










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkJournal of the South African Veterinary Association
versión On-line ISSN 2224-9435
versión impresa ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.85 no.1 Pretoria ene. 2014
ORIGINAL RESEARCH
Development of a flow cytometric bead immunoassay and its assessment as a possible aid to potency evaluation of enterotoxaemia vaccines
Angela BuysI; Raynard MacdonaldI; Jannie CraffordIII; Jacques TheronII
IQuality Control Department, Onderstepoort Biological Products, South Africa
IIDepartment Microbiology and Plant Pathology, University of Pretoria, South Africa
IIIDepartment of Veterinary Tropical Diseases, University of Pretoria, South Africa
ABSTRACT
Enterotoxaemia, an economically important disease of sheep, goats and calves, is caused by systemic effects of the epsilon toxin produced by the anaerobic bacterium Clostridium perfringens type D. The only practical means of controlling the occurrence of enterotoxaemia is to immunise animals by vaccination. The vaccine is prepared by deriving a toxoid from the bacterial culture filtrate and the potency of the vaccine is tested with the in vivo mouse neutralisation test (MNT). Due to ethical, economic and technical reasons, alternative in vitro assays are needed. In this study an indirect cytometric bead immunoassay (I-CBA) was developed for use in vaccine potency testing and the results were compared with those obtained using an indirect enzyme-linked immunosorbent assay (I-ELISA) and the MNT. Sera were collected from guinea pigs immunised with three different production batches of enterotoxaemia vaccine and the levels of anti-epsilon toxin antibodies were determined. Although the intra- and inter-assay variability was satisfactory, epsilon antitoxin levels determined by both the I-ELISA and indirect cytometric bead immunoassay (I-CBA) tests were higher than those of the MNT assay. In contrast to the MNT, all of the serum samples were identified as having antitoxin levels above the required minimum (not less than 5 U/mL). These results indicate that the respective in vitro tests in their current formats are not yet suitable alternatives to the in vivo MNT. The growing demand for a more humane, cost-effective and efficient method for testing the potency of enterotoxaemia vaccines, however, provides a strong impetus for further optimisation and standardisation of the I-CBA assay but further analytical research is required.
Introduction
Enterotoxaemia caused by Clostridium perfringens type D is a fatal disease of sheep, goats and calves (Niilo 1980; Uzal 2004). The major pathological agent is the epsilon toxin that is produced as an inactive prototoxin and activated by proteolytic enzymes (Bhown & Habeeb 1977; Minami et al. 1997; Songer 1996). Since enterotoxaemia in young animals often evolves to a peracute stage, the disease is difficult to treat. Economic losses may, however, be prevented by immunisation through vaccination (Oyston et al. 1998; Titball 2009). In South Africa, a monovalent vaccine containing the epsilon toxoid is manufactured and licensed by Onderstepoort Biological Products Ltd, Pretoria. The requirements for assessing the quality, safety, efficacy and potency of these vaccines are contained in Monograph 01/2008:0363 of the European Pharmacopoeia.
The potency of C. perfringens type D vaccines is measured in terms of their ability to induce antitoxins against the epsilon toxoid and the in vivo mouse neutralisation test (MNT) is the statutory method for determining the level of anti-epsilon toxin antibodies in the sera of vaccinated rabbits or guinea pigs. To be approved, a vaccine must produce an antibody response of at least 5 U/mL of anti-epsilon toxin antibodies (Rosskopf-Streicher, Volkers & Werner 2003). Although the MNT is known to be sensitive, it is cumbersome, expensive and slow and it can be relatively imprecise as often a ranged result is determined (Pfahler et al. 1998; Uzal, Nielsen & Kelly 1997). Moreover, apart from ethical considerations due to the use of large numbers of animals, the MNT may also suffer disadvantages inherent to biological tests such as variation in animal sensitivity and precision of the inoculation technique (Hendriksen 2007; Kozaki et al. 1979; Levings, Henderson & Metz 1993; Pfahler et al. 1998).
Various reports have expressed the need for a reduction in the use of animals for vaccine potency testing (Hendriksen 2007; Kulpa-Eddy et al. 2011). Whist there is no alternative to immunising test animals when assessing the potency of vaccines, the quantification of protective anti-epsilon toxin antibodies may be determined by in vitro methods. Previous studies have reported on the development and evaluation of an indirect enzyme-linked immunosorbent assay (ELISA) and competitive ELISA to measure epsilon antitoxin in the sera of vaccinated rabbits. ELISAs have been reported to offer several advantages over the MNT test (Ebert et al. 1999; Pfahler et al. 1998; Rosskopf-Streicher, Volkers & Werner 2003). In addition to precluding the use of laboratory animals, the ELISA technique allows for greater numbers of samples to be processed at one time and includes fewer complex standardisation processes compared with the MNT (Levings, Henderson & Metz 1993; Uzal, Nielsen & Kelly 1997).
