










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.85 n.1 Pretoria Jan. 2014
http://dx.doi.org/10.4102/jsava.v85i1.1125
ORIGINAL RESEARCH
A model for determining baseline morphometrics of skeletal myofibres
Maurice MarsI; Michael A. GregoryII
IDepartment of TeleHealth, University of KwaZulu-Natal, South Africa
IISchool of Health Sciences, University of KwaZulu-Natal, South Africa
ABSTRACT
The minimum diameter method of morphometry (MDM) is used to measure and detect changes in myofibre diameters (FD). The MDM is used to identify pathology in skeletal muscle. In such studies, an assumption is made that the mean FD in a particular muscle in both limbs is essentially the same. This study explored this premise to determine the accuracy of MDM as a means of morphometric analysis. Muscle biopsies were obtained from the left (G1) and right (G2) tibialis anterior of four vervet monkeys and from the massaged left (G3) and untreated right (G4) tibialis anterior of four animals. Wax sections were prepared for MDM and FD was measured. Three specimens were re-measured on four occasions. The mean FD of each biopsy from G1 and G2 limbs were compared and the number of measurements necessary to produce a meaningful result determined. Repeated measurement showed a difference of < 3.0% in FD means between the first and three subsequent measurements. There was no significant difference of FD means between G1 and G2, whilst the difference between G3 and G4 was 11.2%. When > 175 FD were measured, the difference from the final mean was less than 2.0%. These data show that, (1) FD data derived from a muscle in an untreated limb can be used as a control for experiment mediated changes of FD in the other, (2) MDM is a reliable means of measuring FD and (3) 150-175 FD are needed to provide a dependable result.
Introduction
Skeletal myofibres change in size as a consequence of load (exercise) and pathology. The single parameter of cross-fibre diameter has been used to characterise normal skeletal muscle (Mannion et al. 1997; Toft et al. 2003) and show the effect that various conditions, such as Duchenne's muscular dystrophy (Watkins & Cullen 1982), neuro-muscular diseases (Slavin et al. 1982), juvenile idiopathic arthritis (Lindehammar & Lindvall 2004), spinal cord injury (Scelsi 2001) and diabetes (Hegarty & Rosholt 1981; Hendriksen et al. 1992; Hepple, Ross & Rempfer 2004), have on the myofibres of a particular muscle. In experimental models, change in fibre size has been used to show the effect of exercise (Dimauro, Balnave & Shorey 1992; Fitts, Costill & Gardetto 1989), ageing (Hennessey et al. 2001; Hepple et al. 2004) and nutrition (McKiernan et al. 2004) on myofibres.
Following exercise, drug administration or physical treatment, fibre diameters in a treated limb have been compared with fibre diameters in the untreated limb, the latter data serving as a control. One-legged exercise has been used to show changes in muscle fibre type and diameter (Esbjornsson et al. 1993; Mourtzakis, Gonzalez-Alonso & Graham 2004). Varying degrees of oedema and necrosis in each of three myofibre types have been shown following tourniquet-mediated ischaemic reperfusion injury in vervet monkey tibialis anterior muscle (Gregory & Mars 1992a; Mars & Gregory 1991), an effect that can be ameliorated by pharmacological intervention (Gregory & Mars 1992b). The effect of physical treatment modalities such as deep transverse friction massage (Deane, Gregory & Mars 2002) and compressed air massage on muscle fibre diameters and morphology have been demonstrated (Gregory & Mars 2004).
In all of the above, the rationale has been to measure and use the cross-sectional diameters of fibres in a particular muscle of an untreated limb as a baseline control to quantify morphometric changes of myofibres in the same muscle in a treated limb. The morphometric data have also been correlated with the ultrastructural appearance of myofibres in both control and treated muscle (Gregory & Mars 2004). These studies make the assumption that muscle fibre diameters of any given muscle are essentially the same on both sides of the body prior to the intervention. Any change in size or distribution of myofibre diameters in the muscle of the 'experimental' limb has been deemed to be a consequence of a physical, pharmacological or pathological intervention.
