










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.85 n.1 Pretoria Jan. 2014
ORIGINAL RESEARCH
Assessment of selected biochemical parameters and humoral immune response of Nile crocodiles (Crocodylus niloticus) experimentally infected with Trichinella zimbabwensis
Louis J. La GrangeI, II; Samson MukaratirwaII
IDepartment of Agriculture, Rural Development and Land Administration, Mpumalanga, South Africa
IISchool of Life Sciences, University of KwaZulu-Natal, Westville Campus, South Africa
ABSTRACT
Fifteen crocodiles were randomly divided into three groups of five animals. They represented high-infection, medium-infection and low-infection groups of 642 larvae/kg, 414 larvae/kg and 134 larvae/kg bodyweight, respectively. The parameters assessed were blood glucose, creatine phosphokinase (CPK), lactate dehydrogenase (LDH), aspartate transaminase (AST) and alanine transaminase (ALT). The humoral immune response to Trichinella zimbabwensis infection was evaluated in all three groups by an indirect ELISA method. The results showed deviations from normal parameters of blood glucose, CPK, LDH, AST and ALT when compared with reported levels in uninfected reptiles. Contrary to studies involving mammals, hypoglycaemia was not observed in the infected groups in this study. Peak values of blood glucose were reached on post-infection (PI) Day 49, Day 42 and Day 35 in the high-infection, medium-infection and low-infection groups, respectively. Peak values of LDH and AST were observed on PI Day 56, Day 49 and Day 42 in the high-infection, medium-infection and low-infection groups, respectively. Peak values of CPK were observed on Day 35 PI in all three groups. Peak ALT values were reached on Day 56 in the high-infection group and on Day 28 PI in both the medium-infection and low-infection groups. No correlations between the biochemical parameters and infection intensity were observed. Peak antibody titres were reached on Day 49 PI in the medium-infection group, and on Day 42 PI in both the high-infection and low-infection groups. Infection intensity could not be correlated with the magnitude of the humoral immune response or time to sero-conversion. Results from this study were in agreement with results reported in mammals infected with other Trichinella species and showed that antibody titres could not be detected indefinitely.
Introduction
Apart from their sylvatic and domestic animal hosts, several Trichinella species infect humans; the most important species are Trichinella spiralis and Trichinella britovi (Gottstein et al. 2009). Humans become infected through the consumption of raw or undercooked meat from infected animals (Dupouy-Camet 2000; Gottstein et al. 2009). The zoonotic importance of Trichinella forms the basis for the implementation of measures aimed at the control or eradication of the parasite from the human food chain (Gottstein et al. 2009; Mukaratirwa et al. 2013). However, despite implementation of control measures, trichinellosis remains a major zoonotic threat in many parts of the world (Mukaratirwa et al. 2013; Murrell & Pozio 2011).
Trichinella larvae invade muscle tissue, which causes direct damage to the muscle cell during migration of larvae and indirectly by virtue of the inflammatory response of the host (Bruschi & Chiumiento 2011). This damage also coincides with increased cell membrane permeability and leakage of fluid into the adjacent tissue (Kociecka 2000). This process has been linked to an increase of creatine phosphokinase (CPK), lactate dehydrogenase (LDH), aspartate transaminase (AST) and alanine transaminase (ALT) in the blood (Kociecka 2000).
Serum levels of CPK, LDH and AST are considered to be supplementary in the diagnosis of Trichinella infections in humans (Gottstein et al. 2009). Jongwutiwes et al. (1998) reported elevated levels of CPK, LDH, AST and ALT in human patients infected with Trichinella pseudospiralis. Increased serum levels of ALT in pigs infected with T. spiralis have also been previously reported (Ribicich et al. 2007). However, elevated serum levels of these enzymes are not necessarily indicative of Trichinella infection, as there may be other causes (Koudela & Schanzel 1980; Ribicich et al. 2007; Srivastava & Chosdol 2007; Tassi et al. 1995; Wisniewska 1970). Comparisons between rats and human patients indicated that an increase in enzyme levels is dependent on the individual response from the host rather than being correlated with intensity of infection or clinical severity (Wisniewska 1970).
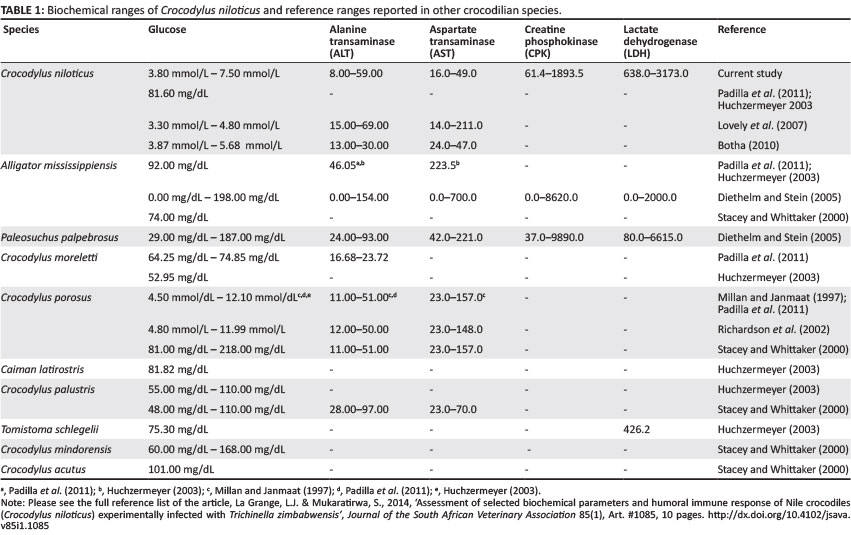
Normal biochemical values for several crocodile species have been previously reported (Lovely et al. 2007; Millan & Janmaat 1997; Padilla et al. 2011; Stacy & Whitaker 2000) but to the authors' knowledge, no studies have been conducted to investigate the effect of Trichinella infection on the biochemical parameters of Nile crocodiles.
Trichinella infection has also been reported to influence blood glucose levels, and hypoglycaemia has been reported in humans (Busila et al. 1968), mice (Nishina & Suzuki 2002; Wu et al. 2009) and dogs (Reina et al. 1989) infected with Trichinella species. The decrease in blood glucose has been attributed to the depletion of blood glucose by the parasite larvae (Wu et al. 2009). In a study involving mice infected with T. spiralis, hypoglycaemia was observed at 10 days post infection (PI) (Nishina & Suzuki 2002).