As an alternative in vitro test to quantify anti-epsilon toxin antibodies in the sera of vaccinated guinea pigs, a flow cytometric microbead-based indirect immunoassay was developed and evaluated in this study. In contrast with the medical field, where cytometric microbead-based immunoassays have been used extensively for diagnostics and research, they have not yet been widely applied in the veterinary field or in veterinary vaccine potency testing (Morgan et al. 2004; Sanchez-Carbayo 2006; Vignali 2000). This technique, which is based on the combination of ELISA-based technology with flow cytometry, uses microbeads as the solid support for a conventional immunosorbent assay with a green-fluorescent reporter dye (Kellar & Iannone 2002). Although the technique incorporates all the advantages of the ELISA, the dynamic range of the cytometric bead-based immunoassay is greater than that of ELISA. Such an increase in the dynamic range reportedly makes these assays more sensitive and accurate than ELISA systems and significantly reduces the sample volumes required (Du Pont et al. 2005; Lal et al. 2004; Morgan et al. 2004).
Due to concerns regarding animal welfare and the large number of animals used in the MNT, there is a need for a more humane and cost-effective method to accurately and reliably determine the potency of enterotoxaemia vaccines. The aim of this study was therefore to develop and evaluate a flow cytometric bead-based assay to detect post-vaccination serum antibodies in an effort to quantify the immunogenicity of enterotoxaemia vaccines. The results were compared with those obtained using a conventional ELISA and the prescribed MNT.
Materials and methods
A vaccine production strain of Clostridium perfringens type D was cultured anaerobically in pulpy kidney medium (PKM) at 37 °C for 24 h. The inoculum was then pumped from the pilot fermenter into the production fermenter containing sterile PKM medium under the same environmental conditions. Anaerobic growth was allowed for 18 h. The epsilon toxin was precipitated from the cell-free culture supernatant with ammonium sulphate, and residual salts were removed by dialysis against sterile ddH2O at 4 °C for 24 h (Habeeb 1969; Thomson 1963; Whitaker & Granum 1980). The epsilon toxin was purified by ion exchange chromatography using DEAE Sepharose resin (GE Healthcare, United Kingdom), as described previously (Laemmli 1970). To assess the purity of the epsilon toxin preparation, samples were analysed by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) (Owens et al. 2000). To confirm the identity of the purified epsilon toxin, an enterotoxaemia ELISA was performed using the BioK 270 kit according to the instructions of the manufacturer (Bio-X Diagnostics, Europe). The concentration of the purified epsilon toxin was determined by the Bradford method, using bovine serum albumin (BSA) as the standard (Bradford 1976).
In this study, sera were obtained from guinea pigs that had been vaccinated with three independently produced batches of enterotoxaemia vaccine (Onderstepoort Biological Products Ltd, Pretoria). For each vaccine batch, eight naive guinea pigs were injected subcutaneously with 2 mL of the vaccine and booster injections were given three weeks following the initial vaccination. Sera were collected prior to immunisation of the guinea pigs and at five weeks post-vaccination. The guinea pig sera were prepared into two serum samples, designated A and B, where A comprised the pooled sera of four randomly selected guinea pigs and B comprised the pooled sera of the remaining four guinea pigs. The serum samples were stored at -22 °C until their use in vaccine potency evaluations. The resulting six test sera groups were designated 198A and 198B, 199A and 199B, and 200A and 200B.
The mouse neutralisation test (MNT) was used to evaluate anti-epsilon toxin serum antibodies from guinea pig test sera for epsilon toxin neutralisation (European Pharmacopia 2008:0363). The guinea pig test sera were serially diluted in filtered saline and each dilution tube was mixed with equal volumes of standardised epsilon toxin. The prepared test samples were incubated at 37 °C for 30 min and 0.2 mL of the samples were injected intravenously into four mice per dilution. The mice were observed for 24 h and the results were noted.