This rationale holds only if, (1) the method of measurement is able to provide reproducible data and (2) the morphometrics and distribution of fibre diameters are in fact the same in both limbs of healthy, untreated animals. There are many studies that use the untreated limb as a control and indeed, the mean, range and distribution of fibre diameters in the left and right limb muscles of healthy individuals are probably not significantly different. However, to the knowledge of the authors, this premise has not been proven and, to date, no study could be found that confirmed that fibre diameters in a given muscle are approximately the same on both sides of the body.
In this study, a light microscope was used to obtain cross-sectional diameters of myofibres within 3 µm haematoxylin and eosin (H&E) stained sections of wax-embedded muscle biopsies using the least cross-fibre diameter method of Dubowitz (1985). The primary objective was to verify that the diameters of myofibres in vervet monkey tibialis anterior in one limb can be used as a control for those in the tibialis anterior of the other. The study also investigated the reproducibility of fibre measurement using the light microscope, digital image capture and Dubowitz's (1985) 'minimum diameter morphometric method' and aimed to establish the number of myofibres that must be counted to attain reproducible results.
Research method and design
Muscle biopsy and preparation
Prior to biopsy, anaesthesia and analgesia were achieved by an intramuscular injection of a combination of 50% ketamine and 50% xylazine, 10 mg/kg of monkey body weight. After biopsy, the eight monkeys selected for the study were observed during recovery from the anaesthetic and then returned to the holding facility.
The biopsy area was at the level of the mid-calf. The hair from ankle to knee was removed with a depilatory cream from the left and right limbs of each of the eight animals to enable observation of any inflammatory reaction and to facilitate treatment. The skin was incised longitudinally over the biopsy site, the fascia opened and wedge biopsies of approximately 1 cm3 were taken from the left and right tibialis anterior of each vervet monkey, at the same distance distal to the knee joint line and lateral to the tibial margin.
Biopsies were taken from the left (Group 1: 1L - 4L) and right (Group 2: 1R - 4R) legs of four animals for comparison of fibre diameters in normal limbs. Four animals underwent compressed air massage for 15 min using a 3 cm multipinhole head and unheated air at 100 kPa to the left tibialis anterior. Biopsies were taken within 10 min of the completion of treatment (Group 3: E1 - E4). To serve as controls, similar biopsies were taken from the right tibialis anterior of each animal within 10 min of completion of treatment (Group 4: C1 - C4).
To reduce biopsy-induced muscle 'supercontraction' artefact that may affect myofibre morphometrics, the entire biopsy was immediately immersed in Karnovsky's fixative (Karnovsky 1965) as per the method of Olmesdahl, Gregory and Cameron (1979). After 5 min immersion, the biopsy was examined with a stereomicroscope to determine the alignment of myofibres and prepared for light microscopy.
Light microscopic morphometry
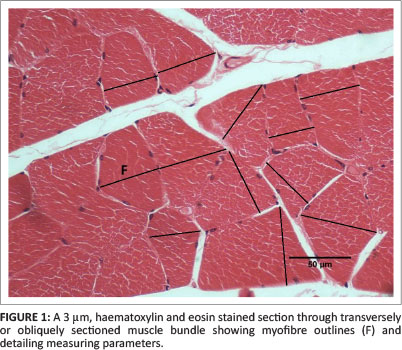
Each biopsy was re-immersed in Karnovsky's fluid for 24 h prior to dehydration and perfusion and embedding in wax. Care was taken to orientate the tissue to facilitate cross-sectioning of myofibres. Sections of 3 μm were cut of the wax-embedded tissue, mounted on glass slides and stained with H&E. The sections were examined using a Leitz Biomed light microscope (Leica Microsystems GmbH, Wetzler, Germany) with 10× objective. Images were collected from the microscope using a Panasonic video camera (WVCP414E) interfaced with a computer using calibrated Soft Imaging Systems (v. 2.1) software. Images displayed on a 35.56 cm (14-inch) monitor had a magnification of 350× and were captured as digital images in JPEG format on hard drive and compact disc. The entire surface area of each section was scanned and random areas captured for subsequent analyses. From five to seven images were captured from each section, with an average 35 fibres per digital image. The morphometric method employed was the 'minimum diameter method' described by Dubowitz and Brooke (1973). In brief, suitable areas containing transverse or obliquely sectioned myofibres were selected for morphometric evaluation (Figure 1). The myofibres were considered to be near cylinders and the diameter of fibres taken to be the minimum distance across each cell. A minimum of 179 and maximum of 240 fibres were measured from Groups 1 and 2 and 186-226 fibres were measured from Groups 3 and 4.
