and the reduction of these during the transport phase of rhinoceros translocation. In a preceding study, overall challenges to animal welfare associated with long-distance rhinoceros translocation were assessed and dehydration, a negative energy balance, muscle damage and a stress-induced immunomodulation identified as the main stressors during transport (Pohlin et al. 2020).
This study was undertaken as a first step towards assessing interventions that could alleviate the main challenges to animal welfare during rhinoceros translocation. The aim of this research was to carry out a preliminary assessment of several interventions to manage stress and its pathophysiological consequences during the transport of free-ranging white rhinoceroses. We hypothesised that the rhinoceroses fed during translocation exhibit no or less physiological challenges to animal welfare than the rhinoceroses transported without being fed.
Material and methods
A total of 32 white rhinoceroses (Ceratotherium simum simum) were included in this study. The animals originated from a 340-hectare private game farm in the Free State in South Africa and comprised 24 adults and eight juveniles. Of the adults, six were males and 18 females. The natural vegetation at the private game farm consisted mainly of wild grasses such as Themeda triandra. As a dietary supplement, rhinoceroses also received a commercially available game feed (0.6 kg/rhinoceros/day; Game macro pack 32, EPOL, South Africa) and a home-made mixture of maize (2.2 kg/rhinoceros/day), lucerne (1.7 kg/rhinoceros/day), molasses (0.5 kg/rhinoceros/day), sunflower oil cake (0.6 kg/ rhinoceros/day) and wheat bran (0.6 kg/rhinoceros/day).
Rhinoceros translocations by road took place for conservation purposes in two separate events. Fifteen rhinoceroses (four juveniles and 11 adults, 11 females and four males) were translocated during the first, and 17 (four juveniles and 13 adults, 12 females and five males) during the second event, five days later. The animals were loaded in separate crates and transported on the back of trucks. Trucks and International Air Transport Association-approved rhinoceros crates were used in both translocations, which followed the practical guidelines for transport of live wild animals (CITES 2013) and rhinoceroses (Emslie et al. 2009; Morkel & Kennedy-Benson 2007). The University of Pretoria Animal Ethics Committee (V067-17) approved opportunistic sample collection from these animals.
Capture
Animals were captured in the early morning hours by darting from a helicopter with a combination of etorphine (Captivon®, Wildlife Pharmaceuticals, 9.8 mg/ml, 3-5 mg/adult or 0.1-1.5 mg/ juvenile), azaperone (Azaperone tartrate, Wildlife Pharmaceuticals, 50 mg/ml, 20-40 mg/adult or 0-10 mg/juvenile) and 5000 IU hyaluronidase (Hyalase®, Kyron Laboratories, Johannesburg, South Africa, adult only) delivered remotely using 2.0 ml darts (Pneu-dart, Inc.®, Williamsport, Pennsylvania, USA) with 63.5 mm barbed needles.
The animals became recumbent within five mins of darting and a blood sample was collected immediately from an auricular vein. If an animal tremored severely (n = 10), butorphanol (Butorphanol tartrate, Wildlife Pharmaceuticals, 50 mg/ml) at 2-5 times the etorphine dose in mg, was administered intravenously in order to mitigate the hypoxaemia associated with muscle tremors (Buss et al. 2018). Additionally, either 5 g vitamin C (n = 4; ascorbic acid, 500 mg/5ml, Fresenius Kabi, Bloemfontein, South Africa) or 500 mg vitamin E and 50 mg selenium (n = 4; vitamin E acetate 17 mg/ml, sodium selenite 1.67 mg/ml, Kyron, Midrand, South Africa) were given intramuscularly to some adult rhinoceroses in an attempt to support the animals' antioxidant defenses during transport (Lykkesfeldt & Svendsen 2007).
Loading and transport
To facilitate loading, in all rhinoceroses, the etorphine's immobilising effects were partially reversed by administering diprenorphine (M5050®, 12 mg/ml, Novartis, Midrand, South Africa, at 0.2-0.8 mg/adult or 0-0.1 mg/juvenile) intravenously. Each rhinoceros, including the juveniles, was loaded into its own transport crate, where adult animals received another 7.2-15 mg of intravenous diprenorphine to complete the reversal of etorphine. Thereafter, the tranquiliser zuclopenthixol acetate (Clopixol-Acuphase®, 50 mg/ml, H. Lundbeck Pty. Ltd., Randburg, South Africa) was administered intramuscularly via hand-injection (100-250/adult or 10-50 mg/juvenile). Transport and the recording of transport time commenced once all rhinoceroses had been captured and loaded. The time between the first and last rhino of the day being loaded for the first and second transport was 4 h 25 min and 3 h 28 min, respectively. The route, trucks, crates and drivers for the two transports were the same.
