










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.90 n.1 Pretoria 2019
http://dx.doi.org/10.4102/jsava.v90i0.1747
ORIGINAL RESEARCH
Genetic diversity of Hepatozoon (Apicomplexa) from domestic cats in South Africa, with a global reassessment of Hepatozoon felis diversity
David J. HarrisI, II; Ali HalajianIII; Joana SantosI; Kgethedi M. RampediIV; Raquel XavierI
ICIBIO/InBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, Vairão, Portugal
IIDepartamento de Biologia, Faculdade de Ciências da Universidade do Porto, Porto, Portugal
IIIDepartment of Biodiversity, University of Limpopo, Sovenga, South Africa
IVAgricultural Research Council Animal Production Institute, Irene, South Africa
ABSTRACT
Genetic diversity within partial 18S rRNA sequences from Hepatozoon protozoan parasites from domestic cats in South Africa was assessed and compared against published data to assess global biogeographic patterns. Multiple distinct haplotypes of Hepatozoon felis were identified, as well as an unrelated Hepatozoon lineage. Hepatozoon felis genetic diversity globally is very high, indicating a likely complex of species. The recently described Hepatozoon apri from wild boars is closely related to some lineages of H. felis. Sarcocystis and Babesia parasites were also detected. Since Hepatozoon felis is apparently a species complex, potential differences between genetically distinct forms need to be assessed. The finding of an unrelated Hepatozoon indicates that felids can be infected by more species of Hepatozoon than currently known, and that trophic interactions may increase the number of Hepatozoon species found in carnivores. Genetic screening again is demonstrated to identify previously unrecognised parasites from vertebrate hosts.
Keywords: 18S rRNA; hepatozoonosis; Felis catus; Sarcocystis; phylogeny.
Introduction
Species of the genus Hepatozoon are apicomplexan parasites infecting most terrestrial vertebrate groups, having haematophagous arthropods as the definitive hosts. Recent phylogenetic assessments have shown a complex pattern of relationships, with the genus Karyolysus forming a clade within a paraphyletic Hepatozoon, although parasites from most mammalian carnivores form a well-defined genetic lineage (Maia et al. 2016). Clinical features of hepatozoonosis are widely reported in canids, but the situation in felids is less clear. Hepatozoon parasites have been identified in many different felids, either through identification of gamonts in blood smears or using DNA sequencing techniques. Reported hosts include lynx, cheetahs, lions, jaguar, tigers, wildcats and domestic cats (Baneth et al. 1998a). Hepatozoon parasites were first reported in a domestic cat from India over one hundred years ago, and until recently, only Hepatozoon felis was considered to occur in felids, with the exception of rare records of Hepatozoon canis, a common parasite of canids (Williams et al. 2014). Hepatozoon felis was recently recovered from ticks infesting humans in Turkey (Karasartova et al. 2018). Reported infection rates vary, for example, from only 0.57% of cats infected from a sample of 1229 from Israel (Baneth et al. 1998b) to 37.9% of 174 cats from Cyprus (Attipa et al. 2017). Several authors have noted 'H. felis-like' infections, with parasites showing genetic similarity to H. felis but some level of morphological or genetic distinction (Giannitti et al. 2012). Recently, a new species was described from the European wildcat, Hepatozoon silvestris (Hodžić et al. 2017:650-661), which was then also found in domestic cats in Italy (Giannelli et al. 2017). However, it is not clear how the different genetic lineages of Hepatozoon from felids are related, while sampling from many geographic regions is limited. This is essential information, because, in canids, different Hepatozoon species can have very different impacts on the health of the host and thus have a different veterinary importance (Vincent-Johnson et al. 1997). Similarly, a recent study identified subclinical infections with the haemoparasite Cytauxzoon felis in cats, suggesting the existence of different strains that may vary in pathogenicity, although this link was not identified with the ITS1 genetic loci (Pollard et al. 2017). Whether different genetic forms of Hepatozoon in felids have different pathological effects remains unknown, although a fatal infection with H. silvestris has recently been reported in a domestic cat (Kegler et al. 2018). Furthermore, 'H. felis-like' genetic lineages were recently identified in a genet, Genetta genetta (Harris et al. 2017), and a white-tailed mongoose, Ichneumia albicauda (Harris et al. 2018), both from South Africa, increasing the number of mammalian families known to be potentially infected. Adding to the already complex taxonomy, Hepatozoon apri was recently described from wild boars in Japan, with haplotypes appearing to be more closely related to H. felis than H. silvestris (Yamamoto et al. 2017). Similarly, Hepatozoon martis was recently described from martens, closely related to H. apri (Hodžić et al. 2018). Other recent studies have also shown that H. felis and H. silvestris may not be closely related (De Azevedo Gomes et al. 2017). Additionally, one study focusing on Hepatozoon from canids and rodents that included three haplotypes from H. felis showed that the species as currently constituted is not monophyletic (Maia et al. 2014).