The Cytometric Bead Array (CBA) kit (Becton Dickinson) was used to develop a flow cytometric-based immunoassay in order to quantify anti-epsilon toxin antibodies in the sera from vaccinated guinea pigs. The optimal antigen concentration for coating the beads was determined by a standard checkerboard titration method prior to performing the indirect cytometric bead immunoassay (Cameron 1982). On the day of the assay, the positive control guinea pig serum used in the standard checkerboard titration was diluted with serum diluent (supplied in the CBA kit) from 50 U/mL to 40 U/mL, 30 U/mL, 20 U/mL, 10 U/mL and 5 U/mL, respectively, in order to construct a standard curve. The purified epsilon toxin was added to a 96-well Nunc microtiter plate at a concentration of 1 mg/mL to a total volume of 50 µL per well. Functional beads (Becton Dickinson) were diluted to a 1 χ working concentration and 50 µL were added to each well. Following incubation of the microtitre plate at room temperature for 1 h, 150 µL of the kit washing buffer was added to each well and the plate was centrifuged at 200 g for 5 min. Unbound epsilon toxin was aspirated using a multichannel pipette. On a separate microtitre plate, each test serum sample was diluted in to a 1/400 concentration using kit serum diluent across the columns of the plate. Fifty (50) µL of the diluted positive control guinea pig serum and 50 µL of each diluted test serum sample was added to respective wells of the plate containing the functional beads and incubated at room temperature for 1 h. After incubation, the plate was washed and the beads pelleted as described above. The supernatant from each well was aspirated using a multichannel pipette and 50 µL of anti-guinea pig FITC-labelled secondary antibody (Sigma-Aldrich), diluted 1/128 in the kit sample diluent, was added to each well, mixed and then incubated at room temperature for 2 h. Samples were washed once, as described above, then re-suspended in 300 µL of wash buffer and analysed in triplicate on three different occasions on a FACSCalibur™ flow cytometer (Becton Dickinson). The reactivity threshold was calculated using negative sera from the guinea pigs obtained pre-vaccination. The mean +1 standard deviations were used to predict an analytical cut-off. Antibody concentrations in each test serum were extrapolated from the standard curve using linear curve fitting.
An antibody-capture immunoassay (ELISA), using purified epsilon toxin antigen and an indirect detection format, was used to quantify antibody levels to the epsilon toxin in guinea pig test sera. For this purpose, 100 µL of the purified epsilon toxin (diluted to 100 µg/mL in 20 M carbonate buffer) was passively adsorbed onto Nunc MaxiSorp ELISA plates overnight at room temperature. The plates were then washed three times with 300 µL per well of TST buffer (0.8 M Tris-HCl, 0.15 M NaCl, 0.05% [v/v] Tween-20; pH 8) to remove unbound epsilon toxin. The coated plates were blocked (as part of the kit) by the addition of 300 µL of blocking buffer at 37 °C for 1 h. After washing three times with TST buffer, 100 µL of each test serum, diluted 1/400 in blocking buffer, was added in triplicate into the coated wells. Following incubation at 37 °C for 1 h, plates were washed and a volume of 100 µL of recombinant protein G conjugated with horseradish peroxidase (Sigma-Aldrich), diluted 1/8000 in blocking buffer, was added to each well. After incubation at 37 °C for 1 h, the plates were washed three times in TST buffer and 100 µL of tetramethylbenzidine (TMB) peroxidase substrate (Invitrogen) was added to each well. The plates were incubated in the dark at room temperature for 10 min. The reactions were stopped by addition of 50 µL per well of 2 N H2SO4 and the optical densities (OD) were measured at 450 nm. For both assays, the samples were performed in duplicate on three different occasions. Antibody concentrations were extrapolated from a standard curve using linear curve fitting. Standard curves were prepared by diluting positive control guinea pig serum with diluent from 50 U/mL to 40 U/mL, 30 U/mL, 20 U/mL, 10 U/mL and 5 U/mL, respectively. The assays were performed with six repetitions. The reactivity threshold was calculated using negative sera from the guinea pigs obtained pre-vaccination. The mean +1 standard deviation were used to predict an analytical cut off. The antitoxin content of the test sera was determined using the equation for a straight line. The means, the standard deviations and the intra-assay and inter-assay coefficients of variation were calculated according to standard statistical literature (Naute 2001; Tijssen 1988).
Results
Purified epsilon toxin was required as antigen for the quantification of anti-epsilon toxin antibodies in post-vaccination guinea pig serum using the MNT, I-CBA and I-ELISA. Analysis of the Coomassie Blue-stained gel loaded with a sample of the purified epsilon toxin indicated the presence of a 33-kDa protein of which the molecular mass is in agreement with that reported for the epsilon toxin. Moreover, the toxin was purified to near homogeneity as was evident by the lack of contaminating proteins (Figure 1). The concentration of the purified epsilon toxin was determined to be 1.126 mg/mL using a Bradford protein assay. To verify the identity of the purified 33-kDa protein, an enterotoxaemia ELISA kit was used that is capable of distinguishing between C. perfringens alpha, beta and epsilon toxins. The results (not shown) indicated that the toxin preparation tested negative for both the alpha and beta toxins but positive for epsilon toxin.