To determine reproducibility of measurement, the cross-sectional diameters of fibres (FD) in images from three specimens, C1, E1 and E4 were counted four times over a period of 1 year by the second author. The specimens were chosen on the basis of having a minimum of 175 fibres at first count.
To determine how many fibres should be counted from individual specimens to provide a reliable result, the random measurements from each biopsy were tabulated and mean FD calculated for the first 25, 50, 75, 100, 125, 150, 175 and 200 measurements. These means were then compared with each specimen's final mean fibre diameter and the mean fibre diameter of the group from which it came.
The mean, standard deviation (s.d.) and 95% confidence intervals (CI) were determined for the fibres in the left and right limbs of individual animals in each group. Comparison of the mean FD between the left and right limbs of the same animal and groups of animals was by unpaired t-test. When calculating the percentage differences in mean FD between individual animals and groups, the lesser of the two values was used as the denominator. One way analysis of variance (ANOVA) was used for comparison of the FD means after repeated measurements. Alpha was set at 5%.
Results
Light microscopy and morphometry
The wax-embedded muscle was well preserved and the fibres were generally transversely orientated. The 3 μm, H&E-stained sections enabled fibre outlines to be clearly discerned using a 10× objective (Figure 1). The morphometric data of fibres in individual specimens obtained from the left and right tibialis anterior muscle in Groups 1 and 2 are shown in Table 1, whilst Groups 3 and 4 are shown in Table 2. In Group 1, the mean FD (n = 851) was 49.4 μm ± 14.1 μm (95% CI: 48.5 μm - 50.4 μm) and ranged from 19.1 μm to 98.7 μm. In Group 2, the mean FD (n = 812) was 49.3 μm ± 13.5 μm (95% CI: 48.4 μm - 50.3 μm) and ranged from 19.6 μm to 97.3 μm. The difference between the mean FD in left and right legs ranged from 2.8% to 3.9%, with an overall left-right mean group difference of 0.2%. The differences in FD between the left and right limbs of each individual animal and the mean fibre diameter for the groups were not significantly different.
In the animals undergoing compressed air massage to the left limb (Group 3), the mean FD (n = 754) was 53.6 μm ± 14.3 μm (95% CI: 52.1 μm - 54.6 μm), with FD ranging from 19.4 μm to 101.0 μm. In the untreated control limb (Group 4), the mean FD (n = 756) was 48.2 μm ± 12.7 μm (95% CI: 47.3 μm - 49.1 μm), with FD ranging from 17.1 μm to 92.2 μm (Table 2). The difference between the mean FD in control and massaged limbs ranged from 3.3% to 18.6%, with an average difference between right (control) and massaged (left) limbs of 11.6%. The difference in mean fibre diameter between Groups 3 and 4 was significant (p < 0.0001). The differences in FD between the control and treated limbs were significantly different in three of the four animals.