During transport, the vehicles stopped at 2-4 hour intervals to allow for additional intramuscular administration of azaperone and/or midazolam (Dazonil®, 50 mg/ml, Wildlife Pharmaceuticals) to restless animals as needed. All white rhinoceroses received at least one top-up dose of azaperone (80-120 mg/ adult or 10-80 mg/juvenile), which was combined with 2.5-15 mg midazolam in the juveniles. Sixteen adult and six juvenile white rhinoceroses required up to three additional top-up doses of azaperone, alone, or in combination with midazolam (10-20 mg/adult) during transport.
During the first translocation, food was not offered to the animals (group non-fed, n = 15). During the second translocation, five days later, all individuals (group fed, n = 17) were offered lucerne. The amount of hay was not weighed, but an estimated 4-6 kg were provided to each animal at the beginning and half-way through the second transport. Water was not provided to the animals, but rhinoceroses were doused with water during the heat of the day. An environmental logger (Kestrel DROP D2AG Livestock Heat Stress Monitor, KestrelMeters, Parker, CO, USA) recorded environmental temperature, relative humidity and heat stress index during both transports at 30-minute intervals.
Release
At the release site, all adult rhinoceroses were re-immobilised with etorphine (3.5-6 mg) and azaperone (20-40 mg) via hand-injection into the nuchal hump while standing in the transport-crate. Juvenile rhinoceroses received 0.5-2.5 mg etorphine and 5 mg midazolam intramuscularly via pole-syringe. Animals were released from their crates and manually restrained with ropes until they became recumbent. Subsequently, blood samples were collected from the immobilised rhinoceroses from the auricular veins. Naltrexone (Trexonil®, 50 mg/ml, Wildlife Pharmaceuticals), at 20 times the etorphine dose in mg, was administered intravenously to reverse the immobilisation and release the rhinoceroses into the national park.
Blood sample analysis
Blood from rhinoceroses was collected directly into serum and potassium oxalate/sodium fluoride (NaF) tubes (BD Vacutainer; Becton and Dickinson, Plymouth, UK), stored in a cooler box with ice packs, and centrifuged within 24 hours. Serum and plasma were aliquoted and stored at -20 °C for one month until they could be transferred to a -80 °C freezer until analysis.
Samples were analysed in the clinical pathology laboratory of the Onderstepoort Veterinary Academic Hospital (Pretoria, South Africa).
Serum clinical chemistry analysis was performed using a Cobas Integra 400 Plus automated biochemistry analyser (Roche Diagnostics Ltd., Rotkreuz, Switzerland) using commercially available kits. Measured analytes included: total serum protein (TSP), albumin, globulin, sodium, chloride, urea, creatinine, potassium, magnesium, phosphorus, total calcium, iron, total bilirubin, cholesterol, triglycerides, alkaline phosphatase (ALP), Y-glutamyl transferase (GGT), glutamate dehydrogenase (GLDH), aspartate aminotransferase (AST), creatine kinase (CK). Plasma glucose and lactate concentrations were measured by spectrophotometric methods with the same analyser. Serum cortisol concentration was assessed by a chemiluminescent immunoassay using the Immulite/Immulite 1000 Cortisol® following manufacturer's instructions (Siemens Healthcare, Erlangen, Germany).
Serum haptoglobin was determined by the haemoglobin-binding method using a commercial kit (PHASE Haptoglobin Assay, Tridelta Development Limited, Kildare, Ireland) with the Cobas Integra 400 Plus analyser. Concentrations of serum amyloid A (SAA) were determined by a solid-phase sandwich enzyme-linked immunoassay using a commercial kit (PHASE SAA Assay, Tridelta Development Limited), previously validated for use in black and white rhinoceroses (Hooijberg et al. 2017; Schook et al. 2015).
Plasma osmolality was calculated using the equine formula (Brownlow & Hutchins 1982): Osmolality (mOsm/kg) = 1.86* (Na+ + K+) + Glucose + Urea + 9 (with all variables in mmol/L).
Statistical analysis
Statistical tests were performed using R 3.6.1 for Windows (The R Foundation, Vienna, Austria). Descriptive tables and scatter plots were generated. Mean and standard deviations were calculated for each analyte and presented for descriptive purposes. Due to the small sample size, nonparametric analyses were used to compare differences in the concentration or activities of measured analytes
1) between capture and after transport within each group (fed and non-fed)
2) between non-fed and fed rhinoceroses after transport
by using the Wilcoxon rank-sum test. Differences in the change (delta δ) in concentration of measured analytes from capture to after transport were compared between non-fed and fed rhinoceroses by using the Wilcoxon rank sum test. A p-value < 0.05 was considered significant.