The dose of toxin used in the MNT was standardised by determining the LD50 in mice, and further confirmation of the toxin dose was established by MNT using a standardised positive control guinea pig serum. All pre-bleed test sera were negative for anti-epsilon toxin antibodies. Five of the six groups of guinea pig test serum samples collected five weeks post-vaccination had antitoxin levels above the required minimum (i.e. not less than 5 U/mL), whereas group 200A did not meet the requirement (Table 1).
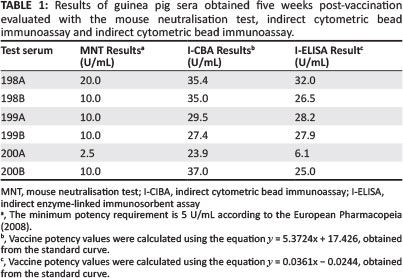
A Cytometric Bead Array (CBA) kit, (Becton Dickinson) was used to develop a flow cytometer-based immunoassay. The CBA system combines a conventional ELISA immunoassay with the sensitivity of fluorescence via flow cytometry (Aitken 2007; Naute 2001). Sample acquisition using CellQuest Pro™ software of the flow cytometer generated dot plots for every sample, which generated median fluorescence intensity (MFI) values for every population of dots (representing the sample). Unconjugated negative control beads were acquired using a dot plot with every assay, thus verifying the instrument setup. Pre-vaccination serum samples were analysed to evaluate the guinea pig starting immunity. The average MFI values for the pre-vaccination serum samples plus 1 standard deviation were used to calculate the cutoff value with 95% confidence. An MFI value of 33.3 was estimated to be the lower level of detection. The results for the guinea pig test serum samples, presented in Table 1, indicated that serum sample 200A had the lowest level of anti-epsilon toxin antibodies, similarly indicated by the results from the MNT.
An indirect ELISA was used as a second in vitro test to semi-quantify anti-epsilon toxin antibodies in guinea pig serum samples five weeks post-vaccination. To evaluate the guinea pig starting immunity, the OD450 values of the pre-vaccination serum samples were determined. The OD450 values for the pre-vaccination serum samples plus 1 standard deviation were used to calculate the cut off value with 95% confidence. An OD450 value of 0.046 was estimated to be the lower level of detection. The results for the guinea pig serum samples obtained five weeks post-vaccination are presented in Table 1. The results indicated that all 6 groups of guinea pig test serum samples had antitoxin levels above the required minimum antitoxin level (i.e. not less than 5 U/mL). However, the value obtained for group 200A was the lowest, as was also indicated by the results from both the MNT and I-CBA.
Discussion
The epsilon toxoid vaccine is prepared from the culture filtrate of a C. perfringens type D isolate and therefore the corresponding vaccine induces antibodies against numerous cellular and extracellular proteins of the bacterial culture. Since the specificity of the indirect cytometric bead assay (I-CBA) and indirect ELISA (I-ELISA) is mainly a function of the purified antigen used in these tests, only epsilon toxin purified by ion exchange chromatography was used in this study. The results demonstrated that with the purification technique used, the antigen obtained was indeed epsilon toxin since it reacted specifically with an anti-epsilon toxin monoclonal antibody. The purified epsilon toxin was subsequently used as the antigen component in the in vivo MNT and the in vitro I-CBA and I-ELISA tests. Moreover, for quantitative immunoassays, standard curves were obtained for the I-CBA and I-ELISA after six assays at different concentrations of a standardised positive control guinea pig serum. The correlation coefficient of R2 = 0.998 obtained for both standard curves serves to confirm the reliability of the results (Naute 2001).
The epsilon antitoxin levels of six guinea pig test serum samples determined in vitro using the I-ELISA and the I-CBA were calculated as mean values of three replicate assays performed independently. The intra-assay coefficient of variation was 7.2% - 15.6% and the inter-assay coefficient of variation was 10.0% - 18.6% for the I-CBA, whereas the intra-and inter-assay coefficients of variation were respectively 4.8% - 13.9% and 2.7% - 19.5% for the I-ELISA. The intra- and inter-assay variability of the respective assays is within the normal range and should not exceed 20.0% - 30.0% and is also comparable to that previously reported (Du Pont et al. 2005; Ebert et al. 1999; Lal et al. 2004; Tijssen 1988). Therefore, the reproducibility of the I-CBA and I-ELISA used in this study was considered to be satisfactory.