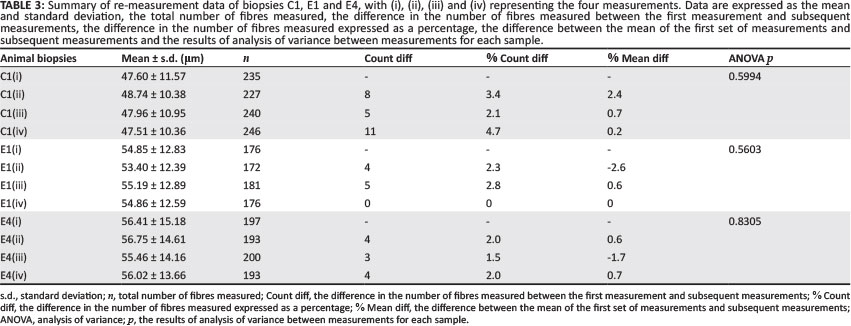
The results of the four repeated measurements of FD in specimens from animals C1, E1 and E4 are shown in Table 3. The maximum difference in the number of fibres counted for each animal was 19 in C1, 9 in E2 and 7 in E4. The number of fibres identified and measured from images of the three specimens over a 1-year period showed a maximum variation of 4.7%, 2.8% and 2.0% respectively from those identified at first count. The maximum difference between the mean of the first set of measurement and any other measurement was 2.4% in C1, -2.6% in E1 and -1.7% in E4. The maximum percentage difference in mean diameter between any of the four sets of means was 2.6% in C1, 3.3% in E1 and 2.0% in E4. The ANOVA showed no significant difference between the four sets of mean diameters for each animal.
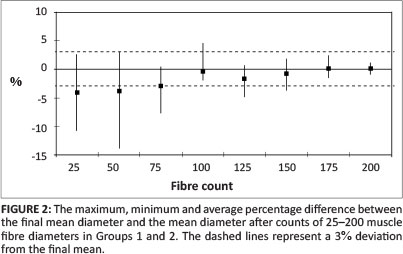
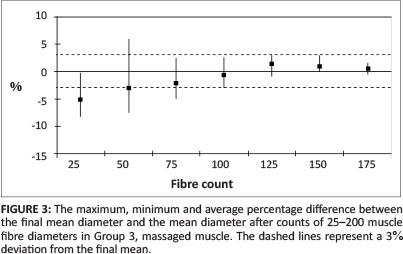
The fibre diameters for all animals in Groups 1 and 2 were measured. The mean fibre diameters after the first 25, 50, 75, 100, 125, 150, 175 and 200 fibres of each specimen were measured, recorded and compared with the final mean. The average percentage difference and the maximum and minimum percentage difference for each group of fibre diameters (25, 50, 75 etc.) are shown in Figure 2. The average percentage difference and the maximum and minimum percentage differences for Group 3 are shown in Figure 3.
Ethical approval
The eight vervet monkeys were studied with the approval of the Ethics and Research Committee of the University of KwaZulu-Natal (permit no. H08/2005).
Discussion
Measurement of fibre diameter is a useful technique to demonstrate normal physiological adaptations and pathological changes in skeletal muscles. The measurement and quantification of myofibre diameters have been reported in a wide range of animal species, including rabbit (Hebling, Scabora & Esquisatto 2009), vervet monkey (Gregory & Mars 1992a, 1992b), sheep (Peinado et al. 2004), rat (Aughsteen, Billah Khair & Suleiman 2006), dog (Kurzon et al. 1989) and even birds (Torrella et al. 1998). Abercrombie (1946) employed a rather complicated method of measuring fibre diameters by the measurement of fibre major and minor profile semi-axes. Recent advances in image processing and software detection of myofibre outlines enable measurement of myofibre diameters to be 'automated' (Garton et al. 2010).