The cuticle of nematodes contains immune-specific antigens targeted by the host immune system (Phillip et al. 1981). Larvae and adults of the genus Trichinella are antigenically heterogeneous (Fabre et al. 2009). Enzyme linked immunosorbent assay (ELISA) is a commonly used method to measure the humoral immune response of the host to the antigen. The method relies on the metabolic excretory/secretory antigens (ESA) comprising of related glycoproteins that are released by the larvae (Gottstein et al. 2009). An important carbohydrate epitope, tyvelose, which is responsible for the induction of the humoral immune response, is situated on the TSL-1 antigen contained within stichocyte cells of the cuticle; a synthetic variant of this carbohydrate is used in the ELISA (Gottstein et al. 2009). Although highly specific, the use of synthetic tyvelose antigen in the ELISA is less sensitive than ESA (Gottstein et al. 2009).
Information is scant about the antibody response of the host against muscle stages of the parasite, but a mixed isotype response of Immunoglobulin G (IgG) 1, IgG2 and IgE has been reported in chronic infections, with IgG1 being the most dominant (Fabre et al. 2009). The use of serological tests as diagnostic tools in animal trichinellosis has previously been evaluated. Enzyme immunoassay tests (Gamble et al. 1996; Soule et al. 1989) and indirect immunofluorescence assays (Reina et al. 1996; Soule et al. 1989) have been conducted in goats and horses. Immunoassays were reported to be useful in horses but the study animals were all euthanased at 12 weeks post infection (Gamble et al. 1996), thus the persistence of antibodies beyond this time frame was never investigated. Similar results were also obtained in a study involving goats (Reina et al. 1996). In a study by Pozio et al. (2002), the immune response of horses against T. spiralis and Trichinella murrelli was evaluated for four-to-five months following infection using both ELISA and Western Blot techniques. In most cases, specific IgG was detectable only between three to seven weeks PI and maximum persistence of antibodies was recorded at 33 weeks PI in one horse infected with 20 000 T. murrelli larvae. Western Blot analysis and ELISA were also developed for pigs and, although relatively high specificity and sensitivity of the tests were reported (Nöckler et al. 2009), samples were not collected over extended periods to evaluate the persistence of antibodies. The aforementioned studies showed the practical application of these techniques to be limited. Specific antibodies against Trichinella do not persist indefinitely and can only be detected for limited periods following infection. Since Trichinella zimbabwensis is a non-encapsulated species, it is expected that the host immune response is stronger and more persistent due to the direct contact between the parasite larvae and host tissue (Huchzermeyer, F.W., pers. comm., 2008). An experimental study to determine the feasibility of the use of ELISA for the detection of T. zimbabwensis infection in crocodiles indicated that antibody titres decreased and eventually disappeared altogether (Ludovisi et al. 2013).
The objective of this study was to evaluate the effect of T. zimbabwensis infection intensity on the levels of blood glucose, AST, ALT, CPK and LDH and on the humoral immune response of experimentally infected crocodiles.
Materials and methods
Source of study animals
Fifteen 7-year-old Nile crocodiles (13 males and 2 females) within the size range of 1.35 m - 1.8 m in length were used in the present study. The crocodiles represented a group of animals not intended for commercial purposes and sourced from a farmed population with no history of T. zimbabwensis infection prior to commencement of the study. The animals were captured on the Wilderness Roads farm in Low's Creek, Mpumalanga Province, South Africa. They were immobilised with 0.4 mL Gallamine triethiodide (40 mg/mL) (Kyron, Johannesburg, South Africa), which was injected intramuscularly on the lateral aspect of the tail base of each animal before being transported to the experimental housing.
Pre-trial data collected from the animals included weight, sex and length of the animals. Clipping the horizontal scutes following a specific pattern on the tail identified each animal. For the high-infection group, both the left and right scutes were clipped in sequence according to the number assigned to the animal. For the low-infection and medium-infection groups, only the scutes on the left or right were clipped, respectively. The experiment was carried out from January 2012 to March 2012, when climatic conditions ensured good feeding and optimal physical condition of the animals.
Animal husbandry and feeding
Animal husbandry and feeding practices for the study animals were followed, as described by the South African National Standard for crocodiles in captivity SANS 631:2009 (SABS Standards Division 2009).
The study animals were housed on a smallholding belonging to the Mpumalanga Tourism and Parks Agency (MTPA) on the outskirts of Nelspruit, Mpumalanga province, South Africa. A fenced enclosure (10 m × 5 m) was constructed using 65 mm diamond mesh fencing, with a single-access gate. The enclosure allowed for a temperature gradient in the water to provide optimal thermoregulation (La Grange & Mukaratirwa 2014). During capture and sampling the pond was drained weekly and replenished with fresh water. Animals were fed with coarsely minced commercially bred chicken carcasses enriched with vitamin and mineral components (Feedmix, Johannesburg, South Africa) that were supplemented as 2.50 kg and 1.25 kg, respectively, in 1000 kg of wet ration.
The study animals were fed approximately 10 kg of food two to three times a week; no food was offered on days immediately prior to testing.
Source and preparation of infective material
As part of a previous study, T. zimbabwensis-infected meat was sourced from a crocodile experimentally infected with T. zimbabwensis (Ludovisi et al. 2013). Muscle tissue was collected from various sites, minced and thoroughly mixed using a ladle. One hundred grams of the homogenised sample was subjected to artificial digestion according to the method of Nockler and Kapel (2007) and infection level was determined to be 30 larvae per gram (LPG) of the homogenised sample. Infective material for each individual animal was calculated and separately packaged and refrigerated at 4 °C until the day of infection.
Infection of study animals
The animals were manually restrained and the infective dose was administered through a stomach tube. The high-infection group of crocodiles were infected with an average of 642 larvae/kg of bodyweight, the medium-infection group with an average of 414 larvae/kg of bodyweight and the low-infection group with an average of 134 larvae/kg of bodyweight. The animals were continuously monitored for at least 30 min following infection for side effects or regurgitation of infective material.
Collection of blood and sera
Blood was collected from each of the animals weekly from the date of infection (Day 0) until eight- weeks post infection (Day 56) to test for blood glucose and levels of CPK, LDH, ALT and AST. A 10 mL syringe and a 21-gauge needle were used to collect blood from the supra-vertebral sinus; approximately 8 mL blood was collected weekly from each animal. In order to separate the serum, blood was allowed to clot and the sample was centrifuged at 10 000 rpm for 15 min.