Results
All animals survived the capture and transport. Rhinoceroses offered lucerne during transport ate and finished their food. Age or sex distribution did not vary significantly between the groups. The total flying time of the helicopter to capture the 15 (non-fed, first transport) rhinoceroses was 174 minutes, resulting in an average of 11.6 minutes of flying time per rhinoceros. The 17 (fed, second transport) rhinoceroses were captured in 78 minutes, resulting in an average of 4.5 minutes of flying time per rhinoceros. The loading process went smoothly, only one animal initially struggling to get up. Transport started once the last rhinoceros of the group was loaded.
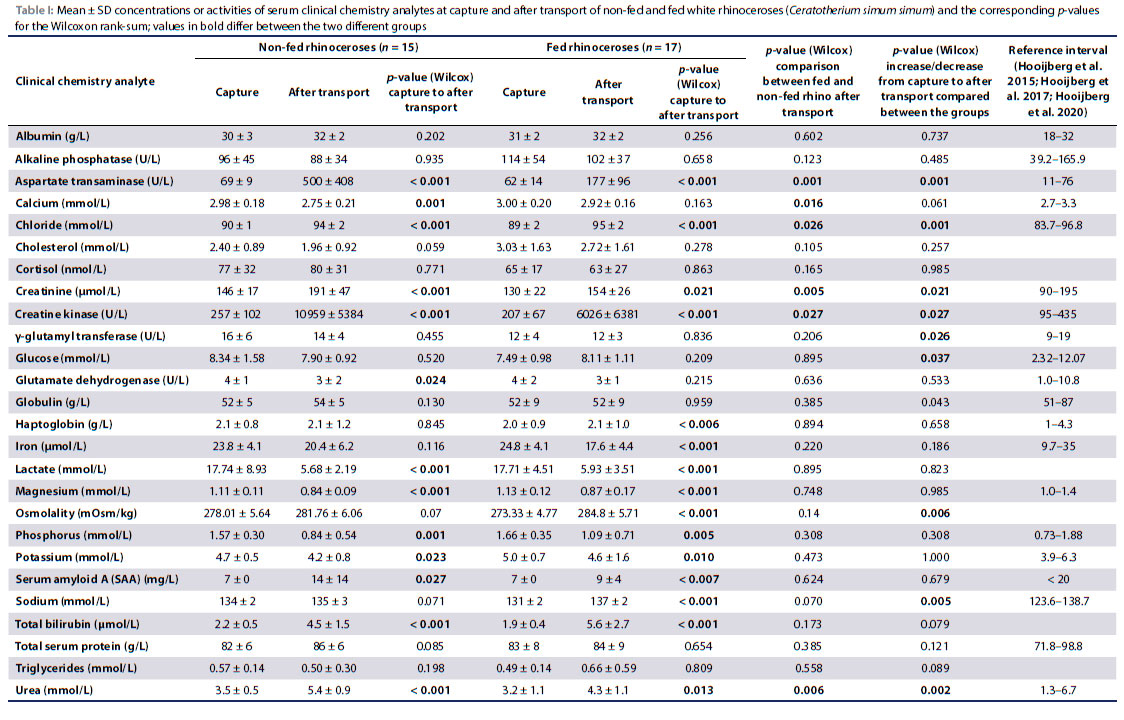
The mean transport time of the first (non-fed) and second (fed) rhinoceros translocation was 37.0 ± 2.4 and 32.2 ± 1.5 hours, respectively (p < 0.001). As the first transport was longer, animals were reinjected on average (mean ± SD) more frequently with a tranquiliser or sedative than during the second transport (2.8 ± 1.1 vs 2.4 ± 1.3 times). The mean ± SD environmental temperature during the first and second transport was 27.3 ± 7.8 °C and 26.8 ± 4.2 °C (p = 0.953), respectively. The mean ± SD relative humidity and heat stress indices during the first and second transport were 19.6 ± 9.8% and 30.2 ± 6.8% (p < 0.001) and 24.6 ± 7.4 and 25.3 ± 4.2 (p = 0.337), respectively. Means ± SD for measured analytes at capture and after transport and the corresponding p-values for the Wilcoxon rank-sum test are shown in Table I for the two different groups. Overall differences in serum analyte concentrations or activities from capture to after transport with respect to the different age-groups are published in Pohlin et al. (2020).
Briefly, both non-fed and fed rhinoceroses experienced an increase in urea (p < 0.001 and p = 0.013, respectively), creatinine (p < 0.001 and p = 0.021, respectively), SAA (p = 0.027 and p < 0.007, respectively) and chloride (p < 0.001 both groups) and a decrease in potassium (p = 0.023 and 0.010, respectively), magnesium (p < 0.001 both groups), phosphorus (p = 0.001 and 0.005, respectively) and lactate (p < 0.001 both groups) from before to after transport. Total bilirubin, AST and CK increased from before to after transport in all rhinoceroses (p < 0.001 all variables, both groups).
Only in non-fed rhinoceroses did the total calcium concentration (p = 0.001) and GLDH activity (p = 0.024) decrease. Only in fed rhinoceroses did the sodium concentration and plasma osmolality increase (p < 0.001) and iron concentration decrease (p < 0.001).