In Garton et al.'s (2010) study, fibre diameters are calculated as a function of surface area, assuming the fibre to be a perfect circle. Whilst providing quick and generally useful results, muscle bundles from test muscle have to be equally cross-sectioned to provide comparative results. Slight oblique sectioning of one or other sample will inevitably cause surface area, and therefore calculated diameters, to differ, thus creating the possibility of comparative anomalies. Whilst it is almost impossible to measure as many fibres 'manually' as with an automated, computerised system, the 'simple', minimum diameter method first promulgated by Dubowitz and Brooke (1973) and later refined by Dubowitz (1985), is still one of the quickest and simplest methods used to determine changes in fibre diameter (Ferini-Strambi et al. 1998; Mannion et al. 1997; McKiernan et al. 2004). There are, however, some potential and inherent difficulties in applying the method that may cause some errors in the results. This study, therefore, had three objectives, (1) to determine whether myofibres in a particular muscle in a healthy untreated limb can be used as a morphometric control for those in a treated limb, (2) to establish the reproducibility of this method of morphometric analysis and (3) to determine how many myofibres should be counted to produce valid results.
In this study, the greatest difference between the mean myofibre diameters in the left and right limbs of individual animals in Groups 1 and 2 was 3.9%. There was no significant difference between the mean diameters of the myofibres in the left and right tibialis anterior of any animal. The difference between the combined data from left and right limbs was 0.2%. These findings strongly support the premise that fibre diameters of a particular muscle in one limb are similar to those in the other. In Groups 3 and 4, the difference between fibre diameters in control right limbs and those in treated left limbs ranged from 3.3% to 18.6%, with the difference between the combined data from left and right limbs being 11.6% (p < 0.0001). These results show that compressed air massage had a significant effect on fibre diameters and strongly suggest that the effect of induced trauma to one limb can be measured by comparing the fibre diameters of both limbs. This leaves the questions of how reliable or valid the minimum diameter method is as a means of measuring muscle fibre size and how many myofibres need to be measured to provide a valid result.
The reproducibility of the minimum diameter method measurements was assessed by re-measuring myofibres and counting the number of myofibres measured on four different occasions. For the most accurate results it is important to measure all discernable fibres in a particular field. In thin sections (< 4 μm), the myofibres in slightly oblique and/or cross-sectioned muscle bundles appear as independent polyhedral objects with from three to six sides separated from each other by a small interfibre space. It is important to identify the outline of the fibre to determine the minimum diameter of the polyhedron. In thicker or more oblique sections, fibre outlines become more difficult to discern, especially those of smaller cells. If some fibres are missed, or, because of preparation artefact or some other cause, some fibres are not measurable, what difference would this have on the mean of the sample?
The number of fibres identified in images of the three specimens measured on four separate occasions in 1 year showed a maximum variation of 4.7%, 2.8% and 2.0% respectively from those identified at first count. This had little effect on the mean diameter recorded. The maximum percentage differences between the mean diameters of the first set of measurements and subsequent measurements for the three samples were 2.3%, 2.7% and 1.6%. These differences were not statistically significant. These results suggest that errors in fibre identification have little effect on averaged morphometric data.
How many skeletal myofibres should be measured to provide a reliable result? Dubowitz (1985:85) suggests that 200 fibres, '… is ordinarily sufficient to give a reproducible and consistent value for the mean diameter and standard deviation', although no supporting data or references are given. The number of fibres counted is often a function of the size of the biopsy. Kurzon et al. (1989) measured a minimum of 20 myofibres of each fibre type using a digitiser tablet. In a study in which the average number of fibres counted per biopsy was 280, the range was 48-850 fibres (Mannion et al. 1997). Hennessey et al. (2001) counted 50 type 1 fibres and 50 type 2 fibres for comparison, accepting a lower limit of 25 fibres if 50 fibres were not obtainable, whereas Hepple et al. (2004) counted between 42 and 274 fibres on average in different regions of the soleus muscle of rats of different ages.