Preservation of samples
For the detection of antibodies, 2 mL serum from each animal was transferred to sterile cryotubes with screw caps and preserved in 0.01% merthiolate solution. The preserved samples were stored at 4 °C until completion of the trial. The remaining sera were frozen at -18 °C and used for the enzyme assays.
Testing of samples
Blood glucose was tested immediately after collection using an Accu-Chek® Active (Roche Diagnostics, Indianapolis, USA) glucometer to minimise the impact of stress (Smith & Marais 2004).
Sera preserved in 0.01% merthiolate were referred to the International Trichinella Reference Centre in Rome, Italy, for testing according to procedure based on Ludovisi et al. (2013).
Sera frozen at -18 °C were submitted to the biomedical research laboratory at the University of KwaZulu-Natal, Westville campus, Durban, South Africa, for testing. The samples were tested for LDH, ALT, AST and CPK enzyme levels. An automated chemistry analyser, Labmax Plenno (Lagoa Santa, Costa Brava, Brazil) was used for the analyses in accordance with manufacturer's recommendations.
Data analysis
Every week, mean blood glucose and serum concentrations of CPK, LDH, ALT and AST were determined for each group of animals over the trial period. Mean serum concentrations of biochemical parameters on Day 0 of the trial were compared, where possible, with normal reference ranges reported for Nile crocodiles (Botha 2010; Huchzermeyer 2003; Lovely et al. 2007; Padilla et al. 2011). Alternatively, where normal reference ranges for Nile crocodiles could not be found, comparisons were made with reference ranges reported for other crocodilian species (Diethelm & Stein 2006; Millan & Janmaat 1997; Padilla et al. 2011) (Table 1). Values from Day 7 PI were expressed as a percentage of the normal values recorded on Day 0 and calculated as follows:

Where Pv is the new percentage value, Iv is the initial value on the specific day and D0 represents the value on Day 0. Data were log transformed (log10[x+1]) and analysis of variance (IBM SPSS Statistics 19) was used to compare results from the different groups. Non-parametric, bivariate correlation (Spearman's rho) analyses (IBM SPSS Statistics 19) were conducted in order to determine relationships between biochemical parameters and infection dose.
Results
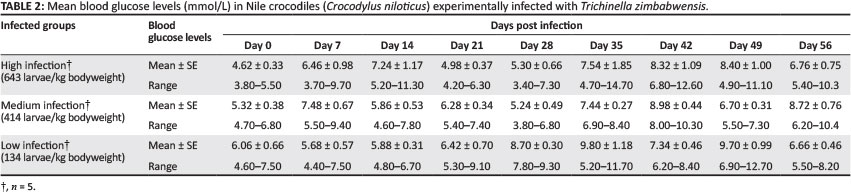
Blood glucose
The mean concentration of blood glucose for the three experimental groups is shown in Table 2. In the high-infection and medium-infection groups, initial peaks in blood glucose values were observed on Day 14 PI and Day 7 PI, respectively. This was followed by a slight decrease the following week. Analysis of variance revealed significant differences in blood glucose concentrations on Day 28 PI between the high-infection and low-infection groups and between the medium-infection and low-infection groups (p < 0.05). No correlation was observed between blood glucose and infection dose.
Alanine transaminase
Mean values of ALT for the three experimental groups are shown in Table 3. The highest increase of ALT was observed in the high-infection group on Day 49 PI and Day 56 PI, and it was 42% higher than the initial concentrations on Day 0. Increases in ALT in the medium-infection and low-infection groups were less pronounced and reached peak values of 8% and 11% higher than Day 0 at Day 28 PI. Analysis of variance showed no significant difference (p > 0.05) in the increases in ALT concentrations among the groups. However, mean ALT concentrations were correlated with mean blood glucose concentrations in the high infection group (R2 = 0.9,p < 0.05).
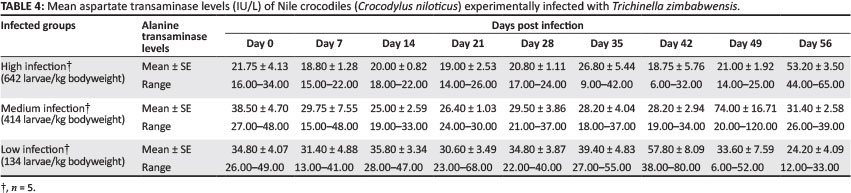
Aspartate transaminase
Mean values of AST for all three experimental groups are shown in Table 4. Maximum values were reached on Day 56 PI in the high-infection group. In the medium-infection group, AST values peaked on Day 49 PI and maximum values of AST were reached in the low-infection group on Day 42 PI. On Day 42 PI, the percentage increase of AST in the low-infection group was significantly higher than that observed in the medium-infection group (p < 0.05). Analysis of variance showed mean AST values in the low-infection group to be significantly higher on Day 14 PI and Day 42 PI (p < 0.05) compared with the high-infection group.
Creatine phosphokinase
The mean values of CPK for all three experimental groups are shown in Table 5. The low-infection group showed significantly lower values of CPK from Day 0 compared with the high-infection and medium-infection groups. Analysis of variance showed significant differences in changes of mean CPK values between the high-infection and low-infection groups from Day 0 until Day 14 of the trial. The percentage increases in CPK values in the low-infection group was significantly higher (p < 0.05). Mean CPK values were significantly increased in the low-infection group compared with the medium-infection group on Day 7 PI, Day 42 PI, and Day 56 PI (p < 0.05). Similarly, CPK in the low-infection group increased to significantly higher levels than those recorded for the high-infection group on Day 42 PI and Day 56 PI (p < 0.05). The mean increase in CPK value was also significantly higher in the medium-infection group compared with the high-infection and low-infection groups on Day 28 PI (p < 0.05). Mean CPK values were significantly correlated with both initial infection dose and mean blood glucose (R2 = 0.9, p < 0.05) in the high-infection group, but a significantly negative correlation was observed with overall infection dose in this group (R2 = -0.9, p < 0.05). Mean CPK was negatively correlated with mean blood glucose in the medium-infection group (R2 = -0.9, p < 0.05). No significant correlation could be established between infection dose and peak CPK values.