Creatine kinase and AST concentrations between animals that received vitamin E/selenium or ascorbic acid and animals that did not receive any supplements did not differ (CK p = 0.699 and 0.752, respectively; AST p = 0.682 and 0.606, respectively). Similarly, there was no difference between the animals that received or did not receive butorphanol and the CK and AST concentrations (CK p = 0.984, AST p = 0.225).
Differences between fed and non-fed rhinoceroses after transport
Urea (p = 0.006) and creatinine (p = 0.005) concentrations were higher and chloride (p = 0.026) and calcium (p = 0.016) concentrations lower in non-fed compared to fed rhinoceroses at the time of release. Serum AST and CK activities were 3.7 times (p = 0.001) and 1.8 times (p = 0.027) higher in non-fed compared to fed study animals, respectively.
Increase/decrease in the concentrations or activities of the different analytes between fed and non-fed rhinoceroses
In non-fed rhinoceroses, urea (p = 0.002), creatinine (p = 0.021), AST (p = 0.001) and CK (p = 0.027) concentrations increased more from capture to after transport than in fed rhinoceroses. Sodium (p = 0.005) and chloride (p = 0.001) concentrations and osmolality (p = 0.006) increased more in fed rhinoceroses whereas serum GGT (p = 0.026) activities decreased more in non-fed rhinoceroses. Glucose (p = 0.037) concentrations decreased in non-fed rhinoceroses, but increased in fed rhinoceroses.
Discussion
The long-duration transport led to changes in serum electrolytes, metabolites and enzymes in all rhinoceroses indicating a loss of total body water, a decreased nutritional intake, stress and fatigue which challenged animal welfare. In fed rhinoceroses that were transported over a shorter time compared to the non-fed rhinoceroses, the changes in serum electrolytes were greater, whereas the changes in metabolite concentrations and enzyme activities were minimal.
The increase in serum chloride, urea and creatinine concentrations in all rhinoceroses likely indicated a decrease in total body water and associated physiological reduction of the renal glomerular filtration rate during transport (DiBartola 2012; Muñoz et al. 2010). In domestic animals, extended transport duration has been identified as a major factor contributing to water loss (Knowles et al. 1999). Interestingly, fed rhinoceroses, which were transported over a shorter duration, showed a greater increase in sodium and chloride concentrations and osmolality, indicating a greater degree of total body water loss. This finding is in agreement with a study in horses, which found that the ingestion of hay without available drinking water leads to a free water deficit, as well as increased sodium concentration, serum osmolarity and thirst (Sufit et al. 1985). The reason for this pathophysiology is the increase of intestinal osmotic pressure associated with food intake and subsequent shift of extracellular water into the gastrointestinal tract (Freeman 2021; Sufit et al. 1985). It is suspected that the type of feed provided during transport plays a significant role in the amount of water required by the gastrointestinal tract. Diets consisting of more digestible ingredients such as grain and hay mixes require less water for excretion and therefore a lower water intake than hay and other diets with high indigestible or high dry matter contents (Freeman 2021; Fonnesbeck 1968; Sufit et al. 1985). These factors and their effect on hydration remain to be investigated in rhinoceroses and are of interest for future studies. However, based on the results of this study, the rhinoceros' water needs are not only influenced by transportation itself, but also by food intake. Thus, maintenance water requirements need to be adjusted accordingly. If rhinoceroses are fed during transport, additional fluid administration should be strongly considered. All rhinoceroses, regardless of being fed or not, experienced a decrease in serum calcium, phosphorus, potassium and magnesium concentrations and an increase in total bilirubin concentrations; changes which are usually attributed to a decreased food intake (Muñoz et al. 2010). Changes similar to those observed in this study have been documented after periods of fasting in domestic animal species such as equids and cattle (Baetz 1976; Fisher et al. 1999; Fisher et al. 2009; Muñoz et al. 2010). Although within normal reference intervals (Mathebula et al. 2012; Hooijberg et al. 2017), calcium concentrations decreased and urea concentrations increased more in the non-fed rhinoceroses. An increased protein catabolism associated with food deprivation and lack of rest during the longer transport could explain these results and demonstrate a beneficial effect of providing food (Doyle 1989; Gounden et al. 2020). In line with these findings, plasma glucose concentrations slightly increased in fed and decreased in non-fed rhinoceroses, likely related to the ingestion of food during transport.