In Groups 1 and 2, all eight samples had 179 or more fibres that could be measured. The means of the first 25, 50, 75, 100, 125, 150 and 175 fibres measured showed an average deviation of -4.0% from the final mean, with a maximum deviation of -11.0% after 25 fibres were measured and -14.0% after 50 fibres were measured, reducing to an average of less than 1.0% after 150 measurements. In Group 3, maximum deviation from the final mean after 25 measurements was 8.4%, reducing to less than 1.0% after 150 measurements respectively. Whilst these represent maximal deviations from the final mean in individual specimens, studies undertaken by this group and earlier researchers generally compared the morphometric parameters of control and treated tissues as groups. When the means of the first 25, 50, 75, 100, 125, 150 and 175 measurements of fibres in each specimen were grouped and compared with the final group mean for both Group 1 and 2 specimens, a maximum variation of 3.3% was extant for all groups of 100 or more measurements. It should be noted that the percentage difference from the final mean has been calculated using positive values, irrespective of whether mean fibre diameter was greater or less than the final mean. When positive and negative values for each specimen were summed, in each group the difference from the final group mean was always less than 3.3% after 100 measurements. This suggests that individual and group measurements from an absolute minimum of 100 to an ideal 175 skeletal myofibres per specimen should provide valid data for comparison of fibre diameter between limbs.
Conclusion
Skeletal muscle is often used for a range of experimental purposes. It is important that the methodologies employed for these studies, including the morphometric evaluation of muscle biopsy, provide useful and accurate information. This study confirms that light microscopic examination of wax-embedded, thin (3 μm) sectioned, H&E-stained material enables skeletal myofibres to be both identified and measured. It also confirms that the minimum diameter method first proposed by Dubowitz in 1973 is adequately accurate and reproducible to morphometrically characterise normal and treated muscle. A minimum of 150 measurements per specimen should provide a reproducible, reliable result, especially if control and experimental specimens are to be grouped. Of particular importance for studies designed to detect change in skeletal myofibres, the results confirm that the morphometrics of myofibres in a particular muscle of one limb can be used as a control for experimentally mediated changes in the other limb of a particular subject. It is hoped that this methodology will be of use to identify and perhaps quantify muscle pathology or where exercise or other experimental procedures are to be studied.
Acknowledgements
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
M.M. (University of KwaZulu-Natal) jointly with M.A.G. (University of KwaZulu-Natal) conceived the project. M.M. took the muscle biopsies and massaged the limbs, helped with the literature search, did the final statistical analysis of the morphometric data, created the tables and helped in the writing of the manuscript. M.A.G. processed all the biopsied muscle, organised the sectioning and staining of the tissue and did all of the microscopy and morphometric measurements and provided rudimentary statistics of the collected data. He wrote the majority of the manuscript prior to submission to M.M. for comment and revision.
References
Abercrombie, M., 1946, 'Estimation of nuclear population from microtome sections', Anatomical Record 94, 239-247. http://dx.doi.org/10.1002/ar.1090940210 [ Links ]