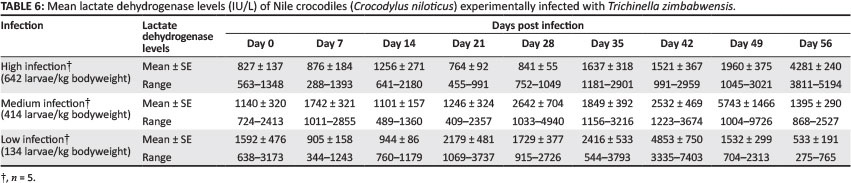
Lactate dehydrogenase
The values of LDH for all three experimental groups are shown in Table 6. No correlation could be established between peak LDH values and infection dose. Mean increase in LDH was significantly higher in the low-infection group on Day 28 PI and Day 35 PI compared with the medium-infection group (p < 0.05). Mean increases in LDH values were significantly higher in the low-infection group compared to the high-infection group on Day 35 PI (p < 0.05). Mean LDH concentrations in the low-infection group reached significantly higher values on Day 21 PI and Day 42 PI compared with the high-infection group (p < 0.05). Mean increase in values of LDH was significantly higher in the high-infection group on Day 56 PI compared with the medium-infection and low-infection groups (p < 0.05). LDH values were also significantly higher in the medium-infection group compared with the high-infection group on Day 28 PI and Day 49 PI (p < 0.05). Mean LDH values were also significantly higher in the medium-infection group on Day 56 PI compared with the low-infection group (p < 0.05). Mean LDH values were negatively correlated with blood glucose in the medium- infection group (R2 = -0.9,p < 0.05). A significant correlation was also observed between mean LDH concentrations and infection dose in the low-infection group (R2 = 0.9, p < 0.05).
Indirect enzyme immnunosorbent assay
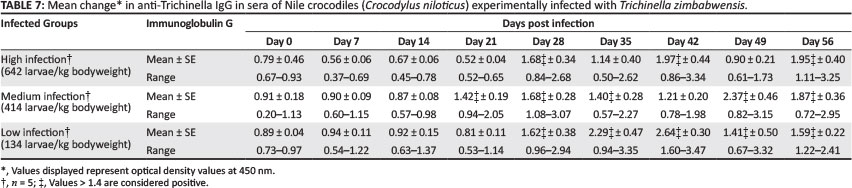
Sero-conversion was observed in all of the experimental animals. Analysis of variance showed no significant differences in the titre levels between the three groups, despite the fact that mean optical density (OD) values in the high-infection group were lower compared with the medium- infection group. Mean OD values were also lower in the medium-infection group compared with the low-infection group. No significant correlation between infection dose and magnitude of the humoral immune response could be established. Peak antibody titres were reached at Day 42 PI in both the high-infection and low-infection groups, whereas the medium-infection group reached its highest value on Day 49 PI. Analysis of variance, however, showed significant differences between the groups on Day 7 PI, Day 21 PI and Day 42 PI. On Day 7, OD values were significantly lower in the high-infection group (p < 0.05) compared with the medium-infection and low-infection groups. On Day 21, OD values in both the high-infection and low-infection groups were significantly lower than in the medium-infection group (p < 0.05). On Day 42, OD values in the low-infection group were significantly higher compared with the high-infection group. However, no significant correlation could be established between infection dose and time to sero-conversion.
Ethical consideration
Ethical approval for the study was obtained through the Animal Ethics Research Committee of the University of KwaZulu-Natal (reference number 035/12/Animal). Where applicable, ARRIVE guidelines for reporting in vivo animal experiments (Kilkenny et al. 2010) have been adhered to.
Discussion
Mean blood glucose ranges of the experimental crocodiles on Day 0 compared favourably with normal reference ranges reported for Nile crocodiles (Botha 2010; Huchzermeyer 2003; Lovely et al. 2007; Padilla et al. 2011). Only four animals in this study showed blood glucose values higher than those reported by previous authors, but they still fell within the ranges reported for estuarine crocodiles Crocodylus porosus (Millan & Janmaat 1997; Padilla et al. 2011).
Wu et al. (2009) reported an initial drop in blood glucose levels between Day 8 PI and Day 28 PI in mice experimentally infected with T. pseudospiralis and T. spiralis. Interestingly, the results reported by Wu et al. (2009) showed that the lowest levels of blood glucose in mice were reached at Day 13 PI and Day 18 PI for T. spiralis and T. pseudospiralis, respectively; T. pseudospiralis-infected mice took longer to return to normal. Similarly, mice infected with T. spiralis in other studies showed hypoglycaemia at Day 10 PI (Nishina & Suzuki 2002; Nishina et al. 2004). The initial decrease in blood glucose observed on Day 7 PI in the low-infection group in the present study is consistent with results obtained from mice experimentally infected with T. spiralis (Nishina & Suzuki 2002). In contrast, blood glucose levels increased between Day 0 PI and Day 7 PI in the high-infection group and between Day 0 PI and Day 14 PI in the medium-infection group in the present study. This suggests that the level or intensity of infection has an effect on blood glucose levels. Modulation of the insulin pathway by the parasite is considered to cause hypoglycaemia, and blood glucose levels correspond to larval growth (Wu et al. 2009). Wu et al. (2009) additionally reported that minimum blood glucose levels in mice experimentally infected with T. spiralis and T. pseudospiralis were reached on Day 13 PI and Day 18 PI, respectively; in the case of the latter, recovery time from hypoglycaemia was longer. These differences suggest that parasite-specific mechanisms are responsible. However, the initial rise in blood glucose levels observed in the high-infection and medium-infection groups in this study cannot be explained. In the study by Wu et al. (2009), blood glucose levels of the infected mice steadily increased following their initial drop and reached almost normal levels by Day 48 PI; they rarely reached levels above normal. In this study, however, blood glucose levels were maintained above the initial levels recorded on Day 0 and never decreased to normal levels during the experimental period. This phenomenon cannot be explained, but it could be speculated that crocodiles possess physiological adaptations that allow them to up-regulate blood glucose concentrations in accordance with the existing demand.
Dolphins have been reported to switch between a normal and hyperglycaemic state when presented with daily feeding and fasting routines (Venn-Watson et al. 2011). This adaptation is necessary to enable these animals to maintain high blood glucose levels, as demanded by their physiology (Venn-Watson et al. 2011). The elevated blood glucose levels observed in this study may also have been influenced by stress associated with capture and restraint (Smith & Marais 2004), but the observed variation in responses between the experimental groups suggests that the effects of infection superseded those caused by other stressors. However, the increase in blood glucose levels observed in the medium-infection and high-infection groups between Day 28 PI and Day 42 PI, and Day 21 PI and Day 35 PI in the low-infection group, may be due to the arrival and infiltration of newborn larvae in the muscle tissue. In this study, minimum and maximum blood glucose levels were not reached at the same time in the three groups, but were marked by a one-week delay between each group; the low-infection group reached the high levels first. Studies with mice and reptiles infected with T. pseudospiralis and T. zimbabwensis, respectively, showed larvae of these two species to be larger in poikilotherms than in mammalian hosts (Pozio et al. 2004). This suggests that the metabolic rate of the host may influence larval growth (Pozio et al. 2004).