Despite the recently identified distinct lineages and species, no studies have yet tried to determine the phylogenetic relations between all known H. felis haplotypes and those of closely related species. The aims of this study were to gather the first genetic data on Hepatozoon from domestic cats from South Africa and to decipher global patterns of diversity, by assessing the genetic variation within H. felis and H. felis-like parasites.
Research methods and design
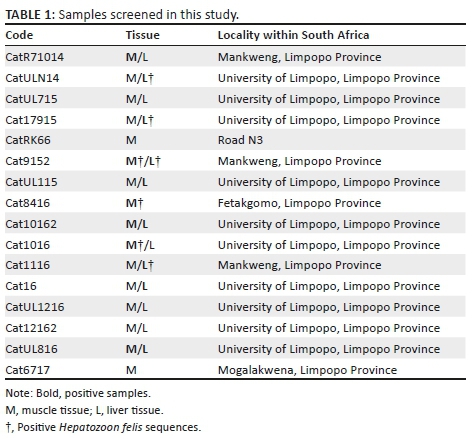
Sampling consisted of specimens either collected as road kills or from tissue banks from various collections in South Africa. Sixteen individuals were assessed, with muscle and/or liver tissues being tested (Table 1). The molecular approach followed standard procedures used in other screening studies, which have already shown that muscle tissue can be effectively used to extract DNA from apicomplexan parasites (Harris et al. 2012). DNA was extracted from different tissues using high salt procedures, which is as effective as commercial kits for extracting parasite DNA (Maia et al. 2014). Polymerase chain reaction (PCR) amplification of part of the 18S rRNA gene was performed using the primers HepF300 and HepR900 (Ujvari et al. 2004) with 35 cycles consisting of 95 ºC (45 seconds [s]), 60 ºC (45 s) and 72 ºC (90 s). Negative and positive controls were run with each reaction, and the products were sequenced by a commercial company (Genewiz, United Kingdom). Electropherograms were checked by eye, and compared against published sequences on GenBank using BLAST. All new sequences were submitted to GenBank (MK301457 to MK301462).
New sequences were aligned against published data from GenBank using the ClustalW software implemented in Geneious 4.8.5 (Biomatters Ltd). Sequences used are given in Supplementary Table 1. Phylogenetic relationships were estimated using Bayesian inference (BI) and maximum likelihood (ML). Models of sequence evolution (GTR+G, 1 partition) were selected by PartitionFinder 1.1.1 (Lanfear et al. 2012:1695-1701). Maximum likelihood estimates were performed using RaxML (Stamatakis 2014) with ten random replicates and supported nodes identified through 1000 nonparametric bootstrap iterations. Bayesian inference was implemented using MrBayes version 3.2.6 (Huelsenbeck & Ronquist 2001) and run for one million generations. After 25% burn-in, remaining trees were combined in a 50% majority rule consensus. Karyolysus paradoxa was used to root the tree. When few characters are available because of the slow evolving nature of the genes examined, networks can also be appropriate ways to represent genetic variation (Posada & Crandall 2001). We therefore estimated a phylogenetic network using the statistical parsimony approach implemented in TCS (Clement et al. 2000), both with a 95% probability criterion for connections and with a 20 connection step limit to visualise relationships of more divergent haplotypes. Although longer fragments were generated in this study, as many of the fragments available on GenBank were much shorter, a final alignment of 255 bps was used.