In the rhinoceroses of both groups, muscle damage was evident, based on the marked increases in AST and CK activities from capture to after transport, both exceeding the AST and CK reference intervals for white rhinoceroses (Hooijberg et al. 2017). However, the non-fed group experienced greater muscle damage as the AST values increased 3.8 times more than in fed rhinoceroses. CK values increased 1.8 times more in non-fed compared to fed rhinoceroses, rising to a serum activity greater than 10 000 U/L. In horses, CK activities above 10 000 U/L may be indicative of clinically relevant rhabdomyolysis (Volfinger et al. 1994). In wildlife, increased CK activities and myoglobin concentrations after high-stress situations are characteristic signs of exertion-induced rhabdomyolysis, also referred to as capture myopathy. Altered blood flow and the exhaustion of normal aerobic energy leads to the breakdown of skeletal muscle fibres and consequently the increase of blood CK and myoglobin (Breed et al. 2019; Paterson 2014). Capture myopathy is a metabolic disease prevalent in various wild animal species with significant morbidity and mortality, often related to chase, capture, prolonged restraint and transport of the animals (Breed et al. 2019; Paterson 2014). The occurrence of the disease remains unpredictable with significant interspecies and individual differences. However, the duration of transport as well as providing food during prolonged transport may influence the occurrence of exertional rhabdomyolysis (Paterson 2014) and, as apparent from this study, may be two of several factors influencing the extent of muscle damage. Other variables, such as the distance run during darting, extent of muscle trauma during injections, and bruising during braking, were not measured in this study but may also cause muscle damage. In agreement with these findings, creatinine concentrations increased more in non-fed compared to fed animals, but did not exceed the reference interval established for white rhinoceroses (Hooijberg et al. 2017). Creatinine is a product of muscle creatine catabolism (Hosten 1990) and an increase of this analyte may indicate muscle protein breakdown and even predict the development of nephrosis as a consequence of myoglobin release from severe myopathy (Breed et al. 2019; Keenan 1967; Paterson 2014; Valberg et al. 1989). The immobilisation and the long duration of transport, spent mostly standing, likely caused a prolonged increase in muscular contraction, particularly in the non-fed rhinoceroses, which were transported over a longer time. This muscular contraction can result in an increased compression of blood vessels reducing muscle perfusion, resulting in muscular hypoxia and ultimately muscle cell damage (Spraker 1993). A previous study in domestic animal species has shown that the extent of muscle exertion and damage seems to not only be associated with the length of the journey but also appears to be highly influenced by knocks and bruising during transport (Fisher et al. 2009). We propose that rhinoceroses may have maintained a better balance while eating and thus experienced less knocks and bruises. Further studies, under more controlled conditions, are required to confirm this assumption and distinguish the effects of transport duration from the effects of feeding on serum muscle enzyme concentrations. In horses, wildlife and other production animals, the type of vehicle motion and the orientation of the animal in relation to the direction of travel have been shown to influence muscle cell damage during transport (Grigor et al. 1998; Padalino & Raidal 2020; Stull 1997). These, and other factors, also remain to be investigated in rhinoceroses.
Because the translocations took place for conservation purposes, independently of this study, a limitation of the study is the lack of ability to standardise the transport times, the tranquilisers used during the transport, the administration of top-up tranquilisers or sedatives or randomise whether animals were fed or not. As already mentioned, non-fed rhinoceroses were captured and transported over a longer time, which could have contributed to the observed differences in muscle enzyme concentration as well as the more frequent repeated intramuscular application of tranquilisers or sedatives.
Further research, under more controlled conditions, is necessary to record the exact amount of food ingested by the individual rhinoceroses as well as the feeding times, and to further elucidate their effects on changes in the various serum analyte concentrations and activities in transported white rhinoceroses.
Despite these limitations, this study provides preliminary insights into the effects of journey duration and feeding during rhinoceros translocation and provides enough evidence to stimulate a dedicated study on this topic.
Furthermore, it shows that animals which are used to being fed supplementary food prior to translocation do eat during transport, a "luxury" that may not occur in wild animals that only eat natural vegetation, which is also important to investigate and might highlight the benefit of letting rhinoceroses adapt to bomas and feed prior to long-distance translocations.
Even though both fed and non-fed animals displayed signs of dehydration, negative energy balance and muscle damage, feeding appeared to have a beneficial effect on reducing the negative effects faced during long-distance translocations. However, feeding lucerne alone did not adequately address the nutritional challenges associated with prolonged transport. Furthermore, since feeding alone seems to exacerbate the loss of total body water in the rhinoceroses, additional fluid administration is advised.
Further research is crucially needed to establish the clinical significance of the observed changes, optimal conditions and interventions during long transport, which will improve animal health and welfare, thus allowing for quick recovery and easy adaptation of rhinoceroses into new environments. Interventions such as parenteral or rectal fluid administration and providing a balanced feed, or parenteral energy supplementation, should be explored to effectively and safely reduce the emergence of energy deficits, dehydration and muscle damage during long duration transportation. Importantly, the efficacy of these, or any other interventions, need to be scientifically assessed in order to develop and improve rhinoceros-specific transport recommendations.