If the growth rate and, subsequently, the metabolic rate of T. zimbabwensis can be altered as a result of variation in host metabolism, similar alterations may be caused in accordance with the availability of nutrients, which may explain the delays observed between the experimental groups of this study. Another hypothesis could be that this strategy is employed by the parasite to prevent the sudden release of large numbers of larvae into the host circulation. Large larval burdens in the blood circulation could potentially restrict blood flow in vital organs or produce acute anaphylaxis causing host fatality, which ultimately is detrimental to survival of the parasite.
Mean ALT of the experimental crocodiles compared favourably on Day 0, with normal reference ranges reported for Nile crocodiles (Botha 2010; Lovely et al. 2007). Only one animal in this study displayed ALT concentrations lower than those reported by previous authors, but still fell within the range reported for American alligators (Alligator mississippiensis) (Diethelm & Stein 2006). Increased levels of ALT are normally more indicative of liver disease and, according to Srivastava and Chosdol (2007), increase in the levels of this enzyme is rarely noted in other cases.
Peak values of ALT were not reached simultaneously in the three groups. Increases in ALT concentrations were less pronounced than those of CPK and LDH, suggesting that ALT is less sensitive to the effects of the parasite on host tissue and that significant increases in this enzyme may only occur in cases of high infection intensity that result in severe tissue damage. This supports the specific nature of this enzyme as an indicator of liver disease rather than muscle damage. Assessment of the individual crocodile results additionally showed high variability within the three groups; this further suggests that the effect of individual host physiology on ALT concentrations supersedes the effect of parasite invasion, especially in lower level infections, further supporting the less sensitive nature of this enzyme compared to CPK and LDH.
Mean AST values of the experimental crocodiles on Day 0 were comparable with normal reference ranges reported for Nile crocodiles (Botha 2010; Lovely et al. 2007). Peak AST concentrations did not appear to coincide with the arrival of newborn larvae in muscles but instead reached peak values much later during the course of infection, which coincided with the response observed in LDH. In contrast to ALT, the response of individual crocodiles in the different groups was also more uniform and this suggests that this enzyme is more specific in its response to the parasitic effects of T. zimbabwensis than ALT, but less sensitive than CPK and LDH. This is in agreement with reports of increased AST concentrations associated with muscle injuries (Srivastava & Chosdol 2007). Normally, AST levels are higher than ALT and the ratio of AST:ALT are reported to be > 1 (Srivastava & Chosdol 2007). Normal values of these enzymes, as recorded for the experimental crocodiles in this study, support this; AST values were on average higher (31.9 IU/L) than ALT (28.7 IU/L). Srivastava and Chosdol (2007) also reported that AST:ALT ratios may change to < 1 in cases where tissue damage is severe. This was also supported by the results of this study, as mean ALT concentrations (38.8 IU/L) exceeded those of AST (31.8 IU/L) over the course of the experiment.
No reference ranges for CPK for Nile crocodiles have been reported to date. However, mean CPK values of animals in this study on Day 0 were comparable with the ranges reported for American alligators and Dwarf caimans (Paleosuchus palpebrosus) (Diethelm & Stein 2006).
The single peak increase in CPK concentrations in the low-infection group on Day 35 PI may correspond with the arrival of larvae and subsequent invasion of muscle tissue. It further suggests that the newborn larvae reached their respective sites of predilection simultaneously, which resulted in a singular, large-scale myopathy event. Analyses of the individual results confirmed that all five animals in this group reached peak CPK concentrations on Day 35 PI. In the medium-infection and high-infection groups, the increase in CPK concentration was not only less pronounced but the subsequent decrease was more protracted compared with the low-infection group. The CPK values remained higher than those recorded on Day 0 in both these groups. This suggests that in these groups, larvae were released into the bloodstream in waves rather than in a single event. This further supports the hypothesis that development of some larvae may have been delayed. In rats and humans, increases in CPK levels were attributed rather to the level of individual response of the host than to specific damage caused by the parasite (Wisniewska 1970). In this study, however, peak concentrations corresponded with the arrival of newborn larvae in the muscles around Day 35 PI. According to Srivastava and Chosdol (2007), CPK levels may rise within 3Â6 h in humans following a heart attack and will return to normal levels within 12Â48 h, if no further damage occurs. The results from this study appear to support this, as in the low-infection group, CPK levels decreased immediately after an initial peak on Day 35 PI. However, in the other two groups, CPK did not reach normal levels. This would suggest that muscle damage continued beyond the peak observed on Day 35 PI and further supports the hypothesis that larvae may have been released in waves in the medium-infection and high-infection groups.
Contrary to the case in human trichinosis, increases in CPK levels in this study did not exceed the concurrent increase of other enzymes (Wisniewska 1970) and in fact was less prominent compared with LDH and AST in this study. The fact that CPK levels in the low-infection group did not increase from their lowest levels on Day 49 PI and Day 56 PI, or persist at concentrations higher than those observed on Day 0, may suggest that all of the larvae had invaded the host musculature earlier. No new events of myopathy occurred after Day 35 PI, thus, CPK concentrations decreased to normal levels. However, in the medium-infection group, there is a possibility that the initial damage was more severe and, thus, the decrease in CPK levels was more protracted. In the high-infection group, however, initial damage caused by the larvae was more severe and after Day 35 PI, new larvae continued to be released from the intestine. This explains the subsequent increase in CPK at Day 49 PI. The negative correlation observed between mean blood glucose and mean CPK in the medium-infection group may be explained by the fact that CPK levels continued to decrease beyond Day 49 PI, following its initial peak on Day 35 PI, whereas blood glucose levels increased again after Day 49 PI. If a second wave of larvae was released in this group, as suggested by the rise in glucose levels on Day 49 PI, their numbers may have been too few to cause a significant increase in the already elevated CPK levels recorded on Day 49 PI. In the high-infection group, however, the second wave of larvae resulted in a secondary increase in CPK levels, which coincided with the increase in blood glucose that resulted in the positive correlation observed in this group.
No reference ranges for LDH could be found for Nile crocodiles. Mean LDH of the experimental crocodiles on Day 0 fell well within normal reference ranges reported for Dwarf caimans, as reported by Diethelm and Stein (2006).