Results
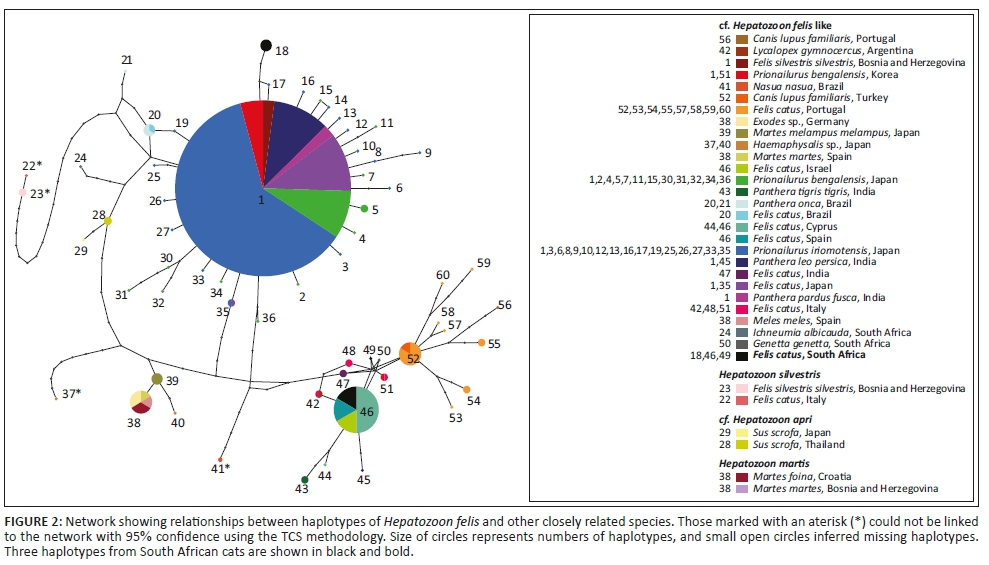
Of the 16 domestic cat samples examined, 11 gave positive amplifications. Of these, six individuals gave rise to three haplotypes of 'H. felis-like' sequences that were included in the estimate of phylogenetic relationships and network with other data from GenBank (Figures 1 and 2). The estimates of relationships derived from ML and BI were similar, so a single tree is presented with support value from both analyses (Figure 1). This shows that H. felis as currently constituted is not monophyletic. Furthermore, the three haplotypes recovered from cats from South Africa were not closely related. In the network (Figure 2), all 'H. felis-like' sequences and samples from wild boars cf. H. apri could be connected with 95% confidence. Hepatozoon silvestris haplotypes could only be included in the network with a higher connection step limit, together with Hepatozoon sp. from a coati, Nasua nasua, from Brazil, and a tick, Haemaphysalis longicornis, collected in Japan (Figure 1). Another domestic cat sample (CatUL115) was positive for Hepatozoon, but it was more similar to a distinct lineage typically identified in rodents and snake predators, with a 99% similarity to Hepatozoon identified from, among others, jerboa, Jaculus jaculus (Maia et al. 2014), or the snake, Psammophis elegans (Tomé et al. 2013). This unrelated Hepatozoon could not be included in the network with 95% confidence and is not represented in Figures 1 or 2, belonging to a lineage that has been proposed as being potentially a distinct genus (Karadjian et al. 2015). An additional sample (CatR71O14) gave a positive PCR amplification, but the resulting sequence was most similar to another family of haemoprotozoan parasites, Sarcocystidae. The most similar available species on GenBank was Sarcocystis nesbitti with 98% similarity. Three samples gave positive PCR amplifications, and the resulting sequences were most similar to parasites of a different apicomplexan order, Piroplasmida, and could be identified as members of the genus Babesia. However, the quality of the sequences was poor, and multiple species of Babesia are known to infect cats and other felids in South Africa (Bosman et al. 2007). Therefore, these samples could not be analysed further.