Acknowledgements
We thank Johannes Eksteen and Thomas Lengate (Kroonstad Dierehospitaal), Comfort Nkgowe, Les Charlisle, Grant Tracy, Rob Jackson, and Caron Botes for helping with sample collection and storage. Our sincere thanks go to Rhinos Without Borders (Great Plains Conservation & Beyond) who organised the translocation and provided access to the rhinoceroses, the Botswana Government, the Department of Wildlife & National Parks Botswana, the Botswana Defence Force, and Rhino Conservation Botswana.
Conflict of interest
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Funding source
This work was supported by the International Rhino Foundation rhino research grant (grant number R-2019-3) and a donation from the South African Veterinary Association Wildlife Group Research Fund, as well as departmental funding.
Ethical approval
All procedures performed in studies involving animals followed all international, national, and institutional guidelines for the care and use of animals. The University of Pretoria Animal Ethics Committee (V067-17) approved opportunistic sample collection from the animals.
ORCID
M Leiberich: https://orcid.org/0000-0001-7381-2446
F Pohlin: https://orcid.org/0000-0002-4236-5847
EH Hooiiberq: https://orcid.org/0000-0002-4367-799X
M Hofmevr: https://orcid.org/0000-0003-4473-3671
D Cooper: https://orcid.org/0000-0002-4026-0240
LCR Meyer: https://orcid.org/0000-0002-5122-2469
References
Baetz, A.L., 1976, The effect of fasting on blood constituents in domestic animals, Ann Rech Vet 7(2), 105-108. [ Links ]
Bapodra, P., Cracknell, J., Wolfe, B.A., 2014, Comparison of butorphanol-detomidine versus butorphanol-azaperone for the standing sedation of captive greater one-horned rhinoceroses (Rhinoceros unicornis), J Zoo Wildl Med 45(1), 60-68. https://doi.org/10.1638/2013-0066R.1. [ Links ]
Berger-Tal, O., Blumstein, D.T., Swaisgood, R.R., 2020, Conservation translocations: a review of common difficulties and promising directions, Animal Conservation 23(2), 121-131. https://doi.org/10.1111/acv.12534. [ Links ]
Breed, D., Meyer, L.C.R., Steyl, J.C.A., et al., 2019, Conserving wildlife in a changing world: Understanding capture myopathy - A malignant outcome of stress during capture and translocation, Conserv Physiol 7(1), coz027. https://doi.org/10.1093/conphys/coz027. [ Links ]
Brownlow, M.A. & Hutchins, D.R., 1982, The concept of osmolality: Its use in the evaluation of "dehydration" in the horse', Equine Vet J 14(2), 106-110. https://doi.org/10.1111/j.2042-3306.1982.tb02358.x. [ Links ]
Buss, P., Miller, M., Fuller, A., et al., 2018, Postinduction butorphanol administration alters oxygen consumption to improve blood gases in etorphine-immobilized white rhinoceros, Vet Anaesth Analg 45(1), 57-67. https://doi.org/10.1016/j.vaa.2017.03.008. [ Links ]
CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora), 2013, Cites guidelines for the non-air transport of live wild animals and plants. Available from: https://cites.org/eng/resources/transport/index.php Accessed 14 Jun 2021. [ Links ]
Dembiec, D.P., Snider, R.J., Zanella, A.J., 2004, The effects of transport stress on tiger physiology and behavior, Zoo Biol 23(4), 335-346.https://doi.org/10.1002/zoo.20012. [ Links ]
DiBartola, S., 2012, Fluid, electrolyte, and acid-base disorders in small animal practice, 4th edn, St. Louis, Missouri: Elsevier Saunders. https://doi.org/10.1016/C2009-0-60133-1. [ Links ]
Dickens, M.J., Delehanty, D.J., Romero, L.M., 2010, Stress: an inevitable component of animal translocation, Biological Conservation 143(6), 1329-1341. https://doi.org/10.1016/j.biocon.2010.02.032. [ Links ]
Dickens, M.J., Delehanty, D.J., Romero, L.M., 2009, Stress and translocation: Alterations in the stress physiology of translocated birds, Proc Biol Sci 276(1664), 2051-2056. https://doi.org/10.1098/rspb.2008.1778. [ Links ]
Doyle, J., 1989, Calcium protein binding (CALCIUM), in Computer Programs in Clinical and Laboratory Medicine, 1st edn, Springer, New York, NY, pp. 138-141. https://doi.org/10.1007/978-1-4612-3576-7_31. [ Links ]
Emslie, R.H., Amin, R., Kock, R., (eds), 2009, Guidelines for the in situ re-introduction and translocation of African and Asian rhinoceros, 1st edn, IUCN, Gland, Switzerland. https://doi.org/10.2305/IUCN.CH.2009.SSC-OP.39.en. [ Links ]