The epsilon antitoxin levels of each guinea pig serum determined by the I-ELISA and I-CBA were subsequently compared with the corresponding results of the MNT. For all serum samples tested, the I-CBA and I-ELISA tests obtained higher antitoxin levels than that of the MNT assay. The higher antitoxin levels determined in the I-CBA and I-ELISA compared with the MNT may be ascribed to the binding of both neutralising and non-neutralising anti-epsilon toxin antibodies present in the sera of vaccinated guinea pigs to the epsilon antigen in the respective in vitro tests. In contrast, the MNT allows for quantification of only neutralising anti-epsilon toxin antibodies that are capable of protecting the mice against the lethal effects of the epsilon toxin and therefore does not take non-neutralising antibodies into account (Levings, Henderson & Metz 1993; Notermans, Dufrenne & Kozaki 1982; Pfahler et al. 1998).
Despite the use of the same antigen and the same serum samples in the I-ELISA and I-CBA tests, the values obtained with the I-CBA were noticeably higher than those obtained in the I-ELISA. It is tempting to speculate that the differences may be due to technical differences between the two test systems. Moreover, an additional factor that may affect the readings is attachment of the capture antigen. In the I-CBA the epsilon toxin is attached firmly to the surface-functional microbeads by covalent bonds via standard amine chemistry (Hashida et al. 1984). In contrast, the antigen is attached to the wells of an ELISA plate primarily by hydrophobic interaction, although some electrostatic forces may also contribute (Engvall & Perlmann 1972). Consequently, it is possible that detachment of the bound antigen and immune complexes during subsequent washing steps can give rise to poorer sensitivity (Howell, Nasser & Schray 1981).
The post-vaccination guinea pig serum samples used in this study had varying levels of potency as assessed in the in vivo MNT. The serum samples showed potencies mostly in the range of 10 U/mL, but serum sample 198A had potency in the range of 20 U/mL and serum sample 200A had potency less than the required 5 U/mL. Of the two in vitro assays, the I-ELISA could discriminate differences in the potency of the vaccines to a greater extent than the I-CBA. In contrast, epsilon antitoxin levels determined in the I-CBA did not differ significantly from each other. Nevertheless, all of the serum samples were unequivocally identified in both in vitro tests as being positive, with more than 5 U/mL epsilon antitoxin content. In contrast, the MNT was the only assay that indicated that serum 200A did not meet the required value (i.e. not less than 5 U/mL). This therefore means that neither the I-CBA nor I-ELISA in their current format can be used as possible alternatives to the MNT for estimating the potency of epsilon toxoid-containing C. perfringens type D vaccines for veterinary use. However, further optimisation of the respective in vitro tests may enable their use in routine potency testing of these vaccines.
In contrast to previous studies, the antitoxin levels determined in the I-ELISA did not compare favourably to those obtained in the MNT (Pfahler et al. 1998). Notably, in the only other study to have used an I-ELISA for potency testing of epsilon toxoid veterinary vaccines, it was reported that best correlation with the MNT results was obtained for plates coated with high amounts of purified antigen (15 mg/mL) and low serum sample dilutions (1/25). Indeed, it was reported that serum dilutions of 1/100 and higher gave recurrently higher results than the MNT. Since only high avidity antibodies are neutralising in vivo, it was suggested that high antigen density and low sample dilution are most likely to enhance the selection of such high affinity antibodies (Pfahler et al. 1998). As an alternative to optimising the current I-ELISA with regard to the above-mentioned parameters, consideration may also be given to the use of a competitive ELISA in future studies. Several reports have demonstrated a high correlation (at least 0.93) in epsilon antitoxin levels determined by competitive ELISA, based on the use of a monoclonal antibody, and the MNT (Ebert et al. 1997; Rosskopf-Streicher, Volkers & Werner 2003; Uzal, Nielsen & Kelly 1997).