Aughsteen, A.A., Billah Khair, A.M. & Suleiman, A., 2006, 'Quantitative morphometric study of the skeletal muscles of normal and streptozoticin-diabetic rats', Journal of the Pancreas 7, 382-389. [ Links ]
Deane, M.N., Gregory, M.A. & Mars, M., 2002, 'Histological and morphometric changes in untraumatised skeletal muscle treated with deep transverse friction', South African Medical Journal58, 28-35. [ Links ]
Dimauro, J., Balnave, R.J. & Shorey, C.D., 1992, 'Effects of anabolic steroids and high intensity exercise on rat skeletal muscle fibres and capilarization. A morphometric study', European Journal of Applied Physiology and Occupational Physiology 64, 204-212. http://dx.doi.org/10.1007/BF00626282 [ Links ]
Dubowitz, V., 1985, 'Normal muscle', in V. Dubowitz, C.A. Sewry & R.B. Fitzsimons (eds.), Muscle biopsy: A practical approach, pp. 82-128, Baillière Tindall, London. [ Links ]
Dubowitz, V. & Brooke, J., 1973, Muscle biopsy: A modern approach, W.B. Saunders, London. [ Links ]
Esbjornsson, M., Jansson, E., Sundberg, C.J., Sylven, C., Eiken, O. & Nygren, A., 1993, 'Muscle fibre types and enzyme activities after training with local leg ischaemia in man', Acta Physiologica Scandinavica 148, 233-241. http://dx.doi.org/10.1111/j.1748-1716.1993.tb09554.x [ Links ]
Ferini-Strambi, L., Smirne, S., Moz, U., Sferraza, B. & Iannaccone, S., 1998, 'Muscle fibre type and obstructive sleep apnea', Sleep Research Online 1, 24-27. [ Links ]
Fitts, R.H., Costill, D.L. & Gardetto, P.R., 1989, 'Effect of swim exercise training on human muscle fiber function', Journal of Applied Physiology 66, 465-475. [ Links ]
Garton, F., Seto, J.T., North, K.N. & Yang, N., 2010, 'Validation of an automated computational method for skeletal muscle fibre morphometry analysis', Neuromuscular Disorders 20, 540-547. http://dx.doi.org/10.1016/j.nmd.2010.06.012 [ Links ]
Gregory, M.A. & Mars, M., 1992a, 'Serial morphological changes in primate skeletal myofibres following 3 hours of ischaemia and 24 hours of reperfusion', South African Medical Journal 81, 473-478. [ Links ]
Gregory, M.A. & Mars, M., 1992b, 'Pharmacological manipulation of the ischaemic reperfusion injury: A histological, morphometric study', Communications of the Electron Microscopy Society of South Africa 21, 217-218. [ Links ]
Gregory, M.A. & Mars, M., 2004, 'The effect of compressed air massage on untraumatised rabbit skeletal muscle - A morphometric and ultrastructural study', South African Medical Journal 60, 19-27. [ Links ]
Hebling, A., Scabora, J.E. & Esquisatto, M.A.M., 2009, 'Muscle fibre types and connective tissue morphometry in frontal muscle of Norfolk rabbits (Oryctolagus cuniculus)', International Journal of Morphology 27, 187-191. http://dx.doi.org/10.4067/S0717-95022009000100032 [ Links ]
Hegarty, P.V. & Rosholt, M.N., 1981, 'Effects of streptozotocin-induced diabetes on the number and diameter of fibres in different skeletal muscles of the rat', Journal of Anatomy 133, 205-211. [ Links ]
Hendriksen, P.H., Oey, P.L., Van Veen, P.K., Wallinga-deJonge, W. & Veldman, H., 1992, 'Muscle conduction velocity and morphology after prolonged hypoxemia and diabetes in rats',Electromyography and Clinical Neurophysiology 32, 491-497. [ Links ]
Hennessey, J.V., Chromiak, J.A., Della Ventura, S., Reinert, S.E., Puhl, J. & Kiel, D.P., 2001, 'Growth hormone administration and exercise effects on muscle fiber type and diameter in moderately frail older people', Journal of the American Geriatrics Society 49, 852-858. http://dx.doi.org/10.1046/j.1532-5415.2001.49173.x [ Links ]