In horses infected with T. spiralis, increases in CPK, LDH and aldolase all peaked at the fifth week PI and LDH was found to be more sensitive than CPK and aldolase (Soule et al. 1989). The sensitivity of LDH was also supported by results from the current study, even though both LDH and AST concentrations peaked later than in the case of CPK. The delayed increase in LDH and AST observed in this study appeared not to coincide with larval migration, but serum concentrations of enzymes might have been influenced by other factors (Srivastava & Chosdol 2007). Although the mean increase in LDH values appeared to be correlated with infection intensity, no direct correlation between these two factors could be established. Koudela and Schanzel (1980) reported that the relationship between levels of LDH and infection intensity was not always apparent when comparing individual animals of the same group. In guinea pigs infected with Trichinella spp. larvae, LDH continued to increase until Day 42 PI (Koudela & Schanzel 1980), which was similar to the results observed in the low-infection group in the current study. The continued increase in LDH observed in the medium-infection and high-infection groups once again suggests that larval development and subsequently larval migration may have been delayed in these groups. Similar to CPK, LDH was also negatively correlated with serum glucose in the medium-infection group. Although LDH concentrations increased beyond Day 35 PI, there was also a sharp decrease immediately following a peak on Day 49 PI. The differences in time to reach peak LDH and AST values observed between the experimental groups suggest that these enzymes are not necessarily indicators of muscle damage. The kinetics of these enzymes appears to correspond better with the migration of larvae rather than with actual damage associated with the larvae, this is in agreement with the hypothesis of Koudela and Schanzel (1980).
An indirect ELISA was developed to detect the immune response of crocodiles to T. zimbabwensis infection (Ludovisi et al. 2013). Immune responses could be detected up to six weeks PI, but not beyond this time, and the ELISA was deemed to be unsuitable for surveillance purposes (Ludovisi et al. 2013). Previous studies have shown that the intensity of infection cannot always be correlated with antibody titres (Ludovisi et al. 2013; Pozio et al. 2002; Vu Thi et al. 2010). In crocodiles, interpretation of antibody titres is also complicated, as the immune response may also be influenced by age, temperature, season, and hormone levels (Brown et al. 2001; Ludovisi et al. 2013). Gamble et al. (1996) reported on the efficacy of enzyme immunoassays to detect light infections and indicated that the time period between infection and sero-conversion of the host was problematic in surveillance. The results from the present study confirm this, since the time from infection to sero-conversion ranged from Day 21 PI to Day 56 PI. Kinetics of the antibody response additionally suggest mean sero-conversion in the high-infection group to have been delayed by a week compared to the medium-infection group and by two weeks compared to the low-infection group. Furthermore, in this study, OD values for 10 of the crocodiles decreased in the week following sero-conversion before increasing again. Thus, even if the initial period of infection is known or suspected, surveillance studies will not be effective where they have to rely on a single sample, as the potential will always exist for false negative results to be obtained. Similar to results obtained from a previous study (Ludovisi et al. 2013), no positive correlation between infection intensity and the time to sero-conversion could be established in this study, as has been reported in previous studies involving mammalian hosts (Gottstein et al. 2009). Results from this study were also in agreement with those from previous studies where no direct correlation could be established between infection intensity in muscles and antibody levels (Ludovisi et al. 2013; Pozio et al. 2002; Vu Thi et al. 2010).
In crocodiles, the potential effects of individual hormonal status, temperature fluctuations and age on the immune response (Brown et al. 2001; Ludovisi et al. 2013) may explain these phenomena. However, despite the observed differences between infection intensity and time to sero-conversion, the lower humoral response associated with higher infection levels may also be the result of larvae being released at different intervals, which could result in smaller but repetitive responses from the host. The results revealed similar problems to those reported in mammals infected with other Trichinella species, which showed that in most cases antibody titres could not be detected indefinitely (Gottstein et al. 2009; Ludovisi et al. 2013; Soule et al. 1989).
Conclusion
For logistical reasons the study design did not make provision for a control group. However, the results remain valuable, as additional new information about biochemical parameters and humoral immune response of Nile crocodiles (Crocodylus niloticus) to infection with Trichinella zimbabwensis was obtained.
The fact that hypoglycaemia was not observed in crocodiles experimentally infected with T. zimbabwensis suggests that Nile crocodiles may up-regulate blood glucose as the demand for energy is increased by the parasites. This phenomenon requires further investigation in order to elucidate the specific mechanisms and pathways involved, and to determine whether this is common among other ectothermic hosts. Future research should be aimed at determining the influence of infection dose on the rate of development and release of larvae, and the impact of these factors on the different haematological and biochemical parameters of the host.
High variability in ALT changes between individual crocodiles supports the finding that increased ALT activity is not reliable as an indicator for muscle damage and that increases in ALT are rarely observed in cases where damage to the liver is not involved. With exception of the high-infection group, it is suggested that differences in ALT concentrations observed in the crocodiles can largely be attributed to variations in individual physiology of the crocodiles.
The more reliable indicator of muscle damage was CPK, as its peak values correspond with the arrival of larvae in the muscle, in spite of previous reports to the contrary. There was lack of correlation between infection intensity and changes in biochemical parameters and this supports the findings that other host factors, such as hormone levels and the age of the animal, as well as environmental factors, including temperature and season, may impact on the immune response of reptiles. The lack of information on normal reference ranges for haematological and biochemical parameters in Nile crocodiles should be addressed in future research.
Acknowledgements
Our sincere gratitude to the following organisations and people without whom this work would not have been possible: Mrs T. La Grange for her assistance with the many late hours of data capturing and laboratory work; Dr J.P. Raath of Wildlife Pharmaceuticals and Wildlifevets.com for the generous financial, technical and logistical support provided; Mpumalanga Tourism and Parks Agency for providing the housing facilities for experimental animals used in this study; Dr D. Brugman for his assistance during the post mortem examinations; the Crocodile Specialist Group for providing financial assistance in the form of a grant through their Student Research Assistance Scheme; Seronera and Wilderness Roads crocodile farms for the provision of experimental animals and food; Dr D. Pfukenyi, Prof. M. Hosie and Ms L. Laubscher for their assistance with the final data analysis; the Community Reference Centre of the European Union and Dr Pozio and Dr Gomez-Morales in Rome, Italy for their assistance in the antibody testing; the staff and students of the Biomedical research laboratory of the University of Kwa-Zulu Natal for conducting the enzyme testing.
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this article.
Authors' contributions
L.J.L.G. (Department of Agriculture, Rural Development and Land Administration) contributed to the study design and field experiments, data analysis, interpretation and drafting of manuscript and tables. S.M. (University of KwaZulu-Natal) also contributed to the study design and supervision of experiments, assisted with interpretation, drafting and editing of manuscript.