Discussion
Overall, the network and phylogenetic trees revealed extremely high diversity within forms typically assigned to H. felis. It seems likely therefore that this represents a species complex. Some tentative biogeographic patterns can be identified. One haplogroup, including haplotype one occurring in several different host species, along with haplotypes that differ from this by just one or two mutations, is mostly found in Japan, India or South America. Most haplotypes from Europe, including Spain, Portugal, Italy, Cyprus but also Israel, form another distinct haplogroup. However, there were exceptions, with, for example, wildcats from Bosnia and Herzegovina having parasites with the common Haplotype 1, while parasites from Indian lions and tigers are more related to the European haplogroup. Haplotypes 29 and 28 were Hepatozoon recovered from ticks from wild boars from Japan (Matsuo et al. 2016) and Thailand (Sumrandee et al. 2015), respectively, with the recently described H. apri also sharing Haplotype 28 (Hodžić et al. 2018), indicating that all the Hepatozoon sequenced from wild boar in this region may correspond to H. apri. On the other hand, no Hepatozoon were identified from wild boar blood samples collected from Spain (Gimenez et al. 2009).
Among samples from South Africa, haplotype diversity was very high. Their positions are dispersed throughout the network and fall within distinct clades within the phylogenetic trees. Therefore, it seems likely that if H. felis as currently constituted represents a species complex, then multiple forms will occur in South Africa. Infection rates also seem to be very high, with 7 out of 16 samples infected with Hepatozoon. Although feline hepatozoonosis has been increasingly reported worldwide over recent years, this is still one of the highest proportions reported (Attipa et al. 2017). Despite the small sample size, this warrants further assessment to determine if this high rate is an artefact of sample size or is truly indicative of high infection rates in South Africa. Hepatozoon infections are sometimes correlated with other infections, for example, Attipa et al. (2017) identified a significant correlation between infection with Hepatozoon and Leishmania in cats from Cyprus. Further testing for other feline infectious agents should also be carried out in South Africa to determine if their prevalence is also high. Hepatozoon canis infections can show seasonal periodicity (Murata et al. 1993), and this should also be assessed in the H. felis complex to determine its impact on reported infection rates. Additionally, identification of the invertebrate final hosts is essential to understand how the parasites spread through the populations.
The finding of another, unrelated type of Hepatozoon in domestic cats is unexpected and increases to four known forms recovered from cats (H. felis, H. silvestris, H. canis) and an unknown Hepatozoon sp. of the 'Bartozoon' lineage (Karadjian et al. 2015). Several studies have shown that trophic pathways can lead to infections, with, for example, the pale fox, Vulpes pallida, sometimes infected with Hepatozoon typical of desert rodents (Maia et al. 2014). The same could be occurring in domestic cats. Whether these are 'dead-end' infections, or if the parasites can continue their life cycle in these predator hosts requires further assessment. However, it demonstrates that studies of Hepatozoon should preferably sequence positive amplifications, rather than assuming that Hepatozoon in felids will correspond to H. felis or H. canis.
The amplification of a Sarcocystis species is not unexpected, as these primers have been shown to amplify these parasites before (Harris et al. 2012) and several species of Sarcocystis are known to use cats as definitive hosts (Levine & Tandros 1980). Although additional data are needed to determine to which species this haplotype might belong, the high similarity with S. nesbitti warrants further attention as this species can cause illness in humans, but is thought to use snakes as the definitive host and humans as occasional intermediate hosts (Lau et al. 2014). It is therefore particularly important, both from a veterinary aspect and for human health assessments, to investigate how the Sarcocystis from a domestic cat may be related to S. nesbitti.
To conclude, H. felis as currently constituted includes highly distinct genetic lineages, and a detailed assessment with additional and more variable genetic markers is needed to determine how many actual species exist under this name. In the meantime, these lineages can be referred to as the H. felis-like complex. Relationships between these H. silvestris, H. martis and H. apri also need to be elucidated. Domestic cats, as well as being infected by three currently described species of Hepatozoon, H. felis, H. silvestris and H. canis, can also be infected by an unrelated lineage typically observed in rodents and reptiles that may be another example of infection via a trophic pathway. Therefore, the full list of Hepatozoon species that can be found in cats may still be incomplete. Similarly, the finding of an unknown Sarcocystis species from a domestic cat indicates that the list of these species infecting cats also needs clarification. Finally, while some general biogeographic patterns are apparent, many areas, including South Africa, can harbour highly distinct genetic lineages.