Fazio, E. & Ferlazzo, A., 2003, Evaluation of stress during transport, Vet Res Commun 27(Suppl 1), 519-524. https://doi.org/10.1023/B:VERC.0000014211.87613.d9. [ Links ]
Fisher, A.D., Colditz, I.G., Lee, C., et al., 2009, The influence of land transport on animal welfare in extensive farming systems, Journal of Veterinary Behavior: Clinical Applications and Research 4(4), 157-162. https://doi.org/10.1016/j.jveb.2009.03.002. [ Links ]
Fisher, A.D., Matthews, L.R., Pearce, P.V., 1999, The effects of long haul transport on pregnant, non-lactating dairy cows, N Z Vet J 47(5), 161-166. https://doi.org/10.1080/00480169.1999.36136. [ Links ]
Fonnesbeck, P.V., 1968, Consumption and excretion of water by horses receiving all hay and hay-grain diets, J Anim Sci 27(5), 1350-1356. https://doi.org/10.2527/jas1968.2751350x. [ Links ]
Freeman, D.E., 2021, Effect of feed intake on water consumption in horses: Relevance to maintenance fluid therapy, Front Vet Sci 8, 626081. https://doi.org/10.3389/fvets.2021.626081. [ Links ]
Gelling, M., McLaren, G.W., Mathews, F., et al., 2009, Impact of trapping and handling on Leukocyte Coping Capacity in bank voles (Clethrionomys glareolus) and wood mice (Apodemussylvaticus), Animal Welfare 18(1), 1-7. [ Links ]
Gounden, V., Bhatt, H., Jialal, I., 2020, Renal function tests, in StatPearls. StatPearls Publishing. [ Links ]
Grandin, T., 1997, Assessment of stress during handling and transport, J Anim Sci 75(1), 249-257. https://doi.org/10.2527/1997.751249x. [ Links ]
Grigor, P.N., Goddard, P.J., Littlewood, C.A., 1998, The behavioural and physiological reactions of farmed red deer to transport: Effects of sex, group size, space allowance and vehicular motion, Appl Anim Behav Sci 56(2-4), 281-295. https://doi.org/10.1016/S0168-1591(97)00085-3. [ Links ]
Hooijberg, E., Du Preez, J.P., Steenkamp, G., et al., 2015, Haematology and serum biochemistry reference intervals for the white rhinoceros, Ceratotherium simum, in South Africa, ESVCP and ECVIM-CA Congress, Lisbon, Portugal, 10-12 September 2015. [ Links ]
Hooijberg, E.H., Steenkamp, G., Buss, P., et al., 2017, Method comparison and generation of plasma biochemistry RIs for the white rhinoceros on a point-of-care and wet chemistry analyzer, Vet Clin Pathol 46(2), 287-298. https://doi.org/10.1111/vcp.12490. [ Links ]
Hooijberg, E.H., Cray, C., Steenkamp, G., et al., 2020, Assessment of the acute phase response in healthy and injured southern white rhinoceros (Ceratotherium simum simum). Front Vet Sci 6, 475. https://doi.org/10.3389/fvets.2019.00475. [ Links ]
Hosten, A.O., 1990, BUN and creatinine, in Walker, H.K., Hall, W.D., and Hurst, J.W. (eds) Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd edn, Boston: Butterworths. Available from: http://www.ncbi.nlm.nih.gov/pubmed/21250147. [ Links ]
International Union for Conservation of Nature/Species Suvival Commission (IUCN/SSC), 2012, IUCN guidelines for reintroductions and other conservation translocations. Available from: https://www.iucn.org/content/guidelines-reintroductions-and-other-conservation-translocations. [ Links ]
Keenan, D.M., 1967, Changes of blood metabolites in horses after racing, with particular reference to uric acid, Aust Vet J 55(2), 54-57. https://doi.org/10.1111/j.1751-0813.1979.tb15159.x. [ Links ]
Knowles, T.G., Warriss, P.D., Braun, S.N., et al., 1999, Effects on cattle of transportation by road for up to 31 hours, Vet Rec 145, 575-582. https://doi.org/10.1136/vr.145.20.575 [ Links ]
Leadon, D., Waran, N., Herholz, C., et al., 2008, Veterinary management of horse transport, Vet Ital 44(1), 149-163. [ Links ]
Linklater, W.L., MacDonald, E.A., Flamand, J.R.B., et al., 2010, Declining and low fecal corticoids are associated with distress, not acclimation to stress, during the translocation of African rhinoceros, Animal Conservation 13(1), 104-111. https://doi.org/10.1111/j.1469-1795.2009.00308x [ Links ]
Lykkesfeldt, J. & Svendsen, O., 2007, Oxidants and antioxidants in disease: oxidative stress in farm animals. Vet J 173, 502-51 1. https://doi.org/10.1016/j.tvjl.2006.06.005. [ Links ]