References
Botha, P.J., 2010, 'The distribution, conservation status and blood biochemistry of Nile crocodiles in the Olifants river system, Mpumalanga, South Africa', Doctoral thesis, Centre for Wildlife Management, Department of Animal and Wildlife Services, University of Pretoria, South Africa. [ Links ]
Brown, D., Schumacher, I., Nogueira, M., Richey, L.J., Zacher, L.A., Schoeb, T.R., Vliet, K.A., Bennet, R.A., Jacobson, E.R. & Brown, M.B., 2001, 'Detection of antibodies to a pathogenic Mycoplasma in American alligators (Alligator mississippiensis), broad-nosed caimans (Caiman latirostris), and Siamese crocodiles (Crocodylus siamensis)', Journal of Clinical Microbiology 39, 285-292. http://dx.doi.org/10.1128/JCM.39.1.285-292.2001 [ Links ]
Bruschi, F. & Chiumiento, L., 2011, 'Trichinella inflammatory myopathy: Host or parasite strategy?', Parasites and Vectors 4, 1-6. http://dx.doi.org/10.1186/1756-3305-4-42 [ Links ]
Busila, V.T., Dragomirescu, M., Dragomirescu, L. & Maager, P., 1968, 'Functional and metabolic alterations in human trichinellosis', Wiadomosci Parazytologiczne 14, 195-200. [ Links ]
Diethelm, G. & Stein, G., 2006, 'Blood chemistry values in reptiles hematologic and serum biochemical values of reptiles', in D.R. Mader (ed.), Reptile Medicine and Surgery, 2nd edn., Saunders Elsevier, St. Louis, 2006. http://dx.doi.org/10.1016/B0-72-169327-X/50092-4 [ Links ]
Dupouy-Camet, J., 2000, 'Trichinellosis: A worldwide zoonosis', Veterinary Parasitology 93, 191-200. http://dx.doi.org/10.1016/S0304-4017(00)00341-1 [ Links ]
Fabre, M.V., Beiting, D.P., Bliss, S.K. & Appleton, J.A., 2009, 'Immunity to Trichinella spiralis muscle infection', Veterinary Parasitology 159, 245-248. http://dx.doi.org/10.1016/j.vetpar.2008.10.051 [ Links ]
Gamble, H.R., Gajadhar, A.A. & Solomon, M.B., 1996, 'Methods for the detection of trichinellosis in horses', Journal of Food Protection 59, 420-425. [ Links ]
Gottstein, B., Pozio, E. & Nöckler, K., 2009 'Epidemiology, diagnosis, treatment, and control of trichinellosis', Clinical Microbiology Reviews 22, 127-145. http://dx.doi.org/10.1128/CMR.00026-08 [ Links ]
Huchzermeyer, F.W., 2003, Crocodiles: Biology, Husbandry and Diseases, CABI Publishing, Wallingford. http://dx.doi.org/10.1079/9780851996561.0000 [ Links ]
Jongwutiwes, S., Chantachum, N., Kraivichian, P., Siriyasatien, P., Putaporntip, C., Tamburrini, A., La Rosa, G., Sreesunpasiriku, C., Yingyourd, P. & Pozio, E., 1998, 'First outbreak of human trichinellosis caused by Trichinella pseudospiralis', Clinical Infectious Diseases 26, 111-115. http://dx.doi.org/10.1086/516278 [ Links ]
Kilkenny, C., Brown, W.J., Cuthill, I.C., Emerson, M. & Altman, D.G., 2010, 'Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research', PLoS Biology 8, e1000412. http://dx.doi.org/10.1371/journal.pbio.1000412 [ Links ]
Kociecka, W., 2000, 'Trichinellosis: Human disease, diagnosis and treatment', Veterinary Parasitology 93, 365-383. http://dx.doi.org/10.1016/S0304-4017(00)00352-6 [ Links ]
Koudela, B. & Schanzel, H., 1980, 'Activity of lactate dehydrogenase in the course of experimental trichinellosis in guinea pigs', Acta Veterinaria Brno 49, 85-89. http://dx.doi.org/10.2754/avb198049010085 [ Links ]
La Grange, L.J. & Mukaratirwa, S., 2014, 'Distribution patterns and predilection muscles of Trichinella zimbabwensis larvae in experimentally infected Nile crocodiles (Crocodylus niloticus Laurenti)', Onderstepoort Journal of Veterinary Research 81(1), Art. #652, 7 pages. http://dx.doi.org/10.4102/ojvr.v81i1.652 [ Links ]
Lovely, C.J., Pittman, J.M. & Leslie, A.J., 2007, 'Normal haematology and blood biochemistry of wild Nile crocodiles (Crocodylus niloticus) in the Okavango Delta, Botswana', Journal of the South African Veterinary Association 78, 137-144. http://dx.doi.org/10.4102/jsava.v78i3.305 [ Links ]
Ludovisi, A., La Grange, L.J., Gomez-Morales, M.A. & Pozio, E., 2013, 'Development of an ELISA to detect the humoral immune response to Trichinella zimbabwensis in Nile crocodiles (C. niloticus)', Veterinary Parasitology 194(2-4), 189-192. http://dx/doi.org/10.1016/j.vetpar.2013.01.053 [ Links ]
Millan, J.M. & Janmaat, A., 1997, 'Reference ranges for biochemical and haematological values in farmed saltwater crocodile (Crocodylus porosus) yearlings', Australian Veterinary Journal 75, 814-817. http://dx.doi.org/10.1111/j.1751-0813.1997. tb15660.x [ Links ]
Mukaratirwa, S.M., La Grange L.J. & Pfukenyi, D.M., 2013, 'Trichinella infections in animals and humans in sub-Saharan Africa: A review', Acta Tropica 125, 82-89. http://dx.doi.org/10.1016/j.actatropica.2012.09.005 [ Links ]
Murrell, K.D. & Pozio, E., 2011, 'The worldwide occurrence and impact of human trichinellosis, 1986-2009', Emerging Infectious Diseases 17, 2194-2202. http://dx.doi.org/10.3201/eid1712.110896 [ Links ]
Nishina, M. & Suzuki, M., 2002 'Biphasic hypothermia in mice infected with a parasitic nematode, Trichinella spiralis', Experimental Animals 51, 207-211. http://dx.doi.org/10.1538/expanim.51.207 [ Links ]