Acknowledgements
The authors thank Daniela Rosado and Daniel Oliveira for help with DNA extractions, and Sello Matjee for assisting A.H. with lab works.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
D.J.H. and A.H. designed the project. A.H. and K.M.R. supplied the samples. J.S. performed the laboratory procedures. J.S. and D.J.H. analysed the data. D.J.H., J.S. and R.X. drafted the manuscript with input from all authors. D.J.H. and R.X. arranged funding. All authors approved the final version of the manuscript.
Ethical considerations
This article followed all ethical standards for research without direct contact with human or animal subjects.
Funding information
D.J.H. and R.X. are supported by the Fundação para a Ciência e a Tecnologia (FCT, Portugal) through IF contracts IF/01627/2014 and IF/00359/2015.
Data availability statement
All new sequences are available on GenBank (MK301457 to MK301462).
Disclaimer
The views expressed in this article are those of the authors and do not necessarily reflect those of the funding organisations.
References
Attipa, C., Papasouliotis, K., Solano-Gallego, L., Baneth, G., Nachum-Biala, Y., Sarvani, E. et al., 2017, 'Prevalence study and risk factor analysis of selected bacterial, protozoal and viral, including vector-borne, pathogens in cats from Cyprus', Parasites & Vectors 10, 130. https://doi.org/10.1186/s13071-017-2063-2 [ Links ]
Baneth, G., Aroch, I., Tal, N. & Harrus, S., 1998a, 'Hepatozoon species infection in domestic cats: A retrospective study', Veterinary Parasitology 79(2), 123-133. [ Links ]
Baneth, G., Breitschwerdt, E.B., Hegarty, B.C., Pappalardo, B. & Ryan, J., 1998b, 'A survey of tick-borne bacteria and protozoa in naturally exposed dogs from Israel', Veterinary Parasitology 74(2-4), 133-142. [ Links ]
Bosman, A.M., Venter, E.H. & Penzhorn, B.L., 2007, 'Occurrence of Babesia felis and Babesia leo in various wild felid species and domestic cats in Southern Africa, based on reverse line blot analysis', Veterinary Parasitology 144(1-2), 33-38. https://doi.org/10.1016/j.vetpar.2006.09.025 [ Links ]
Clement, M., Posada, D.C.K.A. & Crandall, K.A., 2000, 'TCS: A computer program to estimate gene genealogies', Molecular Ecology 9(10), 1657-1659. [ Links ]
De Azevedo Gomes, L., Moraes, L.A., Aguiar, D.C.F., Dias, H.L.T., Ribeiro, A.S.S., Rocha, H.P.C. et al., 2017, 'Genetic diversity of Hepatozoon spp. in Hydrochoerus hydrochaeris and Pecari tajacu from Eastern Amazon', Ticks and Tick-borne Diseases 9(2), 314-318. [ Links ]
Giannelli, A., Latrofa, M.S., Nachum-Biala, Y., Hodžić, A., Greco, G., Attanasi, A. et al., 2017, 'Three different Hepatozoon species in domestic cats from southern Italy', Ticks and Tick-borne Diseases 8(5), 721-724. [ Links ]
Giannitti, F., Diab, S.S., Uzal, F.A., Fresneda, K., Rossi, D., Talmi-Frank, D. et al., 2012, 'Infection with a Hepatozoon sp. closely related to Hepatozoon felis in a wild Pampas gray fox (Lycalopex-Pseudalopex-gymnocercus) co-infected with canine distemper virus', Veterinary Parasitology 186(3-4), 497-502. https://doi.org/10.1016/j.vetpar.2011.11.006 [ Links ]
Gimenez, C., Casado, N., Criado-Fornelio, Á., De Miguel, F.A. & Dominguez-Peñafiel, G., 2009, 'A molecular survey of Piroplasmida and Hepatozoon isolated from domestic and wild animals in Burgos (northern Spain)', Veterinary Parasitology 162(1-2), 147-150. https://doi.org/10.1016/j.vetpar.2009.02.021 [ Links ]