Mathebula, N., Miller, M., Buss, P., et al., 2012, Biochemical values in free-ranging white rhinoceros (Ceratotherium simum) in Kruger National Park, South Africa, J Zoo Wildl Med 43(3), 530-538. https://doi.org/10.1638/2011-0259R.1. [ Links ]
Miller, M., Kruger, M., Kruger, M., et al., 2016, A scoring system to improve decision making and outcomes in the adaptation of recently captured white rhinoceroses (Ceratotherium Simum) to captivity, J Wildl Dis 52(2 Suppl), S78-S85. https://doi.org/10.7589/52.2S.S85. [ Links ]
Modise, A. & Read, F., 2018, The governments of the Republic of South Africa and Chad together with African Parks and SANParks confirm the discovery of two additional black rhino carcasses in Zakouma National Park in Chad. Available from: https://www.gov.za/speeches/governments-republic-south-africa-and-chad-together-african-parks-and-sanparks-confirm. [ Links ]
Montes, I., McLaren, G.W., McDonald, D.W., et al., 2004, The effect of transport stress on neutrophil activation in wild badgers (Meles meles), Animal Welfare 13(3), 355-359. [ Links ]
Morkel, P. & Kennedy-Benson, A., 2007, Translocating black rhino: current techniques for capture, transport, boma care, release and post release monitoring. Report to AfRSG pp. i-v, 1-85. [ Links ]
Muñoz, A. Riber, C., Trigo, P., et al., 2010, Hematology and clinical pathology data in chronically starved horses, J Equine Vet Sci 30(10), 581-589. https://doi.org/10.1016/j.jevs.2010.09.002. [ Links ]
Padalino, B. & Raidal, S.L., 2020, Effects of transport conditions on behavioural and physiological responses of horses, Animals (Basel) 10(1), 1-18. https://doi.org/10.3390/ani10010160. [ Links ]
Paterson, J., 2014, Capture myopathy, in West, G., Heard, D., & Caulkett, N. (eds) Zoo Animal and Wildlife Immobilization and Anesthesia, 2nd edn, Philadelphia: John Wiley & Sons, pp. 171-179. https://doi.org/10.1002/9781118792919.ch12. [ Links ]
Pohlin, F., Hofmeyr, M., Hooijberg, E.H., et al., 2020, Challenges to animal welfare associated with capture and long road transport in boma-adapted black (Diceros Bicornis) and semi-captive white (Ceratotherium Simum) rhinoceroses, J Wildl Dis 56(2), 294-305. https://doi.org/10.7589/2019-02-045. [ Links ]
Save the Rhino, 2018, All rhinos translocated to Tsavo East National Park have died. Available from: https://www.savetherhino.org/africa/kenya/all-rhinos-translocated-to-tsavo-east-national-park-have-died/. [ Links ]
Schook, M.W., Wildt, D.E., Raghanti M.A., et al., 2015, Increased inflammation and decreased insulin sensitivity indicate metabolic disturbances in zoo-managed compared to free-ranging black rhinoceros (Diceros bicornis), Gen Comp Endocrinol 217-218, 10-19. https://doi.org/10.1016/j.ygcen.2015.05.003. [ Links ]
Spraker, T., 1993, Stress and capture myopathy in artiodactylids, in Fowler, M. E. (ed) Zoo and wild animal medicine: Current therapy, 3rd edn, WB Sounders, pp. 481-488. [ Links ]
Stull, C., 1997, Physiology, balance and management of horses during transportation, in Horse Breeders and Owners Conference. [ Links ]
Sufit, E., Houpt, K.A., Sweeting, M., 1985, Physiological stimuli of thirst and drinking patterns in ponies, Equine Vet J 17(1), 12-16. https://doi.org/10.1111/j.2042-3306.1985.tb02028.x. [ Links ]
Teixeira, C.P., De Azevedo, C.S., Mendl, M., et al., 2007, Revisiting translocation and reintroduction programmes: the importance of considering stress, Animal Behaviour 73(1), 1-13. https://doi.org/10.1016/j.anbehav.2006.06.002. [ Links ]
Valberg, S., Gustavson, B.E., Lindholm, A., et al., 1989, Blood chemistry and skeletal muscle metabolic responses during and after different speeds and durations of trotting, Equine Vet J 21(2), 91-95. https://doi.org/10.1111/j.2042-3306.1989.tb02102.x. [ Links ]
Volfinger, L., Lassourd, V., Michaux, J.M., et al., 1994, Kinetic evaluation of muscle damage during exercise by calculation of amount of creatine kinase released, Am J Physiol 266(2 Pt 2), R434-441. https://doi.org/10.1152/ajpregu.1994.266.2.R434. [ Links ]
 Correspondence:
Correspondence:
M Leiberich
Email: marion.leiberich88@gmail.com
† These authors contributed equally to this work


{kind=link}