Nishina, M., Suzuki, M. & Matsushita, K., 2004, 'Trichinella spiralis: Activity of the cerebral pyruvate recycling pathway of the host (mouse) in hypoglycemia induced by the infection', Experimental Parasitology 106, 62-65. http://dx.doi.org/10.1016/j.exppara.2004.01.007 [ Links ]
Nöckler, K. & Kapel, C.M.O., 2007, 'Detection and surveillance for Trichinella: Meat inspection hygiene, and legislation', in J. Dupouy-Camet & K.D. Murrell (eds.), FAO/ WHO/OIE guidelines for the surveillance, management, prevention and control of trichinellosis, pp. 69-97, World Organisation for Animal Health Press, Paris. [ Links ]
Nöckler, K., Reckinger, S., Broglia, A., Mayer-Scholl, A. & Bahn, P., 2009, 'Evaluation of a Western Blot and ELISA for the detection of anti-Trichinella-IgG in pig sera', Veterinary Parasitology 163, 341-347. http://dx.doi.org/10.1016/j.vetpar.2009.04.034 [ Links ]
Padilla, S.C., Weber, M. & Jacobson, E.R., 2011, 'Hematologic and plasma biochemical reference intervals for Morelet's crocodiles {Crocodylus moreletii) in the Northern wetlands of Campeche, Mexico', Journal of Wildlife Diseases 47, 511-522. http://dx.doi.org/10.7589/0090-3558-47.3.511 [ Links ]
Phillip, M., Taylor, P.M., Parkhouse, R.M.E. & Ogilvie, B.M., 1981, 'Immune response to stage specific surface antigens of the parasitic nematode Trichinella spiralis', Journal of Experimental Medicine 154, 210-215. http://dx.doi.org/10.1084/jem.154.1.210 [ Links ]
Pozio, E, Marucci, G., Casulli, A., Sacchi, L., Mukaratirwa, S., Foggin, C.M. & La Rosa, G., 2004, 'Trichinella papuae and Trichinella zimbabwensis induce infection in experimentally infected varans, caimans, pythons and turtles', Parasitology 128, 333-342. http://dx.doi.org/10.1017/S0031182003004542 [ Links ]
Pozio, E., Sofronic-Milosavljevic, L., Gomez-Morales, M.A., Boireau, P. & Nóckler, K., 2002, 'Evaluation of ELISA and Western Blot analysis using three antigens to detect anti-Trichinella IgG in horses, Veterinary Parasitology 108, 163-178. http://dx.doi.org/10.1016/S0304-4017(02)00185-1 [ Links ]
Reina, D., Habela, M., Navarette, I. & Martínez-Gómez, G., 1989, 'Hypoglycaemia in experimental canine trichinellosis', Veterinary Parasitology 33, 289-296. http://dx.doi.org/10.1016/0304-4017(89)90138-6 [ Links ]
Reina, D., Munoz-Ojeda, M. & Serrano, F., 1996, 'Experimental trichinellosis in goats', Veterinary Parasitology 62, 125-132. http://dx.doi.org/10.1016/0304-4017(95)00835-7 [ Links ]
Ribicich, M., Gamble, H., Rosa, A., Sommerfelt, I., Marquez, A., Mira, G., Cardillo, N., Cattaneo, M.L., Falzoni, E. & Franco, A., 2007, 'Clinical, haematological, biochemical and economic impacts of Trichinella spiralis infection in pigs' Veterinary Parasitology 147, 265-270. http://dx.doi.org/10.1016/j.vetpar.2007.04.017 [ Links ]
SABS Standards Division, 2009, South African National Standard Crocodiles in captivity, SANS 631, 1st edn., SABS Standards Division, Pretoria. [ Links ]
Smith, G.A. & Marais, J., 2004, 'Stress in crocodilians - the impact of nutrition', Proceedings of the 12th Working Meeting of the Crocodile Specialist Group, vol. 2, pp. 2-38, IUCN- The World Conservation Union, Gland, Switzerland. [ Links ]
Soule, C., Dupouy-Camet, J., Georges, P., Ancelle, T., Gillet, J.P., Vaissaire, J., Delvigne, A. & Plateau, E., 1989, 'Experimental trichinellosis in horses: Biological and parasitological evaluation', Veterinary Parasitology 31, 19-36. http://dx.doi.org/10.1016/0304-4017(89)90005-8 [ Links ]
Srivastava, T. & Chosdol, K., 2007, Clinical biochemistry: Clinical enzymology and its applications, Department of Biochemistry, All India Institute of Medical Sciences, Ansari Nagar, New Delhi. [ Links ]
Stacy, B.A. & Whitaker, N., 2000, 'Hematology and blood biochemistry of captive mugger crocodiles {Crocodylus palustris)', Journal of Zoo and Wildlife Medicine 31, 339-347. [ Links ]
Tassi, C., Materazzi, L., Pozio, E. & Bruschi, F., 1995, 'Creatine kinase isoenzymes in human trichinellosis', International Journal of Clinical Chemistry 2, 197-202. [ Links ]
Venn-Watson, S., Carlin, K. & Ridgway, S., 2011, 'Dolphins as animal models for type 2 diabetes: Sustained, post-prandial hyperglycemia and hyperinsulinemia', General and Comparative Endocrinology 170, 193-199. http://dx.doi.org/10.1016/j.ygcen.2010.10.005 [ Links ]
Vu Thi, N., Dorny, P., La Rosa, G., To Long, T., Nguyen Van, C. & Pozio, E., 2010, 'High prevalence of anti-Trichinella IgG in domestic pigs of the Son La province, Vietnam', Veterinary Parasitology 168, 136-140. http://dx.doi.org/10.1016/j.vetpar.2009.10.024 [ Links ]
Wisniewska, M., 1970, 'Trichinella spiralis: Diagnostic value of creatine kinase levels in rat and man', Experimental Parasitology 28, 577-584. http://dx.doi.org/10.1016/0014-4894(70)90126-8 [ Links ]
Wu, Z., Nagano, I., Kajita, K., Nishina, M. & Takahashi, Y., 2009, 'Hypoglycaemia induced by Trichinella infection is due to the increase of glucose uptake in infected muscle cells', International Journal for Parasitology 39, 427-434. http://dx.doi.org/10.1016/jJ.ijpara.2008.09.001 [ Links ]
 Correspondence:
Correspondence:
Louis La Grange
Private Bag X11309
Nelspruit 1200
South Africa
Email: croc.research@gmail.com
Received: 11 July 2013
Accepted: 12 Nov. 2013
Published: 21 Aug. 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}