In order to improve the I-CBA assay for use in measuring vaccine potency, further optimisation of the assay is required. Variation of the density of mouse monoclonal capture antibodies on the beads during repeated coupling procedures were tested (Dasso et al. 2002). The authors observed that minor variations in the coupling procedures can markedly affect the density of coupled capture antibodies, which, in turn, can influence the sensitivity of the assays. Therefore, the variability introduced by using bead sets coupled independently with the purified epsilon antigen needs to be determined. Another issue that may require further attention is the choice of the fluorochrome conjugate. It has been reported that in a comparative analysis of different conjugates, conjugates incorporating streptavidin gave low background staining, whereas Alexa488 conjugates were significantly brighter than FITC conjugates and thus increased the sensitivity of the assay (Vignali 2000). The sensitivity of the flow cytometric system may also be enhanced further by reducing the number of beads per test. It has been reported that such reduction results in an increase in the ratio of antigen to antibody in each test without reducing the potential signal strength of the assays (the concentration of capturing antigens per bead) (Dasso et al. 2002; Vignali 2000). It should be noted that although the I-CBA assay incorporates washing steps, a number of studies have reported rapid 'no wash' assays that can be performed in less than two hours (Cook et al. 2001; Tripp et al. 2000). Although this may reduce the time required to perform the assays, the effect of omitting washing steps on the sensitivity of the assay would need to be determined first.
Conclusion
The results indicated that the in vitro I-CBA and I-ELISA tests in their current formats are not yet suitable as alternatives to the in vivo MNT for assessing the potency of C. perfringens type D epsilon toxoid vaccines. However, the I-CBA assay that was developed and evaluated has the potential to become a viable alternative to the MNT for measuring vaccine potency. Besides the advantage of avoiding the use of large numbers of animals as indicators for lethal and sublethal toxicity, the low costs and the simple performance of the I-CBA may allow, on a routine scale, the investigation of serum samples from vaccinated guinea pigs. However, for the I-CBA to be considered as a viable replacement to the MNT, further optimisation of the assay is required as highlighted above. An optimised I-CBA assay will consequently offer the possibility of assay standardisation and validation, which are a precursor for replacement of the MNT. Further incisive studies are therefore required.
Acknowledgements
The authors thank Onderstepoort Biological Products (OBP) for financial support, Mrs E. du Piesanie for assistance with flow cytometry and Dr C. Ellis, Onderstepoort Veterinary Institute, for help in purifying the antigen.
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Authors' contributions
A.B. (Onderstepoort Biological Products) was the project leader responsible for experimental and project design and performed most of the experiments. J.T. (University of Pretoria) and J.C. (University of Pretoria) made conceptual contributions.
References
Aitken, I.D., 2007, 'Clostridial diseases', in C.J. Lewis (ed.), Diseases of sheep, pp. 156167, Blackwell Publishing, Oxford. http://dx.doi.org/10.1002/9780470753316 [ Links ]
Bhown, A.S. & Habeeb, A.F.S.A., 1977, 'Structural studies of epsilon-prototoxin of Clostridium perfringens type D: Localisation of the site of tryptic scission necessary for activation to epsilon-toxin', Biochemical and Biophysical Research Communications 78, 889-896. http://dx.doi.org/10.1016/0006-291X(77)90506-X [ Links ]
Bradford, M.M., 1976, 'Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding', Analytical Biochemistry 72, 248-254. http://dx.doi.org/10.1016/0003-2697(76)90527-3 [ Links ]
Cameron, C.M., 1982, 'Antibody response of guinea-pigs and cattle to a Campylobacter fetus oil emulsion vaccine', Onderstepoort Journal of Veterinary Research 49, 177179. PMid:7177598 [ Links ]