Harris, D.J., Halajian, A., Santos, J.L., Swanepoel, L.H., Taylor, P.J. & Xavier, R., 2018, 'Diversity of haemoprotozoan parasites infecting the wildlife of Southern Africa', Folia Parasitologica 65(15), 015. https://doi.org/10.14411/fp.2018.015 [ Links ]
Harris, D.J., Maia, J.P.M.C. & Perera, A., 2012, 'Molecular survey of Apicomplexa in Podarcis wall lizards detects Hepatozoon, Sarcocystis, and Eimeria species', Journal of Parasitology 98(3), 592-597. https://doi.org/10.1645/JP-GE-2908.1 [ Links ]
Harris, D.J., Pereira, A., Halajian, A., Luus-Powell, W.J. & Kunutu, K.D., 2017, 'Screening for Hepatozoon parasites in gerbils and potential predators in South Africa', Journal of the South African Veterinary Association 88, 1-4. https://doi.org/10.4102/jsava.v88i0.1339 [ Links ]
Hodžić, A., Alić, A., Beck, R., Beck, A., Huber, D., Otranto, D. et al., 2018, 'Hepatozoon martis n. sp. (Adeleorina: Hepatozoidae): Morphological and pathological features of a Hepatozoon species infecting martens (family Mustelidae)', Ticks and Tick-borne Diseases 9(4), 912-920. https://doi.org/0.1016/j.ttbdis.2018.03.023 [ Links ]
Hodžić, A., Alić, A., Prašović, S., Otranto, D., Baneth, G. & Duscher, G.G., 2017, 'Hepatozoon silvestris sp. nov.: Morphological and molecular characterization of a new species of Hepatozoon (Adeleorina: Hepatozoidae) from the European wild cat (Felis silvestris silvestris)', Parasitology 144(5), 650-661. https://doi.org/10.1017/S0031182016002316 [ Links ]
Huelsenbeck, J.P. & Ronquist, F. 2001, 'MRBAYES: Bayesian inference of phylogenetic trees', Bioinformatics 17(8), 754-755. https://doi.org/10.1093/bioinformatics/17.8.754 [ Links ]
Karadjian, G., Chavatte, J.M. & Landau, I., 2015, 'Systematic revision of the adeleid haemogregarines, with creation of Bartazoon n.g., reassignment of Hepatozoon argantis Garnham, 1954 to Hemolivia, and molecular data on Hemolivia stellata', Parasite 22, 31. https://doi.org/10.1051/parasite/2015031 [ Links ]
Karasartova, D., Gurese, A.S., Gokce, T., Celebi, B., Yapar, D., Keskin, A. et al., 2018, 'Bacterial and protozoal pathogens found in ticks collected from humans in Corum province of Turkey', PLoS Neglected Tropical Diseases 12(4), e0006395. https://doi.org/10.1371/journal.pntd.0006395 [ Links ]
Kegler, K., Nufer, U., Alic, A., Posthaus, H., Olias, P. & Basso, W., 2018, 'Fatal infection with emerging apicomplexan parasite Hepatozoon silvestris in a domestic cat', Parasites & Vectors 11(1), 428. https://doi.org/10.1186/s13071-018-2992-4 [ Links ]
Lanfear, R., Calcott, B., Ho, S.Y. & Guindon, S., 2012, 'PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses', Molecular Biology and Evolution 29(6), 1695-1701. https://doi.org/10.1093/molbev/mss020 [ Links ]
Lau, Y.L., Chang, P.Y., Tan, C.T., Fong, M.Y., Mahmud, R. & Wong, K.T., 2014, 'Sarcocystis nesbitti infection in human skeletal muscle: Possible transmission from snakes', American Journal of Tropical Medicine and Hygiene 90(2), 361-364. https://doi.org/10.4269/ajtmh.12-0678 [ Links ]
Levine, N.D. & Tadros, W., 1980, 'Named species and hosts of Sarcocystis (Protozoa: Apicomplexa: Sarcocystidae)', Systematic Parasitology 2(1), 41-59. https://doi.org/10.1007/BF00015094 [ Links ]
Maia, J.P., Alvares, F., Boratyński, Z., Brito, J.C., Leite, J.V. & Harris, D.J., 2014, 'Molecular assessment of Hepatozoon (Apicomplexa: Adeleorina) infections in wild canids and rodents from North Africa, with implications for transmission dynamics across taxonomic groups', Journal of Wildlife Diseases 50(4), 837-848. https://doi.org/10.7589/2013-10-280 [ Links ]