Hepple, R.T., Ross, K.D. & Rempfer, A.B., 2004, 'Fibre atrophy and hypertrophy in skeletal muscle of late middle aged Fischer 344x Brown Norway F1-hybrid rats', Journals of Gerontology, Series A: Biological Sciences and Medical Sciences 49, 108-117. http://dx.doi.org/10.1093/gerona/59.2.B108 [ Links ]
Karnovsky, M.J., 1965, 'A formaldehyde glutaraldehyde fixative of high osmolarity for use in electron microscopy', Journal of Cell Biology 27, 127-139. [ Links ]
Kurzon, W.M. Jr., Rosenblatt, J.D., Pynn, B.R., Marchetti, P.J., Plyley, M.J. & McKee, N.H., 1989, 'A comparative histochemical and morphometric study of canine skeletal muscle', Canadian Journal of Veterinary Research 53, 125-132. [ Links ]
Lindehammar, H. & Lindvall, B., 2004, 'Muscle involvement in juvenile idiopathic arthritis',Rheumatology 43(12), 546-1554. http://dx.doi.org/10.1093/rheumatology/keh381 [ Links ]
Mannion, A.F., Dumas, G.A., Cooper, R.G., Espinosa, M.W. & Stevenson, J.M., 1997, 'Muscle fibre size and type distribution in thoracic and lumbar regions of erector spinae in healthy subjects without back pain: normal values and sex differences', Journal of Anatomy 190, 505-513. http://dx.doi.org/10.1046/j.1469-7580.1997.19040505.x [ Links ]
Mars, M. & Gregory, M.A., 1991, 'A histometric analysis of skeletal myofibers following 90 min of tourniquet ischemia and reperfusion', Journal of Surgical Research 50, 191-195. http://dx.doi.org/10.1016/0022-4804(91)90246-I [ Links ]
McKiernan, S., Bua, E., McGorray, J. & Aitken, J., 2004, 'Early-onset calorie restriction conserves fiber number in aging rat skeletal muscle', The FASEB Journal 18, 580-581. [ Links ]
Mourtzakis, M., Gonzalez-Alonso, J. & Graham, T.E.S.B., 2004, 'Hemodynamics and O2 uptake during maximal knee extensor exercise in untrained and trained human quadriceps muscle: Effects of hyperoxia', Journal of Applied Physiology 97(5), 1796-1802. http://dx.doi.org/10.1152/japplphysiol.00169.2004 [ Links ]
Olmesdahl, P.J., Gregory, M.A. & Cameron, E.W.J., 1979, 'Ultrastructural artefacts in biopsied normal myocardial biopsy in man', Thorax 34, 82-90. http://dx.doi.org/10.1136/thx.34.1.82 [ Links ]
Peinado, B., Latorre, R., Vaquez-Aiton, J.M., Poto, A., Ramirez, G., Lopez-Albors, O. et al., 2004, 'Histochemical skeletal muscle fibre types in sheep', Anatomical Histology and Embryology 33, 236-243. http://dx.doi.org/10.1111/j.1439-0264.2004.00545.x [ Links ]
Scelsi, R., 2001, 'Our 20 year experience and results on skeletal muscle changes in paraplegics, related to functional rehabilitation', Basic and Applied Myology 11, 75-78. [ Links ]
Slavin, G., Sowter, C., Ward, P. & Paton, K., 1982, 'Measurement of striated muscle fibre diameters using interactive computer-aided microscopy', Journal of Clinical Pathology 35, 1268-1271. http://dx.doi.org/10.1136/jcp.35.11.1268 [ Links ]
Toft, I., Lindal, S., Bonaa, K.H. & Jenssen, T., 2003, 'Quantitative measurements of muscle fiber composition in a normal population', Muscle and Nerve 28(1), 101-108. http://dx.doi.org/10.1002/mus.10373 [ Links ]
Torrella, J.R., Fouces, V., Palomeque, J. & Viscor, G., 1998, 'Comparative skeletal muscle fibre morphometry among wild birds with different locomotor behaviour', Journal of Anatomy 192, 211-222. http://dx.doi.org/10.1046/j.1469-7580.1998.19220211.x [ Links ]
Watkins, S.C. & Cullen, M.J., 1982, 'Muscle fibre size and shape in Duchenne muscular dystrophy',Neuropathology and Applied Neurobiology 8, 11-17. http://dx.doi.org/10.1111/j.1365-2990.1982.tb00253.x [ Links ]
 Correspondence:
Correspondence:
Maurice Mars
Private Bag X7
Congella 4013
South Africa
Email: mars@ukzn.ac.za
Received: 18 Oct. 2013
Accepted: 26 Mar. 2014
Published: 14 Nov. 2014


{kind=link}
{kind=link}
{kind=link}