Cook, E.B., Stahl, J.L., Lowe, L., Chen, R., Morgan, E., Wilson, J. et al., 2001, 'Simultaneous measurement of six cytokines in a single sample of human tears using microparticle-based flow cytometry: Allergics vs. non-allergics', Journal of Immunological Methods 254, 109-118. http://dx.doi.org/10.1016/S0022-1759(01)00407-0 [ Links ]
Dasso, J., Lee, J., Bach, H. & Mage, R.G., 2002, 'A comparison of ELISA and flow microsphere-based assays for quantification of immunoglobulins', Journal of Immunological Methods 263, 23-33. http://dx.doi.org/10.1016/S0022-1759(02)00028-5 [ Links ]
Du Pont, N.C., Wang, K.H., Wadhwa, P.D., Culhane, J.F. & Nelson, E.L., 2005, 'Validation and comparison of Luminex multiplex cytokine analysis kits with ELISA: Determinations of a panel of nine cytokines in clinical sample culture supernatants', Journal of Reproductive Immunology 66, 175-191. http://dx.doi.org/10.1016/j.jri.2005.03.005, PMid:16029895 [ Links ]
Ebert, E., Óppling, V., Werner, E. & Cussler, K., 1997, 'Quantification of antibodies against C. perfringens beta and epsilon toxin. The ELISA as an alternative to mouse challenge assay for potency testing of veterinary vaccines', Pharmeuropa Bio 1, 40-46. [ Links ]
Ebert, E., Óppling, V., Werner, E. & Cussler, K., 1999, 'Development and pre-validation of two different ELISA systems for the potency testing of Clostridium perfringens beta and epsilon toxoid containing veterinary vaccines', FEMS Immunology and Medical Microbiology 24, 299-311. PMid:10397315 [ Links ]
Engvall, E. & Perlmann, P., 1972, 'Quantitation of specific antibodies by enzyme-labeled anti-immunoglobulin in antigen-coated tubes', Journal of Immunology 109, 129-135. PMid:4113792 [ Links ]
European Phamacopoeia, 2008, Monograph 01/2008:0363, European Directorate for the Quality of Medicines (EDQM), Strassbourg. [ Links ]
Habeeb, A.F., 1969, 'Studies on epsilon-prototoxin of Clostridium perfringens type D. Purification methods: Evidence for multiple forms of epsilon-prototoxin', Archives of Biochemistry and Biophysics 130, 430-440. http://dx.doi.org/10.1016/0003- 9861(69)90055-1 [ Links ]
Hashida, S., Imagawa, M., Inoue, S., Ruan, K.H. & Ishikawa, E., 1984, 'More useful maleimide compounds for the conjugation of Fab to horseradish peroxidase through thiol groups in the hinge', Journal of Applied Biochemistry 6, 56-63. PMid:6490581 [ Links ]
Hendriksen, C., 2007, 'Three Rs achievements in vaccinology', Alternatives to Animal Testing and Experimentation 14, 575-579. [ Links ]
Howell, E.E., Nasser, J. & Schray, K.J., 1981, 'Coated tube enzyme immunoassay: Factors affecting sensitivity and effects of reversible protein binding to polystyrene', Journal of Immunoassay 2, 205-225. http://dx.doi.org/10.1080/15321818108056978, PMid:7047571 [ Links ]
Kellar, K.L. & Iannone, M.A., 2002, 'Multiplexed microsphere-based flow cytometric assays', Experimental Hematology 30, 1227-1237. http://dx.doi.org/10.1016/S0301-472X(02)00922-0 [ Links ]
Kozaki, S., Dufrenne, J., Hagenaars, A.M. & Notermans, S., 1979, 'Enzyme-linked immunosorbent assay (ELISA) for the detection of Clostridium perfringens type B toxin', Japanese Journal of Medical Science and Biology 32, 199-205. PMid:119079 [ Links ]
Kulpa-Eddy, J., Srinivas, G., Halder, M., Hill, R., Brown, K., Roth, J. et al., 2011, 'Non-animal replacement methods for veterinary vaccine potency testing: State-of-the-science and future directions', Procedía in Vaccinology 5, 60-83. http://dx.doi. org/10.1016/j.provac.2011.10.005 [ Links ]
Laemmli, U.K., 1970, 'Cleavage of structural proteins during the assembly of the head of bacteriophage T4', Nature 227, 680-685. http://dx.doi.org/10.1038/227680a0, PMid:5432063 [ Links ]
Lal, G., Balmer, P., Joseph, H., Dawson, M. & Borrow, R., 2004, 'Development and evaluation of a tetraplex flow cytometric assay for quantitation of serum antibodies to Neisseria meningitides serogroups A, C, Y and W-135', Clinical and Diagnostic Laboratory Immunology 11, 272-279. PMid:15013975, PMCid:PMC371201 [ Links ]
Levings, R.L., Henderson, L.M. & Metz, C.A., 1993, 'In vitro potency assays for nonreplicating veterinary vaccines: Comparison to in vivo assays and considerations in assay development', Veterinary Microbiology 37, 201-219. http://dx.doi.org/10.1016/0378, 1135(93)90023-Z [ Links ]