Maia, J.P., Carranza, S. & Harris, D.J., 2016, 'Comments on the systematic revision of adeleid haemogregarines: Are more data needed?', Journal of Parasitology 102(5), 549-552. https://doi.org/10.1645/15-930 [ Links ]
Matsuo, K., Uetsu, H., Takashima, Y. & Abe, N., 2016, 'High occurrence of Sarcocystis infection in Sika Deer Cervus nippon centralis and Japanese wild boar Sus scrofa leucomystax and molecular characterization of Sarcocystis and Hepatozoon isolates from their muscles', Journal of Zoo and Wildlife Medicine 21(2), 35-40. https://doi.org/10.5686/jjzwm.21.35 [ Links ]
Murata, T., Shimoda, K., Inoue, M., Shiramizu, K., Kanoe, M., Taura, Y. et al., 1993, 'Seasonal periodical appearance of Hepatozoon canis gamont in the peripheral blood', Journal of Veterinary Medical Science 55(5), 877-879. [ Links ]
Pollard, D.A., Reichard, M.V., Cohn, L.A., James, A.M. & Holman, P.J., 2017, 'Genetic variability of cloned Cytauxzoon felis ribosomal RNA ITS1 and ITS2 genomic regions from domestic cats with varied clinical outcomes from five states', Veterinary Parasitology 244, 136-143. https://doi.org/10.1016/j.vetpar.2017.08.002 [ Links ]
Posada, D. & Crandall, K.A., 2001, 'Intraspecific gene genealogies: Trees grafting into networks', Trends in Ecology & Evolution 16(1), 37-45. [ Links ]
Sumrandee, C., Baimai, V., Trinachartvanit, W. & Ahantarig, A., 2015, 'Hepatozoon and Theileria species detected in ticks collected from mammals and snakes in Thailand', Ticks and Tick-borne Diseases 6(3), 309-315. https://doi.org/10.1016/j.ttbdis.2015.02.003 [ Links ]
Stamatakis, A., 2014, 'RaxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies', Bioinformatics 30(9), 1312-1313. https://doi.org/10.1093/bioinformatics/btu033 [ Links ]
Tomé, B., Maia, J.P. & Harris, D.J., 2013, 'Molecular assessment of apicomplexan parasites in the snake Psammophis from North Africa: Do multiple parasite lineages reflect the final vertebrate host diet?', Journal of Wildlife Diseases 99(5), 883-887. https://doi.org/10.1645/12-95.1 [ Links ]
Ujvari, B., Madsen, T. & Olsson, M., 2004, 'High prevalence of Hepatozoon spp. (Apicomplexa, Hepatozoidae) infection in water pythons (Liasis fuscus) from tropical Australia', Journal of Parasitology 90(3), 670-672. [ Links ]
Vincent-Johnson, N.A., Macintire, D.K., Lindsay, D.S., Lenz, S.D., Baneth, G. & Blagburn, B.L., 1997, 'A new Hepatozoon species from dogs: Description of the causative agent of canine hepatozoonosis in North America', Journal of Parasitology 83(6), 1165-1172. [ Links ]
Williams, B.M., Berentsen, A., Shock, B.C., Teixiera, M., Dunbar, M.R., Becker, M.S. et al., 2014, 'Prevalence and diversity of Babesia, Hepatozoon, Ehrlichia, and Bartonella in wild and domestic carnivores from Zambia, Africa', Parasitology Research 1139(3), 911-918. https://doi.org/10.1007/s00436-013-3722-7 [ Links ]
Yamamoto, M., Tokiwa, T., Tobiume, M., Akamatsu, S., Matsuo, K., Moribe, J. et al., 2017, 'Hepatozoon apri n. sp. (Adeleorina: Hepatozoidae) from the Japanese wild boar Sus scrofa leucomystax (Mammalia: Cetartiodactyla)', International Journal of Parasitology: Parasites and Wildlife 6(3), 354-360. https://doi.org/10.1016/j.ijppaw.2017.11.001 [ Links ]
 Correspondence:
Correspondence:
David Harris
james@cibio.up.pt
Received: 10 Oct. 2018
Accepted: 16 Apr. 2019
Published: 20 June 2019


{kind=link}
{kind=link}