Minami, J., Katayama, S., Matsushita, O., Matsushita, C. & Okabe, A., 1997, 'Lambda-toxin of Clostridium perfringens activates the precursor of epsilon-toxin by releasing its N- and C-terminal peptides', Microbiology and Immunology 41, 527535. PMid:9272698 [ Links ]
Morgan, E., Varro, R., Sepulveda, H., Ember, J.A., Apgar, J., Wilson, J. et al., 2004, 'Cytometric bead array: A multiplexed assay platform with applications in various areas of biology', Clinical Immunology 110, 252-266. http://dx.doi.org/10.1016/j.clim.2003.11.017, PMid:15047203 [ Links ]
Naute, J., 2001, 'Statistics in clinical vaccine trials', Springer, Heidelberg, Dordrecht, London, New York. [ Links ]
Niilo, L., 1980, 'Clostridium perfringens in animal disease: A review of current knowledge', Canadian Veterinary Journal 21, 141-148. PMid:6253040, PMCid:PMC1789702 [ Links ]
Notermans, S., Dufrenne, J. & Kozaki, S., 1982, 'The relation between toxicity and toxin-related antigen contents of Clostridium botulinum types C and D cultures as determined by mouse bioassay and ELISA', Japanese Journal of Medical Science and Biology 35, 203-211. PMid:6761472 [ Links ]
Owens, M.A., Vall, G.H., Hurley, A.A. & Wormsley, S.B., 2000, 'Validation and quality control of immunophenotyping in clinical flow cytometry', Journal of Immunological Methods 243, 33-50. http://dx.doi.org/10.1016/S0022-1759(00)00226-X [ Links ]
Oyston, P.C.F., Payne, D.W., Havard, H.L., Williamson, E.D. & Titball, R.W., 1998, 'Production of a non-toxic site directed mutant of Clostridium perfringens epsilon toxin which induces protective immunity in mice', Microbiology 144, 333-341. http://dx.doi.org/10.1099/00221287-144-2-333, PMid:9493371 [ Links ]
Pfahler, W., Bisesti, E., Pereyra, J.B. & Iribarren, F., 1998, 'Enzyme-linked immunosorbent assay for potency testing of vaccines containing Clostridium perfringens type D epsilon-toxoid', Journal of Veterinary Medicine 45, 133-139. http://dx.doi.org/10.1111/j.1439-0450.1998.tb00776.x [ Links ]
Rosskopf-Streicher, U., Volkers, P. & Werner, E., 2003, 'Control of Clostridium perfringens vaccines using an indirect competitive ELISA for the epsilon toxin component', Pharmeuropa Bio 2, 91-96. [ Links ]
Sanchez-Carbayo, M., 2006, 'Antibody arrays: Technical considerations and clinical applications in cancer', Clinical Chemistry 52, 1651-1659. http://dx.doi.org/10.1373/clinchem.2005.059592, PMid:16809399 [ Links ]
Songer, J.G., 1996, 'Clostridial enteric diseases of domestic animals', Clinical Microbiology 9, 216-234. [ Links ]
Thomson, R.O., 1963, 'The fractionation of Clostridium welchii epsilon-antigen on cellulose ion exchangers', Journal of General Microbiology 31, 79-90. http://dx.doi.org/10.1099/00221287-31-1-79, PMid:13981246 [ Links ]
Tijssen, P., 1988, 'Laboratory techniques in biochemistry and molecular biology: Practice and theory of enzyme immunoassays', Elsevier Science, Amsterdam. [ Links ]
Titball, R.W., 2009, 'Clostridium perfringens vaccines', Vaccine 27, suppl. 4, D44-D47. http://dx.doi.org/10.1016/j.vaccine.2009.07.047, PMid:19837285 [ Links ]
Tripp, R.A., Jones, L., Anderson, L.J. & Brown, M., 2000, 'CD40 ligand (CD154) enhances the Th1 and antibody response to respiratory syncytial virus in the BALB/c mouse', Journal of Immunology 164, 5913-5921. PMid:10820273 [ Links ]
Uzal, F.A., Nielsen, K. & Kelly, W.R., 1997, 'Detection of Clostridium perfringens type D epsilon antitoxin in serum of goats by competitive and indirect ELISA', Veterinary Microbiology 51, 223-231. http://dx.doi.org/10.1016/S0378-1135(97)00122-3 [ Links ]
Uzal, F.A., 2004, 'Diagnosis of Clostridium perfringens intestinal infection in sheep and goats', Anaerobe 10, 135-143. http://dx.doi.org/10.1016/j.anaerobe.2003.08.005, PMid:16701510 [ Links ]
Vignali, D.A., 2000, 'Multiplexed particle-based flow cytometric assays', Journal of Immunological Methods 243, 243-255. http://dx.doi.org/10.1016/S0022-1759(00)00238-6 [ Links ]
Whitaker, J.R. & Granum, P.E., 1980, 'Improved method for purification of enterotoxin from Clostridium perfringens type A', Applied Environmental Microbiology 39, 1120-1122. PMid:6250473, PMCid:PMC291492 [ Links ]
 Correspondence:
Correspondence:
Angela Buys
Private Bag X07
Onderstepoort 0110, South Africa
Email: angelagouws@hotmail.com
Received: 09 Jan. 2013
Accepted: 30 May 2013
Published: 10 Mar. 2014

